Introduction
Glioma is the most common brain tumor in adults. Its
ability to evade immune surveillance and impede antitumor responses
may lead to sustained growth and enhanced malignancy (1). Despite attempts at treatment using
surgical resection, radiation and chemotherapy with the alkylating
agent temozolomide, the survival rates of patients with high-grade
gliomas are <10% at 5 years (2). Since current treatment gained little
benefit in the setting of glioma, greater attention has been paid
to the expression of specific molecular markers with the goal of
understanding the main molecular mechanisms of this malignancy and
determining their possible prognostic significance.
The speckle-type POZ protein (SPOP) has been
identified as an autoantigen in scleroderma patients and a
constituent of nuclear speckles in human cells. SPOP is a 374-amino
acid protein that contains a C-terminal POZ (poxvirus and zinc
finger) domain (also known as a BTB domain) and an N-terminal MATH
(meprin and TRAF-C homology) domain (3). The C-terminal POZ domain of SPOP
interacts directly with Cullin3-based E3 ubiquitin ligase complex,
while its N-terminal MATH domain interacts with various substrates
of the complex. In mammals, the MATH-BTB protein SPOP has been
linked to ubiquitination of MacroH2A, to regulate its deposition on
the inactive X-chromosome (4), and
has been shown to act as an adaptor of Daxx in the ubiquitination
process involving Cul3-based ubiquitin ligase in the Hedgehog/Gli
signaling pathway, to regulate transcriptional repression of
pro-apoptotic proteins such as p53 (5). Recent genome-wide somatic mutation
analyses identified that SPOP gene encoding the speckle-type POZ
protein was frequently mutated in some human cancers, including
prostate and endometrial carcinomas (6–9).
Ubiquitin modifications regulate a wide variety of cellular
processes and more or less are involved in cancer pathogenesis
(10). Also, earlier observations
that SPOP, an adaptor for Cul3-based ubiquitination (6–11),
may be a TSG led us to further analyze whether SPOP gene mutations
occurred in other common malignancies. In one recent study, we
found that only 0.2% of hematologic malignancies and none of the
solid tumors (breast, lung and liver cancers) harbored SPOP somatic
mutations, suggesting that somatic mutation of SPOP gene may not
play a role in the development of breast, lung, and liver cancers,
and acute leukemias.
In addition to the somatic mutation, altered
expression of ubiquitination related genes is observed in many
cancers (12,13). An earlier study described analyzed
SPOP protein was expressed in normal gastric, colonic and prostate
epithelial cells, whereas it was lost in 30% of gastric cancer, 20%
of colorectal cancer and 37% of prostate cancer (14).
SPOP plays a key role in the development of
peripheral and central nervous system and tumorigenesis (15). Furthermore, it was recently
demonstrated that knockdown of Drosophila SPOP mRNA
expression by RNA interference (RNAi) and P-element insertion
mutagenesis of the SPOP resulted in severe and consistent
disruption of the peripheral and the CNS (16). Also, our findings support the
genomewide linkage results pointing towards disease loci on
17q12-21.32 and 12p13.33-12.1 for glioma risk in families.
Furthermore, our mapping efforts have identified plausible
candidate genes for further study, one of which is SPOP. The
identification of the causal variants of the gene is likely to
clarify the molecular mechanisms underlying gliomagenesis (17). So far, the prevalence of decreased
SPOP expression has not been extensively reported in glioma,
especially with regard to the effect of decreased SPOP expression
in glioma. Here, in this study, we surveyed the expression of SPOP
in human patient samples. To explore its associated molecular
mechanisms in glioma cells, we examined the effect of targeted
overexpression of SPOP gene on cell viability, migration and
invasion in vitro. These studies will be useful in
identifying potential candidates for targeted therapeutic
intervention of glioma.
Materials and methods
Human tissue samples
A total of 98 paraffin-embedded glioma and 12 NB
tissues samples were obtained from patients who underwent surgical
treatment at Xiangya Hospital during the period from January 2005
to January 2010. These patients or their legal guardian provided
written informed consent to the surgical procedures and gave
permission to use resected tissue specimens for research purposes.
A diagnosis of glioma was confirmed pathologically by two
independent, experienced pathologists. All specimens had confirmed
pathological diagnosis and were classified according to the World
Health Organization (WHO) criteria. The follow-up data of the
glioma patients in this study are available and complete. Overall
survival, which was defined as the time from the operation to the
time of patient death or the last follow-up, was used as a measure
of prognosis. Postoperative follow-up occurred at our outpatient
department and included clinical and laboratory examinations every
3 months for the first 2 years, every 6 months during the 3rd to
5th years, annually for an additional 5 years or until patient
death, whichever occurred first.
Cell culture and transfection
Human glioma cell strains of U87 and U251 were
purchased from a cell bank at the Chinese Academy of Sciences and
grown in Dulbecco’s modified Eagle’s medium (DMEM) (Hyclone, Logan,
UT, USA) supplemented with 10% fetal calf serum (Gibco, Grand
Island, NY, USA). All cell lines were cultured at 37°C in a
humidified atmosphere of 5% CO2. Transfection reagent
Lipofectamine 3000 was purchased from Invitrogen (St. Louis, MO,
USA). For overexpression of SPOP, the full-length SPOP cDNA was
amplified and cloned into the pCDNA3.1(+) expression vector
(Invitrogen; Life Technologies). U87 and U251 cells were then
transfected with a negative empty vector or a SPOP expressing
plasmid using Lipofectamine 3000 according to the manufacturer’s
instructions. Cells were collected after 48–72 h for further
experiments.
Real-time quantitative PCR
Total RNA was extracted from tissues lysate using a
TRIzol kit (Invitrogen, Carlsbad, CA, USA), and cDNA was
subsequently synthesized from total RNA using an Omniscript RT kit
(Qiagen, Valencia, CA, USA) following the supplier’s instructions.
For detecting the mRNA level of SPOP, quantitative real-time PCR
was conducted on the Mastercycler ep realplex (Eppendorf 2S,
Hamburg, Germany). A 25-μl reaction mixture contained 1 μl of cDNA
from samples, 12.5 μl of 2X Fast EvaGreen™ qPCR Master Mix (Biotium
Inc., Hayward, CA, USA), 1 μl primers (10 mM), and 10.5 μl of
RNase/DNase-free water. PCR procedures: incubation at 96°C for 2
min, 40 cycles at 96°C for 15 sec and 60°C for 1 min. The Ct value
was defined as the cycle number at which the fluorescence intensity
reached a certain threshold where amplification of each target gene
was within the linear region of the reaction amplification curves.
Relative expression level for each target gene was normalized by
the Ct value of β-actin (internal control) using a
2−ΔΔCt relative quantification method. The sequences of
the primers for SPOP are as follows: SPOP forward,
5′-TGACCACCAGGTAGACAGCG-3′; SPOP reverse, 5′-CCCGTTTCCCCCAAGTTA-3′.
The β-actin gene served as an internal control. The sequences of
the primers for β-actin are as follows: β-actin forward,
5′-AACTTCCGTTGCTGCCAT-3′; β-actin reverse,
5′-TTTCTTCCACAGGGCTTTG-3′. The 2−ΔΔCt method was used to
calculate relative changes in gene expression.
