Introduction
Although theoretical concepts and technological
approaches have made significant progress, the molecular basis of
carcinogenesis and progression of various types of cancer remains
to be understood. This deficit in knowledge hinders the development
of effective therapies and progress in the treatment of cancers
remains slow. As a result, the curability of cancers is still poor
(1).
To promote the discovery of oncogenic pathways,
investigators have assessed biological networks, such as
transcriptional regulatory networks (TRNs). TRNs (also known as
gene regulatory networks) can offer the possibility to improve the
understanding of the topology and function of gene regulation of
the cellular responses to environmental changes at a system level.
One important local property of biological networks is the ‘network
motifs’, first described by Milo et al (2). They are patterns of interconnections
occurring in complex networks and may reflect a framework in which
particular functions are achieved efficiently. Much experimental
study has been devoted to understanding network motifs in TRNs, as
they define the core of the regulatory machinery of cellular life
and are largely responsible for information processing and decision
making (3).
The transcription network is a collection of DNA
segments in a cell, which interacts with each other indirectly
(through their RNA and protein expression products) and with other
substances in the cell, thereby governing the expression levels of
mRNA and proteins. In the network, a gene serves as the source of a
direct regulatory edge by producing an RNA or protein molecule that
functions as a transcriptional activator or inhibitor of the target
gene. The network consists of network motifs, such as feed-forward
loops (FFLs), feed-back loops (FBLs) and single-input modules. FFLs
have been shown to be one of the most important and promising
classes of transcriptional network motifs (2,4,5).
The FFL, a three-node motif pattern, is composed of
two input elements, one of which regulates the other, both jointly
regulating a target gene. Each of the three interactions in the FFL
can be either activating or repressing (6,7).
As the research is being driven by its promising
prospects, the importance of post-transcriptional processes have
become more evident than previously expected in the regulation of
gene expression. Among the various mechanisms of TRNs,
transcription factors (TFs) and a class of small RNAs, known as
microRNAs (miRNAs or miRs), are frequently observed in numerous TRN
motifs, joining transcriptional and post-transcriptional regulatory
interactions together, so as to play their prominent roles in
regulation. Additionally, as the research regarding cancers
expands, FFLs composed of a TF, an miRNA and their equivalent
target gene are becoming apparent and the number of studies with
such FFLs of different components reported is continuously
increasing (8–12).
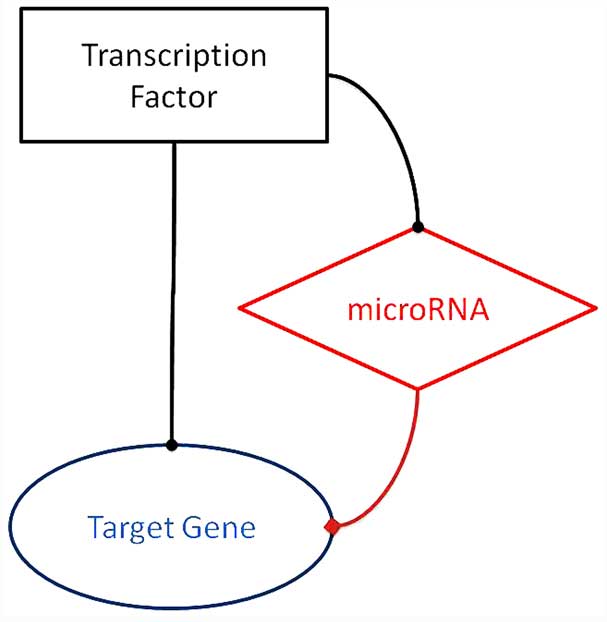
All the possible FFLs involving miRNA, TF and target
gene are shown in Fig. 1. The
TF-miRNA-target gene FFLs, as the network motif of typical TRNs
that we will discuss in the present review, are depicted in
Fig. 1.
