Introduction
Colorectal cancer (CRC) is the third most common
type of cancer with a very poor prognosis predominantly due to its
high rate of tumor invasion and migration, and resistance to
anti-EGFR therapy (1). Although
CRC has been widely studied, the underlying molecular mechanism
remains to be elucidated.
MicroRNAs (miRNAs), a class of small non-coding
RNAs, can negatively regulate gene expression by targeting the
3′-untranslated regions (UTRs) of specific target mRNAs to cause
either mRNA degradation or translational inhibition (2). miRNAs are important in s all
biological activities in mammals (3) and accumulating evidence reveals that
miRNAs are also involved in cancer-associated processes, including
proliferation, migration and apoptosis (4). Therefore, miRNAs are being used as
diagnostic and prognostic biomarkers for certain types of cancer
and also as therapeutic targets. Previously, miR-133b was reported
to be significantly dysregulated in certain types of tumor,
including gastric cancer (5–7),
bladder cancer (8), prostate
cancer (9) and lung cancer
(10,11). In CRC, low expression of miR-133b
was reported to be correlated with poor survival and metastasis
(12). However, the molecular
mechanisms underlying miR-133b in CRC cells remain to be
elucidated.
The EGFR signaling pathway regulates cell
proliferation, differentiation, migration and apoptosis, and widely
participates in tumorigenesis (13–15).
Therefore, EGFR has been used as a therapeutic molecular target for
several metastatic types of cancer, including CRC. Several agents
targeting EGFR, including cetuximab, have been approved and are
currently used for treating patients with CRC (16). Although certain advances in CRC
therapy have been made, patient survival remains poor. This is
predominantly due to the fact that patients with CRC, which are
sensitive to cetuximab almost always develop resistance within
several months. Previous studies have demonstrated that miRNAs are
involved in the EGFR network during cancer oncogenesis (16) and other previous studies revealed
miRNAs to be associated with the therapeutic response in several
cancer cells. For example, miR-221, miR-222, miR-30 and miR-130a
were reported to be involved in gefitinib resistance in non-small
cell lung cancer cells (17–19).
In CRC cells, the cell lines sensitive and resistant to cetuximab
expressed different sets of miRNAs following treatment with
cetuximab (20). This suggests
that certain miRNAs are closely associated with the therapeutic
response in CRC and suggests that they may be important for
overcoming drug resistance in patients with CRC.
The present study determined the involvement of
miR-133b and EGFR in CRC cells. The findings indicated that
miR-133b is downregulated in human CRC tissues and cell lines, and
increased expression levels of miR-133b inhibited the growth and
invasion of CRC cells. In addition, miR-133b directly targeted EGFR
and inhibited its expression in CRC cells. Silencing of EGFR using
TALEN-based knock-out and treatment with cetuximab inhibited CRC
cell growth and invasion in vitro. Additionally,
combinational treatment of miR-133b mimics and cetuximab exhibited
improved inhibitory effects on the growth and invasion of CRC cells
compared with either of treatments alone. Taken together, the
present study characterized the role of the miR-133b/EGFR
interaction in CRC cells and this suggested the combinational
therapy with cetuximab and miR-133b was positive and may be a
potential novel treatment for patients with CRC in the future.
Materials and methods
Tissue samples and cell lines
A total of nine CRC tumor tissue samples and three
normal tissue samples were collected from The Third Xiangya
Hospital of Central South University (Hunan, China). The present
study was approved by the Independent Ethical Committee of Central
South University. The samples were stored at −80°C until used. A
total of five CRC cell lines, HT-29, SW480, SW620, Caco-2 and
HCT-116, (American Type Culture Collection, Rockville, MA, USA)
were used. The cells were grown routinely in H-Dulbecco's modified
Eagle's medium (DMEM; Gibco Life Technologies, Carlsbad, CA, USA),
containing 10% fetal bovine serum (Gibco Life Technologies) and
cultured at 37°C in humidified air of 5% CO2.
Antibody and cetuximab
Rabbit polyclonal anti-EGFR antibody (1:200) was
purchased from Abcam (Hong Kong, China) and Cetuximab (100
μg/ml−1; 2 mg/ml−1) was purchased from
Merck Pharma GmbH (Erbitux, Darmstadt, Germany).
RNA isolation and reverse transcription
quantitative poly- merase chain reaction (RT-qPCR)
The total RNA was extracted from cells and the
patient samples using TRIzol reagent (Invitrogen Life Technologies,
Carlsbad, CA, USA) and ~500 ng extracted total RNA was
reverse-transcribed into cDNA using the Primer Script RT reagent
kit (Takara Bio, Inc., Otsu, Japan). The relative mRNA expression
levels of EGFR were detected using a SYBR-Green qPCR assay (Takara
Bio, Inc.) performed on an ABI Prism 7700 (Applied Biosystems,
Foster City, CA, USA). β-actin was used as a control for
normalization. The specific primer sequences used were: EGFR,
forward 5′-CTTCACACATACTCCTCCTC-3′ and reverse
5′-TCTCCATCACTTATCTCCTT-3′; β-actin, forward
5′-AGGGGCCGGACTCGTCATACT-3′ and reverse
5′-GGCGGCACCACCATGTACCCT-3′. The relative expression levels of
miR-133b were determined using an mirVana™ qRT-PCR miRNA Detection
kit (Ambion, Austin, TX, USA), according to the manufacturer's
instructions. The specific primers for miRNA-133b and U6 were
purchased from GeneCopoeia, Inc. (Rockville, MD, USA). The
expression levels of U6 were used as an endogenous control. All
experiments were performed in triplicate and the relative
expression levels were calculated using the 2−ΔΔCt
method.