Western blot analysis
The homogenized glioma cancer samples, including
tumor and nomal brain tissues, as well as cell lines, were lysed in
RIPA lysis buffer, and the lysates were harvested by centrifugation
(12,000 rpm) at 4°C for 20 min. Approximately 20 μg protein samples
were then separated by electrophoresis in a 12% sodium dodecyl
sulfate polyacrylamide gel and transferred onto a polyvinylidene
fluoride membrane. After blocking the non-specific binding sites
for 60 min with 5% non-fat milk, the membranes were incubated
overnight at 4°C with a goat polyclonal antibody against SPOP
(1:500, sc-66649, Santa Cruz Biotechnology, USA). The membranes
were then washed three times with TBST (Tris-buffered saline with
Tween-20) for 10 min and probed with the horseradish peroxidase
(HRP)-conjugated donkey anti-goat IgG antibody (1:2,000, Immunology
Consultants Laboratory, USA) at 37°C for 1 h. After three washes,
the membranes were developed by an enhanced chemiluminescence
system (Cell Signaling Technology, Danvers, MA, USA). The band
intensity was measured by densitometry using the Quantity One
software (Bio-Rad Laboratories, Inc. Hercules, CA, USA). The
protein levels were normalized to β-actin.
Immunohistochemistry analysis
The tissue sections were deparaffinized with
dimethylbenzene and rehydrated with 100, 95, 90, 80, 70 and 50%
ethanol. After three washes in phosphate-buffered saline (PBS), the
slides were boiled in antigen retrieval buffer containing 0.01 M
sodium citrate-hydrochloric acid (pH 6.0) for 15 min in a microwave
oven. After rinsing with PBS, the tissue sections were incubated
with goat polyclonal anti-human SPOP antibody (1:100, sc-66649,
Santa Cruz Biotechnology, Inc., USA) and then rinsed in 3%
peroxidase quenching solution (Invitrogen) to block endogenous
peroxidase. The sections were then incubated with a donkey
anti-goat second antibody conjugated horseradish peroxidase
(1:5,000; Abcam, Cambridge, UK) at 4°C overnight and after washing
in PBS, the visualization signal was developed with 3,
3′-diaminobenzidine (DAB) solution, and all of the slides were
counterstained with hematoxylin. As negative controls, adjacent
sections were processed as described above except that they were
incubated overnight at 4°C in blocking solution without the primary
antibody. The specimens were analyzed by two observers who were
blinded to the patients’ clinical outcomes. Discrepancies between
the observers were found in <10% of the examined slides, and a
consensus was reached after further review. The total SPOP
immunostaining score was calculated as the sum of the percentage of
positively stained tumor cells and the staining intensity and
ranged from 0 to 9. Briefly, the percentage of positive staining
was scored as 0 (0–9%, negative), 1 (10–25%, sporadic), 2 (26–50%,
focal) or 3 (51–100%, diffuse), and the intensity was scored as 0
(no staining), 1 (weak staining), 2 (moderate staining) or 3
(strong staining). The expression level of SPOP was defined as
follows: ‘−’ (negative, score of 0), ‘+’ (weakly positive, score of
1–3), ‘++’ (positive, score of 4–6), ‘+++’ (strongly positive,
score of 7–9). We defined strong SPOP expression as a total score
of >3, and weak SPOP expression as a total score of ≤3.
Cell viability and proliferation
assay
The tetrazolium-based cell viability (MTT) assay was
performed to test cell proliferation. Cells transfected SPOP
plasmid or empty vector were seeded in a 96-well plate at
1×103 cells/well, 100 μl of sterile MTT dye (0.5 mg/ml,
Sigma) was added to each well and cultured for another 4 h. The
supernatant was discarded and then 150 μl of dimethyl sulphoxide
(DMSO) (Sigma, St. Louis, MO, USA) was added to each well, the
spectrophotometric absorbance was measured for each sample at 490
nm, all the experiments were performed in triplicate and repeated 3
times, and the average was calculated.
Cell migration assays
The cell migratory capacity was determined using
transwell chambers (Corning, Corning, NY, USA). Briefly, cells
(1×105/well) were suspended in 100 μl serum-free medium
and then added to the upper chamber of the inserts, DMEM medium
(Gibco) containing 10% FBS (500 μl) was added to the lower chamber
as the chemotactic factor. After culture for 24 h at 37°C,
non-migrated cells on the upper surface were removed gently with a
cotton swab and cells that migrated to the lower side of the
department were fixed and dyed with 0.1% crystal violet. The number
of migrated cells were calculated by counting five different views
by microscopy. Independent experiments were repeated three times,
with triplicates in each experiment.
Statistical analysis
All quantified data represent an average of at least
triplicate samples. SPSS 18.0 (SPSS Inc, Chicago, IL, USA) was used
for statistical analysis. Data are presented as mean ± SD. One-way
ANOVA or two-tailed Student’s t-test was used for comparisons
between groups. Chi-square test or Fischer’s were used to identify
differences between categorical variables. Survival analysis was
performed using Kaplan-Meier method. Multivariate Cox proportional
hazards method was used for analyzing the relationship between the
variables and patient’s survival time. Values of P<0.05 were
considered to indicate statistically significant results in all
cases.