In molecular biology and genetics, a TF (sometimes
known as a sequence-specific DNA-binding factor) is a protein that
binds to specific DNA sequences, thereby controlling the flow (or
transcription) of genetic information from DNA to messenger RNA
(mRNA) (13). Characterized by
containing one or more DNA-binding domains, which attach to
specific sequences of DNA adjacent to the genes that they regulate,
TFs are essential for the regulation of gene expression and are
found in all living organisms. TFs can read and interpret the
genetic ‘blueprint’ in the DNA. They bind to the DNA and help
initiate a program of increased or decreased gene transcription. As
such, they are vital for numerous important cellular processes
(14–16).
The most well-known TF, p53, regulates genes, such
as p21, cdc25c, bax and puma, which are
elucidated to have an indispensable role in cell cycle arrest,
apoptosis or cell senescence, and all these can eliminate or
reverse the presence of progenitor cancer cells in the body
(17). p53 function appears to be
crucial in tumors: i) p53 limits the first steps of transformation
by preventing the proliferation of cells with damaged genomes or
dysregulated growth. ii) p53 may act as an emergency brake at later
stages of tumor progression, by preventing cells from accumulating
multiple mutations and developing an invasive phenotype. Taken
together, these mechanisms explain the effects of TP53
mutation in numerous types of human cancer, detectable sometimes as
an early event in precursor lesions or as a later event at the
transition from in situ to invasive cancer (18). Cancer associated miRNAs, such as
miR-34, miR-221 and miR-15/16, harbor p53 consensus binding sites
and are already confirmed to be regulated by p53 and thus control
downstream genes, including Bcl2, p27, E2F3
and CDK6, to carry out anti-tumorigenesis function (19). In 2008, Brosh et al (20) reported an FFL constructed by p53, E2F
and miR-106b/93/25 polycistron, of which the target gene E2F
was also a TF.
Numerous other TFs, such as WT1, TAL1/SCL and Myc,
have also been proven to be associated with their regulating miRNA
or target gene in cancer (21,22).
Increasing attention has focused on miRNAs as they
have been indicated in various types of human cancer. miRNAs are
small, evolutionarily conserved, endogenous non-coding RNAs of
18–25 nucleotides (nts) in length that have an important function
in gene regulation by pairing to the mRNAs of protein-coding genes
to direct their post-transcriptional repression. Their silencing
effects are exerted by cleavage of their target mRNAs and by
inhibition of their translation. Each miRNA can target a large
number of genes (mRNAs) and each mRNA can be targeted by several
miRNAs. It is generally observed that miRNAs only have a minor
influence on the protein levels of their targets; however, miRNAs
can have a profound influence on cell-fate determination. It can
even change a phenotype by modulating a single miRNA. miRNAs are
now known to repress thousands of target genes and coordinate
normal processes, such as developmental timing, pattern formation,
embryogenesis, differentiation, organogenesis, growth control and
cell death. This discovery established a new paradigm of gene
regulation (23–29). The alteration of miRNAs also
contributes to a range of human pathologies, including cancer.
Different associations between miRs and cancer have accumulated
since the first evidence of an oncogene, KRAS, being targeted by an
miRNA, the let-7 family, was reported in 2005 (30).
Beyond the impact of somatic, genetic and epigenetic
lesions, the altered expression of miRNAs in cancer can arise
through the aberrant activity of TFs that control their expression.
Of note, the same TFs are often targets of miRNA-mediated
repression, which gives rise to complex regulatory circuits
(24).
Following this, as the understanding of miRNAs
improved due to high-throughput miRNA expression profiling,
bioinformatic prediction and other advanced technologies, their
functions as regulators in signaling pathways and transcription
networks have been revealed step by step. The maps of the networks
have been completed gradually.
Directed by the theoretical discovery, research on
signal flow inside the cell has increased. Researchers have carried
out various experiments on the typical transcription network
motifs. Subsequently, more hypotheses and their respective
confirmative experiments were conducted and the TF-miRNA-target
gene FFL motif theory was proven repeatedly, particularly in
various types of human cancer (31–34).
Reported TF-miRNA-target gene FFLs in
cancer
One of the best-characterized oncogenic miRNAs is
miR-17-92, a polycistronic miRNA cluster also designated by He
et al (35) as oncomiR-1 in
2005. This was the first time that the concept of an ‘oncogenic
miRNA (oncomiR)’ became apparent. As more miRNAs have been
identified to act as oncogenes, tumor suppressors and important
modulators in cellular pathways have been divided into two classes:
Increased activity of oncomiRs leads to inhibition of
tumor-suppressor genes, facilitating cell proliferation and tumor
progression. Decreased activity of tumor-suppressor miRNAs (tsmiRs)
leads to increased oncogene translation, contributing to tumor
formation (36,37).
Since the same TF can act as an activator or a
repressor under different conditions, its directly-regulating
downstream miRNA can be either an oncomiR or a tsmiR regardless of
whether the TF is an oncogene or a tumor repressor. For instance,
Myc, the c-Myc oncogenic TF, is known to directly upregulate a
pro-tumorigenic group of miRNAs known as the miR-17-92 cluster,
however, the predominant consequence of Myc activation is
widespread repression of miRNA expression (38). The involved miRNAs, including miR-26a,
miR-150 and miR-195/miR-497, whose tumor-suppressing properties are
to be confirmed (39–41).
TF-miRNA-target gene FFL circuits with
its TF as an oncogene in cancer
In 2005, O'Donnell et al (42) reported that the loop consisting of
c-Myc, miR-17-5p and miR-20a cluster and E2F1 modulates cellular
proliferation in P493-6 cells. c-Myc simultaneously activates E2F1
transcription and limits its translation by upregulating miR-17-5p
and miR-20a, allowing a tightly controlled proliferative
signal.
Burk et al (31)
reported an FFL in 2008. In the FFL, zinc-finger E-box binding
homeobox 1 (ZEB1) directly suppressed the transcription of
miRNA-200 family members, miR-141 and miR-200c, which suppress
target gene transforming growth factor-β2 (TGFβ2) and strongly
activate epithelial differentiation so as to repress the
epithelial-mesenchymal transition (EMT) in pancreatic, colorectal
and breast cancer cells. As less-suppressed TGFβ2 in return
upregulates ZEB1, ZEB1 triggers this miRNA-mediated FFL that
stabilizes EMT and promotes invasion. TF ZEB1 itself is a crucial
inducer of EMT in various human tumors and was shown to promote
invasion and metastasis of tumor cells (43).
Following this, KRAS was proved to be a target for
several miRNAs and KRAS activation indicated the repression
of several miRNAs. For example, in pancreatic cancer with mutant
KRAS, RAS-responsive element-binding protein 1 (RREB1)
represses the miR-143 and miR-145 promoter and at the same time
KRAS and RREB1 are targets of miR-143 and miR-145, revealing a
feed-forward regulatory circuit that increases the effect of RAS
signaling (44).
El Baroudi et al (45) made a summary of simple and mixed FFLs
involving c-Myc in 2011. There are various complex circuits with
c-Myc involved, such as the MYC/PTEN/miR-106b, miR-93, miR-25,
miR-19a, miR-22, miR-26a, miR-193b and miR-23b circuit, acting as a
noise-buffering circuit to guarantee a steady level of the PTEN
protein as a tumor-suppressor gene. The MYC/retinoblastoma 1
(RB1)/miR-106a, miR-106b and miR-17 circuit has a critical role in
the pathogenesis of solid cancer by repressing the transcription
and translation of tumor-suppressor gene RB1. The
MYC/vascular endothelial growth factor (VEGF)/miR-106b, miR-106a,
miR-93, miR-34a, miR-20a, miR-17, miR-16 and miR-15a circuit, can
be classified as a coherent or incoherent loop, depending on the
different functional roles of VEGF ranging from cell migration to
apoptosis.
In the year 2013, Polioudakis et al (46) confirmed that miR-22, activated by the
TF Myc when quiescent cells enter proliferation, could inhibit the
Myc transcriptional repressor MXD4, mediating an FFL to elevate Myc
expression levels in HeLa cells and human foreskin fibroblasts.
Also in 2013, Zhao et al (41) published a study that identified a
MYC-miRNAs-EZH2 FFL linking overexpression of MYC, EZH2 and miR-26a
repression in aggressive B cell lymphomas.
TF-miRNA-target gene FFL circuits with
its TF as a tumor repressor in cancer
He et al (47)
reported that the loop formed by p53 and miR-34a-c promotes cell
cycle arrest and inhibits inappropriate cell proliferation in 2007.
They proved direct regulation of the association between p53 and
miR-34a, miR-34a and its target genes CDK4 and MET;
however, in 2011, Hwang et al (32) established the exact p53-regulated FFL:
Regulation of cancer-invasion-promoting gene MET by
wild-type p53 consists of miR-34-dependent and -independent
mechanisms. p53 activates miR-34, which represses MET and
p53 can repress MET itself.
Untypical or uncertain reported FFLs
are associated with cancer
In 2008, Lin et al (48) showed that TF c-Myc directly activates
transcription of the 3 subunits of eIF4F (eIF4E, eIF4AI and
eIF4GI), which is thought to be the rate-limiting phase of
translation. Increased eIF4F levels result in stimulation of c-Myc
mRNA translation specifically. This FFL involving c-Myc and eIF4F
that serves to link transcription and translation could contribute
to the effects of c-Myc on cell proliferation and neoplastic
growth. The following year, the investigators published another
study confirming the FFL association and highlighted that the
regulators of the transcription and translation that affect Myc
function (such as Mad1 or antisense approaches) or eIF4F activity
(such as mammalian target of rapamycin) are expected to act as
rheostats during normal growth and development to fine-tune the
outcomes of the Myc/eIF4F FFL, representing promising targets for
cancer therapy (49).
By studying head and neck squamous cell carcinoma in
2009, Cohen and Rosner (50) and Cohen
et al (51) identified an FFL
in cell cycle regulation involving protein kinase Cα (PKCα) that
activates mitogen-activated protein kinase (MAPK), as well as
cyclin E translation via inhibition of miR-15a. Of note, while one
arm of the network entails classic transcriptional regulation of
cyclins by the MAPK pathway, the other arm involves regulation of
miR15a inhibiting cyclin E translation. The FFL is constitutively
driven by PKCα activation, leading to the unabated proliferation
inherent to cancer cells. Although the specific elements may
differ, their results suggest that FFL networks could play a
fundamental role in controlling DNA synthesis and cell cycle
progression in tumor cells. In this FFL, PKCα is not typically a
TF, the mechanism of how it downregulates miR-15a remains to be
elucidated.
Using a coculture model system, Rokavec et al
(52) showed a feed-forward
inflammatory signaling circuit in breast cancer in 2012. The
circuit was composed of miR-200c, p65, c-Jun N-terminal kinase 2
(JNK2), heat-shock factor 1 (HSF1) and interleukin 6 (IL-6).
Suppression of miR-200c by IL-6 constitutively activates p65/RelA
and JNK2, and the latter phosphorylates and activates HSF1. In
turn, HSF1 triggers demethylation of the IL-6 promoter that
facilitates the binding of p65 and c-Jun, which together drive
constitutive IL-6 transcription, promoting transformation in human
cancer cells and in a mouse model of ErbB2-driven breast
cancer.
Finding and confirming a specific
TF-miRNA-target gene FFL
Research has accumulated in this field of the
typical TF-miRNA-target protein coding gene formed FFL involved in
cancers. During the past decades, investigators have developed an
accession of procedure to predict, investigate and verify the
interaction of the 3 elements of a special FFL circuit. A series of
databases appeared with respectively different algorithms to offer
bioinformatic support of predicted connections.
miRNAs repress the translation of target genes by
binding, in a Watson-Crick complementary manner, to 7-nt long
sequences present at the 3′-untranslated region (3′-UTR) of the
regulated genes. The binding usually involves 2–8 nts of the miRNA,
known as the ‘seed’. The large amount of research associated with
the discovery of TF binding sites suggest that transcriptional and
post-transcriptional regulatory interactions could be predicted
in silico by searching over-represented short sequences of
nts present in promoters or 3′-UTRs and by filtering the results
with suitable evolutionary or functional constraints.
Independent computational evidence for the
regulatory interactions of the TF-miRNA-target gene FFL can be
extracted from the ECRbase, miRBase, PicTar and TargetScan
databases, with relevance to cancer of the TF-miRNA-target gene FFL
as deduced from their intersection with the oncomiR and cancer gene
census databases. In addition, Gene Ontology enrichment provides
detailed information regarding the joint targets of the loop
(5).
Establishment of the miRNA-target gene
association
Significant progress has been made in computational
algorithms for miRNA target prediction during the last decade.
Currently, there are databases such as TargetScan, miRanda, PicTar,
RNA22 and DIANA for target gene prediction (53–55)
(Table I). When miRNA and its
potential target gene emerge from bioinformatics, gain- and
loss-of-function methods, most commonly transfection, are ready to
be applied to evaluate the effect of the miRNA in respective human
cell lines. Immunology or molecular biology experiments, such as
western blot analysis, quantitative polymerase chain reaction
(qPCR), and miRNA and mRNA microarray expression profiling, are
also involved to verify the linking dynamic expression changes of
the miRNA, or its mimics and its target genes' mRNA and protein.
For microarray analysis, the stand-alone software tool CoExpress
(freely available at www.bioinformatics.lu/CoExpress), could be chosen to
perform interactive detection of correlated profiles in large
expression data sets. Finally, a luciferase reporter assay will
confirm associations between the miRNA and its targets (56). Publications of validated miR-target
correlations are recorded in TarBase.
 | Table I.Examples of common databases for
miRNA-TG and TF-TG interactions. |
Table I.
Examples of common databases for
miRNA-TG and TF-TG interactions.
Establishment of the TF-target gene
association
In an effort to further dissect the molecular
pathways regulated by a particular TF, several genome-wide screens
have been used to identify its transcriptional targets. The
transcription regulation databases include TRANSFAC, JASPAR, TRED,
DBTSS and TRRD (57–61). Among these databases mentioned,
TRANSFAC is the most commonly used (Table
I). First, researchers use bioinformatics to select consensus
DNA elements situated in the 5′ regulatory region of genes and
subsequently they measure TF binding to those sequences in
vivo by, most commonly, quantitative chromatin
immunoprecipitation (ChIP). Other experimental methods include
using the luciferase reporter gene, electrophoretic mobility shift
assays and DNase footprinting. Recently, as the technology of
microarray and new generation sequencing develop, high-throughput
methods based on ChIP are emerging. These advanced approaches are
ChIP-chip and ChIP-seq, which are expensive but extremely
promising. Classic ChIP results from electrophoretic analysis of
PCR amplification products, and this method can only observe some
specific target genes. However, the emergence of ChIP-chip and
ChIP-seq technology has made the observation on the whole genome of
protein and DNA combination possible (62–66).
Establishment of the TF-miRNA
association
Based on bioinformatic prediction, using a miRNA
microarray containing different miRNAs and a set of miRNA qPCR
assays to validate the microarray results, the correct miRNAs can
be identified that are induced by a special TF in vitro,
whether upregulated or downregulated with the TF amplification. The
TransmiR database can also be used, which is a TF-miRNA regulatory
database built by researchers from Peking University (Beijing
China) who manually surveyed ~5,000 reports in the literature and
identified 243 TF-miRNA regulatory associations, which were
supported experimentally from 86 studies (67).
In 2012, Yan et al (68) described a novel method for integrating
gene and miRNA expression profiles in cancer using FFLs consisting
of TFs, miRNAs and their common target genes. This was the
dChip-GemiNi (Gene and miRNA Network-based Integration) method,
available at www.canevolve.org/dChip-GemiNi/usergemini.php. It
statistically ranks computationally predicted TF-miRNA-target gene
FFLs by their explanatory power to account for differential gene
and miRNA expression between two biological conditions, such as
normal and cancer. Compared to existing approaches, GemiNI also
computationally derives information regarding TF-target gene and
miRNA-mRNA interactions. The integrated modeling of expression data
and FFLs better identifies cancer-related TFs and miRNAs.
All the connections of these TF-miRNA-target gene
FFLs were based on experimentally validated interactions referenced
in the Ingenuity Knowledge Base, based on which the powerful
software Ingenuity Pathway Analysis (IPA) was built.
Conclusions
As the concepts of transcription network motifs and
TF-miRNA-target gene FFLs emerge, our understanding of the
molecular deregulatory mechanism of the cancer step delves further
into the unknown field. Researchers could study the complicated
deregulation system involving numerous elements at a higher level.
Associated TF-miRNA-target gene FFLs can be confirmed and collected
to form a regulatory network, similar to a jigsaw puzzle. Key nodes
may be used therapeutically as a target for drugs or as the drug
itself. More experimental analyses in vivo and more accurate
network constituent data analyses may lead to the discovery of
crucial principles of cancer, which indicates curability and hope
of overcoming malignancy.
However, since miRNAs can also regulate other
non-coding RNAs (for example, long non-coding RNAs), which have a
role in cancer development and vice versa (69), and TFs are also involved in the
regulation of other cancer-associated non-coding RNAs. Each member
as a node of a special FFL can be another motifs' indispensable
element, which indicates that the overlaying of TF-miRNA-target
gene FFLs should put other types of motifs (such as FBL) into
consideration. These two elements make the profile of transcription
networks more complex and have a larger role than expected, which
is currently unknown. New strategies to identify and characterize
the entire targets of individual miRNAs and TFs, with an improved
high-throughput, and to determine how they function in combination
to regulate specific targets, will be required to understand their
action on cancer pathology.
Acknowledgements
The present study was supported by grants from the
National Natural Science Foundation of China (nos. 81101824 and
81302112) and from the Outstanding Youth Science Foundation of
Tongji Hospital (no. YXQN005).
References
|
1
|
Siegel R, Naishadham D and Jemal A: Cancer
statistics, 2012. CA Cancer J Clin. 62:10–29. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Milo R, Shen-Orr S, Itzkovitz S, Kashtan
N, Chklovskii D and Alon U: Network motifs: Simple building blocks
of complex networks. Science. 298:824–827. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Widder S, Solé R and Macía J: Evolvability
of feed-forward loop architecture biases its abundance in
transcription networks. BMC Syst Biol. 6:72012. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Shen-Orr SS, Milo R, Mangan S and Alon U:
Network motifs in the transcriptional regulation network of
Escherichia coli. Nat Genet. 31:64–68. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Re A, Corá D, Taverna D and Caselle M:
Genome-wide survey of microRNA-transcription factor feed-forward
regulatory circuits in human. Mol Biosyst. 5:854–867. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Ma HW, Kumar B, Ditges U, Gunzer F, Buer J
and Zeng AP: An extended transcriptional regulatory network of
Escherichia coli and analysis of its hierarchical structure and
network motifs. Nucleic Acids Res. 32:6643–6649. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Mangan S and Alon U: Structure and
function of the feed-forward loop network motif. Proc Natl Acad Sci
USA. 100:11980–11985. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
He XX, Guo AY, Xu CR, et al:
Bioinformatics analysis identifies miR-221 as a core regulator in
hepatocellular carcinoma and its silencing suppresses tumor
properties. Oncol Rep. 32:1200–1210. 2014.PubMed/NCBI
|
|
9
|
Yan JW, Lin JS and He XX: The emerging
role of miR-375 in cancer. Int J Cancer. 135:1011–1018. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Li Y, Liang C, Easterbrook S, Luo J and
Zhang Z: Investigating the functional implications of reinforcing
feedback loops in transcriptional regulatory networks. Mol Biosys.
10:3238–3248. 2014. View Article : Google Scholar
|
|
11
|
Fujita Y, Komatsu N, Matsuda M and Aoki K:
Fluorescence resonance energy transfer based quantitative analysis
of feedforward and feedback loops in epidermal growth factor
receptor signaling and the sensitivity to molecular targeting
drugs. FEBS J. 281:3177–3192. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Hsieh WT, Tzeng KR, Ciou JS, Tsai JJ,
Kurubanjerdjit N, Huang CH, et al: Transcription factor and
microRNA-regulated network motifs for cancer and signal
transduction networks. BMC Syst Biol. 9:(Suppl 1). S52015.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Latchman DS: Transcription factors: An
overview. Int J Biochem Cell Biol. 29:1305–1312. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Mitchell PJ and Tjian R: Transcriptional
regulation in mammalian cells by sequence-specific DNA binding
proteins. Science. 245:371–378. 1989. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Ptashne M and Gann A: Transcriptional
activation by recruitment. Nature. 386:569–577. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
van Nimwegen E: Scaling laws in the
functional content of genomes. Trends Genet. 19:479–484. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Lane D and Levine A: p53 Research: The
past thirty years and the next thirty years. Cold Spring Harb
Perspect Biol. 2:a0008932010. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Hainaut P and Wiman KG: 30 years and a
long way into p53 research. Lancet Oncol. 10:913–919. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Hermeking H: p53 enters the microRNA
world. Cancer Cell. 12:414–418. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Brosh R, Shalgi R, Liran A, Landan G,
Korotayev K, Nguyen GH, Enerly E, Johnsen H, Buganim Y, Solomon H,
et al: p53-Repressed miRNAs are involved with E2F in a feed-forward
loop promoting proliferation. Mol Syst Biol. 4:2292008. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Altshuler ML, Severin SE and Glukhov AI:
The tumor cell and telomerase. Biochemistry (Mosc). 68:1275–1283.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Sanda T, Lawton LN, Barrasa MI, Fan ZP,
Kohlhammer H, Gutierrez A, Ma W, Tatarek J, Ahn Y, Kelliher MA, et
al: Core transcriptional regulatory circuit controlled by the TAL1
complex in human T cell acute lymphoblastic leukemia. Cancer Cell.
22:209–221. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Bartel DP: MicroRNAs: Target recognition
and regulatory functions. Cell. 136:215–233. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Lujambio A and Lowe SW: The microcosmos of
cancer. Nature. 482:347–355. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Calin GA and Croce CM: MicroRNA-cancer
connection: The beginning of a new tale. Cancer Res. 66:7390–7394.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Lewis BP, Burge CB and Bartel DP:
Conserved seed pairing, often flanked by adenosines, indicates that
thousands of human genes are microRNA targets. Cell. 120:15–20.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Eiring AM, Harb JG, Neviani P, Garton C,
Oaks JJ, Spizzo R, Liu S, Schwind S, Santhanam R, Hickey CJ, et al:
miR-328 functions as an RNA decoy to modulate hnRNP E2 regulation
of mRNA translation in leukemic blasts. Cell. 140:652–665. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Moretti F, Thermann R and Hentze MW:
Mechanism of translational regulation by miR-2 from sites in the 5′
untranslated region or the open reading frame. RNA. 16:2493–2502.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Shalgi R, Brosh R, Oren M, Pilpel Y and
Rotter V: Coupling transcriptional and post-transcriptional miRNA
regulation in the control of cell fate. Aging (Albany NY).
1:762–770. 2009.PubMed/NCBI
|
|
30
|
Johnson SM, Grosshans H, Shingara J, Byrom
M, Jarvis R, Cheng A, Labourier E, Reinert KL, Brown D and Slack
FJ: RAS is regulated by the let-7 microRNA family. Cell.
120:635–647. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Burk U, Schubert J, Wellner U, Schmalhofer
O, Vincan E, Spaderna S and Brabletz T: A reciprocal repression
between ZEB1 and members of the miR-200 family promotes EMT and
invasion in cancer cells. EMBO Rep. 9:582–589. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Hwang CI, Choi J, Zhou Z, Flesken-Nikitin
A, Tarakhovsky A and Nikitin AY: MET-dependent cancer invasion may
be preprogrammed by early alterations of p53-regulated feedforward
loop and triggered by stromal cell-derived HGF. Cell Cycle.
10:3834–3840. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Avraham R and Yarden Y: Regulation of
signalling by microRNAs. Biochem Soc Trans. 40:26–30. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Kobayashi K, Sakurai K, Hiramatsu H, Inada
K, Shiogama K, Nakamura S, et al: The miR-199a/Brm/EGR1 axis is a
determinant of anchorage-independent growth in epithelial tumor
cell lines. Sci Rep. 5:84282015. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
He L, Thomson JM, Hemann MT,
Hernando-Monge E, Mu D, Goodson S, Powers S, Cordon-Cardo C, Lowe
SW, Hannon GJ, et al: A microRNA polycistron as a potential human
oncogene. Nature. 435:828–833. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Garzon R, Marcucci G and Croce CM:
Targeting microRNAs in cancer: Rationale, strategies and
challenges. Nat Rev Drug Discov. 9:775–789. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Song S and Ajani JA: The role of microRNAs
in cancers of the upper gastrointestinal tract. Nat Rev
Gastroenterol Hepatol. 10:109–118. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Chang TC, Yu D, Lee YS, Wentzel EA, Arking
DE, West KM, Dang CV, Thomas-Tikhonenko A and Mendell JT:
Widespread microRNA repression by Myc contributes to tumorigenesis.
Nat Genet. 40:43–50. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Jiang X, Huang H, Li Z, Li Y, Wang X,
Gurbuxani S, et al: Blockade of miR-150 maturation by
MLL-fusion/MYC/LIN-28 is required for MLL-associated leukemia.
Cancer Cell. 22:524–535. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Musilova K and Mraz M: MicroRNAs in B-cell
lymphomas: How a complex biology gets more complex. Leukemia.
29:1004–1017. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Zhao X, Lwin T, Zhang X, Huang A, Wang J,
Marquez VE, Chen-Kiang S, Dalton WS, Sotomayor E and Tao J:
Disruption of the MYC-miRNA-EZH2 loop to suppress aggressive B-cell
lymphoma survival and clonogenicity. Leukemia. 27:2341–2350. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
42
|
O'Donnell KA, Wentzel EA, Zeller KI, Dang
CV and Mendell JT: c-Myc-regulated microRNAs modulate E2F1
expression. Nature. 435:839–843. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Ning Z, Wang A, Liang J, et al: USP22
promotes epithelial-mesenchymal transition via the FAK pathway in
pancreatic cancer cells. Oncol Rep. 32:1451–1458. 2014.PubMed/NCBI
|
|
44
|
Kent OA, Chivukula RR, Mullendore M,
Wentzel EA, Feldmann G, Lee KH, Liu S, Leach SD, Maitra A and
Mendell JT: Repression of the miR-143/145 cluster by oncogenic Ras
initiates a tumor-promoting feed-forward pathway. Genes Dev.
24:2754–2759. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
El Baroudi M, Corà D, Bosia C, Osella M
and Caselle M: A curated database of miRNA mediated feed-forward
loops involving MYC as master regulator. PLoS One. 6:e147422011.
View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Polioudakis D, Bhinge AA, Killion PJ, Lee
BK, Abell NS and Iyer VR: A Myc-microRNA network promotes exit from
quiescence by suppressing the interferon response and cell-cycle
arrest genes. Nucleic Acids Res. 41:2239–2254. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
He L, He X, Lim LP, de Stanchina E, Xuan
Z, Liang Y, Xue W, Zender L, Magnus J, Ridzon D, et al: A microRNA
component of the p53 tumour suppressor network. Nature.
447:1130–1134. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Lin CJ, Cencic R, