1. Introduction
Histone chaperones serve a pivotal role in histone
metabolism, facilitating histone binding to DNA during processes
such as DNA replication and repair (1). Originally, Laskey et al
(2) revealed that nucleophosmin
in Xenopus eggs can bind histones and promote nucleosome
formation independently of ATP. These two properties remain the
only shared, and thus defining, characteristics of histone
chaperones. Currently, histone chaperones are deemed to aid histone
transfer but remain separate from the final histone-DNA complex
(3). Extensive research has been
conducted on the relationship between histone chaperones and human
diseases (3-5). Their abnormal expression patterns,
not only in tumors, but also in cardiovascular diseases, autoimmune
disorders and neurodegenerative diseases, highlight their
prognostic value in various human diseases. Given their role in
chromatin dynamics and disease contexts, they present potential
therapeutic targets and diagnostic markers in various human
diseases.
Retinoblastoma binding protein (RBBP)4 and RBBP7 are
recognized as histone chaperones, both of which belong to the WD40
family (6). The WD40 domain is a
distinct protein motif that typically adopts a β-propeller
architecture, mediating protein-protein interactions (PPIs).
RBBP4/7 typically exist in complexes, such as the nucleosome
remodeling and deacetylase (NuRD) complex (7) and polycomb repressive complex 2
(PRC2) (8), to regulate
chromatin remodeling and gene expression through the interactions
of their WD40 repeats with the H4 α1 helix and H3 tail (9,10). Dysregulation of RBBP4/7 may
disrupt the normal chromatin remodeling process, resulting in
altered gene expression patterns that contribute to the initiation
and progression of cancer and other human diseases (11). Notably, alterations in the
expression levels of RBBP4/7 have been observed in different types
of human disease, such as esophageal squamous cell carcinoma (ESCC)
and colorectal cancer (CRC). These changes are closely related to
clinicopathological features (12-15).
The present review summarizes the pivotal role of
RBBP4/7 in regulating cell fate, assessing their expression,
functions, clinical features and associated mechanisms in human
diseases.
2. Molecular structure of RBBP4 and
RBBP7
RBBP4, also known as RbAp48 or NURF55, is integral
to multiple chromatin-modifying and remodeling complexes. It was
originally discovered as a binding partner of the tumor suppressor
retinoblastoma protein (RB) in yeast (16), and subsequently revealed to
cofractionate with histone deacetylase (HDAC)1 (17). Located on chromosome 1p35.1, the
RBBP4 gene encodes a 425-amino acid protein ubiquitously
present across human tissues (18). The molecular structure of the
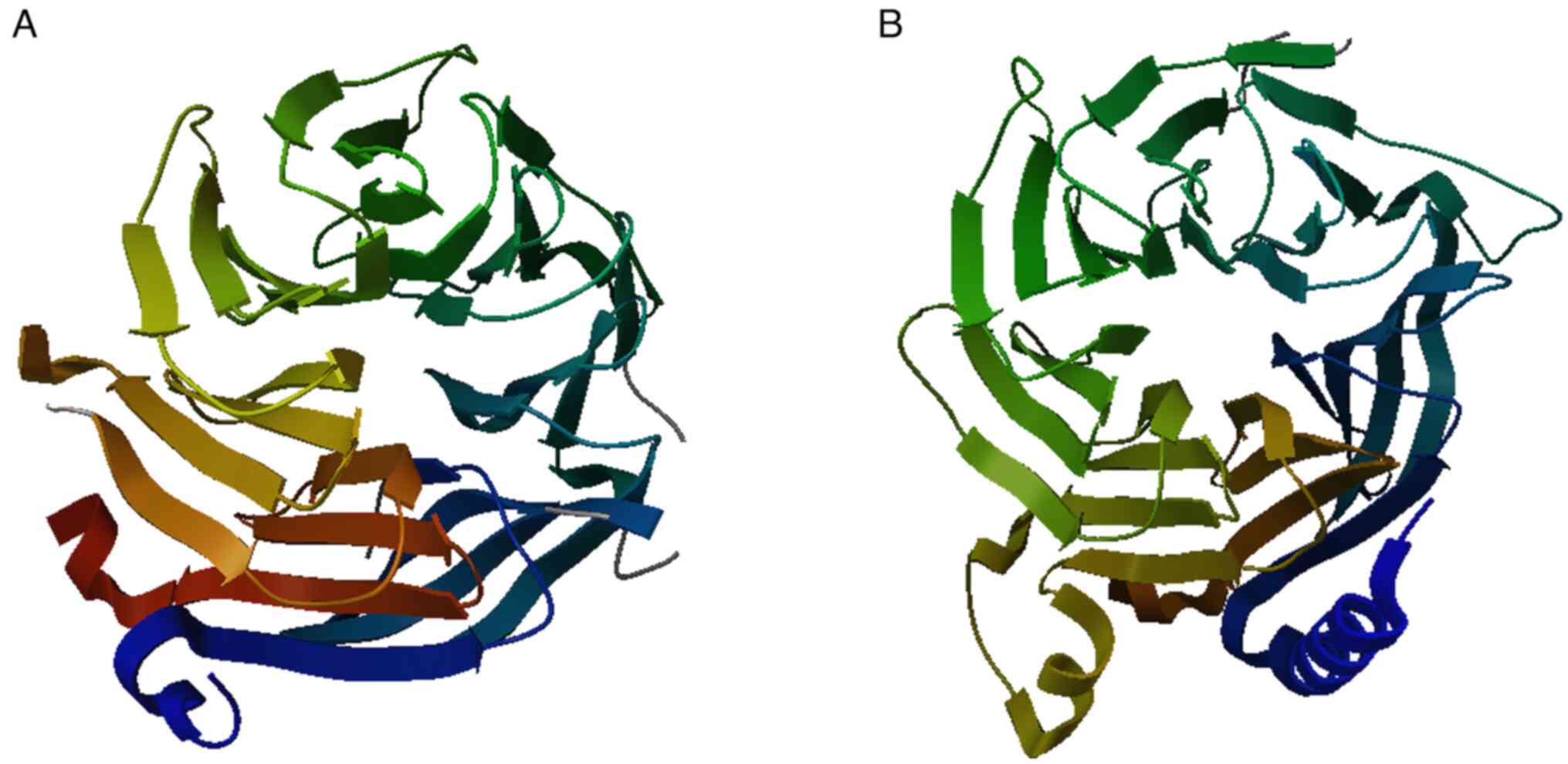
RBBP4 protein is shown in Fig.
1A.
RBBP7, alternatively known as RbAp46, is a nuclear
protein ubiquitously expressed across various cell types. Located
on chromosome 3p25.1, the RBBP7 gene encodes a 425-amino
acid protein that is universally expressed in human tissues
(18). RBBP4 and RBBP7 are 92%
identical (9); however, they
differ in certain amino acid sequences, which may lead to subtle
structural variations, especially in domains that interact with
other proteins. The molecular structure of RBBP7 is displayed in
Fig. 1B. Structurally, both
RBBP4 and RBBP7 possess a seven-bladed WD40 repeat domain,
indicating that RBBP4 and RBBP7 can serve multiple roles in
chromatin remodeling, histone modification and transcriptional
regulation (19). Notably, both
RBBP4 and RBBP7 are integral subunits of the NuRD complex (7), the switch-independent 3A complex
(20) and PRC2 (8). Additionally, RBBP4 has been
identified as a subunit of the chromatin assembly factor 1 (CAF-1)
complex (21) and a core
component of the MuvB complex (22), while RBBP7 is known to be an
essential component of the histone acetyltransferase 1 (HAT1)
complex (23). As part of these
multisubunit protein complexes, RBBP4 and RBBP7 are believed to
function as chromatin adapters, mediating direct interactions with
histone H3/H4 (24).
3. The biological functions of RBBP4/7
The functions of RBBP4/7 in the cell
cycle
RBBP4/7 have a pivotal role in cell cycle
regulation, with their absence leading to dysregulation of cell
cycle genes and cycle arrest (25). Specifically, RBBP4 deficiency
results in S phase defects and inhibits mitotic exit (M to
G1 transition) (26),
whereas RBBP7 deficiency causes G2/M phase arrest in 293
cells (27). In addition,
RBBP4/7 (LIN-53) are crucial for centromere protein A (CENP-A)
localization to centromeres (28). It has also been suggested that
the Cullin-4 (CUL4) RING ligase (CRL4) complex containing RBBP7
might regulate mitosis by promoting ubiquitin-dependent loading of
newly synthesized CENP-A during the G1 phase (29).
RBBP4/7 are members of the RB family (30). Studies conducted in yeast and
cultured cells have shown that RBBP4 appears to function as a tumor
suppressor along with RB, leading to inhibition of cell cycle
progression and cell growth (16,18). Nevertheless, Schultz-Rogers et
al (31) indicated that
RBBP4 is essential for the cell cycle progression of neural
progenitor cells and the initiation of G0, irrespective
of the involvement of RB. The E2F family plays a key role in cell
cycle regulation, and all RB family members interact with typical
E2F proteins to form transcriptional inhibition complexes (32). RB primarily inhibits E2F
transcription factor 1, whereas RBBP4 can be directed to RB via
HDAC1 (33). In the
G1 phase, RBBP4/7 and RB inhibit E2F target gene
activation, preventing entry into S phase. Cyclin
D-cyclin-dependent kinase (CDK)4/6 can monophosphorylate RB,
whereas cyclin E-CDK1/2 are involved in poly-phosphorylation or
hyper-phosphorylation of RB (34). Once hyperphosphorylated, RBBP4/7
and RB dissociate from E2F, activating target genes and recruiting
chromatin remodelers (35)
(Fig. 2A).
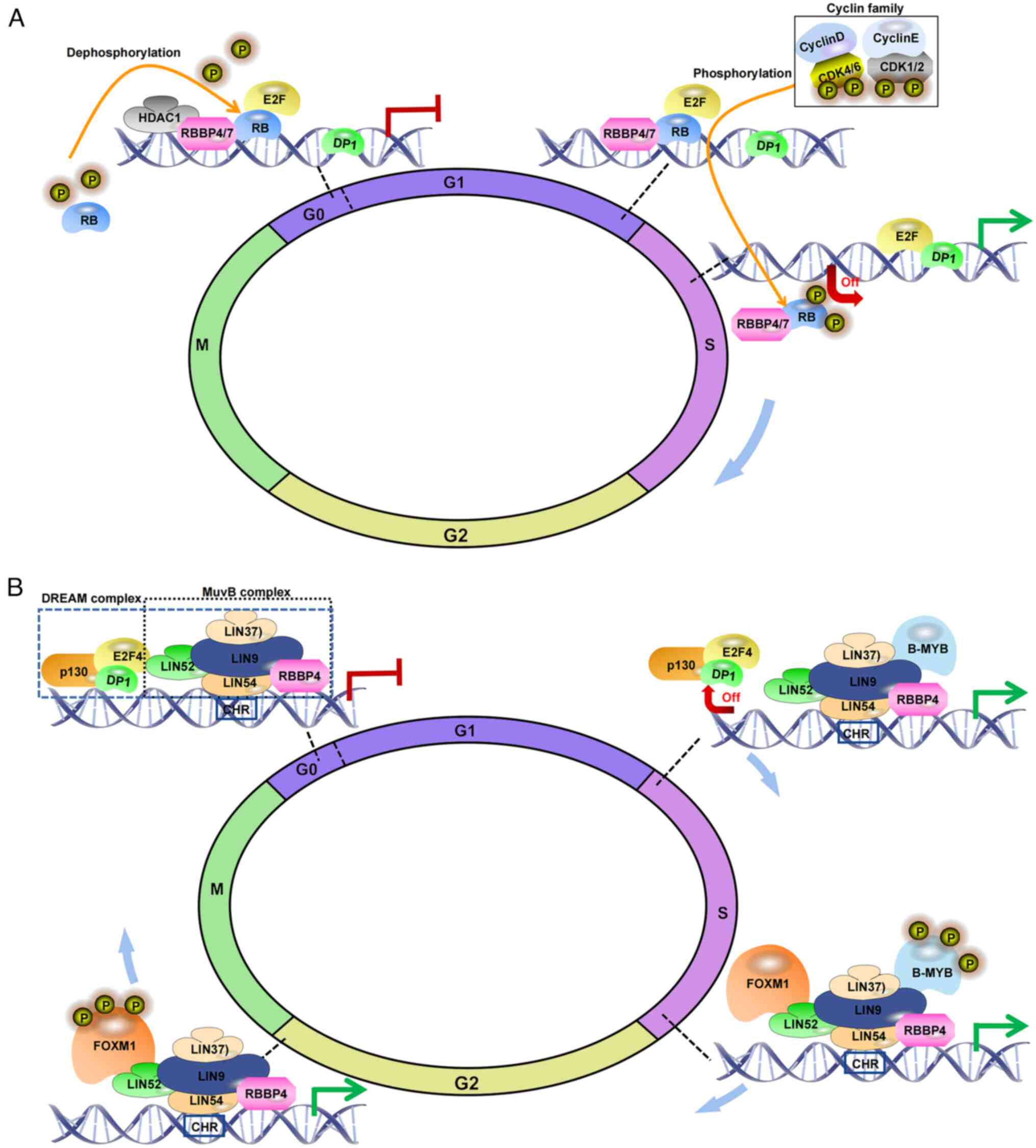 | Figure 2Molecular mechanism underlying how
RBBP4/7 affects the cell cycle. (A) Involvement of RBBP4/7 in cell
cycle regulation via RB interaction: Guided by HDAC1, RBBP4/7 is
oriented to interact with RB. During the
G0/G1 phase of the cell cycle, RB restrains
the expression of genes regulated by transcription factors of the
E2F family via binding to them, thus the dephosphorylated RB
prevents cell entry into S phase. As the cell prepares to
transition from the G1 phase to the S phase, RB is
phosphorylated by CDKs, and cyclins D/E. This phosphorylation leads
to the disassociation of RB together with RBBP4 from E2F and gene
promotors. Through dephosphorylation, RB regains its activity,
binds to E2F once again, and inhibits excessive progression of the
cell cycle. (B) During the G0/G1 phase, the
MuvB complex, composed of LIN9, LIN37, LIN52, LIN54 and RBBP4,
forms the DREAM complex through association with the RB-like
protein p130, E2F4 and DP1 under quiescent conditions. This complex
suppresses the expression of cell cycle regulatory genes during the
G0 phase. RBBP4, through its interactions with complex
members, modulates the localization of the MuvB complex and
suppresses the transcription of specific genes. When transitioning
to the S phase, the p130/E2F4/DP1 module dissociates from the DREAM
complex, resulting in the loss of suppression. Subsequently, the
MuvB complex, with RBBP4 as the core complex, cooperates with B-MYB
to form the MMB complex, activating the expression of genes
required for the S/G2 phase. FOXM1 is then recruited to
MMB, forming the MMB-FOXM1 complex during the transition of
S/G2. B-MYB undergoes phosphorylation during the late S
phase, leading to its dissociation from the MMB-FOXM1 complex as
the cell cycle advances, while MuvB and FOXM1 persist in the DNA
until mitosis. RBBP, RB binding protein. |
In addition, RBBP4 is a core component of the MuvB
complex, collaboratively working with LIN9, LIN37, LIN52 and LIN54
to regulate the cell cycle (22). During the
G0/G1 phase, the RBBP4-containing MuvB
complex associates with p130/E2F4/DP1 to form the dimerization
partner, RB-like, E2F and MuvB (DREAM) complex (36), which inhibits expression of cell
cycle regulatory genes, maintaining the cell in a quiescent state.
Within the DREAM complex, RBBP4 directly binds to LIN9 and LIN37
within the complex, playing a pivotal role in its assembly process
(22). Additionally, RBBP4
collaborates with p130, E2F4 and DP1 to inhibit the transcriptional
activity of E2F target genes (36). As the cell cycle progresses,
phosphorylation by CDK leads to the disassembly of the DREAM
complex, releasing p130/E2F4/DP1 from MuvB. Subsequently, the MuvB
complex, inclusive of RBBP4, interacts with activated transcription
factors B-MYB (also known as MYB proto-oncogene like 2) and
forkhead box M1 (FOXM1), culminating in the formation of the
activated MYB-MuvB-FOXM1 complex. This complex further regulates
the expression of cell cycle genes, especially during the
G2 phase and mitosis (37) (Fig. 2B). Thus, RBBP4, through its
positioning and recruitment roles in the MuvB complex, serves a
critical role in the regulation of various stages of cell cycle
gene expression. Notably, the DREAM complex collaborates with RB
during quiescence to suppress cell cycle gene expression (38,39). Furthermore, in response to the
p53 tumor suppressor and genotoxic stress, the involvement of RBBP4
within the DREAM complex and its collaboration with RB becomes
evident. Specifically, the DREAM complex and RB utilize the p53-p21
pathway to induce p21 and consequently arrest the cell cycle
(39,40). The dual association of RBBP4 with
the RB family and the DREAM complex indicates that it has a crucial
role in maintaining cell cycle arrest.
Finally, RBBP4/7 serve as components of
chromatin-remodeling complexes, such as NuRD and PRC2, further
modulating the cell cycle via nucleosome acetylation and
methylation regulation (8,41).
In summary, RBBP4/7, as components of the RB family
or MuvB complex, have a pivotal role in cell cycle regulation.
Their association with chromatin remodeling further influences cell
cycle progression. While the role of RBBP4/7 in the cell cycle is
evident, the specifics of their mechanism require further
exploration.
The multifaceted role of RBBP4/7 in
chromatin remodeling
Chromatin remodeling is a critical process that
governs the accessibility of DNA to various cellular machineries,
influencing gene transcription, replication and repair (42). RBBP4/7 can affect histone
conformation by directly binding to histone H4 and H3 (43).