Results
Downregulated expression of SPOP gene in
glioma tissues
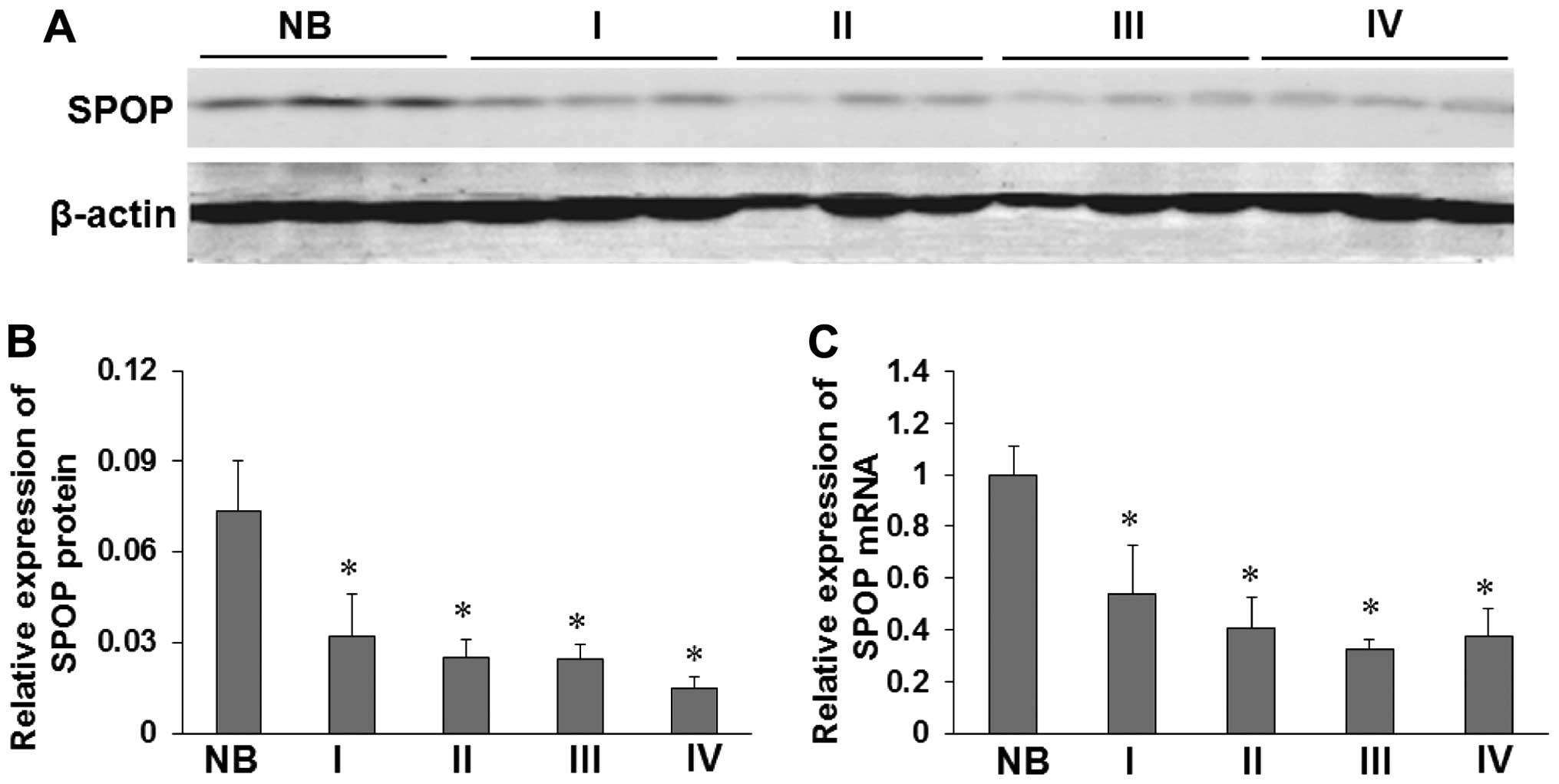
In order to assess the role of SPOP in glioma, we
performed western blotting and real-time PCR to measure the
expression of SPOP in 35 freshly collected glioma tissues and 12
freshly collected NB tissues. SPOP protein was found to be markedly
downregulated in 24 cases of glioma (WHO I–IV) compared with 6 NB
tissues with an increase in pathologic grade of the brain gliomas
(P<0.05, Fig. 1A and B).
Compared with NB tissues, glioma tissues exhibited lower expression
levels of SPOP mRNA (P<0.05) (Fig.
1C). We also measured the expression levels and subcellular
localization of SPOP protein in 98 archived paraffin-embedded
glioma samples and 12 NB tissues using immunohistochemical staining
(Fig. 2). SPOP protein showed low
expression in 62.2% (61/98) of glioma samples, while high
expression in 75% (9/12) of NB samples, and the difference was
statistically significant (P=0.014; Table I).
 | Table ISPOP expression compared in glioma and
NB tissues. |
Table I
SPOP expression compared in glioma and
NB tissues.
| | Expression | |
|---|
| |
| |
|---|
| Group | Value (n) | Low expression
(n) | High expression
(n) | P-value |
|---|
| Glioma | 98 | 61 | 37 | 0.014 |
| Normal | 12 | 3 | 9 | |
The correlations between the expression
of SPOP and various clinicopathological characteristics
The relationship between clinicopathological
characteristics and SPOP expression levels in individuals with
glioma were analyzed by the Chi-square analysis. We found no
significant association between SPOP expression levels and the
patient age, gender, increased ICP, cystic change, tumor necrosis,
resection degree, chemotherapy or radiotherapy in the 98 glioma
cases. However, we observed that the expression level of SPOP was
positively correlated with mean tumor diameter (MTD) (P=0.021) and
the status of tumor grade and histological type (WHO I, II, III and
IV) (P=0.032) in glioma patients (Table II).
| Table IICorrelation between the
clinicopathological characteristics and expression of SPOP in
glioma. |
Table II
Correlation between the
clinicopathological characteristics and expression of SPOP in