Dual luciferase reporter assay
The 3′UTR of the wild-type EGFR (position 50–56 of
EGFR 3′UTR; GGACCAA), and a variant containing mutations
(GCAGCTA) in the putative binding site, were
inserted downstream of the firefly luciferase reporter into the
psiCHECK-2 vector (Promega, Madison, WI, USA). The corresponding
mutant construct was created by mutating the seed regions of the
miR-133b binding sites and was termed 3′-UTR mut EGFR. The primer
sequences used were: 3′UTR EGFR, forward
5′-CCGCTCGAGCCACGGAGGATAGTAT GAG-3′ and reverse
5′-GTTGCGGCCGCGGAAGCCTT GAAGCAGAAC-3′; 3′-UTR mut EGFR, forward
5′-GTTGCG GCCGCCCACGGAGGATAGTATGAG-3′ and reverse
5′-CCGCTCGAGGGAAGCCTTGAAGCAGAAC-3′. The cells were transfected
using lipofectamine 2000 (Invitrogen Life Technologies) and were
co-transfected with reporter constructs with pre-miR-133b,
anti-miR-133b, pre-scramble or anti-scramble. An untreated group
was used as a control. Luciferase activity was detected 48 h after
transfection using a dual-luciferase reporter gene assay kit
(Promega), and the luciferase activities in each group were
determined using an LD400 luminometer (Promega.). Renilla
luciferase activity was normalized to firefly luciferase activity.
Flow cytometery was performed to determine cell apoptosis using an
Annexin V-fluoresecin isothiocyanate (FITC) Apoptosis Detection kit
(BD Biosciences, Frankin Lakes, New Jersey, United State). At 24 h
post-transfection, the cells were harvested and washed twice with
cold PBS. Subsequently, 106 cells were resuspended in
200 μl binding buffer with 10 μl Annexin-FITC and 5
μl propidium iodide, followed by incubation in the dark for
30 min. Finally, 300 μl binding buffer was added and the
cells were assessed using flow cytometric analysis (Moflo XDP;
Beckman Coulter, Brea, CA, USA).
Cell proliferation assay
The cells were seeded into 96-well plates at a
density of 5×103 cells/well and were incubated for 24,
48 and 72 h at 37°C with 5% CO2. The viability of the
cells was assessed using an MTT assay. Briefly, MTT (10 mg/ml) was
added to the cells and incubated for 3 h at 37°C The reaction was
terminated by removal of the supernatant, followed by the addition
of 200 μl dimethyl sulfoxide and gently pipetting. Following
a 2 h incubation, the optical density of each well at 570 nm was
measured with a BioRad microplate reader (Bio-Rad, Hercules, CA,
USA). Each assay was performed in triplicate.
Cell cycle analysis by flow cytometric
analysis
The cells were digested, washed with
phosphate-buffered saline (PBS), and subsequently fixed in 70%
ethanol at 4°C overnight. The fixed cells (1×106) were
washed with PBS and resuspended in staining solution, containing 50
μg/ml propidium iodide, 1 mg/ml RNase A and 0.1% Triton
X-100 in PBS. Following incubation for 30 min at 4°C, the stained
cells were analyzed on a flow cytometer (Beckman Coulter).
Cell invasion assay
Cell invasion was determined using a transwell
assay. Briefly, the cells were suspended in serum-free medium and
aliquots (1×105 cells) of the prepared cell suspension
were added into the upper chamber. The lower chamber was filled
with 1 ml DMEM, containing fetal bovine serum (10%). Following
incubation for 24 h at 37°C, the cells remaining on the upper side
of the membrane were removed using a cotton swab, while the cells,
which had migrated through the membrane were fixed with 75% alcohol
and stained with 1% crystal violet for 20 min. The invasive cells
were subsequently counted and images were captured using an
inverted microscope (Nikon, Tokyo, Japan).
Transcription activator-like effector
nucleases (TALEN)- mediated knockout of the EGFR gene in Caco-2
cells
The present study used TALEN technology to knock out
the EGFR gene in Caco-2 cells. TALENs designed to target the
EGFR gene were purchased from Sidansai Biotechnology
(Shanghai, China). The cells in 24-well plates were transfected
with 400 ng TALEN expression plasmids (EGFR-TALEN) or negative
control (TALEN-NC) using Lipofectamine 2000 (Invitrogen Life
Technologies), according to the manufacturer's instructions.
Western blotting was performed to determine the protein expression
levels of EGFR and confirm the efficiency of the TALEN mediated
knockout.