The NuRD complex, comprising several subunits,
including the HDAC complex, chromodomain helicase DNA binding
protein 3/4 ATPase, methyl-CpG binding domain protein 2/3, RBBP4/7,
metastasis associated 1/2/3 (MTA1/2/3) and GATA zinc finger domain
containing 2A/B, is a multifunctional entity with nucleosome
remodeling and deacetylase activities (44). These activities enable the NuRD
complex to alter chromatin structure, thereby regulating gene
expression. Mu et al (45) suggested that RBBP4 may contribute
to controlling the acetylation (ac) of lysine 27 on histone H3
(H3K27ac) levels at enhancer elements by promoting the deacetylase
activity of the HDAC complex, effectively removing acetyl groups
from H3K27 (Fig. 3A). RBBP4/7
also have regulatory chromatin remodeling effects independent of
NuRD complexes. RBBP4 promotes H3K27ac by maintaining p300 levels
(46), and together with RBBP7,
mediates H4K5ac and H4K12ac to enable CENP-A deposition into
centromeres (47). A network
including SIN3 transcription regulator family member
A-HDSAC3-RBBP4-H4 recognizes and deacetylates histones during
chromatin assembly (48). RBBP4
also controls histone deacetylation at H3K4, H4K8, H4K12 and H4K16
during meiosis I (49).
Additionally, RBBP4/7 collaborate with HAT1 in the site-specific
de novo acetylation of histone H4 (50), facilitating its nuclear delivery
and folding (51), which may be
crucial in chromatin assembly and gene expression regulation.
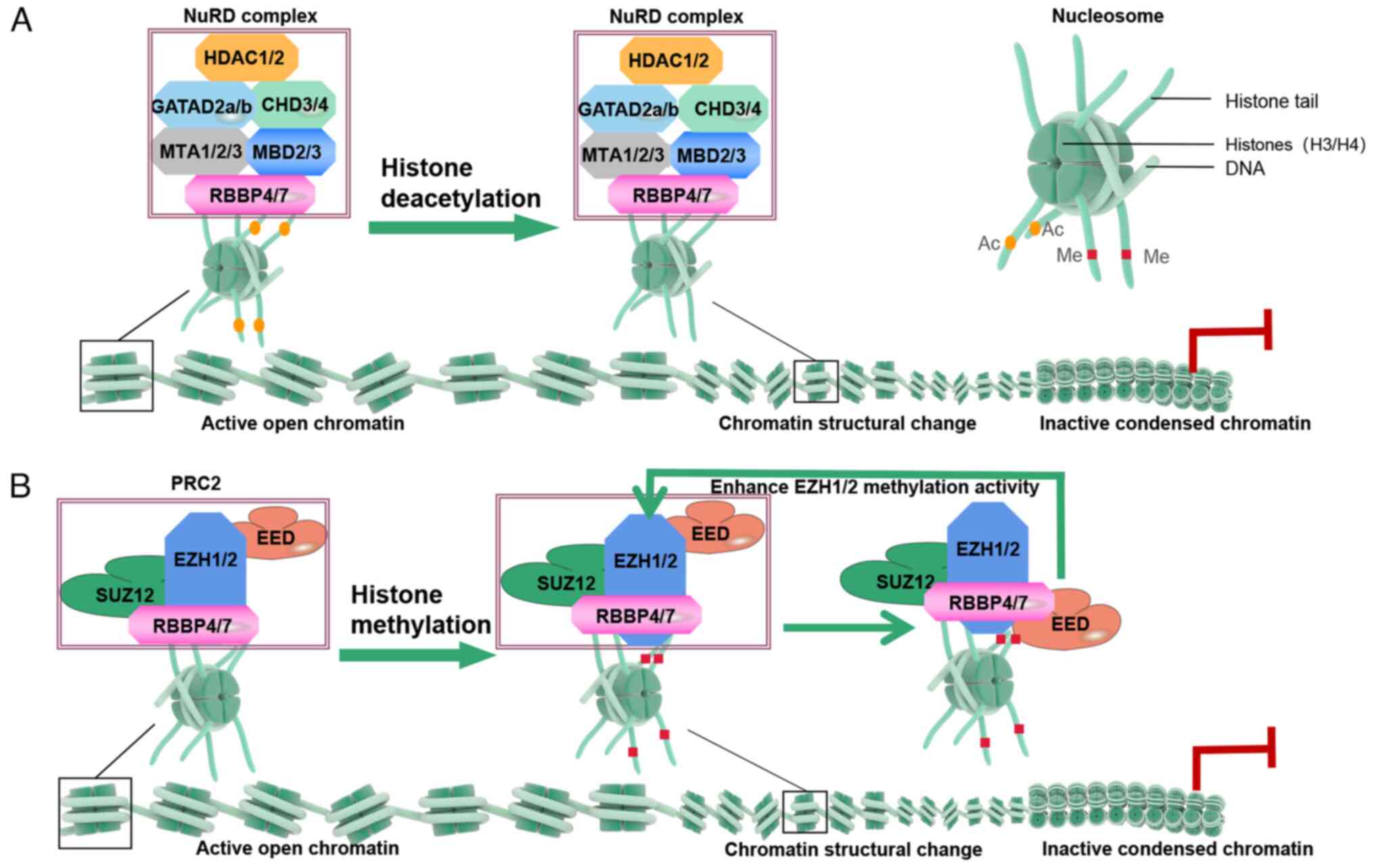 | Figure 3Mechanism of RBBP4/7 in chromatin
remodeling. (A) Deacetylation: RBBP4/7 interacts with the tails of
histones H3 and H4, promoting the deacetylase activity of the HDAC
complex, resulting in the removal of acetyl groups from H3/H4. This
leads to chromatin compaction, preventing gene promoter
transcription and inhibiting gene expression. (B) Methylation: In
PRC2, RBBP4 recruits SUZ12 to PRC2 target sites. The EZH2 subunit
serves as the methyltransferase active center, performing primary,
secondary and tertiary methylation on the 27th lysine of histone
H3, resulting in H3K27me3 formation. EED binds to the H3K27me3 mark
and further enhances the methyltransferase activity of EZH2.
PRC2-mediated methylation induces chromatin compaction, limiting
the binding of transcription factors and RNA polymerases, thus
enabling gene silencing. RBBP, RB binding protein. |
PRC2, consisting of core subunits SUZ12 polycomb
repressive complex 2 subunit (SUZ12), embryonic ectoderm
development (EED), RBBP4/7 and enhancer of zeste homolog (EZH)2 or
EZH1, is the sole confirmed methyltransferase responsible for the
mono-, di- and trimethylation of H3K27, generating the H3K27me3
mark (52). RBBP4/7 interact
with various molecules to facilitate PRC2 recruitment and activity
modulation. Studies have shown that RBBP4 can recruit SUZ12 to PRC2
target sites, and methylate H3K27 or H1K26 with the histone lysine
N-methyltransferase EZH2 (45,53). Simultaneously, EED can interact
with the H3K27me3 mark, thereby activating the methyltransferase
activity of EZH2 and influencing the overall activity of PRC2.
Consequently, this process promotes the 'spreading' of H3K27me3
(54) (Fig. 3B). Notably, the absence of the
SUZ12-RBBP4 complex influences H3K27me3 (55). Therefore, as pivotal constituents
of PRC2, RBBP4/7 emerge as determinants of site-specific H3K27me3
and other histone methylations across the genome.
RBBP4/7 also exhibit methyltransferase activity
independent of PRC2. Kitange et al (56) showed that RBBP4 knockdown
can enhance the levels of H3K9 trimethylation. RBBP4/7 interact
with SUV39H1 to specifically methylate lysine 9 on histone H3,
leading to heterochromatin silencing or RB transcriptional
repression (57). Furthermore,
RBBP7 inhibits DNA methyltransferase 1, affecting DNA methylation
and limiting transcription factor access to promoters (58).
In summary, RBBP4/7 serve an important role in
regulating histone deacetylation and methylation, which are key
processes in chromatin remodeling and the regulation of gene
expression. The precise mechanisms underlying these regulatory
activities of RBBP4/7 both inside and outside the aforementioned
complexes remain an area of ongoing research and exploration.
The role of RBBP4/7 in the DNA damage
response
RBBP4/7 have been identified as crucial components
of several protein complexes involved in DNA repair, making them
essential players in maintaining genome integrity.
The NuRD complex governs gene expression and DNA
damage repair by modulating nucleosome RNA polymerase accessibility
at transcription factor binding sites, enhancers and promoters
(59). RBBP4/7, as subunits of
the NuRD complex, mediate the interaction of NuRD with histone
tails and transcription factors (59). Yang et al (60) showed that breast cancer (BC)
anti-estrogen resistance 1 and RBBP4 can form a complex, be
recruited to chromatin, and jointly occupy the promoter regions of
some DNA repair genes, and promote DNA damage repair. Similarly,
RBBP4/7 specifically interact with the C-terminal domain of BC type
1 susceptibility protein (BRCA1) and inhibit its transactivation
activity (61). The association
between BRCA1 and RBBP7 is disrupted in cells treated with
DNA-damaging agents (62).
Therefore, the interaction between RBBP4/7 and BRCA1 might be the
key to regulating DNA damage repair.
Li et al (63) demonstrated that RBBP4 disruption
results in heightened DNA damage and apoptosis in glioblastoma
(GBM) cells post-temozolomide (TMZ) and radiotherapy. Additionally,
in MCF10AT3B cells, which are neoplastigenic breast epithelial
cells derived from a model of human proliferative breast disease,
high levels of RBBP7 might induce the growth arrest- and DNA
damage-induced (GADD) gene, GADD45 (64). Consequently, aberrant expression
of RBBP4/7 has implications for the DNA damage repair response.
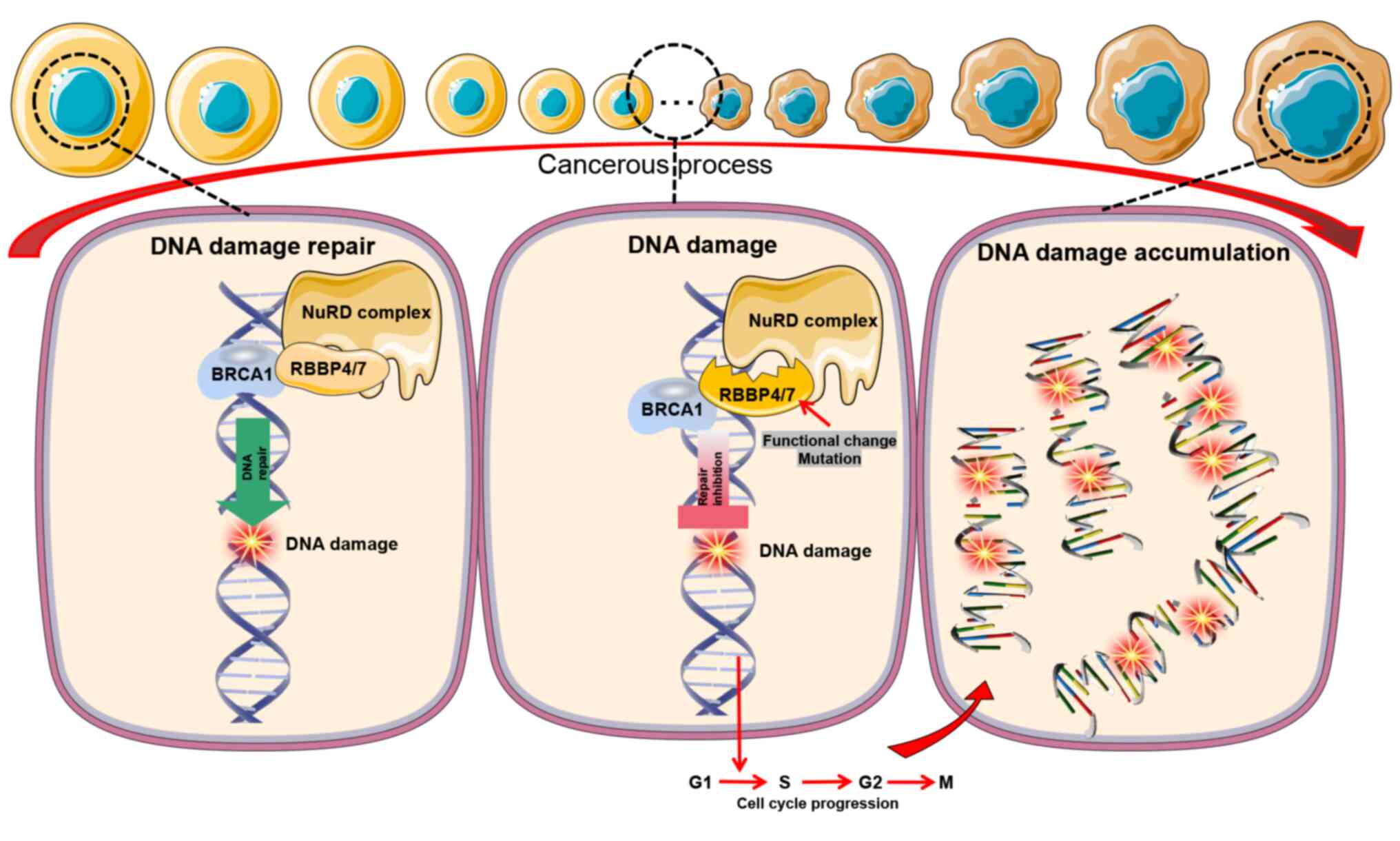
In summary, RBBP4/7 serve a pivotal role in DNA
damage repair and gene regulation, particularly in collaboration
with BRCA1. Their interaction with BRCA1 facilitates DNA repair,
and aberrant expression may affect this response, leading to the
accumulation of DNA damage, as cell cycle progression and increased
carcinogenic risk (Fig. 4). The
complexity of these interactions warrants further
investigation.
The role of RBBP4/7 in cell development,
differentiation, maturation and senescence
In mouse oocytes, RBBP4 is crucial for bipolar
spindle formation, and its deficiency can lead to mitotic
abnormalities (49), suggesting
its importance in cell division, a fundamental process of cell
growth. RBBP7 is strongly expressed in the kidneys and brain from
embryonic day 9.5 (65),
regulating Kisspeptin-1 expression to participate in reproductive
development (66). Giri et
al (67) revealed that RBBP7
can be suppressed in relation to Ras-associated cell proliferation
through stabilization by small ubiquitin like modifier 1.
RBBP4 is indispensable in heterochromatin assembly
and serves as a crucial barrier in inducing cell fate transition
from pluripotency to totipotency. Ping et al (68) demonstrated that the deletion of
RBBP4 enhances the transition of mouse embryonic stem cells to
trophectoderm cells. In the NuRD complex, RBBP4/7 and MTA interact
with friend of GATA protein 2 (FOG-2) and are involved in
FOG-2-mediated inhibition of GATA binding protein 4 activity,
preventing the aberrant cell differentiation that leads to cardiac
malformations (69). Moreover,
the RBBP4 homolog DjRbAp48 in planarians (Dugesia japonica)
regulates stem cell differentiation (70). In addition, notable discoveries
have been made in the study of RBBP7. RBBP7 is involved in
regulating histone acetylation and the expression of cyclin D3 in
post-implantation trophoblast matrix cells (71), and it interacts with the
pregnancy-induced non-coding RNA, inhibiting the differentiation of
alveolar cells during pregnancy (72). Xin et al (73) showed that RBBP4/7 may be
indirectly involved in the differentiation process of bone marrow
cells by affecting the expression of the long noncoding RNA
HOTAIRM1. Finally, in kidney development, RBBP7, as a target
gene of the transcription factor Wilms tumor 1, exhibits decreased
expression, reflecting podocyte dedifferentiation (74). These data suggested that RBBP4/7
may have a key role in cell differentiation processes.
During cell growth, RBBP4 influences cell morphology
and cytoskeleton organization by enhancing K-Ras activity and
mitogen-activated protein kinase signaling (75). Gasca et al (76) proposed that RBBP7 is involved in
the maturation of oocytes. Likewise, RBBP7 also contributes to
histone deacetylation during oocyte maturation (77). By contrast, Guan et al
(78) showed that RBBP7 has
strong growth inhibitory activity in the developing kidney and
gonads. In summary, RBBP4/7 play diverse roles in cell maturation,
influencing cell morphology, oocyte maturation and organ
growth.
In aging human fibroblasts, a decrease in RBBP4
expression leads to chromatin defects (79). Concurrently, Hunt et al
(80) demonstrated that
ubiquitin protein ligase E3 component N-recognin 4 deficiency
prevents skeletal muscle cell aging and atrophy by reducing the
ubiquitination and degradation of the HAT1/RBBP4/RBBP7
histone-binding complex. Additionally, decreased RBBP4 expression
in the aging hippocampus is associated with memory loss (81). Tsujii et al (82) further illustrated that
RBBP4 knockdown might inhibit nuclear transport and induce
cellular aging. Furthermore, RBBP7 has been reported to be
consistently upregulated in the lobules of degenerated mammary
glands and to be associated with hormone induction (83). Collectively, RBBP4/7 are closely
associated with cellular senescence, affecting chromatin defects,
skeletal muscle cell aging and memory loss.
In summary, RBBP4/7 exhibit diverse functions
throughout cell growth and development, impacting essential
cellular processes and providing valuable insights into cellular
differentiation, aging and chromatin regulation.