glioma.
| Characteristics | Value (n) | SPOP expression Low
(n) | SPOP expression High
(n) | P-value |
|---|
| Gender | −98 | 61 | 37 | 0.421 |
| Male | 58 | 38 | 20 | |
| Female | 40 | 23 | 17 | |
| Age | | | | 0.752 |
| ≤50 | 31 | 20 | 11 | |
| >50 | 67 | 41 | 26 | |
| MTD (cm) | | | | 0.021a |
| ≤5 | 41 | 31 | 10 | |
| ≥5 | 57 | 30 | 27 | |
| Increased ICP | | | | 0.226 |
| No | 63 | 42 | 21 | |
| Yes | 35 | 19 | 16 | |
| Cystic change | | | | 0.578 |
| No | 71 | 43 | 28 | |
| Yes | 27 | 18 | 9 | |
| Tumor necrosis | | | | 0.136 |
| No | 79 | 52 | 27 | |
| Yes | 19 | 9 | 10 | |
| Resection
degree | | | | 0.334 |
| Gross total
resection | 80 | 48 | 32 | |
| Subtotal
resection | 18 | 13 | 5 | |
| Tumor grade and
histological type | | | | 0.032a |
| WHO grade I |
| Pilocytic
astrocytoma | 14 | 9 | 5 | |
| WHO grade II |
| Astrocytoma | 19 | 16 | 3 | |
|
Oligodendroglioma | 7 | 4 | 3 | |
| WHO grade III |
| Anaplastic
astrocytoma | 25 | 18 | 7 | |
| WHO grade IV |
| Glioblastoma
multiforme | 33 | 14 | 19 | |
| Chemotherapy | | | | 0.176 |
| No | 15 | 7 | 8 | |
| Yes | 83 | 54 | 29 | |
| Radiotherapy | | | | 0.384 |
| No | 17 | 9 | 8 | |
| Yes | 81 | 52 | 29 | |
The role of SPOP in proliferation and
migration in the cell lines
Because our above results indicated that SPOP
expression was reduced in glioma and that SPOP might act as a tumor
suppressor, we next tried to explore the function of SPOP in glioma
development. To evaluate the effects of SPOP on cell viability, the
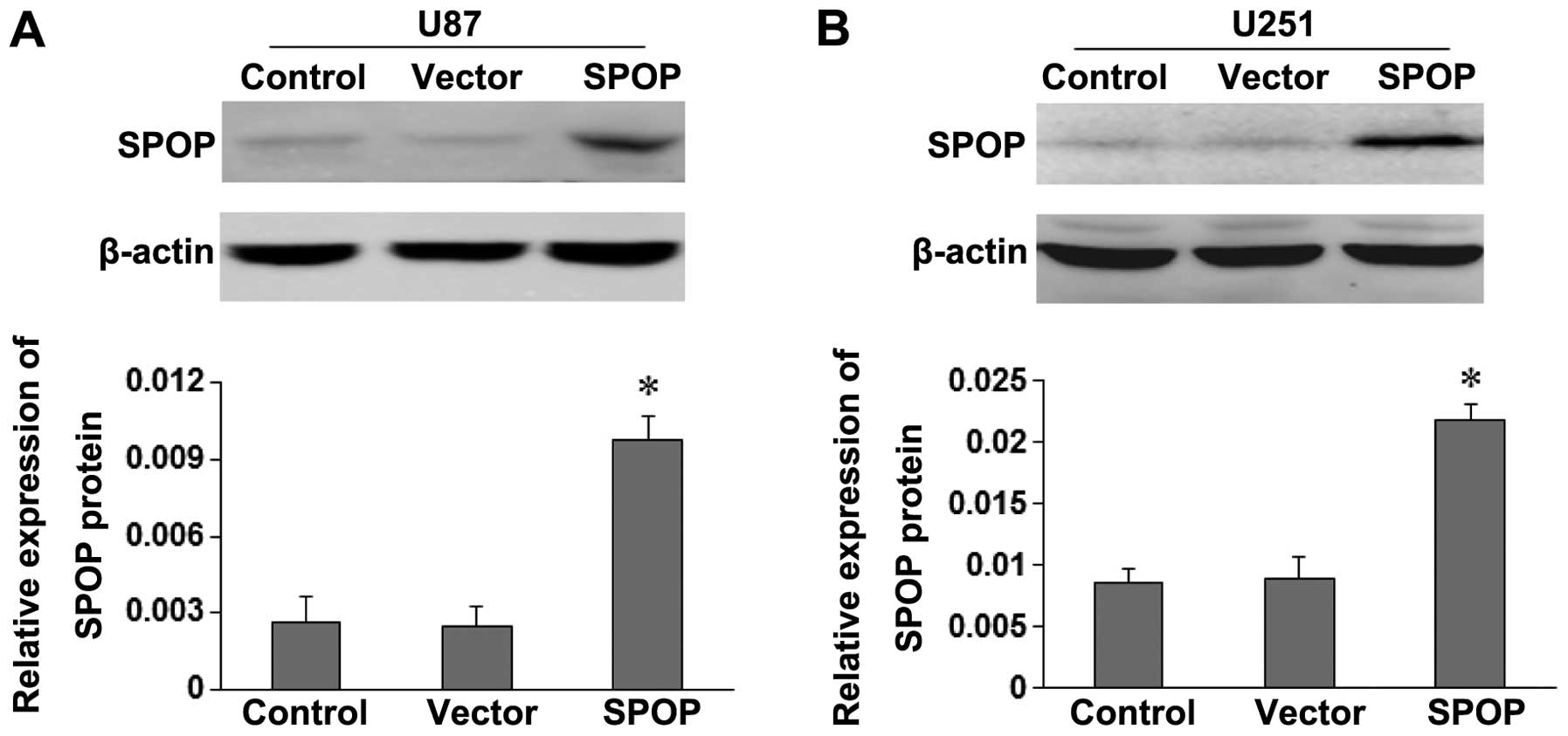
SPOP expression vector or the empty vector were, respectively,
transfected into U87 and U251 cells. SPOP expression in transfected
cells were detected by western blotting (Fig. 3). The expression of SPOP protein in
pCDNA3.1-SPOP group (SPOP) was significantly higher than control
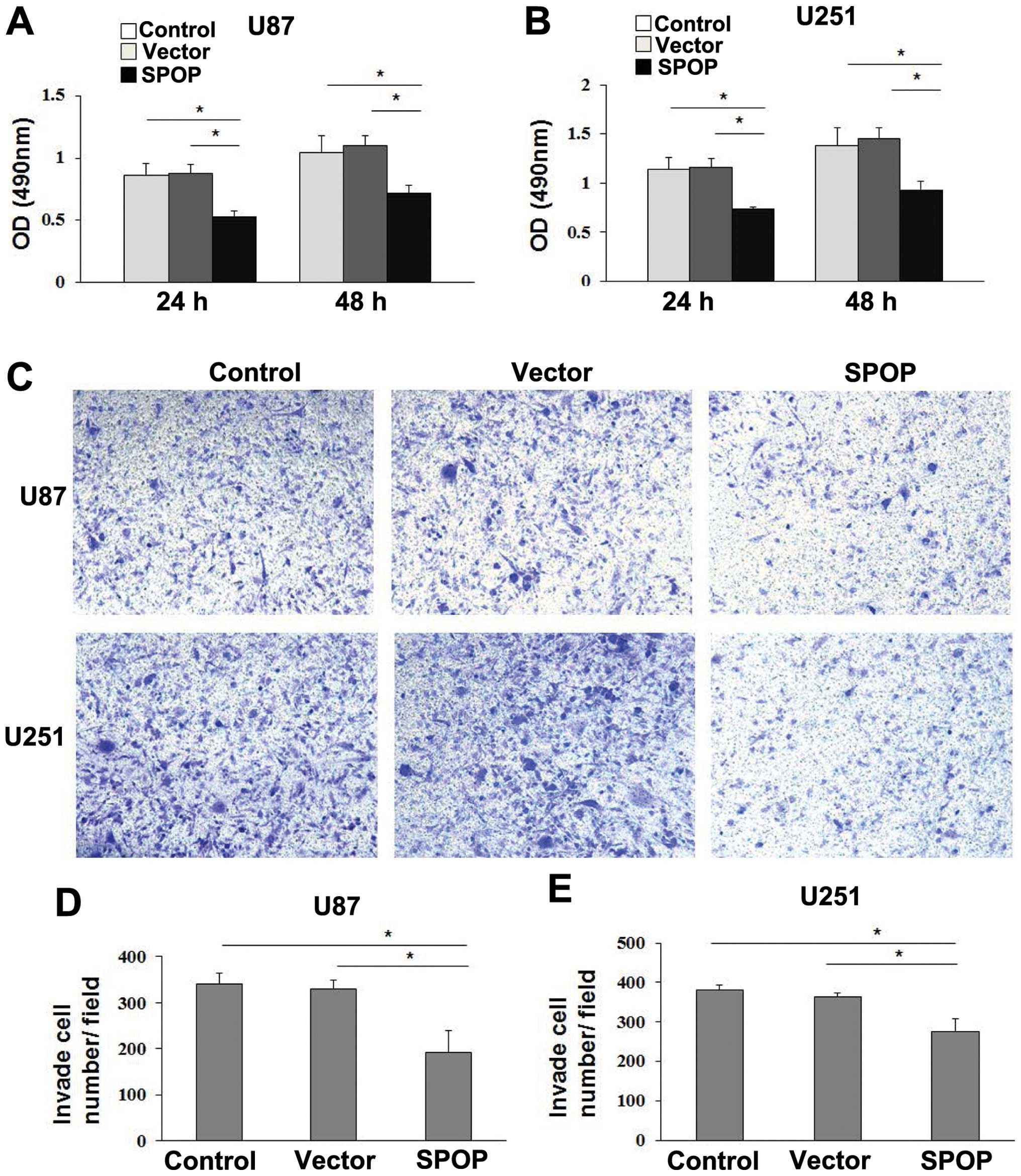
and empty vector groups. The cell growth by MTT assay revealed that
cell growth rate in SPOP-transfected glioma cells were
significantly lower than control or empty vector-transfected glioma
cells (Fig. 4A and B, P<0.05),
which showed that SPOP significantly repressed the viability of
glioma cells.
We evaluated the potential role of SPOP on cellular
migration by transwell assays. U87 and U251 cells were transfected
with SPOP overexpressing or empty vector plasmid and their
migratory abilities were determined 24 h later. The results showed
overexpression of SPOP was associated with a significant reduction
of migration compared to the empty vector (Fig. 4C–E, P<0.01).
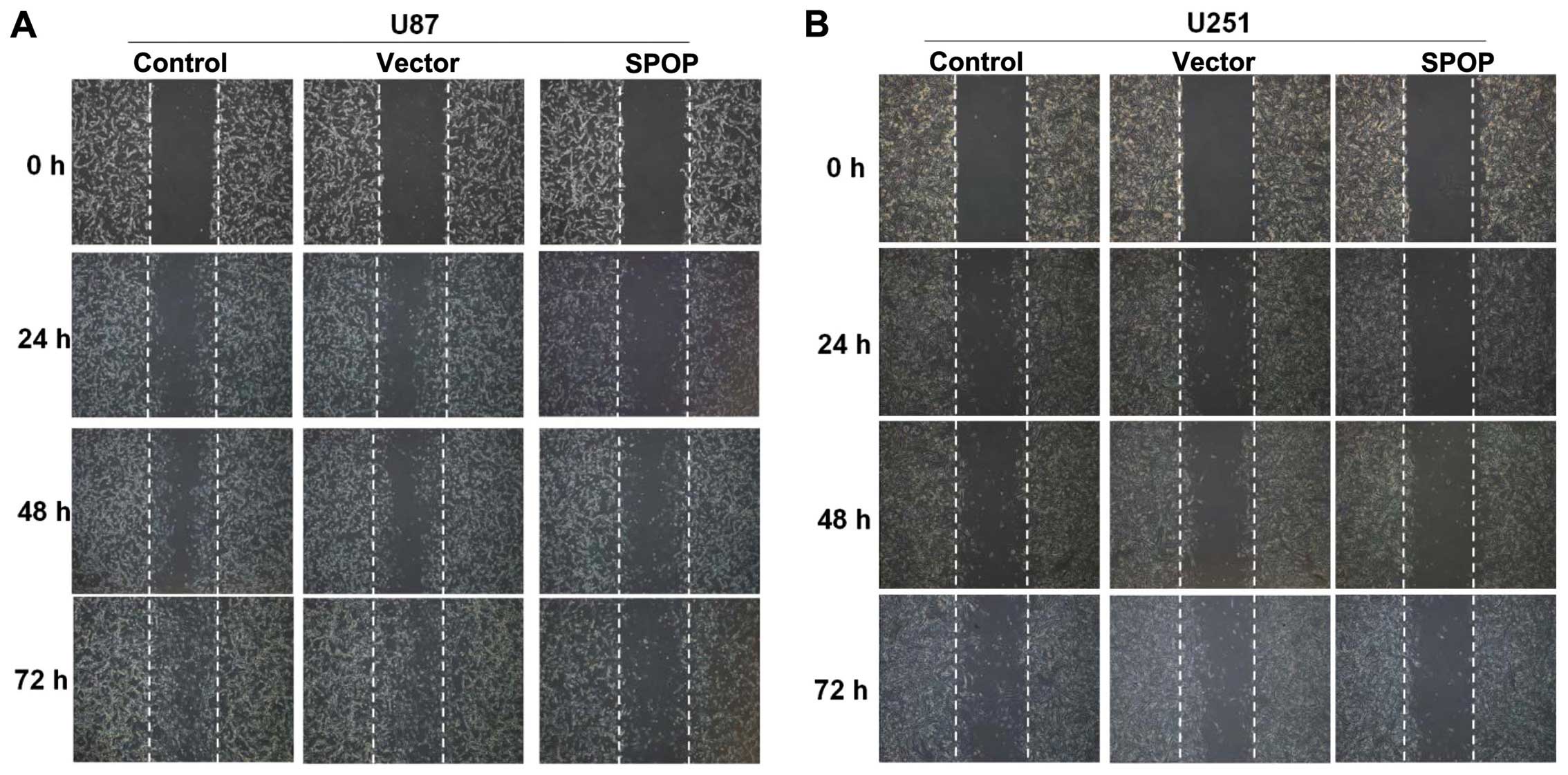
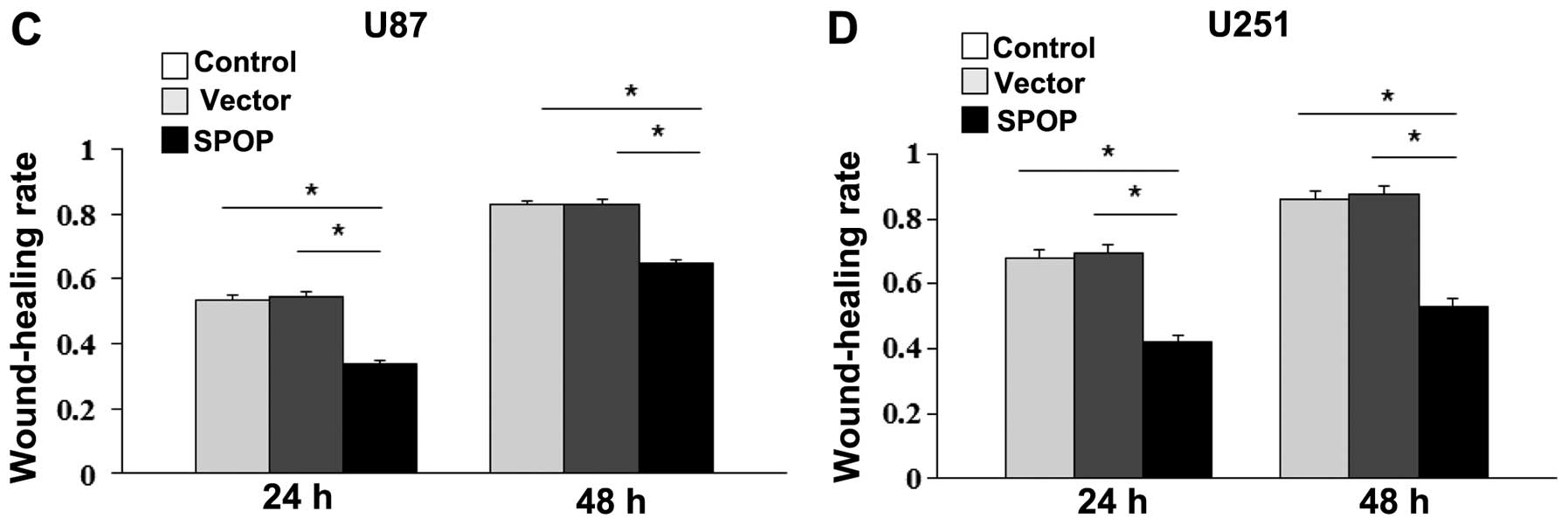
These findings were further confirmed by the wound
healing assay. The overexpression of SPOP significantly inhibited
the migration of U87 and U251 cells at 48 h after transfection
(Fig. 5, P<0.05).
Expression of SPOP and clinical
outcome
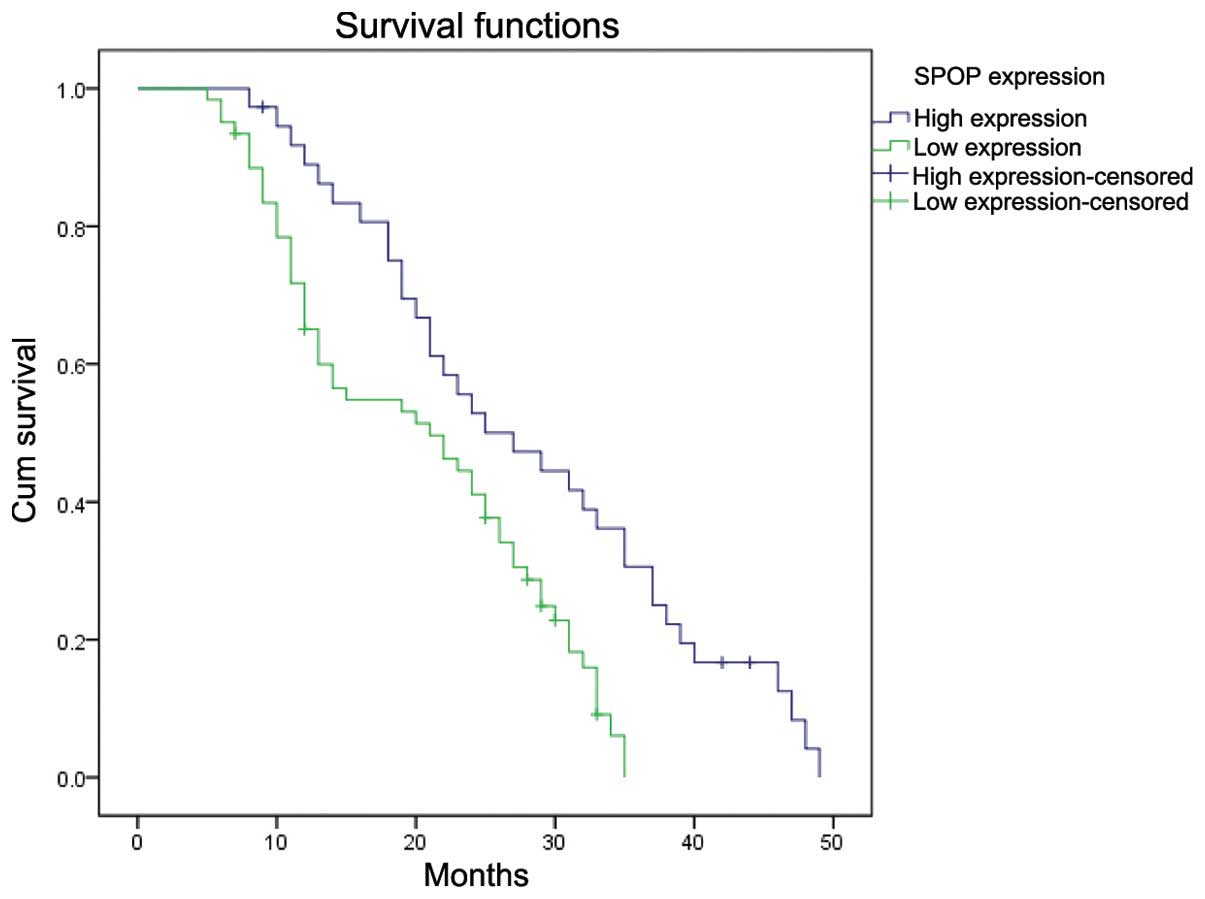
The overall survival of patients with low SPOP
expression was significantly worse than that of SPOP-high patients
(P=0.001, log-rank test, Fig. 6).
Univariate Cox regression analyses showed that MTD and SPOP
expression were significantly associated with overall survival
(Table III). Furthermore, a
multivariate Cox regression analysis confirmed the MTD and SPOP
expression as independent predictors of the overall survival of
glioma cancer patients (Table
III).
| Table IIIUnivariate and multivariate analyses
of overall survival of glioma patients. |
Table III
Univariate and multivariate analyses
of overall survival of glioma patients.
| Univariate
analysis | | Multivariate
analysis | |
|---|
|
| |
| |
|---|
| Variables | HR | 95% CI | P-value | HR | 95% CI | P-value |
|---|
| Gender (male vs
female) | 2.138 | 1.437–3.516 | 0.132 | 2.053 | 1.267–3.326 | 0.223 |
| Age (≤50 vs
>50) | 0.842 | 0.433–1.668 | 0.460 | 0.765 | 0.384–1.537 | 0.414 |
| MTD (cm) (≤5 vs
≥5) | 1.031 | 0.716–1.514 | 0.034a | 1.329 | 0.887–1.915 | 0.027a |
| Increased ICP (no
vs yes) | 0.977 | 0.606–1.575 | 0.942 | 1.227 | 0.749–1.908 | 0.417 |
| Cystic change (no
vs yes) | 0.525 | 0.342–0.805 | 0.287 | 0.611 | 0.393–0.950 | 0.512 |
| Tumor necrosis (no
vs yes) | 0.546 | 0.316–0.944 | 0.430 | 0.445 | 0.268–0.739 | 0.362 |
| Resection degree
(gross total vs subtotal) | 0.616 | 0.391–1.004 | 0.183 | 0.624 | 0.387–1.078 | 0.112 |
| WHO (I vs II vs III
vs IV) | 2.118 | 1.328–3.379 | 0.142 | 2.896 | 1.719–4.878 | 0.131 |
| Chemotherapy (no vs
yes) | 0.896 | 0.787–1.106 | 0.357 | 0.799 | 0.689–1.010 | 0.610 |
| Radiotherapy (no vs
yes) | 0.832 | 0.743–1.018 | 0.474 | 0.762 | 0.613–1.008 | 0.762 |
| SPOP expression
(low vs high) | 0.548 | 0.346–0.893 | 0.018a | 0.439 | 0.298–0.764 | 0.012a |
Discussion
Glioma, the most common type of primary brain tumors
in adults, is subcategorized by histopathologic evaluation and
clinical criteria into four grades (I–IV), according to the current
World Health Organization (WHO) guidelines (18). Despite multimodal treatment
including surgery, chemotherapy and radiation, the average survival
time of glioma patients is 10–14 months after diagnosis in the last
5 years (19). The extremely poor
prognosis of these patients may be due to the biological
characteristics of glioma cells which include unrestricted
proliferation and extensive invasion (20). Therefore, understanding the main
molecular mechanisms of this malignancy is the key for the
development of novel and effective therapeutic strategies for
glioma. In this study, we investigated the expression of
speckle-type POZ domain protein (SPOP) in glioma. We found that the
expression of SPOP was significantly reduced at both mRNA and
protein levels in glioma compared with normal brain tissues. We
also found that glioma patients with low expression of SPOP showed
shorter postsurgical survival than high SPOP expression patients.
Notably, we show for the first time that SPOP is expressed in
normal brain, supporting further research on the potential role of
SPOP in brain development. Instead, our data, obtained by
modulating SPOP expression, establish a role for SPOP in modulating
the biological properties of glioma cells, including proliferation,
migration and in vitro aggressiveness. Overexpression of
SPOP resulted in suppression of cell proliferation, arrest of cell
migration. This evidence suggests a potential role for SPOP in the
regulation of glioma cell growth and proliferation which are in
agreement with previous studies (11).
In humans, the roles for SPOP in regulating the Hh
and TNF pathways have been conserved (21,22),
and several other SPOP substrates have been identified as well,
including the death domain-associated protein (Daxx) (5), the polycomb group protein BMI-1, the
phosphatidylinosital phosphate kinase PIPKIIβ (23), the transcription factor Gli
(24), the Jun-kinase phosphatase
Puckered (14) and Pdx1 (25), and the histone variant MacroH2A
(4). Together, these previous
results indicate that SPOP plays important roles during cell
apoptosis, proliferation, and animal development. Many proteins
implicated in the ubiquitin pathways are related to tumor
progression. In recent years, there have been many approaches to
target the ubiquitin system in cancer therapy (26,27).
The E3 ubiquitin ligases are considered the most important
components in these approaches because they bind directly to the
target proteins (26). In the
present study, we evaluated the role of SPOP as an adaptor in the
Cullin3-based E3 ubiquitin ligase complex, which indicated that
SPOP might play a role in tumorigenesis as a tumor suppressor gene.
Moreover, our experimental results from breast cancer cell
proliferation, invasion, anchorage-independent growth, and tumor
growth in nude mice indicate that SPOP inhibits cancer cell
functions. We also found that shRNA knockdown of SPOP in MDA-MB-231
cells increases cancer cell size (11), consistent with previous reports
(28).
Considering pro-apoptotic and other
tumor-suppressive functions of SPOP identified in earlier studies
(11,29), we suggest that downregulation of
SPOP in the cancers compared with normal cells might possibly
decrease its functions as a tumor suppressor gene and might
contribute to cancer development. Although most of the identified
functions of SPOP are related to tumor suppression, there is
evidence that SPOP serves as a regulatory hub to promote clear cell
renal cell carcinoma (ccRCC) tumorigenesis suggesting a possibility
that tumor-related functions of SPOP might vary depending on tissue
and cellular contexts (30).
However, in this study, we have reasons to believe that SPOP could
be a new therapeutic target for improving the treatment efficiency
of glioma.
In conclusion, in this study, we investigated the
role of SPOP in human glioma for the first time. Our results
suggested that SPOP was downexpressed in glioma compared with
normal brain tissues, and low expression of SPOP was significantly
associated with poorer overall survival. Additionally, we provide
compelling evidence that overexpression of SPOP leads to suppressed
cell viability, migration and invasion in glioma cell lines in
vitro. SPOP expression may have significant value as an
unfavorable progression indicator for glioma patients.
References
|
1
|
Louis DN: Molecular pathology of malignant
gliomas. Annu Rev Pathol. 1:97–117. 2006. View Article : Google Scholar
|
|
2
|
Stupp R, Hegi ME, Mason WP, van den Bent
MJ, Taphoorn MJ, Janzer RC, Ludwin SK, Allgeier A, Fisher B,
Belanger K, Hau P, Brandes AA, Gijtenbeek J, Marosi C, Vecht CJ,
Mokhtari K, Wesseling P, Villa S, Eisenhauer E, Gorlia T, Weller M,
Lacombe D, Cairncross JG and Mirimanoff RO; European Organisation
for Research and Treatment of Cancer Brain Tumour and Radiation
Oncology Groups; National Cancer Institute of Canada Clinical
Trials Group: National Cancer Institute of Canada Clinical Trials
Group. Effects of radiotherapy with concomitant and adjuvant
temozolomide versus radiotherapy alone on survival in glioblastoma
in a randomised phase III study: 5-year analysis of the EORTC-NCIC
trial. Lancet Oncol. 10:459–466. 2009.
|
|
3
|
Nagai Y, Kojima T, Muro Y, Hachiya T,
Nishizawa Y, Wakabayashi T and Hagiwara M: Identification of a
novel nuclear speckle-type protein, SPOP. FEBS Lett. 418:23–26.
1997. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Hernández-Muñoz I, Lund AH, van der Stoop
P, Boutsma E, Muijrers I, Verhoeven E, Nusinow DA, Panning B,
Marahrens Y and van Lohuizen M: Stable X chromosome inactivation
involves the PRC1 Polycomb complex and requires histone MACROH2A1
and the CULLIN3/SPOP ubiquitin E3 ligase. Proc Natl Acad Sci USA.
102:7635–7640. 2005.PubMed/NCBI
|
|
5
|
Kwon JE, La M, Oh KH, Oh YM, Kim GR, Seol
JH, Baek SH, Chiba T, Tanaka K, Bang OS, Joe CO and Chung CH: BTB
domain-containing speckle-type POZ protein (SPOP) serves as an
adaptor of Daxx for ubiquitination by Cul3-based ubiquitin ligase.
J Biol Chem. 281:12664–12672. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Kan Z, Jaiswal BS, Stinson J, Janakiraman
V, Bhatt D, Stern HM, Yue P, Haverty PM, Bourgon R, Zheng J,
Moorhead M, Chaudhuri S, Tomsho LP, Peters BA, Pujara K, Cordes S,
Davis DP, Carlton VE, Yuan W, Li L, Wang W, Eigenbrot C, Kaminker
JS, Eberhard DA, Waring P, Schuster SC, Modrusan Z, Zhang Z, Stokoe
D, de Sauvage FJ, Faham M and Seshagiri S: Diverse somatic mutation
patterns and pathway alterations in human cancers. Nature.
466:869–873. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Berger MF, Lawrence MS, Demichelis F,
Drier Y, Cibulskis K, Sivachenko AY, Sboner A, Esgueva R, Pflueger
D, Sougnez C, Onofrio R, Carter SL, Park K, Habegger L, Ambrogio L,
Fennell T, Parkin M, Saksena G, Voet D, Ramos AH, Pugh TJ,
Wilkinson J, Fisher S, Winckler W, Mahan S, Ardlie K, Baldwin J,
Simons JW, Kitabayashi N, MacDonald TY, Kantoff PW, Chin L, Gabriel
SB, Gerstein MB, Golub TR, Meyerson M, Tewari A, Lander ES, Getz G,
Rubin MA and Garraway LA: The genomic complexity of primary human
prostate cancer. Nature. 470:214–220. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Barbieri CE, Baca SC, Lawrence MS,
Demichelis F, Blattner M, Theurillat JP, White TA, Stojanov P, Van
Allen E, Stransky N, Nickerson E, Chae SS, Boysen G, Auclair D,
Onofrio RC, Park K, Kitabayashi N, MacDonald TY, Sheikh K, Vuong T,
Guiducci C, Cibulskis K, Sivachenko A, Carter SL, Saksena G, Voet
D, Hussain WM, Ramos AH, Winckler W, Redman MC, Ardlie K, Tewari
AK, Mosquera JM, Rupp N, Wild PJ, Moch H, Morrissey C, Nelson PS,
Kantoff PW, Gabriel SB, Golub TR, Meyerson M, Lander ES, Getz G,
Rubin MA and Garraway LA: Exome sequencing identifies recurrent
SPOP, FOXA1 and MED12 mutations in prostate cancer. Nat Genet.
44:685–689. 2012. View
Article : Google Scholar : PubMed/NCBI
|
|
9
|
Le Gallo M, O’Hara AJ, Rudd ML, Urick ME,
Hansen NF, O’Neil NJ, Price JC, Zhang S, England BM and Godwin AK:
Exome sequencing of serous endometrial tumors identifies recurrent
somatic mutations in chromatin-remodeling and ubiquitin ligase
complex genes. Nat Genet. 44:1310–1315. 2012.PubMed/NCBI
|
|
10
|
Ande SR, Chen J and Maddika S: The
ubiquitin pathway: an emerging drug target in cancer therapy. Eur J
Pharmacol. 625:199–205. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Li C, Ao J, Fu J, Lee DF, Xu J, Lonard D
and O’Malley BW: Tumor-suppressor role for the SPOP ubiquitin
ligase in signal-dependent proteolysis of the oncogenic
co-activator SRC-3/AIB1. Oncogene. 30:4350–4364. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Nakayama KI and Nakayama K: Ubiquitin
ligases: cell-cycle control and cancer. Nat Rev Cancer. 6:369–381.
2006. View
Article : Google Scholar : PubMed/NCBI
|
|
13
|
Bode AM and Dong Z: Post-translational
modification of p53 in tumorigenesis. Nat Rev Cancer. 4:793–805.
2004. View
Article : Google Scholar : PubMed/NCBI
|
|
14
|
Kim MS, Je EM, Oh JE, Yoo NJ and Lee SH:
Mutational and expressional analyses of SPOP, a candidate tumor
suppressor gene, in prostate, gastric and colorectal cancers.
APMIS. 121:626–633. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Dahmane N, Sánchez P, Gitton Y, Palma V,
Sun T, Beyna M, Weiner H and Ruiz i Altaba A: The Sonic
Hedgehog-Gli pathway regulates dorsal brain growth and
tumorigenesis. Development. 128:5201–5212. 2001.PubMed/NCBI
|
|
16
|
Liu J, Ghanim M, Xue L, Brown CD, Iossifov
I, Angeletti C, Hua S, Nègre N, Ludwig M, Stricker T, Al-Ahmadie
HA, Tretiakova M, Camp RL, Perera-Alberto M, Rimm DL, Xu T,
Rzhetsky A and White KP: Analysis of Drosophila segmentation
network identifies a JNK pathway factor overexpressed in kidney
cancer. Science. 323:1218–1222. 2009. View Article : Google Scholar
|
|
17
|
Liu Y, Melin BS, Rajaraman P, Wang Z,
Linet M, Shete S, Amos CI, Lau CC, Scheurer ME, Tsavachidis S,
Armstrong GN, Houlston RS, Hosking FJ, Claus EB, Barnholtz-Sloan J,
Lai R, Il’yasova D, Schildkraut J, Sadetzki S, Johansen C,
Bernstein JL, Olson SH, Jenkins RB, LaChance D, Vick NA, Wrensch M,
Davis F, McCarthy BJ, Andersson U, Thompson PA, Chanock S and Bondy
ML; Gliogene Consortium. Insight in glioma susceptibility through
an analysis of 6p22.3, 12p13.33-12.1, 17q22-23.2 and 18q23 SNP
genotypes in familial and non-familial glioma. Hum Genet.
131:1507–1517. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Louis DN, Ohgaki H, Wiestler OD and
Cavenee WK: WHO Classifcation of Tumours of the Central Nervous
System. 4th edition. IARC Press; Lyon: 2007
|
|
19
|
Van Meir EG, Hadjipanayis CG, Norden AD,
Shu HK, Wen PY and Olson JJ: Exciting new advances in
neuro-oncology: the avenue to a cure for malignant glioma. CA
Cancer J Clin. 60:166–193. 2010.PubMed/NCBI
|
|
20
|
Mrugala MM: Advances and challenges in the
treatment of glioblastoma: a clinician’s perspective. Discov Med.
15:221–230. 2013.PubMed/NCBI
|
|
21
|
Zhang Q, Zhang L, Wang B, Ou CY, Chien CT
and Jiang J: A hedgehog-induced BTB protein modulates hedgehog
signaling by degrading Ci/Gli transcription factor. Dev Cell.
10:719–729. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Zhuang M, Calabrese MF, Liu J, Waddell MB,
Nourse A, Hammel M, Miller DJ, Walden H, Duda DM, Seyedin SN,
Hoggard T, Harper JW, White KP and Schulman BA: Structures of
SPOP-substrate complexes: insights into molecular architectures of
BTB-Cul3 ubiquitin ligases. Mol Cell. 36:39–50. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Bunce MW, Boronenkov IV and Anderson RA:
Coordinated activation of the nuclear ubiquitin ligase Cul3-SPOP by
the generation of phosphatidylinositol 5-phosphate. J Biol Chem.
283:8678–8686. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Chen MH, Wilson CW, Li YJ, Law KK, Lu CS,
Gacayan R, Zhang X, Hui CC and Chuang PT: Cilium-independent
regulation of Gli protein function by Sufu in Hedgehog signaling is
evolutionarily conserved. Genes Dev. 23:1910–1928. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Claiborn KC, Sachdeva MM, Cannon CE, Groff
DN, Singer JD and Stoffers DA: Pcif1 modulates Pdx1 protein
stability and pancreatic β cell function and survival in mice. J
Clin Invest. 120:3713–3721. 2010. View
Article : Google Scholar : PubMed/NCBI
|
|
26
|
Hoeller D and Dikic I: Targeting the
ubiquitin system in cancer therapy. Nature. 458:438–444. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Cohen P and Tcherpakov M: Will the
ubiquitin system furnish as many drug targets as protein kinases.
Cell. 143:686–693. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Torres-Arzayus MI, Font de Mora J, Yuan J,
Vazquez F, Bronson R, Rue M, Sellers WR and Brown M: High tumor
incidence and activation of the PI3K/AKT pathway in transgenic mice
define AIB1 as an oncogene. Cancer Cell. 6:263–274. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Byun B, Tak H and Joe CO: BTB/POZ domain
of speckle-type POZ protein (SPOP) confers proapoptotic function in
HeLa cells. Biofactors. 31:165–169. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Li G, Ci W, Karmakar S, Chen K, Dhar R,
Fan Z, Guo Z, Zhang J, Ke Y, Wang L, Zhuang M, Hu S, Li X, Zhou L,
Li X, Calabrese MF, Watson ER, Prasad SM, Rinker-Schaeffer C,
Eggener SE, Stricker T, Tian Y, Schulman BA, Liu J and White KP:
SPOP promotes tumorigenesis by acting as a key regulatory hub in
kidney cancer. Cancer Cell. 25:455–468. 2014. View Article : Google Scholar : PubMed/NCBI
|