Results
miR-133b is decreased in CRC tumor
samples and cell lines
To determine the potential clinical and pathological
implications of an altered expression of miR-133b in CRC tumor
samples, the present study investigated the expression levels of
miR-133b in nine CRC tumor samples and three normal tissue samples
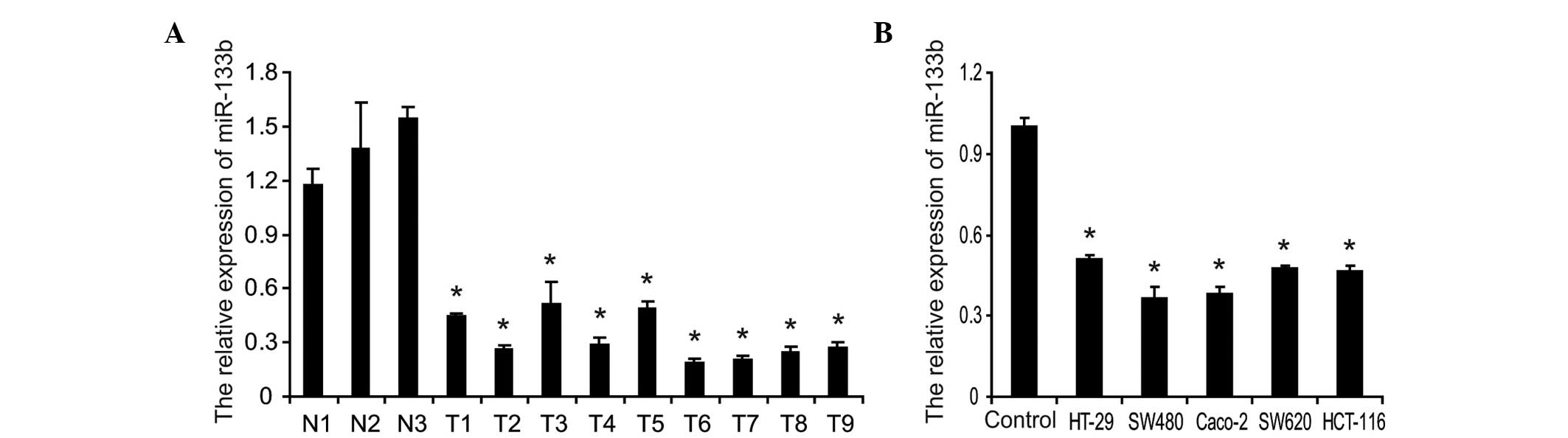
using RT-qPCR. As shown in Fig.
1A, the expression levels of miR-133b were significantly lower
in CRC tumor samples compared with the normal tissue samples.
Furthermore, the abundance of miR-133b in five CRC cell lines was
demonstrated. Compared with normal colonic mucosa epithelial cells
isolated and pooled from three adjacent non-cancerous tissue
samples, the expression levels of miR-133b in the five CRC cell
lines were significantly reduced to different extents, markedly
lower in the SW480 and Caco-2 cells (Fig. 1B). These results suggested that the
expression levels of miR-133b were significantly decreased in human
CRC specimens and cell lines, which suggested that it may be
involved in the pathogenesis of CRC.
miR-133b targets and represses the
expression of EGFR in CRC cells in vitro
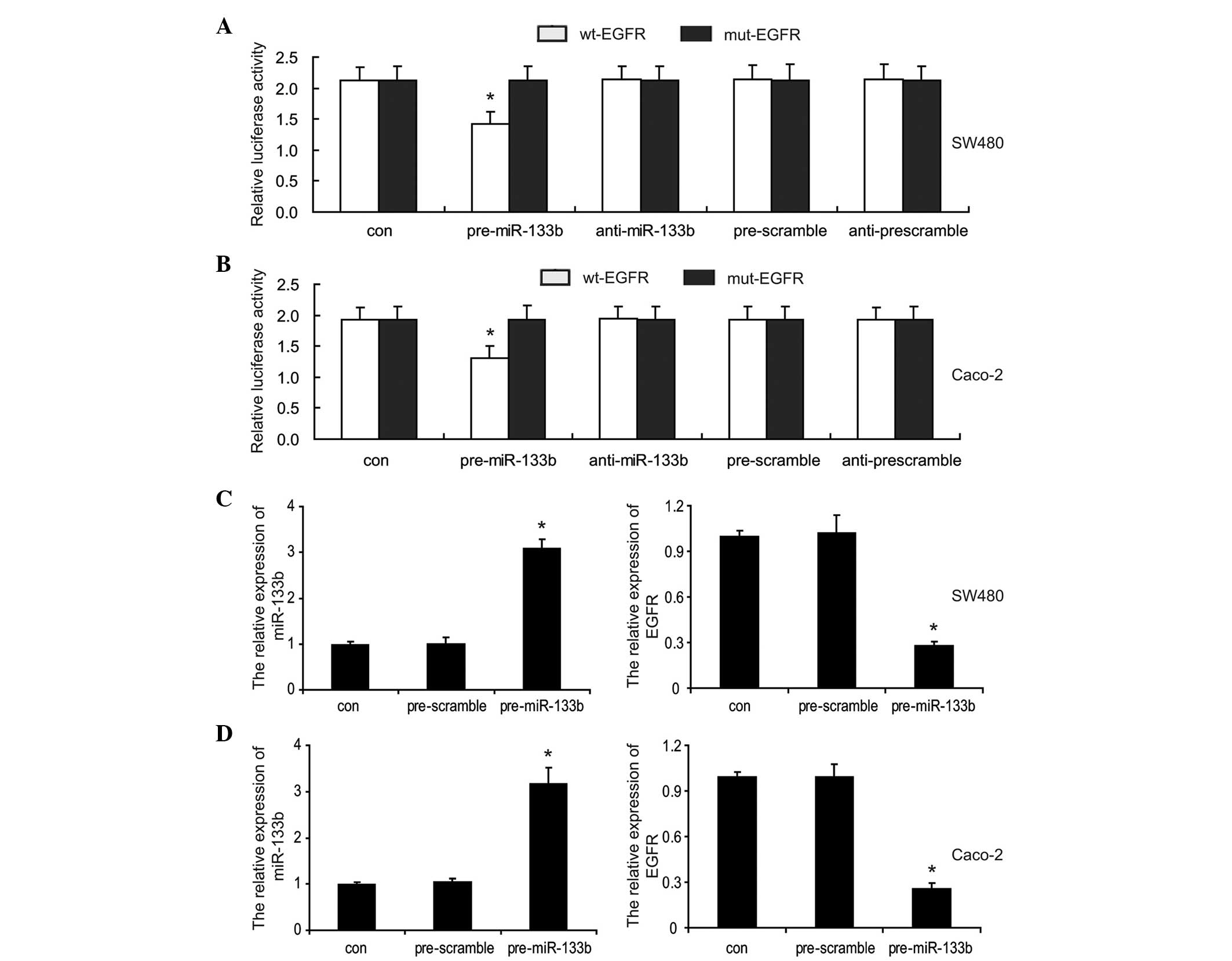
To determine whether miR-133b directly targets EGFR
in CRC cells, a luciferase assay was performed. A region of 501 bp
length of the wild-type 3′-UTR of EGFR, containing the predicted
miR-133b target sites, was amplified and inserted into the
resulting amplicon downstream of a lucife rase reporter gene
(termed wt-EGFR). A mutant version of the 3′-UTR of EGFR lacking
the miR-133b binding sites (named mut-EGFR), was also constructed.