4. Expression and function of RBBP4/7 in
diseases
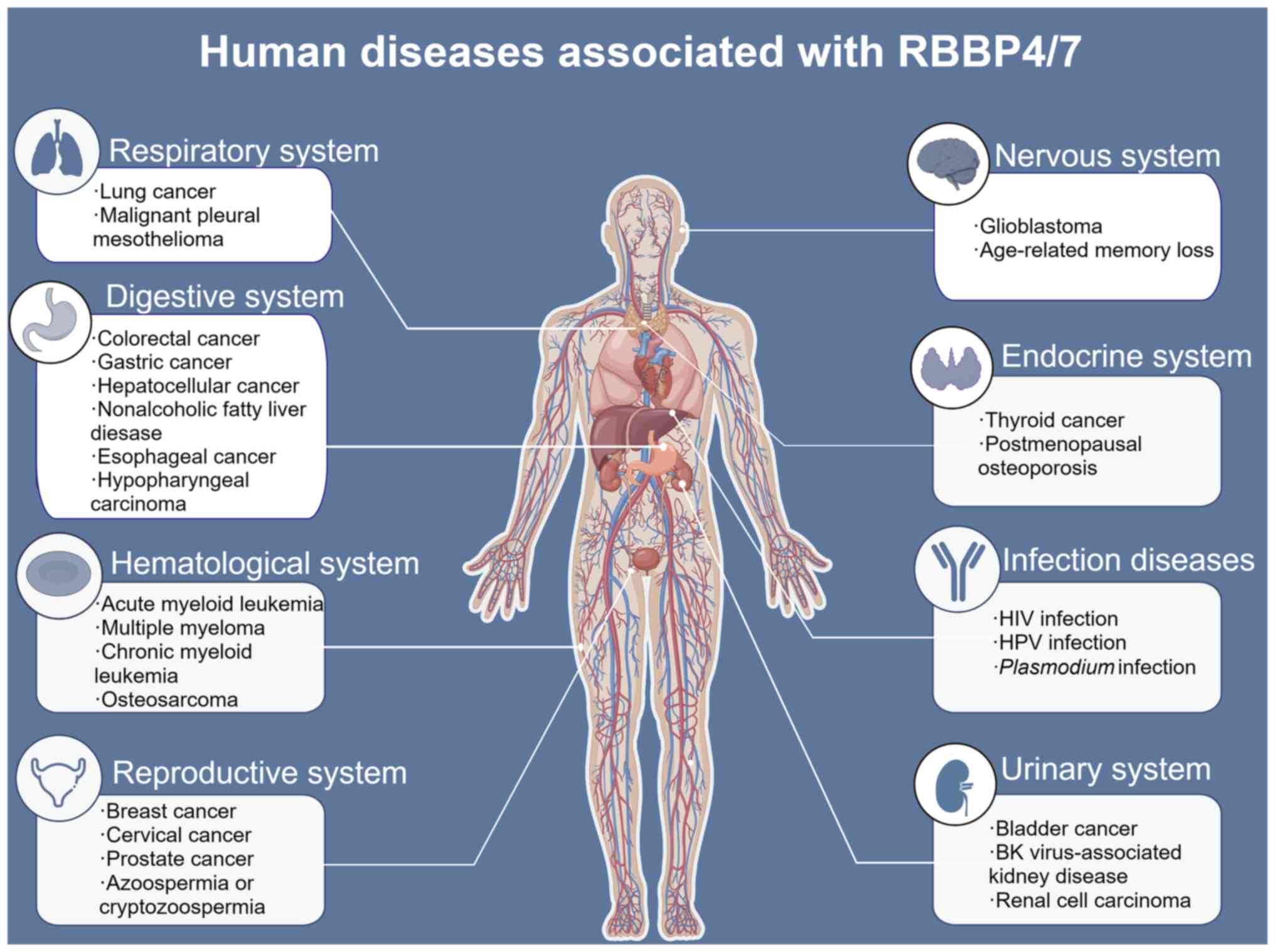
Aberrant expression of RBBP4/7 has been observed in
several diseases. The present review summarizes the various
diseases regulated by RBBP4/7 (Fig.
5), the clinical characteristics of RBBP4 and RBBP7 in human
diseases (Tables I and II), and their roles and mechanisms in
disease development (Tables
III and IV).
 | Table IExpression of RB binding protein 4 in
human diseases and relative clinical significance. |
Table I
Expression of RB binding protein 4 in
human diseases and relative clinical significance.
| First author,
year | Disease type | Expression | Samples | Clinical
characteristics | (Refs.) |
|---|
| Gao M, 2023 | NSCLC | Upregulated | LUAD: 54 adjacent
normal, 497 tumor tissues; LUSC: 49 adjacent normal, 502 tumor
tissues | Poor prognosis | (84) |
| Cao X, 2021 | NSCLC | Upregulated | / | / | (13) |
| Hao D, 2023 | NSCLC | Upregulated | 54 NSCLC tissues
and 54 adjacent normal tissues | / | (85) |
| Wang N, 2021 | NSCLC | / | mRNA expression
profiles of 43 patients with NSCLC | Poor prognosis,
tumor recurrence | (86) |
| Vavougios GD,
2015 | MPM | Upregulated | 40 MPM tissues and
9 control tissues (5 pleura tissues and 4 lung tissues) | / | (92) |
| Shou J, 2021 | GBM | Upregulated | 33 GBM tissues and
adjacent normal tissues | / | (94) |
| Li J, 2023 | GBM | Upregulated | / | Poor prognosis | (63) |
| Li D, 2018 | NB | Upregulated | Tissues from 42
primary cases of NB | Poor prognosis,
tumor stage, survival rate | (97) |
| Pavlopoulos E,
2013 | Age-related memory
loss | Downregulated | Entorhinal cortex
and DG of 10 healthy human brains, mouse DG tissues | Memory loss | (81) |
| Kosmidis S,
2018 | Discriminative | Downregulated | Mouse DG tissues
memory deficits | Discriminative
memory, spatial memory | (100) |
| Khateb A, 2021 | AML | Upregulated | / | Overall survival,
tumor development | (106) |
| Casas S, 2003 | AML | Upregulated | Bone marrow
aspirate of 15 patients with AML and 5 healthy individuals | / | (107) |
| Sakhinia E,
2005 | AML | Upregulated | Bone marrow
aspirate of 26 patients with AML, 12 patients with AML in remission
and 9 individuals with morphologically normal bone marrow | AML remission | (108) |
| Sakhinia E,
2005 | ALL | Upregulated | Bone marrow
aspirate of 5 patients with ALL and 9 individuals with
morphologically normal bone marrow | / | (108) |
| Li YD, 2019 | CRC | Upregulated | Colon cancer
tissues, para-colon cancer tissues and haptic metastatic cancer
tissues from 80 patients with CRC | Haptic metastases,
poor prognosis | (117) |
| Ding L, 2019 | GC | Upregulated | 142 GC tissues and
adjacent normal tissues | / | (120) |
| Song H, 2004 | HCC | Upregulated | Tissue from a
patient with primary HCC | / | (124) |
| Zhi S, 2022 | NAFLD | Downregulated | Liver tissues of
patients with NAFLD | / | (130) |
| Chen L, 2022 | ESCC | Upregulated | ESCC tissues and
corresponding normal tissues from 111 patients | / | (131) |
| Pacifico F,
2007 | TC | Upregulated | / | / | (135) |
| Guo Q, 2020 | BC | Upregulated | 240 BC tumor
tissues | Overall survival,
lymph node metastasis, tumor development | (141) |
| Gong X, 2020 | BC | Upregulated | / | / | (142) |
| Zheng Z, 2022 | TNBC | Upregulated | / | / | (144) |
| Barreiro-Alonso A,
2021 | PCa | Upregulated | 494 prostate
adenocarcinoma tissues | Progression-free
survival | (160) |
| Lohavanichbutr P,
2009 | OSCC | Upregulated | 124 OSCC patient
tissues and 45 normal tissues | Radiosensitivity,
chemosensitivity | (174) |
| Wurlitzer M,
2020 | HPV-positive
OPSCC | Upregulated | 8 HPV-positive and
9 HPV-negative oropharyngeal tumor tissues | / | (175) |
| Table IIExpression of RB binding protein 7 in
human diseases and relative clinical significance. |
Table II
Expression of RB binding protein 7 in
human diseases and relative clinical significance.
| First author,
year | Disease type | Expression | Samples | Clinical
characteristics | (Refs.) |
|---|
| Wang CL, 2009 | NSCLC | Upregulated | 154 lung cancer
tissues and adjacent normal tissues | Distant
metastasis | (90) |
| Wang H, 2022 | LUAD | Upregulated | 334 LUAD samples
and 59 adjacent normal lung samples, and 23 cancer tissues and
adjacent normal tissues from patients with LUAD | Relapse-free
survival, poor prognosis, TNM stage | (89) |
| Zhu H, 2022 | LUAD | / | / | Poor prognosis | (88) |
| Vavougios GD,
2015 | MPM | Upregulated | 40 MPM tissues and
9 control tissues (5 pleura tissues and 4 lung tissues) | / | (92) |
| Crea F, 2010 | Anaplastic
astrocytoma | Upregulated | / | / | (98) |
| Crea F, 2010 | Anaplastic
oligodendroglioma | Upregulated | / | / | (98) |
| Dave N, 2021 | AD | Downregulated | 89 AD brain tissues
and 98 normal brain tissues | CERAD (neuritic
plaque density), Braak stage, brain weight | (105) |
| Hu SY, 2005 | AL | Upregulated | Bone marrow cells
from 98 patients with AL, 5 patients with relapsing AL, 8 patients
with CR-AL and 32 healthy individuals | / | (113) |
| Hu SY, 2005 | CML-BC | Upregulated | Bone marrow cells
from 13 patients with CML-CP, patients with CML-BC and 32 healthy
individuals | Tumor
progression | (113) |
| Yu N, 2018 | ESCC | Upregulated | 126 ESCC tissues,
72 of which had adjacent non-neoplastic tissues | Poor
differentiation, lymph node invasion and progression, pathological
TNM stage, poor prognosis, overall survival | (14) |
| Wang R, 2022 | EC | Upregulated | 182 EC tissues and
286 normal tissues | Overall survival,
relapse-free survival, tumor stage | (133) |
| Thakur A, 2007 | BC | Upregulated | 20 breast cancer
and adjacent benign or normal breast tissue | / | (150) |
| Ebata A, 2012 | pDCIS | Upregulated | 53 pDCIS and 27 IDC
tissues | / | (151) |
| Barreiro-Alonso A,
2021 | PCa | Upregulated | 494 prostate
adenocarcinoma tissues | Progression-free
survival | (160) |
| Riera-Escamilla A,
2022 | Azoospermia | / | X-linked
protein-coding genes in 2,354 men with idiopathic
NOA/cryptozoospermia |
Spermatogenesis | (163) |
| Yeh HH, 2015 | Bladder cancer | Upregulated | Tissues from 4
patients with clinical bladder cancer | / | (166) |
| Wang Y, 2022 | BKVN | Upregulated | / | Immune cell
infiltration, graft rejection, diagnosis | (167) |
| Wurlitzer M,
2020 | HPV-positive
OPSCC | Upregulated | 8 HPV-positive and
9 HPV-negative oropharyngeal tumor tissues | / | (175) |
| Table IIIThe functions and mechanisms of RB
binding protein 4 in diseases. |
Table III
The functions and mechanisms of RB
binding protein 4 in diseases.
| First author,
year | Disease type | Cell lines | Role | Functions | Upstream
regulators | Target/interacting
genes | (Refs.) |
|---|
| Cao X, 2021 | NSCLC | A549, HC827 | Oncogene | Promotes cell
proliferation, invasion and migration | hsa_circ_0102231,
miR-145 | / | (13) |
| Hao D, 2023 | NSCLC | A549, H1299 | Oncogene | Promotes cell
proliferation, migration, invasion, glycolysis and drug
resistance | circ_0110498,
miR-1287-5p | / | (85) |
| Jin X, 2022 | LUAD | A549, H1299 | Oncogene | / | / | CBX3, ARHGAP24 | (87) |
| Shou J, 2021 | GBM | U87, U251 | Oncogene | Promotes cell
proliferation, viability and adhesion, inhibits cell apoptosis | HOXA-AS2,
miR-885-5p | / | (94) |
| Mladek AC,
2022 | GBM | U251 | Oncogene | TMZ resistance | / | p300, c-MYC,
RAD51 | (95) |
| Kitange GJ,
2016 | GBM | GBM12, GBM22 | Oncogene | Promotes tumor
growth in orthotopic xenografts and TMZ resistance | / | p300, CBP, RAD51,
MGMT | (56) |
| Li J, 2023 | GBM | U87MG, T98G, U118,
U251 | Oncogene | Reduces sensitivity
to RT and TMZ, promotes DNA damage repair | / | MRE11, RAD50, NBS1,
ELF-1 | (63) |
| Li D, 2018 | NB | BE (2)-C, IMR32,
SH-SY5Y | Oncogene | Promotes cell
growth and invasion | / | ARMC12 | (97) |
| Kosmidis S,
2018 | Age-related memory
loss | / | / | Maintains normal
brain function | / | GPR158, BDNF | (100) |
| Khateb A, 2021 | AML | MOLM-13, U937 | Oncogene | Promote cell
growth | RNF5 | ANXA1, NCF1,
CDKN1A | (106) |
| Hu SY, 2006 | CML | K562 | Tumor
suppressor | Inhibits cell
growth, arrests cell cycle | / | IGFBP-rP1 | (111) |
| Zhu X, 2023 | CRC | HT29, HCT116, LOVO,
SW620, RKO | Oncogene | Promotes cell
proliferation and invasion | circAGO2,
miR-1-3p | HSPB8 | (118) |
| Li YD, 2020 | CRC | SW620, HT29, LoVo,
SW480, HCT-116 | Oncogene | Promotes cell
proliferation, migration and invasion, inhibits cell apoptosis | / | Wnt/β-catenin | (12) |
| Zhuo FF, 2022 | CRC | HCT116 | Oncogene | Promotes cell
proliferation and migration | / | / | (119) |
| Ding L, 2019 | GC | BGC-823, AGS | Oncogene | Promotes cell
growth and reduces apoptosis | circ-DONSON | SOX4 | (120) |
| Jin X, 2018 | GC | AGS | Tumor
suppressor | Promotes apoptosis,
inhibits cell proliferation, enhances radiosensitivity and cell
cycle arrest | / | PI3K/Akt | (121) |
| Song H, 2004 | HCC | L02, Bel-7404,
HepG2, Bel-7402 and HuH7 | / | / | / | / | (124) |
| Li L, 2015 | HCC | EPCAM+
and EPCAM− HCCLM3 | Oncogene | Promotes cell
proliferation, self-renewal, chemotherapy resistance and
tumorigenesis | miR-429 | E2F1, OCT4 | (126) |
| Zhi S, 2022 | NAFLD | Mouse AML12
hepatocytes | Reduces hepatic
steatosis | Regulates lipid
metabolism and reduces lipid accumulation in liver cells | / | Cpt1α, Acox1 | (130) |
| Chen L, 2022 | ESCC | Kyse150, Kyse170,
Eca109, TE1 | Oncogene | Promotes EMT
transition | KTN1-AS1 | HDAC1 | (131) |
| Bai X, 2015 | Hypopharyngeal
carcinoma | FaDu | Tumor
suppressor | Inhibits cell
proliferation, colony formation and tumor formation. Promotes
apoptosis and regulates tumor suppressors | / | p53, RB, Bax,
caspase-3, caspase-8, caspase-9 | (132) |
| Pacifico F,
2007 | TC | FRO | Oncogene | Promotes cell
proliferation and cell cycle arrest | NF-κB | / | (135) |
| Yang C, 2019 | PMOP | / | / | Regulates
mitochondrial function | / | ESR1 | (138) |
| Ishimaru N, 2006;
Ishimaru N, 2008 | Autoimmune exocrine
disease | / | / | Promotes exocrine
cell apoptosis | / | p53 | (139, 140) |
| Gong X, 2020 | BC | MCF-7,
MDA-MB-231 | Oncogene | Promotes cell
proliferation, migration and invasion, inhibits cell apoptosis | / | LCPAT1, MFAP2 | (142) |
| Creekmore AL,
2008 | BC | MCF-7 | / | Regulates the
expression of estrogen-responsive genes and estrogen signaling | / | ERα | (143) |
| Zheng Z, 2022 | TNBC | / | Oncogene | Promotes cell
proliferation, invasion and migration, and regulates EMT
activity | / | / | (144) |
| Moody RR, 2018 | TNBC | / | Oncogene | Promotes
tumorigenesis | / | BCL11A | (145) |
| Kong L, 2007 | Cervical
cancer | Caski, H8 | Tumor
suppressor | Inhibits cell
proliferation and colony formation, promotes cell senescence and
prevents tumor formation | / | RB, p53, caspase-3,
caspase-8, E6, E7, CCND1, c-MYC | (153) |
| Wu S, 2017 | Cervical
cancer | SiHa, HeLa | Tumor
suppressor | Promotes cell
apoptosis and inhibits cell proliferation | / | HPV E6/E7, RB, p53,
caspase-3 | (155) |
| Zhong J, 2015 | Cervical
cancer | MS751 | Oncogene | Promotes cell
migration and invasion, inhibits EMT | / | SNAIL, TWIST | (156) |
| Zheng L, 2013 | Cervical
cancer | SiHa, Caski,
HeLa | Tumor
suppressor | Promotes apoptosis,
inhibits cell proliferation and enhances radiosensitivity | / | / | (157) |
| Cai L, 2014 | PCa | LNCaP | Tumor
suppressor | Promotes apoptosis
and inhibits cell proliferation | / | EAF2 | (159) |
| Wang J, 2016 | HIV infection | 293T, TZM-bl,
CEM-ss | Antiviral | Inhibits the
expression of HIV-1 | / | / | (169) |
| Wang J, 2019 | HIV infection | HIV-1 infected T
cells, J-lat | Antiviral | Suppresses
transcription and remodels chromatin | / | NR2F1, HDAC1/2 | (170) |
| Biswas S, 2018 | HIV infection | HIV-2-infected
MDMs | / | / | / | / | (171) |
| Xu W, 2020 | HIV infection | 293T | antiviral | Inhibits the
activity of LTR, affect the nuclear translocation of p65 | / | P65, NF-κB
pathway | (172) |
| Table IVThe functions and mechanisms of RB
binding protein 7 in diseases. |
Table IV
The functions and mechanisms of RB
binding protein 7 in diseases.
| First author,
year | Disease type | Cell lines | Roles | Functions | Upstream
regulators | Target/interacting
genes | (Refs.) |
|---|
| Wang CL, 2009 | NSCLC | CL1-0, CL1-5 | Oncogene | Promotes cell
migration | / | / | (90) |
| Dave N, 2021 | AD | HT-22 | / | Inhibits cell
death | / | tau, p300 | (105) |
| Duan WM, 2004 | / | U937 | Tumor
suppressor | Inhibits leukemic
tumor growth | / | / | (114) |
| Zhang TF, 2003 | Osteosarcoma | Saos-2 | Tumor
suppressor | Promotes apoptosis
and inhibits the growth of tumor grafts | / | JNK | (116) |
| Guo L, 2023 | CRC | / | Tumor
suppressor | Promotes
apoptosis | / | p53, FAS, FIT | (15) |
| Yu N, 2018 | ESCC | TE1, KYSE30,
KYSE150, Eca109 | Oncogene | Promotes cell
invasion and migration | / | / | (14) |
| Wang R, 2022 | EC | Eca109,
KYSE450 | Oncogene | Promotes cell
viability and proliferation | HIF1α | CDK4 | (133) |
| Xie ZF, 2020 | EC | ECA-109,
KYSE-510c | Oncogene | Promotes cell
proliferation, migration, invasion and glycolysis | circ_0006168,
miR-384 | S6K/S6, | (134) |
| Creekmore AL,
2008 | BC | MCF-7 | / | Regulates the
expression of estrogen-responsive genes and estrogen signaling | / | ERα | (143) |
| Moody RR, 2018 | TNBC | / | Oncogene | Promotes
tumorigenesis | / | BCL11A | (145) |
| Zhang TF, 2003 | BC | MCF-7, MDA-MB-231,
MDA-MB-43 | Tumor
suppressor | Inhibits tumor
formation | / | / | (146) |
| Zhang TF, 2007 | BC | MCF10AT3B | Tumor
suppressor | Inhibits tumor
formation and cell proliferation | / | β-catenin | (147) |
| Li Gc, 2003 | BC | MCF10AT3B | Tumor
suppressor | Promotes apoptosis
and inhibits cell growth | / | JNK | (64) |
| Li GC, 2006 | BC | MCF10AT3B | Oncogene | Promotes cell
migration, invasion and EMT | / | N-cadherin,
E-cadherin, α/β/γ-catenin | (149) |
| Fu J, 2011 | BC | MDA-MB-435 | Oncogene | Promotes cell
migration, invasion and EMT | / | E-cadherin,
TWIST | (148) |
| Li HJ, 2016 | Cervical
cancer | HeLa, SiHa | Tumor
suppressor | Inhibits cell
invasion and EMT | NKX6.1 | Vimentin,
N-cadherin | (158) |
| Cai L, 2014 | PCa | LNCaP | Tumor
suppressor | Promotes apoptosis
and inhibits cell proliferation | / | EAF2 | (159) |
| Wang J, 2020 | PCa | DU145 | Tumor
suppressor | Inhibits cell
migration and EMT | / | HNF1B | (161) |
| Yeh HH, 2015 | Bladder cancer | T24, NIH3T3 | Oncogene | Promotes cell
invasion and tumor metastasis | Ras | RECK, MMP-9, HDAC1,
Sp1 | (166) |
Respiratory system Lung cancer (LC)
RBBP4 is associated with genetic susceptibility to
LC (84). Elevated RBBP4
expression in non-small cell LC (NSCLC) tissues amplifies cell
proliferation and invasion, and is concomitantly linked to an
adverse clinical outcome (13,84). RBBP4 is also increased in
cisplatin-resistant NSCLC, affecting drug resistance (85), and is associated with enhanced
DNA damage sensitivity and repair pathway activity (86). Additionally, in lung
adenocarcinoma (LUAD) cells, RBBP4 interacts with chromobox homolog
3, which is found to be upregulated in current smokers with LUAD,
thereby promoting LUAD progression (87). Thus, RBBP4 could be a potential
biomarker or therapeutic target for LUAD recurrence and prognosis
post-platinum treatment.
Research has revealed that the expression levels of
RBBP7 in NSCLC are higher than those in normal lung tissues, and
this elevated expression is associated with distant metastasis,
poor prognosis and tumor immune response in NSCLC, serving as a
predictor for the recurrence of early-stage NSCLC (88-90).
Despite these significant findings, the
understanding of the functions of RBBP4/7 in LC is limited and more
in-depth research is needed to develop new treatment
strategies.
Malignant pleural mesothelioma (MPM)
MPM has an average survival of 1 year
post-diagnosis, urgently necessitating improved treatment methods
(91). Vavougios et al
(92) showed that RBBP4/7
interact with Parkinson disease protein 7 and are upregulated in an
array of 18 different sarcoma types. However, the mechanism by
which RBBP4/7 functions in MPM remains unknown.
Nervous system
GBM and other brain tumors
RBBP4 plays an indispensable role in the disease
development of GBM (93), with
its mRNA expression universally upregulated in GBM tissues where it
acts as an oncogene to promote GBM malignancy, countering the
tumor-suppressive effects of microRNA (miR)-885-5p (94). In GBM cells, the RBBP4/p300
complex governs pro-survival genes and influences the
responsiveness to TMZ (95).
This suggests that disrupting RBBP4 or p300 might enhance
sensitivity to TMZ. In addition, RBBP4 increases TMZ resistance by
regulating the expression of the MRN complex (MRE11 homolog, RAD50
double-strand break repair protein, and nibrin), thereby reducing
sensitivity to radiotherapy and TMZ (63). Therefore, RBBP4 potentially
enhances cancer progression or drug resistance through DNA repair
and might be considered a new therapeutic target for future GBM
treatment.
RBBP4 is required during brain development, and
RBBP4 is upregulated in RB1-mutated embryonic brain tumors, serving
as a potential target for inducing apoptosis in RB1-mutated brain
cancer cells (96).
Additionally, in neuroblastoma, RBBP4 is upregulated and is
associated with poor patient prognosis (97).
By contrast, limited research has been conducted on
the involvement of RBBP7 in GBM. Notably, Crea et al
(98) explored the Oncomine
database and observed an upregulation of RBBP7 in anaplastic
astrocytoma and anaplastic oligodendroglioma.
Age-related memory loss
Loss of RBBP4 is key to age-related memory decline.
Its expression in human and mouse brains declines with age,
affecting memory formation (81). Another study demonstrated that
RBBP4 regulates the expression of brain-derived neurotrophic factor
and G protein-coupled receptor 158, key components of the mouse
hippocampal osteocalcin (OCN) signaling pathway (99). Inhibition of RBBP4 disrupts the
cognitive benefits of OCN and leads to discriminative memory
deficits (100). Furthermore,
certain genetic variants in a cAMP element binding
protein-dependent histone acetylation pathway, associated with
RBBP4, influence memory performance in cognitively healthy elderly
individuals (101). Therefore,
RBBP4 could serve as a potential therapeutic target for age-related
memory loss.
Current research has explored RBBP4/7 as therapeutic
targets to address age-related memory loss. The functional role of
RBBP4 in Alzheimer's disease (AD) might be influenced by the
instability of the RBBP4-FOG1 complex (102). Huang et al (103) identified three traditional
Chinese medicine compounds (bittersweet alkaloid ii, eicosanedioic
acid and perivine), which could enhance the stability of the
RBBP4-FOG1 complex, offering potential therapeutic benefits for AD.
However, another study showed that RBBP4/7 did not contribute to
the neuroprotective effects of green tea polyphenols (104). Thus, the mechanism of RBBP4/7
as a target for age-related memory loss requires further
investigation.
Dave et al (105) discovered that RBBP7 mRNA
expression is diminished in AD cases, with significant negative
correlations with the Consortium to Establish a Registry for AD and
Braak stage. Moreover, this previous study revealed that high RBBP7
expression mitigates tau acetylation and phosphorylation, thereby
preventing tau pathologies (105).
In summary, RBBP4/7 play crucial roles in
age-related memory deficits, and present promising therapeutic
targets for future interventions in cognitive aging and associated
diseases.
Hematological (blood and bone marrow)
system diseases Acute myeloid leukemia (AML)
In AML, elevated RBBP4 expression is linked to
poorer survival and disease progression (106-108). Moreover, AML primary blasts
with lower levels of ring finger protein 5/RBBP4 have demonstrated
increased sensitivity to the HDAC inhibitor FK228. These findings
suggest that the abundance of RBBP4 may serve as valuable marker to
stratify patients with AML who might benefit from treatment with
HDAC inhibitors (106).
However, to the best of our knowledge, there is no currently
research indicating an association between RBBP7 and AML.
Multiple myeloma (MM)
Gao et al (109) showed that RBBP4 is a
core gene in MM, and its dysregulation is evident in the epigenetic
modifications of MM. Thus, RBBP4 emerges as a potential focal point
in MM research, providing promising avenues to understand the
pathogenesis of MM and develop more effective therapeutic
strategies. However, to the best of our knowledge, no studies have
revealed a role for RBBP7 in MM.
Chronic myeloid leukemia (CML)
CML is a malignant hematological disorder. It has
been shown that BMI1 (encoding B lymphoma Mo-MLV insertion
region 1 homolog) transcript levels are significantly increased in
CML cells, and the expression of RBBP4 is decreased after
BMI1 silencing (110).
In addition, RBBP7 in K562 leukemic cells inhibits growth by
inducing IGFBP7 expression (111). These findings revealed that
RBBP4/7 might serve an important role in the biological process of
CML. However, the expression levels of RBBP7 were significantly
elevated in patients with acute leukemia and CML in blast crisis
compared with healthy donors and those with CML in chronic phase,
indicating that it might be involved in the occurrence of leukemia
(112,113). In addition, RBBP7
overexpression slows growth in the U937 leukemia cell line
(114).
Osteosarcoma
Osteosarcoma originates from primitive mesenchymal
cells in the bone, rarely in soft tissue, and if untreated, can
lead to local and often metastatic progression (115). Zhang et al (116) showed that the inducible
expression of RBBP7 can activate the c-Jun N-terminal kinase
signaling pathway, and trigger apoptosis in Saos-2 osteosarcoma
cells, while also strongly suppressing the formation of tumor
grafts in nude mice and significantly reducing the growth of
established osteosarcoma xenografts.
Digestive system
CRC
RBBP4 has been identified as a key factor in CRC,
with studies showing its upregulation in colon cancer tissues, and
linking increased RBBP4 expression to poor prognosis and liver
metastasis (117). Reducing
RBBP4 levels can hinder the growth and migration, and increase the
apoptosis of HCT116 and SW620 colon cancer cells, and also suppress
the Wnt/β-catenin pathway (12).
Concurrently, knocking down RBBP4 also inhibits H3K27ac
acetylation and HSPB8 gene transcription (118). Another study demonstrated that
RBBP4 is a key target of protopanaxadiol (PPD), a major ginseng
metabolite (119), suggesting
its potential as a new diagnostic and therapeutic target for
CRC.
RBBP7 has been reported to form a trimeric complex
with long non-coding RNA FIT and p53, enhancing p53-mediated FAS
gene transcription, which promotes CRC cell apoptosis (15). This suggests the
apoptosis-inducing role of RBBP7 in CRC, which differs from the
role of RBBP4. However, the precise expression and mechanism of
RBBP7 in CRC warrant further investigation.
Gastric cancer (GC)
Ding et al (120) revealed that the expression of
RBBP4 is increased in GC tissues, and knocking down RBBP4
can significantly inhibit GC cell proliferation, migration and
invasion, and promote cell apoptosis. Radiation can also increase
RBBP4 expression in AGS GC cells, leading to G2 phase
arrest. Moreover, RBBP4 enhances the radiosensitivity of these
cells by inhibiting the PI3K/Akt pathway (121). These results underscore the
potential of RBBP4 as a promising target for future gene therapy
interventions in the treatment of GC.
Currently, research on the role of RBBP7 in GC is
limited. Src-suppressed C-kinase substrate (SSeCKS), a crucial
substrate for protein kinase C, is significantly downregulated in
GC (122). Liu et al
(123) demonstrated that the
re-expression of SSeCKS induces RBBP7, suggesting a potential
connection. However, the specific function and significance of
RBBP7 in GC remains to be further determined.
Hepatocellular carcinoma (HCC)
It has been shown that RBBP4 is highly expressed in
liver tumor tissues, and is associated with clinical severity and
disease prognosis (124). In
HCC cells, Liu et al (125) identified that RBBP4 interacts
with the N-terminal peptide of Sal-like protein 4 (SALL4),
contributing to the silencing of tumor suppressor genes, such as
PTEN. Furthermore, a potent SALL4 peptide antagonist (FFW)
targeting RBBP4 significantly inhibits HCC cell growth. These
studies have shown that RBBP4 plays a role in promoting HCC, which
is expected to be a potential target for future HCC treatment.
By contrast, Li et al (126) observed a decrease in RBBP4
expression in HCC tissues; it was revealed that RBBP4
knockdown may enhance the self-renewal and tumorigenic potential of
epithelial cell adhesion molecule-positive liver tumor-initiating
cells, suggesting that RBBP4 serves as a downstream target of
miR-429. Furthermore, epigallocatechin gallate (EGCG) can reverse
multidrug resistance (MDR) in HCC, and RBBP4 has been reported to
be significantly upregulated after EGCG treatment (127), suggesting that RBBP4 could be a
potential anti-MDR target in HCC.
Nonalcoholic fatty liver disease
Hepatic steatosis, crucial in nonalcoholic
steatohepatitis development, elevates the risk of cirrhosis and HCC
(128). Within the NuRF
complex, the subunit RBBP4 assumes a role in the regulation of
lipid droplet size by transcriptionally suppressing target genes
(129). Zhi et al
(130) suggested that RBBP4
exerts a favorable influence on liver cell steatosis by promoting
the expression of genes associated with fatty acid β-oxidation.
These findings illustrate a protective role of RBBP4 in the context
of hepatic steatosis.
Esophageal cancer (EC) and
hypopharyngeal carcinoma
Research has shown that RBBP4 is upregulated in
ESCC and promotes the epithelial-mesenchymal transition (EMT)
process (131). By contrast,
Bai et al (132)
demonstrated that in hypopharyngeal carcinoma, a distinct squamous
cell carcinoma impacting the upper aerodigestive tract,
RBBP4 overexpression curtails proliferation, colony
formation and tumorigenesis in the FaDu hypopharyngeal carcinoma
cell line. The role of RBBP4 in hypopharyngeal carcinoma growth
appears linked to its modulation of tumor suppressors. This
contrasting role of RBBP4, from facilitating tumor progression in
ESCC to inhibiting growth in hypopharyngeal carcinoma, demonstrates
its functional variability across different types of cancer,
highlighting its potential complexity as a therapeutic target.
Yu et al (14) observed that higher RBBP7
expression in ESCC tissues is correlated with poor differentiation,
advanced lymph node involvement, higher tumor-node-metastasis
stage, reduced survival, and increased cell invasion and migration.
Mechanistically, hypoxia can induce high expression of RBBP7, which
in turn upregulates CDK4 expression and promotes tumor progression
(133). Furthermore, RBBP7 is
upregulated in EC. As a target of miR-384, RBBP7 mRNA levels
are elevated by circ_0006168 through its interaction with miR-384.
This, in turn, promotes cell proliferation, migration, invasion and
glycolysis in EC (134).
Collectively, RBBP4/7 are crucial in EC progression
and prognosis, with their complex molecular interactions making
them promising for future therapeutic and diagnostic developments
in esophageal oncology.
Endocrine system
Thyroid cancer (TC)
Pacifico et al (135) detected increased RBBP4
expression in primary human TC via immunohistochemical analysis,
particularly in undifferentiated TC samples and cell lines. In
addition, RBBP4 knockdown was shown to reduce FRO anaplastic
thyroid carcinoma cell colony formation, suggesting RBBP4 as a
target of nuclear factor (NF)-κB and a potential therapeutic target
for NF-κB-dependent TC. Therefore, RBBP4 might be a potential
target for TC therapy, particularly for NF-κB-dependent cases.
Postmenopausal osteoporosis (PMOP) and
other estrogen-influenced conditions
PMOP represents a significant global public health
concern. RBBP4 expression is elevated in estrogen-deficient rats
and patients with PMOP, decreasing after treatment with Liuwei
Dihuang pills (136,137). Moreover, RBBP4 has been
reported to be upregulated in the blood of patients with PMOP,
suggesting that it could serve as a potential diagnostic biomarker
of PMOP (138). The interaction
between RBBP4 and estrogen receptor 1 (ESR1) is believed to serve a
crucial role in the pathogenesis of PMOP (138). In summary, RBBP4 serves as a
potential biomarker for PMOP diagnosis and its interaction with
ESR1 suggests a fundamental mechanism in PMOP pathogenesis.
Additionally, estrogen deficiency-induced
overexpression of RBBP4 triggers p53-mediated apoptosis in exocrine
cells, implying a link to autoimmune exocrine disorders in
postmenopausal women (139,140). Therefore, RBBP4 represents a
novel immunotherapeutic target for preventing the development of
sex-based autoimmune exocrine disorders.
Reproductive system
BC
RBBP4 is highly expressed in BC and is associated
with poorer overall survival and a greater likelihood of lymph node
metastasis (141). Knockdown of
RBBP4 inhibits the proliferation, migration and invasion of
BC cells, while affecting the transcription of tumor-related genes,
such as microfibril-associated protein 2, which is activated via
the interaction between RBBP4 and the long noncoding RNA
LCPAT1 (142). Another
study showed that RBBP4/7 interact with DNA-bound estrogen receptor
α to alter the expression of estrogen-responsive genes in MCF-7
cells (143). Thus, RBBP4 is
implicated in BC progression, in which it influences survival,
metastasis and estrogen-responsive gene expression.
RBBP4 expression has also been shown to be
significantly elevated in triple-negative BC (TNBC) cells and
tissues; and its knockdown markedly inhibits TNBC cell
proliferation, invasion and migration, and concurrently
downregulates EMT regulatory activities (144). In addition, Moody et al
(145) demonstrated that BCL11
transcription factor A (BCL11A), which possesses the ability to
promote BC progression, can interact with RBBP4/7; therefore,
targeting RBBP4-BCL11A binding may have therapeutic potential.
The role of RBBP7 in BC appears complex and
contradictory. Zhang et al (146) observed decreased expression
levels of RBBP7 in BC cell lines, and its dysregulation was shown
to contribute to BC tumorigenesis. Further research has revealed
that RBBP7 is related to estrogen regulation and may affect the
early development of BC (64,147). As a component of the Mi2/NuRD
complex, RBBP7 regulates TWIST-mediated repression of E-cadherin
expression and inhibits BC cell metastasis (148). By contrast, Li and Wang
(149) found that recombinant
RBBP7 induces EMT and enhances mammary epithelial cell migration.
Thus, the mechanism of RBBP7 in BC progression and metastasis
requires further investigation.
Notably, in contrast to the observation of Zhang
et al (146) of
decreased RBBP7 expression in BC, Thakur et al (150) showed that RBBP7 expression was
upregulated in 79% of BC cases and its expression was positively
correlated with malignancy. RBBP7 may also be involved in the
pathogenesis of estrogen receptor-positive pure ductal carcinoma
in situ (151). In
addition, Mieczkowska et al (152) found that RBBP7 was
downregulated in parental G-2 cells from the WAP-T transgenic
breast cancer line after surviving traditional cytotoxic
combination therapy, suggesting that RBBP7 might be considered a
potential therapeutic target for BC in the future.
In summary, both RBBP4 and RBBP7 have demonstrated
significant roles in the progression, metastasis and therapeutic
potentialities of BC; however, their precise mechanisms and
interactions in various BC subtypes warrant deeper exploration.
Cervical cancer
Kong et al (153) observed that RBBP4
overexpression inhibits cervical cancer growth and affects human
papillomavirus (HPV)16 transformation by regulating tumor
suppressors and oncogenes. Notably, 5-aminole-vulinic acid
photodynamic therapy (ALA-PDT), an effective treatment for
HPV-related conditions, has been reported to elevate RBBP4
expression in HPV16 immortalized cervical epithelial H8 cells
(154). A subsequent decrease
in RBBP4 can mitigate the inhibitory effects of ALA-PDT-induced
cell proliferation and apoptosis in cervical cancer cells (155). These studies demonstrated that
RBBP4 may function as a tumor suppressor in cervical cancer and
could serve as a promising therapeutic target for future cervical
cancer intervention.
However, studies have also indicated that RBBP4
promotes cervical cancer, influencing EMT and radiotherapy outcomes
(156,157). This implicates RBBP4 as a
prospective target to boost radiotherapeutic outcomes in patients
with cervical cancer. In addition, RBBP7 can be recruited by NK6
homeobox 1, thereby inhibiting the invasive ability of cervical
cancer cells (158).
In conclusion, RBBP4/7 have complex roles in
cervical cancer and may be potential therapeutic targets. However,
the role of RBBP4 in cervical cancer remains controversial, and its
mechanism requires further study.
Prostate cancer
Cai et al (159) showed that there is a physical
interaction between the tumor suppressor gene EAF2 (encoding
ELL associated factor 2) and RBBP4/7, where their overexpression
induces cell death in LNCaP prostate cancer cells. High RBBP4/7
expression in prostate adenocarcinoma is also linked to shorter
progression-free survival, with RBBP7 interacting with high
mobility group box 1 to regulate RNA processing (160). Furthermore, overexpression of
RBBP7 suppresses SLUG1/EMT in DU145 cells and exerts tumor
suppressive functions in presence of hepatocyte nuclear factor 1β
(161). Thus, RBBP4/7 are
pivotal in prostate cancer progression and potential therapeutic
targets.
Azoospermia or cryptozoospermia
Male infertility, affecting ~7% of men in the
general population, is often due to factors such as azoospermia or
cryptozoospermia (162). The X
chromosome is vital for male reproductive health. Riera-Escamilla
et al (163) linked
RBBP7 mutations to early spermatogenic failure in an
analysis of 2,354 men with azoospermia/cryptozoospermia, revealing
that these mutations were more prevalent in this infertile group
compared with in control individuals with normozoospermia. RBBP7
forms the CRL4B-RBBP7 complex with CUL4B (encoded by another
mutated gene found in infertile men), which contributes to the
degradation of HUWE1 and is associated with non-obstructive
azoospermia (164). In
conclusion, RBBP7 has a central role in male infertility,
highlighting the importance of genetic factors in reproductive
health.
Urinary system
Bladder cancer
Bladder cancer, affecting >440,000 individuals
annually worldwide (165),
shows high RBBP7 expression in specimens. Mechanistically, RBBP7
can bind to HDAC1 and specificity protein 1 (SP1), and then bind to
the RECK (encoding reversion inducing cysteine rich protein
with kazal motifs) promoter at the SP1 site, thereby inhibiting the
expression of RECK, which in turn leads to matrix
metalloproteinase-9 activation and metastasis, thereby
participating in Ras-induced experimental lung metastasis (166). Therefore, RBBP7 could be used
as a therapeutic target for Ras-related cancer; however, to the
best of our knowledge, there are currently no studies on RBBP4 in
bladder cancer.
BK virus-associated kidney
disease
Wang et al (167) demonstrated that RBBP7 is highly
enriched in BK virus-associated nephropathy (BKVN) tissues and is
associated with alterations in various immune cells, such as CD8
naïve cells, induced regulatory T cells, neutrophils and
CD8+ T cells. Furthermore, RBBP7 serves as a molecular
biomarker for the precise diagnosis of BKVN, effectively
distinguishing transplant rejection responses. Thus, targeting
RBBP7 as a diagnostic tool may offer novel therapeutic and
prognostic opportunities for BKVN in transplant recipients.
Renal cell carcinoma (RCC)
Kim et al (168) showed that RBBP7 is highly
expressed in the chromaffin subtype of RCC, but not in traditional
RCC. Therefore, RBBP7 could be used as a candidate biomarker in
RCC, and its existence and expression patterns might be related to
the pathological characteristics of RCC subtypes, providing a novel
direction for the diagnosis and treatment of RCC.
Infectious diseases
HIV infection
Wang et al (169) reported increased RBBP4
expression following HIV-1 infection in cell culture models, with
RBBP4 knockdown enhancing HIV infection and viral
production. RBBP4 suppresses HIV-1 transcriptionally by binding to
its long terminal repeats, recruiting nuclear receptor subfamily 2
F group member 1 and HDAC1/2, leading to H3 deacetylation and
replication control (170).
Similarly, Biswas et al (171) observed elevated RBBP4 levels in
HIV-2-infected monocyte-derived macrophages, and Xu et al
(172) reported that
thieno[3,4-d] pyrimidine treatment in infected cells increases
RBBP4 levels and activates the NF-κB pathway, suppressing HIV-1.
Collectively, these findings demonstrate a critical role for RBBP4
in the regulation of HIV infection and suggest its potential as a
therapeutic target for HIV management.
HPV infection
Oral squamous cell carcinoma (OSCC) and
oropharyngeal squamous cell carcinoma (OPSCC) constitute a major
global public health burden, and there is an association between
infection with high-risk types of HPV and OSCC risk (173). Lohavanichbutr et al
(174) identified differential
expression of RBBP4 in HPV-positive vs. HPV-negative oropharyngeal
cancer. Wurlitzer et al (175) performed a mass spectrometric
comparison of eight HPV-positive and nine HPV-negative OPSCC cases,
and found that RBBP4/7 was expressed at higher levels in
HPV-positive OPSCC.
In cervical cancer, a major HPV-related cancer,
RBBP4 mediates the transforming activity of HPV16 (153) and is upregulated by ALA-PDT in
HPV16 immortalized cervical cells (154). These findings indicated that
RBBP4 plays a key role in HPV infection; however, the specific
mechanism still needs further exploration. Moreover, current
research on the role of RBBP7 in HPV infection is insufficient.
Plasmodium infection
Kaushik et al (176) discovered that the homologs of
RBBP4/7 in Plasmodium falciparum (PfRBBP4/7, PF3D7_0110700)
retain the β-helical conformation and binding interfaces, exhibit
significant interspecies differences, and show stage-specific
expression in the asexual blood stages of the parasite, increasing
from the ring stage to the schizont stage, and localizing in the
nucleus. Furthermore, PfRBBP4/7 have been shown to interact with
histone H4, suggesting their role in chromatin assembly and
remodeling pathways in P. falciparum. As CAF-1 family
members, they show structural and functional consistency. PfRBBP4,
central in malaria biology with 108 PPIs, emerges as a potential
antimalarial drug target (177). Thus, the function of PfRBBP4/7
in P. falciparum illustrates their potential as targets to
develop novel antimalarial interventions.
5. RBBP4/7 as potential targets for human
disease treatment
In the realm of targeted therapy research focused
on RBBP4/7, these proteins have demonstrated significant potential
in the treatment of various diseases, particularly in modulating
therapeutic outcomes. For example, increased expression of RBBP4
has been linked to mitigating lead-induced neuronal apoptosis,
suggesting a potential role in alleviating lead poisoning and
related neurological disorders (178). Additionally, the interaction of
RBBP4 with the efficacy of multiple drugs has been extensively
studied, including its role in enhancing the sensitivity of GBM
cells to TMZ (56,95), suppression of LC cell malignancy
via ropivacaine by downregulating RBBP4 (179), and the identification of the
circ-0110498/miR-1287-5p/RBBP4 axis as a novel target for
overcoming cisplatin resistance in NSCLC (85). RBBP4 is also considered a
potential target for treating CRC with PPD (119). In therapeutic contexts, RBBP4
expression is significantly increased in cervical cancer cell lines
treated with ALA-PDT (155),
and upregulation of RBBP4 has been found to induce radiosensitivity
in BC, melanoma and TNBC (180). Conversely, reduced levels of
RBBP7 may be associated with survival rates and chemoresistance
phenotypes in basal-like BC (152).
PPIs play a pivotal role in cellular functions, and
modulating PPIs offers a novel therapeutic avenue. It has been
reported that blocking the interaction between BCL11A and RBBP4
reduces the cancer stem cell population in TNBC (145). Furthermore, compounds, such as
bittersweet alkaloid II, may aid in AD treatment by stabilizing the
RBBP4-FOG1 complex (103).
Additionally, peptides designed by Hart et al targeting the
RBBP4/MTA1 interaction interface show potential as future
therapeutic strategies for disrupting epigenetic regulation
mechanisms in various types of cancer (181). Despite the potential of small
molecules or peptides targeting RBBP4/7, challenges such as low
oral bioavailability and poor in vivo stability remain,
necessitating further research to overcome these obstacles.
Emerging research has consistently linked elevated
RBBP4/7 expression to poorer prognosis across various cancer types
(Table V), underscoring their
pivotal role in disease therapy. Despite the evident potential of
RBBP4/7 in treating various diseases, the success of targeted
strategies remains elusive, possibly due to the complex biological
roles of RBBP4/7 and their involvement in multiple protein
complexes. Future research should investigate the mechanisms of
RBBP4/7 to develop targeted and effective treatment approaches.
This includes a deeper understanding of the specific roles of
RBBP4/7 in different cell types and disease states, identifying the
molecular networks interacting with RBBP4/7, studying their
expression and functional variations across diseases, and
validating therapeutic interventions targeting RBBP4/7 in
preclinical and clinical studies. Through these efforts, the
scientific groundwork may be laid for novel treatment methods based
on RBBP4/7, offering more personalized and effective therapeutic
options for patients.
| Table VPrognostic significance of RBBP4/7 in
cancer. |
Table V
Prognostic significance of RBBP4/7 in
cancer.
A, RBBP4
|
|---|
| First author,
year | Disease type | Omics type | Samples or sample
sources | Analysis
methods | Prognostic
relevance | (Refs.) |
|---|
| Gao M, 2023 | NSCLC |
Transcriptomics | LUAD: 54 adjacent
normal, 497 tumor samples; LUSC: 49 adjacent normal, 502 tumor
samples | KM-plotter | Poor prognosis:
Reduced OS | (84) |
| Wang N, 2021 | LUAD |
Transcriptomics | KM website | KM-plotter | Poor prognosis:
Reduced OS | (86) |
| Jia W, 2023 | LUAD |
Transcriptomics | Patients with LUAD
from the KM-plotter database | KM-plotter | Poor prognosis:
Reduced OS, FP and PPS | (179) |
| Li J, 2023 | GBM |
Transcriptomics | Samples from
patients withMGMT-negative GBM in TCGA database | KM-plotter | Poor prognosis:
Reduced OS and PFS | (63) |
| Li D, 2018 | NB | Transcriptomics,
proteomics | Tissues from 42
primary cases of NB, GSE14340 and GSE16476 datasets | Log-rank test | Poor prognosis:
Poor differentiation, reduced survival probability | (97) |
| Khateb A, 2021 | AML |
Transcriptomics | GEPIA and TCGA | Log-rank test | Poor prognosis:
Reduced OS | (106) |
| Li YD, 2019 | CRC | Proteomics | Tumor tissues of 80
patients with CRC | KM-plotter,
log-rank test | Poor prognosis:
Reduced OS | (117) |
| Guo Q, 2020 | BC | Proteomics | 240 BC tumor
tissues | KM-plotter,
log-rank test | Poor prognosis:
Reduced OS | (141) |
| Barreiro-Alonso A,
2021 | PCa |
Transcriptomics | 494 prostate
adenocarcinoma tissues | Log-rank test | Poor prognosis:
Reduced PFS | (160) |
B, RBBP7
|
|---|
| First author,
year | Disease type | Omics type | Samples or sample
sources | Analysis
methods | Prognostic
relevance | (Refs.) |
|---|
| Wang H, 2022 | LUAD |
Transcriptomics | Samples of patients
with early-stage LUAD from different cohorts (TCGA, GSE30219,
GSE31210, GSE37745, GSE50081) | KM-plotter,
log-rank test, meta-analysis | Poor prognosis:
Reduced RFS | (89) |
| Zhu H, 2022 | LUAD |
Transcriptomics | / | LASSO regression,
forest plots | Poor prognosis | (88) |
| Yu N, 2018 | EC | Transcriptomics,
proteomics | 126 ESCC tissues,
182 patients with EC from TCGA database | KM-plotter,
log-rank test | Poor prognosis:
Reduced OS and DFS | (14) |
| Wang R, 2022 | EC |
Transcriptomics | Patients with EC in
GEPIA database and TCGA database | KM-plotter | Poor prognosis:
Reduced OS and DFS | (133) |
| Barreiro-Alonso A,
2021 | PCa |
Transcriptomics | 494 prostate
adenocarcinoma tissues | Log-rank test | Poor prognosis:
Reduced PFS | (160) |
6. Conclusion and future perspectives
RBBP4/7 are conserved proteins ubiquitously present
in various organisms, which function in chromatin modification and
gene regulation across species. However, their specific structure,
expression patterns and molecular mechanisms may differ depending
on the organism. For example, in P. falciparum,
Drosophila, zebrafish and Saccharomyces cerevisiae,
the structure of RBBP4 might resemble that in humans; however,
there could be unique structural domains or adjustments (18,80,96,176). Moreover, in these organisms,
RBBP4/7 primarily function during developmental and reproductive
stages. By contrast, in humans, RBBP4/7 are expressed across
diverse cells and tissues, and are associated with cell cycle
regulation and gene transcription.
As histone chaperones, RBBP4/7 regulate various
cellular processes and are implicated in a variety of human
diseases, thus the future of RBBP4/7 research is promising.
However, the expression patterns of RBBP4/7 exhibit significant
variability in certain tumor types. This variability is attributed
to the diverse roles of RBBP4/7 within multiple functional
complexes, whose impact on tumorigenesis is intricately linked to
the specific actions of these complexes, which vary with the
cellular context and tumor type. In addition, research into gene
mutations and DNA methylation abnormalities of RBBP4/7 in diseases
remains limited, with mutations in RBBP7 identified only in cases
of early spermatogenic failure (163). This underscores an important
area for further investigation. Further studies of the complex
molecular functions of RBBP4/7 may improve the understanding of
cellular processes and disease pathways, leading to the development
of innovative therapies for a variety of human diseases and
cancers. Furthermore, exploring RBBP4/7 as potential biomarkers
could improve diagnostic accuracy, enabling early detection and
personalized medicine approaches.
Availability of data and materials
Not applicable.
Authors' contributions
YZ was primarily responsible for writing, reviewing
and revising this review. AY, XS, NT, ZZ and YC participated in the
literature review and provided feedback for this review. JW and WW
provided guidance throughout the preparation of this manuscript and
made significant revisions to the text. Data authentication is not
applicable. All authors read and approved the final version of the
manuscript
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Abbreviations:
|
RBBP
|
RB binding protein
|
|
NuRD
|
nucleosome remodeling and
deacetylase
|
|
PRC2
|
polycomb repressive complex 2
|
|
RB
|
retinoblastoma protein
|
|
CAF-1
|
chromatin assembly factor 1
|
|
HAT1
|
histone acetyltransferase 1
|
|
CENP-A
|
centromere protein A
|
|
HDAC
|
histone deacetylase
|
|
CDK
|
cyclin-dependent kinase
|
|
FOXM1
|
forkhead box M1
|
|
MTA1
|
metastasis associated 1
|
|
SUZ12
|
SUZ12 polycomb repressive complex 2
subunit
|
|
EED
|
embryonic ectoderm development
|
|
EZH
|
enhancer of zeste homolog
|
|
BRCA1
|
breast cancer type 1 susceptibility
protein
|
|
GBM
|
glioblastoma
|
|
TMZ
|
temozolomide
|
|
FOG-2
|
friend of GATA protein 2
|
|
LC
|
lung cancer
|
|
NSCLC
|
non-small cell LC
|
|
CBX3
|
chromobox homolog 3
|
|
LUAD
|
lung adenocarcinoma
|
|
MPM
|
malignant pleural mesothelioma
|
|
NB
|
neuroblastoma
|
|
OCN
|
osteocalcin
|
|
AD
|
Alzheimer's disease
|
|
AML
|
acute myeloid leukemia
|
|
MM
|
multiple myeloma
|
|
CML
|
chronic myeloid leukemia
|
|
CRC
|
colorectal cancer
|
|
PPD
|
protopanaxadiol
|
|
GC
|
gastric cancer
|
|
SSeCKS
|
Src-suppressed C-kinase substrate
|
|
HCC
|
hepatocellular carcinoma
|
|
SALL4
|
Sal-like protein 4
|
|
EGCG
|
epigallocatechin gallate
|
|
MDR
|
multidrug resistance
|
|
ESCC
|
esophageal squamous cell
carcinoma
|
|
EMT
|
epithelial-mesenchymal transition
|
|
EC
|
esophageal cancer
|
|
TC
|
thyroid cancer
|
|
NF-κB
|
nuclear factor κB
|
|
PMOP
|
postmenopausal osteoporosis
|
|
ESR1
|
estrogen receptor 1
|
|
BC
|
breast cancer
|
|
TNBC
|
triple-negative BC
|
|
BCL11A
|
BCL11 transcription factor A
|
|
HPV
|
human papillomavirus
|
|
ALA-PDT
|
5-aminole-vulinic acid photodynamic
therapy
|
|
SP1
|
specificity protein 1
|
|
BKVN
|
BK virus-associated nephropathy
|
|
RCC
|
renal cell carcinoma
|
|
OSCC
|
oral squamous cell carcinoma
|
|
OPSCC
|
oropharyngeal squamous cell
carcinoma
|
|
PPI
|
protein-protein interaction
|
Acknowledgments
Not applicable.
Funding
This work was supported by the National Natural Science
Foundation of China (grant nos. 82004007 and 81774026).
References
|
1
|
Hammond CM, Strømme CB, Huang H, Patel DJ
and Groth A: Histone chaperone networks shaping chromatin function.
Nat Rev Mol Cell Biol. 18:141–158. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Laskey RA, Honda BM, Mills AD and Finch
JT: Nucleosomes are assembled by an acidic protein which binds
histones and transfers them to DNA. Nature. 275:416–420. 1978.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
De Koning L, Corpet A, Haber JE and
Almouzni G: Histone chaperones: An escort network regulating
histone traffic. Nat Struct Mol Biol. 14:997–1007. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Burgess RJ and Zhang Z: Histone chaperones
in nucleosome assembly and human disease. Nat Struct Mol Biol.
20:14–22. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Yang C, Shao Y, Wang X and Wang J, Wang P,
Huang C, Wang W and Wang J: The effect of the histone chaperones
HSPA8 and DEK on tumor immunity in hepatocellular carcinoma. Int J
Mol Sci. 24:26532023. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Xu C and Min J: Structure and function of
WD40 domain proteins. Protein Cell. 2:202–214. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Millard CJ, Varma N, Saleh A, Morris K,
Watson PJ, Bottrill AR, Fairall L, Smith CJ and Schwabe JW: The
structure of the core NuRD repression complex provides insights
into its interaction with chromatin. Elife. 5:e139412016.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Ciferri C, Lander GC, Maiolica A, Herzog
F, Aebersold R and Nogales E: Molecular architecture of human
polycomb repressive complex 2. Elife. 1:e000052012. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Murzina NV, Pei XY, Zhang W, Sparkes M,
Vicente-Garcia J, Pratap JV, McLaughlin SH, Ben-Shahar TR,
Verreault A, Luisi BF and Laue ED: Structural basis for the
recognition of histone H4 by the histone-chaperone RbAp46.
Structure. 16:1077–1085. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Schmitges FW, Prusty AB, Faty M, Stützer
A, Lingaraju GM, Aiwazian J, Sack R, Hess D, Li L, Zhou S, et al:
Histone methylation by PRC2 is inhibited by active chromatin marks.
Mol Cell. 42:330–341. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Feinberg AP, Koldobskiy MA and Göndör A:
Epigenetic modulators, modifiers and mediators in cancer aetiology
and progression. Nat Rev Genet. 17:284–299. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Li YD, Lv Z and Zhu WF: RBBP4 promotes
colon cancer malignant progression via regulating Wnt/β-catenin
pathway. World J Gastroenterol. 26:5328–5342. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Cao X, Li F, Shao J, Lv J, Chang A, Dong W
and Zhu F: Circular RNA hsa_circ_0102231 sponges miR-145 to promote
non-small cell lung cancer cell proliferation by up-regulating the
expression of RBBP4. J Biochem. 169:65–73. 2021. View Article : Google Scholar
|
|
14
|
Yu N, Zhang P, Wang L, He X, Yang S and Lu
H: RBBP7 is a prognostic biomarker in patients with esophageal
squamous cell carcinoma. Oncol Lett. 16:7204–7211. 2018.PubMed/NCBI
|
|
15
|
Guo L, Xia Y, Li H, Wang Z, Xu H, Dai X,
Zhang Y, Zhang H, Fan W, Wei F, et al: FIT links c-Myc and P53
acetylation by recruiting RBBP7 during colorectal carcinogenesis.
Cancer Gene Ther. 30:1124–1133. 2023. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Qian YW, Wang YC, Hollingsworth RE Jr,
Jones D, Ling N and Lee EY: A retinoblastoma-binding protein
related to a negative regulator of Ras in yeast. Nature.
364:648–652. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Nicolas E, Morales V, Magnaghi-Jaulin L,
Harel-Bellan A, Richard-Foy H and Trouche D: RbAp48 belongs to the
histone deacetylase complex that associates with the retinoblastoma
protein. J Biol Chem. 275:9797–9804. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Qian YW and Lee EY: Dual
retinoblastoma-binding proteins with properties related to a
negative regulator of ras in yeast. J Biol Chem. 270:25507–25513.
1995. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Brasen C, Dorosz J, Wiuf A, Boesen T,
Mirza O and Gajhede M: Expression, purification and
characterization of the human MTA2-RBBP7 complex. Biochim Biophys
Acta Proteins Proteom. 1865:531–538. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Zhang Y, Iratni R, Erdjument-Bromage H,
Tempst P and Reinberg D: Histone deacetylases and SAP18, a novel
polypeptide, are components of a human Sin3 complex. Cell.
89:357–364. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Sauer PV, Gu Y, Liu WH, Mattiroli F, Panne
D, Luger K and Churchill ME: Mechanistic insights into histone
deposition and nucleosome assembly by the chromatin assembly
factor-1. Nucleic Acids Res. 46:9907–9917. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Müller GA, Asthana A and Rubin SM:
Structure and function of MuvB complexes. Oncogene. 41:2909–2919.
2022. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Verreault A, Kaufman PD, Kobayashi R and
Stillman B: Nucleosomal DNA regulates the core-histone-binding
subunit of the human Hat1 acetyltransferase. Curr Biol. 8:96–108.
1998. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Suganuma T, Pattenden SG and Workman JL:
Diverse functions of WD40 repeat proteins in histone recognition.
Genes Dev. 22:1265–1268. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Xiao L, Dang Y, Hu B, Luo L, Zhao P, Wang
S and Zhang K: Overlapping functions of RBBP4 and RBBP7 in
regulating cell proliferation and histone H3.3 deposition during
mouse preimplantation development. Epigenetics. 17:1205–1218. 2022.
View Article : Google Scholar :
|
|
26
|
Satrimafitrah P, Barman HK, Ahmad A,
Nishitoh H, Nakayama T, Fukagawa T and Takami Y: RbAp48 is
essential for viability of vertebrate cells and plays a role in
chromosome stability. Chromosome Res. 24:161–173. 2016. View Article : Google Scholar
|
|
27
|
Guan LS, Li GC, Chen CC, Liu LQ and Wang
ZY: Rb-associated protein 46 (RbAp46) suppresses the tumorigenicity
of adenovirus-transformed human embryonic kidney 293 cells. Int J
Cancer. 93:333–338. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Lee BC, Lin Z and Yuen KW:
RbAp46/48(LIN-53) is required for Holocentromere assembly in
Caenorhabditis elegans. Cell Rep. 14:1819–1828. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Mouysset J, Gilberto S, Meier MG, Lampert
F, Belwal M, Meraldi P and Peter M: CRL4(RBBP7) is required for
efficient CENP-A deposition at centromeres. J Cell Sci.
128:1732–1745. 2015.PubMed/NCBI
|
|
30
|
Huang S, Lee WH and Lee EY: A cellular
protein that competes with SV40 T antigen for binding to the
retinoblastoma gene product. Nature. 350:160–162. 1991. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Schultz-Rogers LE, Thayer ML, Kambakam S,
Wierson WA, Helmer JA, Wishman MD, Wall KA, Greig JL, Forsman JL,
Puchhalapalli K, et al: Rbbp4 loss disrupts neural progenitor cell
cycle regulation independent of Rb and leads to Tp53 acetylation
and apoptosis. Dev Dyn. 251:1267–1290. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Dick FA and Rubin SM: Molecular mechanisms
underlying RB protein function. Nat Rev Mol Cell Biol. 14:297–306.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Brehm A, Miska EA, McCance DJ, Reid JL,
Bannister AJ and Kouzarides T: Retinoblastoma protein recruits
histone deacetylase to repress transcription. Nature. 391:597–601.
1998. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Narasimha AM, Kaulich M, Shapiro GS, Choi
YJ, Sicinski P and Dowdy SF: Cyclin D activates the Rb tumor
suppressor by mono-phosphorylation. Elife. 3:2014. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Gubern A, Joaquin M, Marquès M, Maseres P,
Garcia-Garcia J, Amat R, González-Nuñez D, Oliva B, Real FX, de
Nadal E and Posas F: The N-Terminal phosphorylation of RB by p38
bypasses its inactivation by CDKs and prevents proliferation in
cancer cells. Mol Cell. 64:25–36. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Guiley KZ, Liban TJ, Felthousen JG,
Ramanan P, Litovchick L and Rubin SM: Structural mechanisms of
DREAM complex assembly and regulation. Genes Dev. 29:961–974. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Müller GA, Quaas M, Schümann M, Krause E,
Padi M, Fischer M, Litovchick L, DeCaprio JA and Engeland K: The
CHR promoter element controls cell cycle-dependent gene
transcription and binds the DREAM and MMB complexes. Nucleic Acids
Res. 40:1561–1578. 2012. View Article : Google Scholar :
|
|
38
|
Mages CF, Wintsche A, Bernhart SH and
Müller GA: The DREAM complex through its subunit Lin37 cooperates
with Rb to initiate quiescence. Elife. 6:e268762017. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Uxa S, Bernhart SH, Mages CFS, Fischer M,
Kohler R, Hoffmann S, Stadler PF, Engeland K and Müller GA: DREAM
and RB cooperate to induce gene repression and cell-cycle arrest in
response to p53 activation. Nucleic Acids Res. 47:9087–9103. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Schade AE, Fischer M and DeCaprio JA: RB,
p130 and p107 differentially repress G1/S and G2/M genes after p53
activation. Nucleic Acids Res. 47:11197–11208. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Zhang Y, Ng HH, Erdjument-Bromage H,
Tempst P, Bird A and Reinberg D: Analysis of the NuRD subunits
reveals a histone deacetylase core complex and a connection with
DNA methylation. Genes Dev. 13:1924–1935. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Bao Y and Shen X: Chromatin remodeling in
DNA double-strand break repair. Curr Opin Genet Dev. 17:126–131.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Zhang W, Tyl M, Ward R, Sobott F, Maman J,
Murthy AS, Watson AA, Fedorov O, Bowman A, Owen-Hughes T, et al:
Structural plasticity of histones H3-H4 facilitates their
allosteric exchange between RbAp48 and ASF1. Nat Struct Mol Biol.
20:29–35. 2013. View Article : Google Scholar :
|
|
44
|
Torchy MP, Hamiche A and Klaholz BP:
Structure and function insights into the NuRD chromatin remodeling
complex. Cell Mol Life Sci. 72:2491–2507. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Mu W, Murcia NS, Smith KN, Menon DU, Yee D
and Magnuson T: RBBP4 dysfunction reshapes the genomic landscape of
H3K27 methylation and acetylation and disrupts gene expression. G3
(Bethesda). 12:jkac0822022. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Mei S, Liu Y, Bao Y, Zhang Y, Min S, Liu
Y, Huang Y, Yuan X, Feng Y, Shi J and Yang R: Dendritic
cell-associated miRNAs are modulated via chromatin remodeling in
response to different environments. PLoS One. 9:e902312014.
View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Shang WH, Hori T, Westhorpe FG, Godek KM,
Toyoda A, Misu S, Monma N, Ikeo K, Carroll CW, Takami Y, et al:
Acetylation of histone H4 lysine 5 and 12 is required for CENP-A
deposition into centromeres. Nat Commun. 7:134652016. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Vermaak D, Wade PA, Jones PL, Shi YB and
Wolffe AP: Functional analysis of the SIN3-histone deacetylase
RPD3-RbAp48-histone H4 connection in the Xenopus oocyte. Mol Cell
Biol. 19:5847–5860. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Balboula AZ, Stein P, Schultz RM and
Schindler K: RBBP4 regulates histone deacetylation and bipolar
spindle assembly during oocyte maturation in the mouse. Biol
Reprod. 92:1052015. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Yue Y, Yang WS, Zhang L, Liu CP and Xu RM:
Topography of histone H3-H4 interaction with the Hat1-Hat2
acetyltransferase complex. Genes Dev. 36:408–413. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Pardal AJ and Bowman AJ: A specific role
for importin-5 and NASP in the import and nuclear hand-off of
monomeric H3. Elife. 11:e817552022. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Conway E, Healy E and Bracken AP: PRC2
mediated H3K27 methylations in cellular identity and cancer. Curr
Opin Cell Biol. 37:42–48. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Kuzmichev A, Jenuwein T, Tempst P and
Reinberg D: Different EZH2-containing complexes target methylation
of histone H1 or nucleosomal histone H3. Mol Cell. 14:183–193.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Oksuz O, Narendra V, Lee CH, Descostes N,
LeRoy G, Raviram R, Blumenberg L, Karch K, Rocha PP, Garcia BA, et
al: Capturing the onset of PRC2-Mediated repressive domain
formation. Mol Cell. 70:1149–1162.e5. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Chen S, Jiao L and Liu X, Yang X and Liu
X: A Dimeric structural scaffold for PRC2-PCL targeting to CpG
island chromatin. Mol Cell. 77:1265–1278.e7. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Kitange GJ, Mladek AC, Schroeder MA,
Pokorny JC, Carlson BL, Zhang Y, Nair AA, Lee JH, Yan H, Decker PA,
et al: Retinoblastoma binding protein 4 modulates temozolomide
sensitivity in glioblastoma by regulating DNA repair proteins. Cell
Rep. 14:2587–2598. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Vaute O, Nicolas E, Vandel L and Trouche
D: Functional and physical interaction between the histone methyl
transferase Suv39H1 and histone deacetylases. Nucleic Acids Res.
30:475–481. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Marin TL, Gongol B, Zhang F, Martin M,
Johnson DA, Xiao H, Wang Y, Subramaniam S, Chien S and Shyy JY:
AMPK promotes mitochondrial biogenesis and function by
phosphorylating the epigenetic factors DNMT1, RBBP7, and HAT1. Sci
Signal. 10:eaaf74782017. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Li DQ and Kumar R: Mi-2/NuRD complex
making inroads into DNA-damage response pathway. Cell Cycle.
9:2071–2079. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Yang B, Zhang W, Sun L, Lu B, Yin C, Zhang
Y and Jiang H: Creatine kinase brain-type regulates BCAR1
phosphorylation to facilitate DNA damage repair. iScience.
26:1066842023. View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Yarden RI and Brody LC: BRCA1 interacts
with components of the histone deacetylase complex. Proc Natl Acad
Sci USA. 96:4983–4988. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Chen GC, Guan LS, Yu JH, Li GC, Choi Kim
HR and Wang ZY: Rb-associated protein 46 (RbAp46) inhibits
transcriptional transactivation mediated by BRCA1. Biochem Biophys
Res Commun. 284:507–514. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Li J, Song C, Gu J, Li C, Zang W, Shi L,
Chen L, Zhu L, Zhou M, Wang T, et al: RBBP4 regulates the
expression of the Mre11-Rad50-NBS1 (MRN) complex and promotes DNA
double-strand break repair to mediate glioblastoma
chemoradiotherapy resistance. Cancer Lett. 557:2160782023.
View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Li GC, Guan LS and Wang ZY: Overexpression
of RbAp46 facilitates stress-induced apoptosis and suppresses
tumorigenicity of neoplastigenic breast epithelial cells. Int J
Cancer. 105:762–768. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Yang J, Kiefer S and Rauchman M:
Characterization of the gene encoding mouse retinoblastoma binding
protein-7, a component of chromatin-remodeling complexes. Genomics.
80:407–415. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
66
|
Horihata K, Inoue N, Uenoyama Y, Maeda KI
and Tsukamura H: Retinoblastoma binding protein 7 is involved in
Kiss1 mRNA upregulation in rodents. J Reprod Dev. 66:125–133. 2020.