The SW480 and Caco-2 cells were co-transfected with wt-EGFR or
mut-EGFR vectors and either the miR-133b mimics (pre-miR-133b),
miR-133b inhibitor (anti-miR-133b) or their respective scrambled
controls (pre-scramble and anti-scramble). The results showed that
miR-133b significantly suppressed the luciferase activity of the
reporter gene, containing the 3′UTR of EGFR compared with the
controls, however, this was significantly rescued when the miR-133b
binding sites were absent in the SW480 and Caco-2 cells (Fig. 2A and B). Furthermore, the
inhibitory effect of miR-133b on EGFR expression was assessed.
RT-qPCR analysis demonstrated that an sincreased expression of
miR-133b significantly decreased the mRNA expression levels of EGFR
compared with the cells transfected with the control (Fig. 2C and D). Taken together, these
results confirmed that miR-133b directly targeted EGFR and
repressed its expression levels in CRC cells.
miR-133b inhibits the growth and invasion
of CRC cells
To investigate the roles of miR-133b in the CRC
cells, the expression of miR-133b was restored in SW480 and Caco-2
cells. This resulted in a lower expression of miR-133b in the
selected CRC cell lines, by liposomal delivery. The effect of
miR-133b on cell growth and invasion was investigated. An MTT assay
demonstrated that the overexpression of miR-133b decreased the cell
viability in SW480 and Caco-2 cells (Fig. 3A and B). It was also revealed that
miR-133b marginally increased the percentage of G1 phase cells in
SW480 and Caco-2 cells (Fig. 3C and
D), as determined by flow cytometric analysis. A Transwell
assay indicated the forced expression of miR-133b significantly
inhibited the cell invasion ability compared with the control group
in SW480 and Caco-2 cells (Fig. 3E and
F). These results suggested that miR-133b, which serves as a
tumor suppressor, is important in the control of growth and
invasion of CRC cells in vitro.
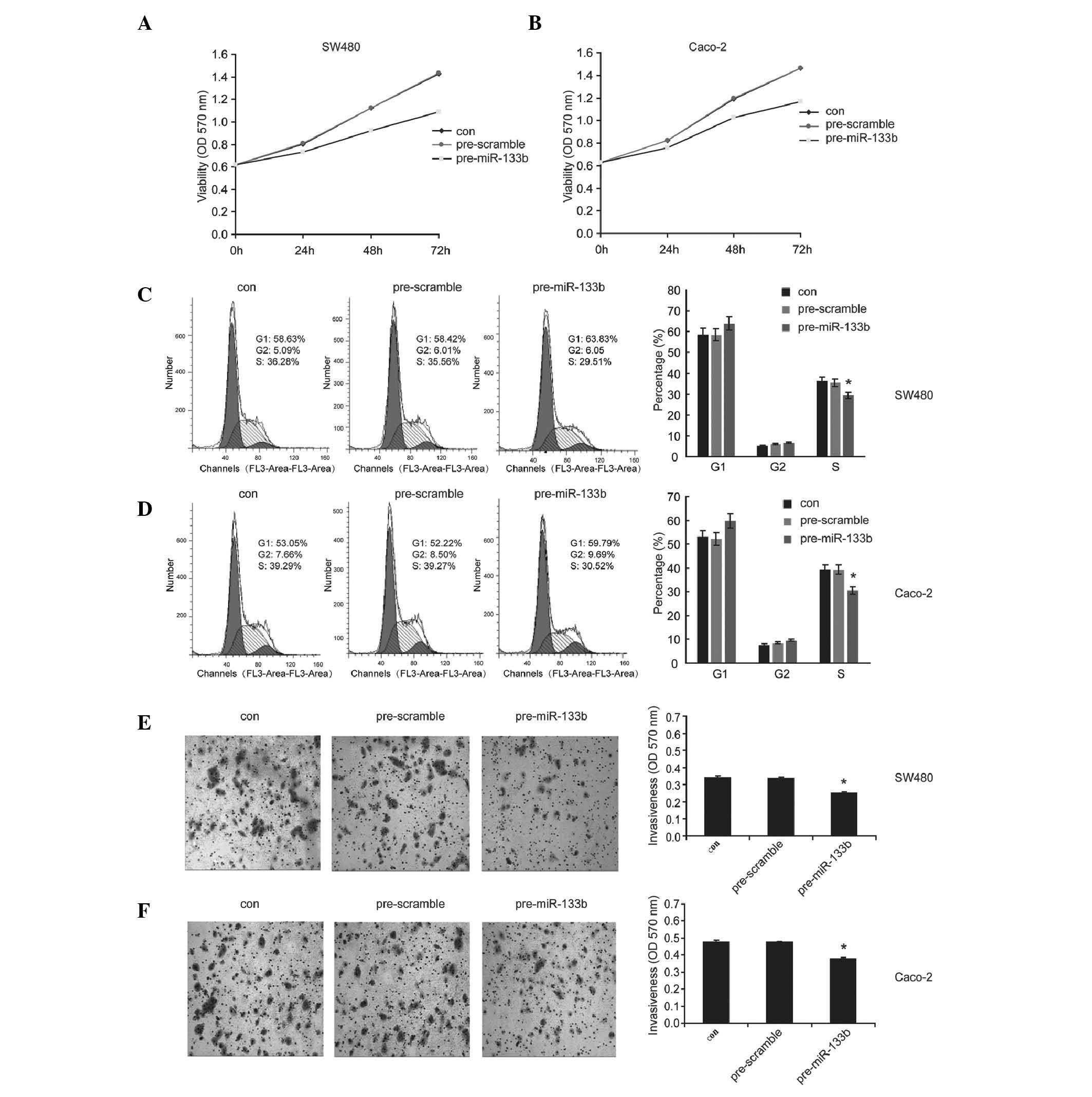 | Figure 3miR-133b inhibits the growth and
invasion of colorectal cancer cells. Following transfection with
pre-miR-133b, pre-scramble or control in SW480 and Caco-2 cells,
tha (A and B) cell viability, (C and D) the percentage of cells in
the G1, G2 and S phases and (E and F) the cell invasiveness were
determined using MTT, flow cytometry and transwell assays,
respectively (magnification, ×100). The data are presented as the
mean ± standard deviation (*P<0.05, compared with the
control). miR, microRNA; con, control; OD, optical density. |
Silencing of EGFR by TALEN inhibits the
growth and invasion of Caco-2 cells
Based on the above finding that miR-133b inhibited
the cell growth and invasion, at least in part, by downregulating
the expression levels of EGFR, the present study hypothesized that
inhibiting the expression of EGFR by other approaches would also
have similar inhibitory effects. EGFR was knocked out using TALEN
technology, a highly effective approach for the targeted knockout
of genes in mammalian cells. Western blotting was performed to
confirm the TALEN-mediated knock-out efficiency in Caco-2 cells. As
shown in Fig. 4A, the EGFR gene
was silenced effectively in the EGFR-TALEN vector transfected
cells. As expected, silencing of EGFR by TALEN decreased the cell
viability (Fig. 4B), increased the
percentage of cells in the G1 phase (Fig. 4C) and inhibited the invasiveness
(Fig. 4D) of Caco-2 cells. This
suggested that EGFR silencing may be a common effective strategy to
inhibit the growth and invasion in CRC cells.
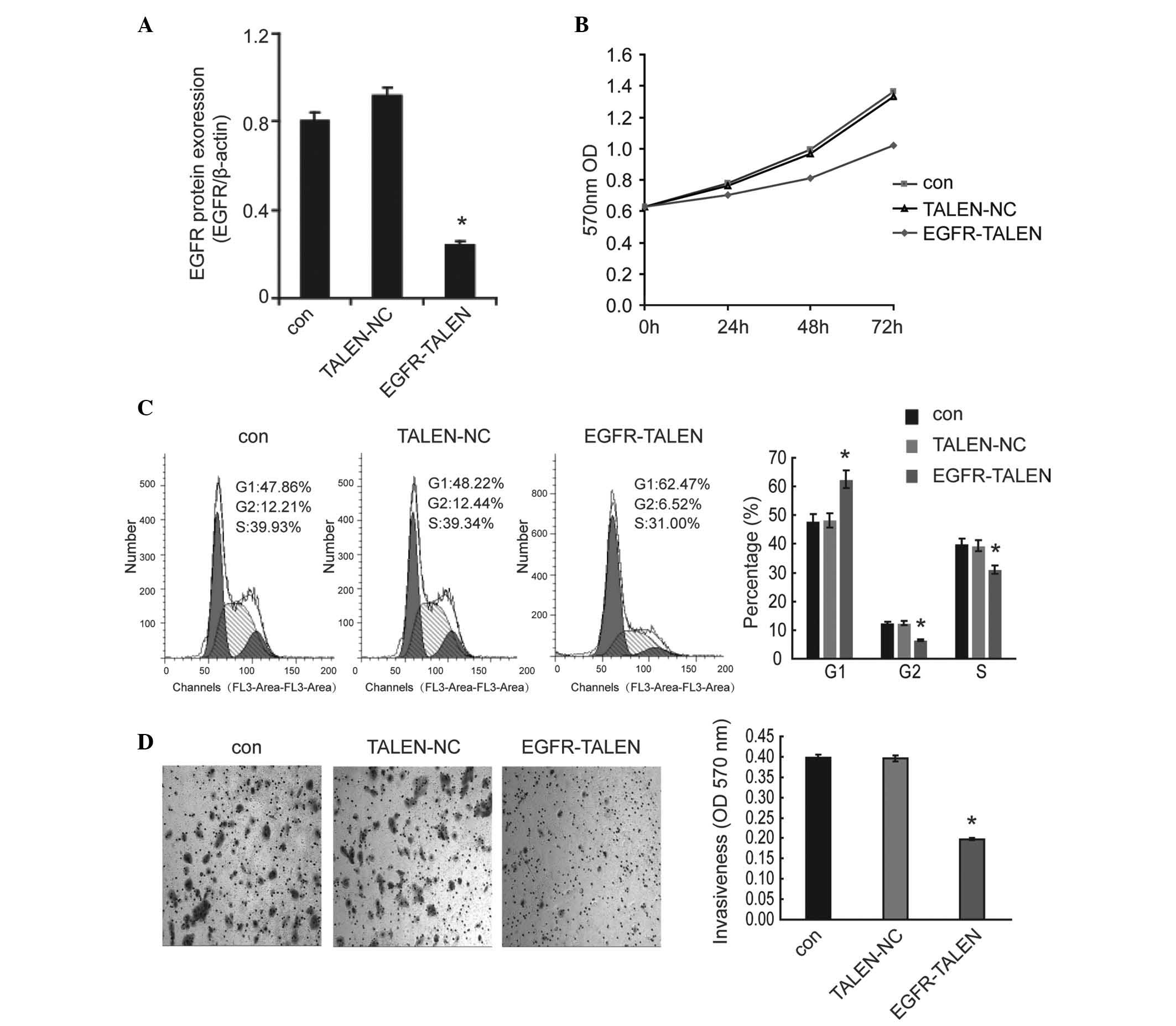 | Figure 4Silencing of EGFR by TALEN inhibits
the growth and invasion of Caco-2 cells. (A) Following knock-out
using TALEN technology in Caco-2 cells, the protein expression
levels of EGFR were determined by western blotting to confirm the
TALEN-mediated knock-out efficiency. (B) The cell viability, (C)
the percentage of cells in the G1, G2 and S phases and (D) the cell
invasiveness were determined by MTT assay, flow cytometric analysis
and transwell assay, respectively (magnification, ×100). The data
are shown as the mean ± standard deviation (*P<0.05,
compared with the control). TALEN, transcription activator-like
effector nucleases; con, control; OD, optical density; NC, negative
control; EGFR, epidermal growth factor receptor. |
Combination of miR-133b and cetuximab
increases the inhibitory effects on the growth and invasion of CRC
cells
Cetuximab is a chimeric monoclonal antibody, which
binds to EGFR and competitively inhibits ligand binding to suppress
tumor proliferation. Cetuximab has been approved and is used for
the treatment of patients with CRC. The present study aimed to
determine whether the combination of miR-133b and cetuximab
provided an improved antitumor effect in CRC cells. To carry this
out, the SW480 and Caco-2 cells were treated with cetuximab
following transfection with the miR-133b mimics or scramble
control. Notably, the combination treatment of miR-133b and
cetuximab exhibited an increased inhibitory effect on the cell
viability (Fig. 5A and B) and
invasion (Fig. 5E and F) of the
SW480 and Caco-2 cells compared with treatment with either miR-133b
or cetuximab alone. It was also demonstrated that combinational
treatment had a marginally increased inhibitory effect on cell
cycle progression into S phase, as determined by flow cytometric
analysis (Fig. 5C and D). These
data suggested that this was a promising option of a combinational
therapy for patients with CRC.
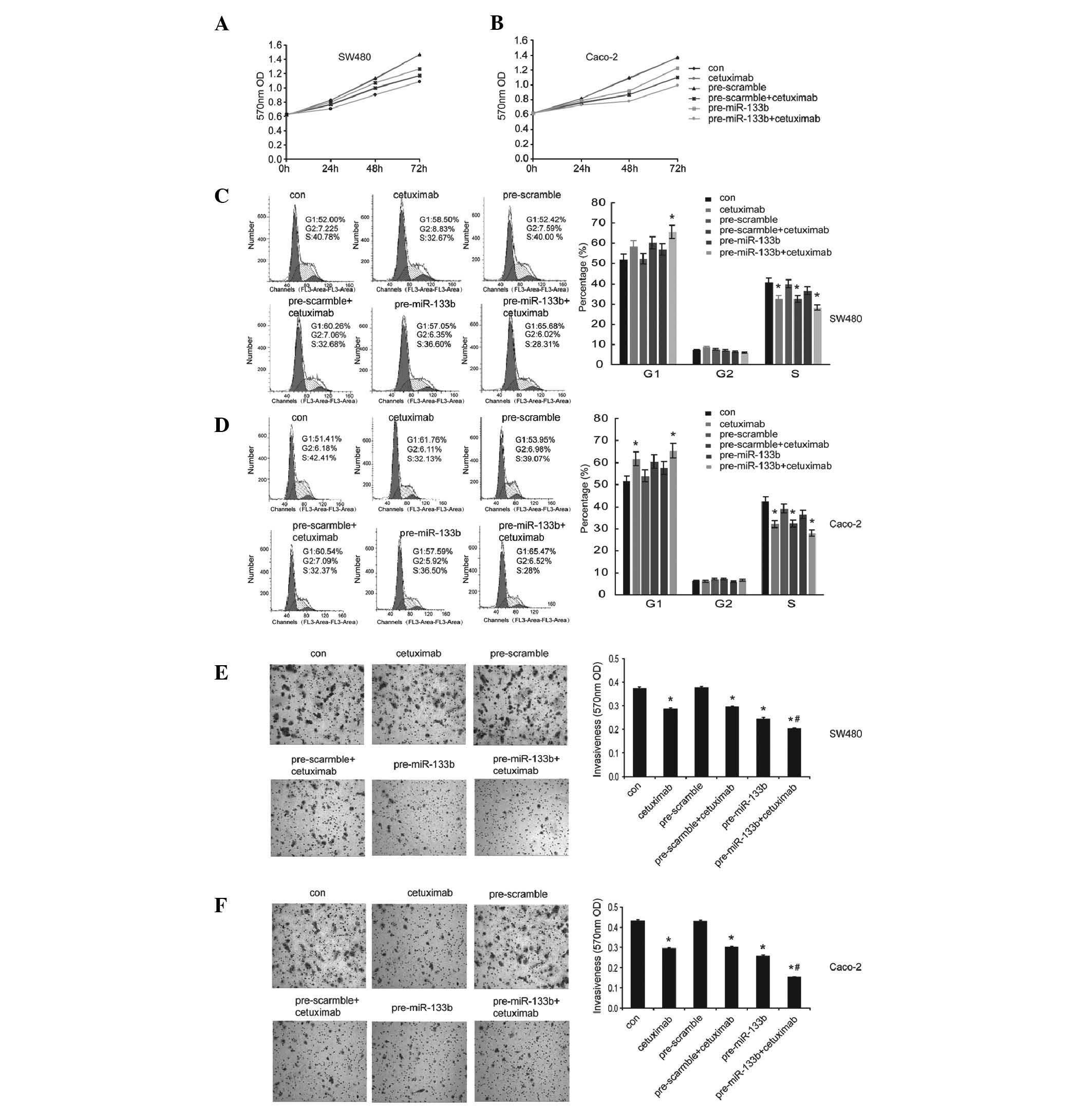 | Figure 5Combination of miR-133b and cetuximab
increases the inhibitory effects on the growth and invasion of
colorectal cancer cells. SW480 and Caco-2 cells were transfected
with pre-miR-133b or pre-scramble, followed by treatment with
cetuximab (10 μM) for 24 h. An untreated group were used as
a control. (A and B) The cell viability, (C and D) the percentage
of cells in the G1, G2 and S phases and (E and F) the cell
invasiveness were determined by MTT, flow cytometric analysis and
transwell assay, respectively (magnification, ×100). The data are
shown as the mean ± standard deviation (*P<0.05,
compared with the control; #P<0.05, compared with the
cetuximab treated groups or the pre-miR-133b transfected groups).
miR, microRNA; con, control; OD, optical density. |
Discussion
The present study demonstrated that miR-133b is
down-regulated in CRC tissue samples and cell lines compared with
normal tissue samples and cells. When the expression levels of
miR-133b were restored, the growth and invasion of CRC cells was
inhibited, suggesting that miR-133b may function as a tumor
suppressor in the development of CRC. It was also revealed that
EGFR is a direct functional target of miR-133b in CRC cells and
restored expression of miR-133b repressed the expression of EGFR.
Therefore, the present study speculated that low expression levels
of miR-133b contributed, at least in part, to EGFR-mediated cell
growth and migration in CRC cells. The loss of EGFR by TALEN-based
knockout had similar inhibitory effects on the growth and invasion
of Caco-2 cells in vitro, which suggested that EGFR
silencing may be a common effective strategy to inhibit the growth
and invasion in CRC cells. Additionally, combinational treatment
with miR-133b mimics and the EGFR monoclonal antibody, cetuximab,
demonstrated an increased inhibitory effect on the cell growth and
invasion of SW480 and Caco-2 cells compared with either miR-133b or
cetuximab alone. These data suggested that this combinational
therapy may be a promising choice for treating patients with CRC in
the future.
miRNA alterations are involved in the initiation and
progression of human cancer. miRNA acts as a tumor suppressor or
promoter depending on its target genes (21). Therefore, the miRNA and its targets
can shed light on the molecular mechanism underlying cancer
progression and provide useful potential therapeutic targets for
the clinical treatment of certain types of cancer. miR-133b, a
muscle-specific miRNA (22), has
previously been reported to be involved in other types of tumor
(4–9). The expression of miR-133b is
frequently decreased and acts as a tumor suppressor in gastric
cancer (5,6), osteosarcoma (23) and esophageal squamous cell
carcinoma (24), by negatively
regulating the expression of FGFR1, MET and FSCN1, respectively.
However, miRNA-133b is a key promoter of the development of
cervical carcinoma through the activation of the ERK and AKT1
pathways (25). The present study
confirmed that miR-133b inhibited the cell growth and invasion in
CRC cells by targeting EGFR, consistent with previous findings that
miR-133b inhibits cell growth and invasion in prostate cancer
(8), bladder cancer (7) and non-small-cell lung cancer
(9) by downregulating EGFR. Taken
together, the present study suggested that miR-133b may broadly
inhibit EGFR-induced tumorigenesis.
Cetuximab is an EGFR monoclonal antibody and is
currently used in the therapy of patients with metastatic CRC.
However, several patients treated with cetuximab fail to respond
and those who do initially respond often acquire resistance. This
may be a result of the extensive involvement of miRNAs in the
regulation of the EGFR signaling pathway and certain miRNAs,
including miR-133b, close involvement in therapeutic response of
CRC cells treated with cetuximab. CRC SW480 and Caco-2 cells were
treated with cetuximab and miR-133b mimics to investigate the
efficiency of the combinational therapy. Notably, the growth and
invasion of SW480 cells sensitive to cetuximab and Caco-2 cells
resistant to cetuximab, were effectively inhibited by the
combinational treatment compared with either of the treatments
alone. Taken together, these findings suggested that anti-EGFR
therapy with cetuximab in combination with miR-133b or other
miRNAs, which inhibit EGFR, may provide therapeutic benefit for
patients with metastatic CRC and other EGFR driven types of
tumor.
In conclusion, results of the present study
confirmed the miR-133b/EGFR association in CRC cells, in which the
loss of miR-133b may result in the increased expression levels of
EGFR. This increased expression may endow CRC cells with the
ability of improved growth and invasion capacity. The restoration
of miR-133b and combinational treatment with cetuximab may be a
promising therapeutic strategy for the treatment of metastatic CRC
in the future.
Acknowledgments
This study was supported by the Key Program for
International Cooperation Projects of Hunan Province (no.
2011WK2011) and the Natural Science Foundation of China (no.
81172298).
References
|
1
|
Asbagh LA, Vazquez I, Vecchione L, et al:
The tyrosine phosphatase PTPRO sensitizes colon cancer cells to
anti-EGFR therapy through activation of SRC-mediated EGFR
signaling. Oncotarget. 5:10070–10083. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Lewis BP, Burge CB and Bartel DP:
Conserved seed pairing, often flanked by adenosines, indicates that
thousands of human genes are microRNA targets. Cell. 120:15–20.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Kato M and Slack FJ: microRNAs: small
molecules with big roles-C. elegans to human cancer. Biol Cell.
100:71–81. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Jansson MD and Lund AH: MicroRNA and
cancer. Mol Oncol. 6:590–610. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Qiu T, Zhou X, Wang J, et al: MiR-145,
miR-133a and miR-133b inhibit proliferation, migration, invasion
and cell cycle progression via targeting transcription factor Sp1
in gastric cancer. FEBS Lett. 588:1168–1177. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Wen D, Li S, Ji F, et al: miR-133b acts as
a tumor suppressor and negatively regulates FGFR1 in gastric
cancer. Tumour Biol. 34:793–803. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Zhao Y, Huang J, Zhang L, et al: MiR-133b
is frequently decreased in gastric cancer and its overexpression
reduces the metastatic potential of gastric cancer cells. BMC
Cancer. 14:342014. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Zhou Y, Wu D, Tao J, Qu P, Zhou Z and Hou
J: MicroRNA-133 inhibits cell proliferation, migration and invasion
by targeting epidermal growth factor receptor and its downstream
effector proteins in bladder cancer. Scand J Urol. 47:423–432.
2013. View Article : Google Scholar
|
|
9
|
Tao J, Wu D, Xu B, et al: microRNA-133
inhibits cell proliferation, migration and invasion in prostate
cancer cells by targeting the epidermal growth factor receptor.
Oncol Rep. 27:1967–1975. 2012.PubMed/NCBI
|
|
10
|
Liu L, Shao X, Gao W, et al: MicroRNA-133b
inhibits the growth of non-small-cell lung cancer by targeting the
epidermal growth factor receptor. FEBS J. 279:3800–3812. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Wu J, Yang T, Li X, et al: Alteration of
serum miR-206 and miR-133b is associated with lung carcinogenesis
induced by 4-(methylnitrosamino)-1-(3-pyridyl)-1-butanone. Toxicol
Appl Pharmacol. 267:238–246. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Akcakaya P, Ekelund S, Kolosenko I, et al:
miR-185 and miR-133b deregulation is associated with overall
survival and metastasis in colorectal cancer. Int J Oncol.
39:311–318. 2011.PubMed/NCBI
|
|
13
|
Davis NM, Sokolosky M, Stadelman K, et al:
Deregulation of the EGFR/PI3K/PTEN/Akt/mTORC1 pathway in breast
cancer: possibilities for therapeutic intervention. Oncotarget.
5:4603–4650. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Centuori SM and Martinez JD: Differential
regulation of EGFR-MAPK signaling by deoxycholic acid (DCA) and
ursodeoxycholic acid (UDCA) in colon cancer. Dig Dis Sci.
59:2367–2380. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Wang F, Xiao W, Sun J, Han D and Zhu Y:
MiRNA-181c inhibits EGFR-signaling-dependent MMP9 activation via
suppressing Akt phosphorylation in glioblastoma. Tumour Biol.
35:8653–8658. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Mlcochova J, Faltejskova P, Nemecek R,
Svoboda M and Slaby O: MicroRNAs targeting EGFR signalling pathway
in colorectal cancer. J Cancer Res Clin Oncol. 139:1615–1624. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Garofalo M, Romano G, Di Leva G, et al:
EGFR and MET receptor tyrosine kinase-altered microRNA expression
induces tumorigenesis and gefitinib resistance in lung cancers. Nat
Med. 18:74–82. 2011.PubMed/NCBI
|
|
18
|
Gu YF, Zhang H, Su D, et al: miR-30b and
miR-30c expression predicted response to tyrosine kinase inhibitors
as first line treatment in non-small cell lung cancer. Chin Med J
(Engl). 126:4435–4439. 2013.
|
|
19
|
Zhou YM, Liu J and Sun W: MiR-130a
overcomes gefitinib resistance by targeting met in non-small cell
lung cancer cell lines. Asian Pac J Cancer Prev. 15:1391–1396.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Ragusa M, Majorana A, Statello L, et al:
Specific alterations of microRNA transcriptome and global network
structure in colorectal carcinoma after cetuximab treatment. Mol
Cancer Ther. 9:3396–3409. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Calin GA and Croce CM: MicroRNA signatures
in human cancers. Nat Rev Cancer. 6:857–866. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Nielsen S, Scheele C, Yfanti C, et al:
Muscle specific microRNAs are regulated by endurance exercise in
human skeletal muscle. J Physiol. 588:4029–4037. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Novello C, Pazzaglia L, Cingolani C, et
al: miRNA expression profile in human osteosarcoma: role of miR-1
and miR-133b in proliferation and cell cycle control. Int J Oncol.
42:667–675. 2013.
|
|
24
|
Kano M, Seki N, Kikkawa N, et al: miR-145,
miR-133a and miR-133b: Tumor-suppressive miRNAs target FSCN1 in
esophageal squamous cell carcinoma. Int J Cancer. 127:2804–2814.
2010. View Article : Google Scholar
|
|
25
|
Qin W, Dong P, Ma C, et al: MicroRNA-133b
is a key promoter of cervical carcinoma development through the
activation of the ERK and AKT1 pathways. Oncogene. 31:4067–4075.
2012. View Article : Google Scholar
|