View Article : Google Scholar : PubMed/NCBI
|
|
67
|
Giri R, Yeh HH, Wu CH and Liu HS: SUMO-1
overexpression increases RbAp46 protein stability and suppresses
cell growth. Anticancer Res. 28:3749–3756. 2008.
|
|
68
|
Ping W, Sheng Y, Hu G, Zhong H, Li Y, Liu
Y, Luo W, Yan C, Wen Y, Wang X, et al: RBBP4 is an epigenetic
barrier for the induced transition of pluripotent stem cells into
totipotent 2C-like cells. Nucleic Acids Res. 51:5414–5431. 2023.
View Article : Google Scholar : PubMed/NCBI
|
|
69
|
Garnatz AS, Gao Z, Broman M, Martens S,
Earley JU and Svensson EC: FOG-2 mediated recruitment of the NuRD
complex regulates cardiomyocyte proliferation during heart
development. Dev Biol. 395:50–61. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
70
|
Bonuccelli L, Rossi L, Lena A, Scarcelli
V, Rainaldi G, Evangelista M, Iacopetti P, Gremigni V and Salvetti
A: An RbAp48-like gene regulates adult stem cells in planarians. J
Cell Sci. 123:690–698. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
71
|
He H, Kong S, Liu F, Zhang S, Jiang Y,
Liao Y, Jiang Y, Li Q, Wang B, Zhou Z, et al: Rbbp7 is required for
uterine stromal decidualization in mice. Biol Reprod. 93:132015.
View Article : Google Scholar : PubMed/NCBI
|
|
72
|
Shore AN, Kabotyanski EB, Roarty K, Smith
MA, Zhang Y, Creighton CJ, Dinger ME and Rosen JM:
Pregnancy-induced noncoding RNA (PINC) associates with polycomb
repressive complex 2 and regulates mammary epithelial
differentiation. PLoS Genet. 8:e10028402012. View Article : Google Scholar : PubMed/NCBI
|
|
73
|
Xin J, Li J, Feng Y, Wang L, Zhang Y and
Yang R: Downregulation of long noncoding RNA HOTAIRM1 promotes
monocyte/dendritic cell differentiation through competitively
binding to endogenous miR-3960. Onco Targets Ther. 10:1307–1315.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
74
|
Rico M, Mukherjee A, Konieczkowski M,
Bruggeman LA, Miller RT, Khan S, Schelling JR and Sedor JR:
WT1-interacting protein and ZO-1 translocate into podocyte nuclei
after puromycin aminonucleoside treatment. Am J Physiol Renal
Physiol. 289:F431–F441. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
75
|
Scuto A, Zhang H, Zhao H, Rivera M,
Yeatman TJ, Jove R and Torres-Roca JF: RbAp48 regulates
cytoskeletal organization and morphology by increasing K-Ras
activity and signaling through mitogen-activated protein kinase.
Cancer Res. 67:10317–10324. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
76
|
Gasca S, Pellestor F, Assou S, Loup V,
Anahory T, Dechaud H, De Vos J and Hamamah S: Identifying new human
oocyte marker genes: A microarray approach. Reprod Biomed Online.
14:175–183. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
77
|
Balboula AZ, Stein P, Schultz RM and
Schindler K: Knockdown of RBBP7 unveils a requirement of histone
deacetylation for CPC function in mouse oocytes. Cell Cycle.
13:600–611. 2014. View Article : Google Scholar
|
|
78
|
Guan LS, Rauchman M and Wang ZY: Induction
of Rb-associated protein (RbAp46) by Wilms' tumor suppressor WT1
mediates growth inhibition. J Biol Chem. 273:27047–27050. 1998.
View Article : Google Scholar : PubMed/NCBI
|
|
79
|
Pegoraro G, Kubben N, Wickert U, Göhler H,
Hoffmann K and Misteli T: Ageing-related chromatin defects through
loss of the NURD complex. Nat Cell Biol. 11:1261–1267. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
80
|
Hunt LC, Stover J, Haugen B, Shaw TI, Li
Y, Pagala VR, Finkelstein D, Barton ER, Fan Y, Labelle M, et al: A
key role for the ubiquitin ligase UBR4 in myofiber hypertrophy in
drosophila and mice. Cell Rep. 28:1268–1281.e1266. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
81
|
Pavlopoulos E, Jones S, Kosmidis S, Close
M, Kim C, Kovalerchik O, Small SA and Kandel ER: Molecular
mechanism for age-related memory loss: The histone-binding protein
RbAp48. Sci Transl Med. 5:200ra1152013. View Article : Google Scholar : PubMed/NCBI
|
|
82
|
Tsujii A, Miyamoto Y, Moriyama T, Tsuchiya
Y, Obuse C, Mizuguchi K, Oka M and Yoneda Y: Retinoblastoma-binding
protein 4-regulated classical nuclear transport is involved in
cellular senescence. J Biol Chem. 290:29375–29388. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
83
|
Ginger MR, Gonzalez-Rimbau MF, Gay JP and
Rosen JM: Persistent changes in gene expression induced by estrogen
and progesterone in the rat mammary gland. Mol Endocrinol.
15:1993–2009. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
84
|
Gao M, Li Y, Huang H, Fan Y, Shi R, Su L,
Chen C, Li X, Zhu G and Wu D: Exploring the association between
PRC2 genes variants and lung cancer risk in chinese han population.
Onco Targets Ther. 16:499–513. 2023. View Article : Google Scholar : PubMed/NCBI
|
|
85
|
Hao D, Li Y, Shi J and Jiang J:
Circ_0110498 facilitates the cisplatin resistance of non-small cell
lung cancer by mediating the miR-1287-5p/RBBP4 axis. Thorac Cancer.
14:662–672. 2023. View Article : Google Scholar : PubMed/NCBI
|
|
86
|
Wang N, Wang W, Mao W, Kuerbantayi N, Jia
N, Chen Y, Zhou F, Yin L and Wang Y: RBBP4 enhances platinum chemo
resistance in lung adenocarcinoma. Biomed Res Int.
2021:69059852021.PubMed/NCBI
|
|
87
|
Jin X, Zhang B, Zhang H and Yu H:
Smoking-associated upregulation of CBX3 suppresses ARHGAP24
expression to activate Rac1 signaling and promote tumor progression
in lung adenocarcinoma. Oncogene. 41:538–549. 2022. View Article : Google Scholar :
|
|
88
|
Zhu H, Zheng C, Liu H, Kong F, Kong S,
Chen F and Tian Y: Significance of macrophage infiltration in the
prognosis of lung adenocarcinoma patients evaluated by scRNA and
bulkRNA analysis. Front Immunol. 13:10284402022. View Article : Google Scholar : PubMed/NCBI
|
|
89
|
Wang H, Lu X and Chen J: Construction and
experimental validation of an acetylation-related gene signature to
evaluate the recurrence and immunotherapeutic response in
early-stage lung adenocarcinoma. BMC Med Genomics. 15:2542022.
View Article : Google Scholar : PubMed/NCBI
|
|
90
|
Wang CL, Wang CI, Liao PC, Chen CD, Liang
Y, Chuang WY, Tsai YH, Chen HC, Chang YS, Yu JS, et al: Discovery
of retinoblastoma-associated binding protein 46 as a novel
prognostic marker for distant metastasis in nonsmall cell lung
cancer by combined analysis of cancer cell secretome and pleural
effusion proteome. J Proteome Res. 8:4428–4440. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
91
|
Tsao AS, Pass HI, Rimner A and Mansfield
AS: New era for malignant pleural mesothelioma: Updates on
therapeutic options. J Clin Oncol. 40:681–692. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
92
|
Vavougios GD, Solenov EI, Hatzoglou C,
Baturina GS, Katkova LE, Molyvdas PA, Gourgoulianis KI and
Zarogiannis SG: Computational genomic analysis of PARK7 interactome
reveals high BBS1 gene expression as a prognostic factor favoring
survival in malignant pleural mesothelioma. Am J Physiol Lung Cell
Mol Physiol. 309:L677–L686. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
93
|
Barzegar Behrooz A, Latifi-Navid H, da
Silva Rosa SC, Swiat M, Wiechec E, Vitorino C, Vitorino R,
Jamalpoor Z and Ghavami S: Integrating Multi-Omics analysis for
enhanced diagnosis and treatment of glioblastoma: A comprehensive
Data-Driven approach. Cancers (Basel). 15:31582023. View Article : Google Scholar : PubMed/NCBI
|
|
94
|
Shou J, Gao H, Cheng S, Wang B and Guan H:
LncRNA HOXA-AS2 promotes glioblastoma carcinogenesis by targeting
miR-885-5p/RBBP4 axis. Cancer Cell Int. 21:392021. View Article : Google Scholar : PubMed/NCBI
|
|
95
|
Mladek AC, Yan H, Tian S, Decker PA,
Burgenske DM, Bakken K, Hu Z, He L, Connors MA, Carlson BL, et al:
RBBP4-p300 axis modulates expression of genes essential for cell
survival and is a potential target for therapy in glioblastoma.
Neuro Oncol. 24:1261–1272. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
96
|
Schultz LE, Haltom JA, Almeida MP, Wierson
WA, Solin SL, Weiss TJ, Helmer JA, Sandquist EJ, Shive HR and
McGrail M: Epigenetic regulators Rbbp4 and Hdac1 are overexpressed
in a zebrafish model of RB1 embryonal brain tumor, and are required
for neural progenitor survival and proliferation. Dis Model Mech.
11:dmm0341242018. View Article : Google Scholar : PubMed/NCBI
|
|
97
|
Li D, Song H, Mei H, Fang E, Wang X, Yang
F, Li H, Chen Y, Huang K, Zheng L and Tong Q: Armadillo repeat
containing 12 promotes neuroblastoma progression through
interaction with retinoblastoma binding protein 4. Nat Commun.
9:28292018. View Article : Google Scholar : PubMed/NCBI
|
|
98
|
Crea F, Hurt EM and Farrar WL: Clinical
significance of Polycomb gene expression in brain tumors. Mol
Cancer. 9:2652010. View Article : Google Scholar : PubMed/NCBI
|
|
99
|
Khrimian L, Obri A, Ramos-Brossier M,
Rousseaud A, Moriceau S, Nicot AS, Mera P, Kosmidis S, Karnavas T,
Saudou F, et al: Gpr158 mediates osteocalcin's regulation of
cognition. J Exp Med. 214:2859–2873. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
100
|
Kosmidis S, Polyzos A, Harvey L, Youssef
M, Denny CA, Dranovsky A and Kandel ER: RbAp48 protein is a
critical component of GPR158/OCN signaling and ameliorates
Age-Related memory loss. Cell Rep. 25:959–973.e6. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
101
|
Barral S, Reitz C, Small SA and Mayeux R:
Genetic variants in a 'cAMP element binding protein'
(CREB)-dependent histone acetylation pathway influence memory
performance in cognitively healthy elderly individuals. Neurobiol
Aging. 35:2881.e7–2881.e10. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
102
|
Lejon S, Thong SY, Murthy A, AlQarni S,
Murzina NV, Blobel GA, Laue ED and Mackay JP: Insights into
association of the NuRD complex with FOG-1 from the crystal
structure of an RbAp48•FOG-1 complex. J Biol Chem. 286:1196–1203.
2011. View Article : Google Scholar
|
|
103
|
Huang HJ, Lee CC and Chen CY: Lead
discovery for Alzheimer's disease related target protein RbAp48
from traditional Chinese medicine. Biomed Res Int. 2014:7649462014.
View Article : Google Scholar : PubMed/NCBI
|
|
104
|
Ramis MR, Sarubbo F, Tejada S, Jiménez M,
Esteban S, Miralles A and Moranta D: chronic Polyphenon-60 or
catechin treatments increase brain monoamines syntheses and
hippocampal SIRT1 levels improving cognition in aged rats.
Nutrients. 12:3262020. View Article : Google Scholar : PubMed/NCBI
|
|
105
|
Dave N, Vural AS, Piras IS, Winslow W,
Surendra L, Winstone JK, Beach TG, Huentelman MJ and Velazquez R:
Identification of retinoblastoma binding protein 7 (Rbbp7) as a
mediator against tau acetylation and subsequent neuronal loss in
Alzheimer's disease and related tauopathies. Acta Neuropathol.
142:279–294. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
106
|
Khateb A, Deshpande A, Feng Y, Finlay D,
Lee JS, Lazar I, Fabre B, Li Y, Fujita Y and Zhang T: The ubiquitin
ligase RNF5 determines acute myeloid leukemia growth and
susceptibility to histone deacetylase inhibitors. Nat Commun.
12:53972021. View Article : Google Scholar : PubMed/NCBI
|
|
107
|
Casas S, Ollila J, Aventín A, Vihinen M,
Sierra J and Knuutila S: Changes in apoptosis-related pathways in
acute myelocytic leukemia. Cancer Genet Cytogenet. 146:89–101.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
108
|
Sakhinia E, Faranghpour M, Liu Yin JA,
Brady G, Hoyland JA and Byers RJ: Routine expression profiling of
microarray gene signatures in acute leukaemia by real-time PCR of
human bone marrow. Br J Haematol. 130:233–248. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
109
|
Gao H, Wang H and Yang W: Identification
of key genes and construction of microRNA-mRNA regulatory networks
in multiple myeloma by integrated multiple GEO datasets using
bioinformatics analysis. Int J Hematol. 106:99–107. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
110
|
Merkerova M, Bruchova H, Kracmarova A,
Klamova H and Brdicka R: Bmi-1 over-expression plays a secondary
role in chronic myeloid leukemia transformation. Leuk Lymphoma.
48:793–801. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
111
|
Hu SY, Chen ZX, Cen JN, Gu M, Zhao Y, Shen
HL and Wang W: RbAp46 gene activates the expression of IGFBP-rP1
gene in K562 leukemic cells. Zhonghua Xue Ye Xue Za Zhi.
27:107–110. 2006.In Chinese. PubMed/NCBI
|
|
112
|
Hu SY, Chen ZX, Gu WY, Cen JN, Zhao Y and
Gu M: Detection of RbAp46 expression in bone marrow cells of
leukemia patients by real-time quantitative RT-PCR. Zhonghua Xue Ye
Xue Za Zhi. 26:417–420. 2005.In Chinese. PubMed/NCBI
|
|
113
|
Hu SY, Chen ZX, Gu WY, Cen JN and Zhao Y:
High expression of RbAp46 gene in patients with acute leukemia or
chronic myelogenous leukemia in blast crisis. Chin Med J (Engl).
118:1295–1298. 2005.PubMed/NCBI
|
|
114
|
Duan WM, Chen ZX, Wang W, Fu JX, Bai X,
Zhao XJ and Yao L: Establishment and characterization of leukemic
cell line U937 stably expressing exogenous RbAp46. Zhongguo Shi Yan
Xue Ye Xue Za Zhi. 12:416–419. 2004.In Chinese. PubMed/NCBI
|
|
115
|
Ritter J and Bielack SS: Osteosarcoma. Ann
Oncol. 21(Suppl 7): vii320–vii325. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
116
|
Zhang TF, Yu SQ, Loggie BW and Wang ZY:
Inducible expression of RbAp46 activates c-Jun NH2-terminal
kinase-dependent apoptosis and suppresses progressive growth of
tumor xenografts in nude mice. Anticancer Res. 23:4621–4627.
2003.
|
|
117
|
Li YD, Lv Z, Xie HY and Zheng SS:
Retinoblastoma binding protein 4 up-regulation is correlated with
hepatic metastasis and poor prognosis in colon cancer patients.
Hepatobiliary Pancreat Dis Int. 18:446–451. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
118
|
Zhu X, Luo X, Long X, Jiang S, Xie X,
Zhang Q and Wang H: CircAGO2 promotes colorectal cancer progression
by inhibiting heat shock protein family B (small) member 8 via
miR-1-3p/retinoblastoma binding protein 4 axis. Funct Integr
Genomics. 23:782023. View Article : Google Scholar : PubMed/NCBI
|
|
119
|
Zhuo FF, Guo Q, Zheng YZ, Liu TT, Yang Z,
Xu QH, Jiang Y, Liu D, Zeng KW and Tu PF: Photoaffinity
Labeling-Based chemoproteomic strategy reveals RBBP4 as a cellular
target of protopanaxadiol against colorectal cancer cells.
Chembiochem. 23:e2022000382022. View Article : Google Scholar : PubMed/NCBI
|
|
120
|
Ding L, Zhao Y, Dang S, Wang Y, Li X, Yu
X, Li Z, Wei J, Liu M and Li G: Circular RNA circ-DONSON
facilitates gastric cancer growth and invasion via NURF complex
dependent activation of transcription factor SOX4. Mol Cancer.
18:452019. View Article : Google Scholar : PubMed/NCBI
|
|
121
|
Jin X, Jiang R, Xiang Y, Fan Z, Wu Z, Yang
B, Yang L, Wei S and Yang Y: Overexpression of
retinoblastoma-binding protein 4 contributes to the
radiosensitivity of AGS gastric cancer cells via
phosphoinositide3-kinase/protein kinase B pathway suppression. Mol
Med Rep. 18:1571–1581. 2018.PubMed/NCBI
|
|
122
|
Choi MC, Jong HS, Kim TY, Song SH, Lee DS,
Lee JW, Kim TY, Kim NK and Bang YJ: AKAP12/Gravin is inactivated by
epigenetic mechanism in human gastric carcinoma and shows growth
suppressor activity. Oncogene. 23:7095–7103. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
123
|
Liu Y, Gao L and Gelman IH:
SSeCKS/Gravin/AKAP12 attenuates expression of proliferative and
angiogenic genes during suppression of v-Src-induced oncogenesis.
BMC Cancer. 6:1052006. View Article : Google Scholar : PubMed/NCBI
|
|
124
|
Song H, Xia SL, Liao C, Li YL, Wang YF, Li
TP and Zhao MJ: Genes encoding Pir51, Beclin 1, RbAp48 and aldolase
b are up or down-regulated in human primary hepatocellular
carcinoma. World J Gastroenterol. 10:509–513. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
125
|
Liu BH, Jobichen C, Chia CSB, Chan THM,
Tang JP, Chung TXY, Li J, Poulsen A, Hung AW, Koh-Stenta X, et al:
Targeting cancer addiction for SALL4 by shifting its transcriptome
with a pharmacologic peptide. Proc Natl Acad Sci USA.
115:E7119–E7128. 2018.PubMed/NCBI
|
|
126
|
Li L, Tang J, Zhang B, Yang W, LiuGao M,
Wang R, Tan Y, Fan J, Chang Y, Fu J, et al: Epigenetic modification
of MiR-429 promotes liver tumour-initiating cell properties by
targeting Rb binding protein 4. Gut. 64:156–167. 2015. View Article : Google Scholar
|
|
127
|
Tang HH, Zhou M and Liang G: Impact of
epigallocatechin gallate on gene expression profiles of human
hepatocellular carcinoma cell lines BEL7404/ADM and BEL7402/5-FU.
Ai Zheng. 27:1056–1064. 2008.In Chinese. PubMed/NCBI
|
|
128
|
Younossi ZM: Non-alcoholic fatty liver
disease-A global public health perspective. J Hepatol. 70:531–544.
2019. View Article : Google Scholar
|
|
129
|
Yao Y, Li X, Wang W, Liu Z, Chen J, Ding M
and Huang X: MRT, Functioning with NURF complex, regulates lipid
droplet size. Cell Rep. 24:2972–2984. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
130
|
Zhi S, Congcong Z, Zhiling G, Yihan Q,
Yijing X, Guanjie L, Fang W, Xuehua S, Hongjie L, Xiaoni K and
Yueqiu G: Quantitative proteomics of HFD-induced fatty liver
uncovers novel transcription factors of lipid metabolism. Int J
Biol Sci. 18:3298–3312. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
131
|
Chen L, Lu J, Xu T, Yan Z, Guo Y, Dong Z
and Guo W: KTN1-AS1, a SOX2-mediated lncRNA, activates
epithelial-mesenchymal transition process in esophageal squamous
cell carcinoma. Sci Rep. 12:201862022. View Article : Google Scholar : PubMed/NCBI
|
|
132
|
Bai X, Wang D, Ji H, Zheng L, Lu Y, Tang
W, Zhang H, Xu W, Li J, Fei Z and Wang H: RbAp48 is critical for
the proliferation of hypopharyngeal carcinoma. ORL J
Otorhinolaryngol Relat Spec. 77:310–319. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
133
|
Wang R, Huang Z, Lin Z, Chen B, Pang X, Du
C and Fan H: Hypoxia-induced RBBP7 promotes esophagus cancer
progression by inducing CDK4 expression. Acta Biochim Biophys Sin
(Shanghai). 54:179–186. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
134
|
Xie ZF, Li HT, Xie SH and Ma M: Circular
RNA hsa_ circ_0006168 contributes to cell proliferation, migration
and invasion in esophageal cancer by regulating miR-384/RBBP7 axis
via activation of S6K/S6 pathway. Eur Rev Med Pharmacol Sci.
24:151–163. 2020.PubMed/NCBI
|
|
135
|
Pacifico F, Paolillo M, Chiappetta G,
Crescenzi E, Arena S, Scaloni A, Monaco M, Vascotto C, Tell G,
Formisano S and Leonardi A: RbAp48 is a target of nuclear
factor-kappaB activity in thyroid cancer. J Clin Endocrinol Metab.
92:1458–1466. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
136
|
Saul D, Ninkovic M, Komrakova M, Wolff L,
Simka P, Gasimov T, Menger B, Hoffmann DB, Rohde V and Sehmisch S:
Effect of zileuton on osteoporotic bone and its healing, expression
of bone, and brain genes in rats. J Appl Physiol (1985).
124:118–130. 2018. View Article : Google Scholar
|
|
137
|
Gong R, Ren S, Chen M, Wang Y, Zhang G,
Shi L, Zhang C, Su R and Li Y: Bioinformatics analysis reveals the
altered gene expression of patients with postmenopausal
osteoporosis using liuweidihuang pills treatment. Biomed Res Int.
2019:19079062019.PubMed/NCBI
|
|
138
|
Yang C, Ren J, Li B, Jin C, Ma C, Cheng C,
Sun Y and Shi X: Identification of gene biomarkers in patients with
postmenopausal osteoporosis. Mol Med Rep. 19:1065–1073. 2019.
|
|
139
|
Ishimaru N, Arakaki R, Omotehara F, Yamada
K, Mishima K, Saito I and Hayashi Y: Novel role for RbAp48 in
tissue-specific, estrogen deficiency-dependent apoptosis in the
exocrine glands. Mol Cell Biol. 26:2924–2935. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
140
|
Ishimaru N, Arakaki R, Yoshida S, Yamada
A, Noji S and Hayashi Y: Expression of the retinoblastoma protein
RbAp48 in exocrine glands leads to Sjögren's syndrome-like
autoimmune exocrinopathy. J Exp Med. 205:2915–2927. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
141
|
Guo Q, Cheng K, Wang X, Li X, Yu Y, Hua Y
and Yang Z: Expression of HDAC1 and RBBP4 correlate with
clinicopathologic characteristics and prognosis in breast cancer.
Int J Clin Exp Pathol. 13:563–572. 2020.PubMed/NCBI
|
|
142
|
Gong X, Dong T, Niu M, Liang X, Sun S,
Zhang Y, Li Y and Li D: lncRNA LCPAT1 upregulation promotes breast
cancer progression via enhancing MFAP2 transcription. Mol Ther
Nucleic Acids. 21:804–813. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
143
|
Creekmore AL, Walt KA, Schultz-Norton JR,
Ziegler YS, McLeod IX, Yates JR and Nardulli AM: The role of
retinoblastoma-associated proteins 46 and 48 in estrogen receptor
alpha mediated gene expression. Mol Cell Endocrinol. 291:79–86.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
144
|
Zheng Z, Yao X and Liu Y: RBBP4 plays a
vital role in the malignant progression of triple-negative breast
cancer by regulating epithelial-mesenchymal transition. Genes
Genomics. 44:1301–1309. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
145
|
Moody RR, Lo MC, Meagher JL, Lin CC,
Stevers NO, Tinsley SL, Jung I, Matvekas A, Stuckey JA and Sun D:
Probing the interaction between the histone
methyltransferase/deacetylase subunit RBBP4/7 and the transcription
factor BCL11A in epigenetic complexes. J Biol Chem. 293:2125–2136.
2018. View Article : Google Scholar :
|
|
146
|
Zhang TF, Yu SQ, Deuel TF and Wang ZY:
Constitutive expression of Rb associated protein 46 (RbAp46)
reverts transformed phenotypes of breast cancer cells. Anticancer
Res. 23:3735–3740. 2003.PubMed/NCBI
|
|
147
|
Zhang TF, Yu SQ and Wang ZY: RbAp46
inhibits estrogen-stimulated progression of neoplastigenic breast
epithelial cells. Anticancer Res. 27:3205–3209. 2007.PubMed/NCBI
|
|
148
|
Fu J, Qin L, He T, Qin J, Hong J, Wong J,
Liao L and Xu J: The TWIST/Mi2/NuRD protein complex and its
essential role in cancer metastasis. Cell Res. 21:275–289. 2011.
View Article : Google Scholar
|
|
149
|
Li GC and Wang ZY: Constitutive expression
of RbAp46 induces epithelial-mesenchymal transition in mammary
epithelial cells. Anticancer Res. 26:3555–3560. 2006.PubMed/NCBI
|
|
150
|
Thakur A, Rahman KW, Wu J, Bollig A,
Biliran H, Lin X, Nassar H, Grignon DJ, Sarkar FH and Liao JD:
Aberrant expression of X-linked genes RbAp46, Rsk4, and Cldn2 in
breast cancer. Mol Cancer Res. 5:171–181. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
151
|
Ebata A, Suzuki T, Takagi K, Miki Y,
Onodera Y, Nakamura Y, Fujishima F, Ishida K, Watanabe M, Tamaki K,
et al: Oestrogen-induced genes in ductal carcinoma in situ: Their
comparison with invasive ductal carcinoma. Endocr Relat Cancer.
19:485–496. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
152
|
Mieczkowska IK, Pantelaiou-Prokaki G,
Prokakis E, Schmidt GE, Müller-Kirschbaum LC, Werner M, Sen M,
Velychko T, Jannasch K, Dullin C, et al: Decreased PRC2 activity
supports the survival of basal-like breast cancer cells to
cytotoxic treatments. Cell Death Dis. 12:11182021. View Article : Google Scholar : PubMed/NCBI
|
|
153
|
Kong L, Yu XP, Bai XH, Zhang WF, Zhang Y,
Zhao WM, Jia JH, Tang W, Zhou YB and Liu CJ: RbAp48 is a critical
mediator controlling the transforming activity of human
papillomavirus type 16 in cervical cancer. J Biol Chem.
282:26381–26391. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
154
|
Wu SX, Ren XY, Pan YL, Guan DD, Liu S, Zou
XY, Shi CG, Li YZ and Zhang YZ: Effects of ALA-PDT on
HPV16-immortalized cervical epithelial cell. Neoplasma. 64:175–181.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
155
|
Wu S, Wang L, Ren X, Pan Y, Peng Y, Zou X,
Shi C and Zhang Y: Involvement of retinoblastoma-associated protein
48 during photodynamic therapy of cervical cancer cells. Mol Med
Rep. 15:1393–1400. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
156
|
Zhong J, Yang X, Mai M, Wang D, Lv L and
Rao J: Effect of RbAp48 knockdown on migration and invasion of
human cervical cancer cell line MS751 in vitro. Nan Fang Yi Ke Da
Xue Xue Bao. 35:1564–1569. 2015.In Chinese. PubMed/NCBI
|
|
157
|
Zheng L, Tang W, Wei F, Wang H, Liu J, Lu
Y, Cheng Y, Bai X, Yu X and Zhao W: Radiation-inducible protein
RbAp48 contributes to radiosensitivity of cervical cancer cells.
Gynecol Oncol. 130:601–608. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
158
|
Li HJ, Yu PN, Huang KY, Su HY, Hsiao TH,
Chang CP, Yu MH and Lin YW: NKX6.1 functions as a metastatic
suppressor through epigenetic regulation of the
epithelial-mesenchymal transition. Oncogene. 35:2266–2278. 2016.
View Article : Google Scholar
|
|
159
|
Cai L, Wang D, Fisher AL and Wang Z:
Identification of a genetic interaction between the tumor
suppressor EAF2 and the retinoblastoma protein (Rb) signaling
pathway in C. elegans and prostate cancer cells. Biochem Biophys
Res Commun. 447:292–298. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
160
|
Barreiro-Alonso A, Lamas-Maceiras M,
Lorenzo-Catoira L, Pardo M, Yu L, Choudhary JS and Cerdán ME: HMGB1
Protein interactions in prostate and ovary cancer models reveal
links to RNA processing and ribosome biogenesis through NuRD, THOC
and septin complexes. Cancers (Basel). 13:46862021. View Article : Google Scholar : PubMed/NCBI
|
|
161
|
Wang J, He C, Gao P, Wang S, Lv R, Zhou H,
Zhou Q, Zhang K, Sun J, Fan C, et al: HNF1B-mediated repression of
SLUG is suppressed by EZH2 in aggressive prostate cancer. Oncogene.
39:1335–1346. 2020. View Article : Google Scholar :
|
|
162
|
Tournaye H, Krausz C and Oates RD: Novel
concepts in the aetiology of male reproductive impairment. Lancet
Diabetes Endocrinol. 5:544–553. 2017. View Article : Google Scholar
|
|
163
|
Riera-Escamilla A, Vockel M, Nagirnaja L,
Xavier MJ, Carbonell A, Moreno-Mendoza D, Pybus M, Farnetani G,
Rosta V, Cioppi F, et al: Large-scale analyses of the X chromosome
in 2,354 infertile men discover recurrently affected genes
associated with spermatogenic failure. Am J Hum Genet.
109:1458–1471. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
164
|
Liu F, Cao L, Zhang T, Chang F, Xu Y, Li
Q, Deng J, Li L and Shao G: CRL4BRBBP7 targets HUWE1 for
ubiquitination and proteasomal degradation. Biochem Biophys Res
Commun. 501:440–447. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
165
|
Sung H, Ferlay J, Siegel RL, Laversanne M,
Soerjomataram I, Jemal A and Bray F: Global cancer statistics 2020:
GLOBOCAN estimates of incidence and mortality worldwide for 36
cancers in 185 countries. CA Cancer J Clin. 71:209–249. 2021.
View Article : Google Scholar : PubMed/NCBI
|
|
166
|
Yeh HH, Tseng YF, Hsu YC, Lan SH, Wu SY,
Raghavaraju G, Cheng DE, Lee YR, Chang TY, Chow NH, et al: Ras
induces experimental lung metastasis through up-regulation of
RbAp46 to suppress RECK promoter activity. BMC Cancer. 15:1722015.
View Article : Google Scholar : PubMed/NCBI
|
|
167
|
Wang Y, Wang Y, Zhang D, Zhang H, Wang W
and Hu X: Identifying RBBP7 as a promising diagnostic biomarker for
BK virus-associated nephropathy. J Immunol Res. 2022:69347442022.
View Article : Google Scholar : PubMed/NCBI
|
|
168
|
Kim DS, Choi YP, Kang S, Gao MQ, Kim B,
Park HR, Choi YD, Lim JB, Na HJ, Kim HK, et al: Panel of candidate
biomarkers for renal cell carcinoma. J Proteome Res. 9:3710–3719.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
169
|
Wang J, Yang J, Yang Z, Lu X, Jin C, Cheng
L and Wu N: RbAp48, a novel inhibitory factor that regulates the
transcription of human immunodeficiency virus type 1. Int J Mol
Med. 38:267–274. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
170
|
Wang J, Yang Z, Cheng L, Lu L, Pan K, Yang
J and Wu N: Retinoblastoma binding protein 4 represses HIV-1 long
terminal repeat-mediated transcription by recruiting NR2F1 and
histone deacetylase. Acta Biochim Biophys Sin (Shanghai).
51:934–944. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
171
|
Biswas S, Haleyurgirisetty M, Ragupathy V,
Wang X, Lee S, Hewlett I and Devadas K: Differentially expressed
host long intergenic noncoding RNA and mRNA in HIV-1 and HIV-2
infection. Sci Rep. 8:25462018. View Article : Google Scholar : PubMed/NCBI
|
|
172
|
Xu W, Wu Y, Zhao J, Chen J and Zhang W:
Human immunodeficiency virus type 1 transcription is regulated by
thieno[3,4-d] pyrimidine. Exp Ther Med. 19:3090–3096.
2020.PubMed/NCBI
|
|
173
|
Schwartz SM, Daling JR, Doody DR, Wipf GC,
Carter JJ, Madeleine MM, Mao EJ, Fitzgibbons ED, Huang S, Beckmann
AM, et al: Oral cancer risk in relation to sexual history and
evidence of human papillomavirus infection. J Natl Cancer Inst.
90:1626–1636. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
174
|
Lohavanichbutr P, Houck J, Fan W, Yueh B,
Mendez E, Futran N, Doody DR, Upton MP, Farwell DG, Schwartz SM, et
al: Genomewide gene expression profiles of HPV-positive and
HPV-negative oropharyngeal cancer: Potential implications for
treatment choices. Arch Otolaryngol Head Neck Surg. 135:180–188.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
175
|
Wurlitzer M, Möckelmann N, Kriegs M, Vens
M, Omidi M, Hoffer K, Bargen CV, Möller-Koop C, Witt M, Droste C,
et al: Mass spectrometric comparison of HPV-Positive and
HPV-Negative oropharyngeal cancer. Cancers (Basel). 12:15312020.
View Article : Google Scholar : PubMed/NCBI
|
|
176
|
Kaushik M, Nehra A, Gakhar SK, Gill SS and
Gill R: The multifaceted histone chaperone RbAp46/48 in Plasmodium
falciparum: Structural insights, production, and characterization.
Parasitol Res. 119:1753–1765. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
177
|
Kaushik M, Nehra A, Gill SS and Gill R:
Unraveling CAF-1 family in Plasmodium falciparum: Comparative
genome-wide identification and phylogenetic analysis among
eukaryotes, expression profiling and protein-protein interaction
studies. 3 Biotech. 10:1432020. View Article : Google Scholar : PubMed/NCBI
|
|
178
|
Chen H, Zhang W, Luo S, Li Y, Zhu Q, Xia
Y, Tan H, Bian Y, Li Y, Ma J, et al: Lead exposure induces neuronal
apoptosis via NFκB p65/RBBP4/Survivin signaling pathway.
Toxicology. 499:1536542023. View Article : Google Scholar
|
|
179
|
Jia W, Shen J, Wei S, Li C, Shi J, Zhao L
and Jia H: Ropivacaine inhibits the malignant behavior of lung
cancer cells by regulating retinoblastoma-binding protein 4. PeerJ.
11:e164712023. View Article : Google Scholar : PubMed/NCBI
|
|
180
|
Torres-Roca JF, Eschrich S, Zhao H, Bloom
G, Sung J, McCarthy S, Cantor AB, Scuto A, Li C, Zhang S, et al:
Prediction of radiation sensitivity using a gene expression
classifier. Cancer Res. 65:7169–7176. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
181
|
Hart P, Hommen P, Noisier A, Krzyzanowski
A, Schüler D, Porfetye AT, Akbarzadeh M, Vetter IR, Adihou H and
Waldmann H: Structure based design of bicyclic peptide inhibitors
of RbAp48. Angew Chem Int Ed Engl. 60:1813–1820. 2021. View Article : Google Scholar
|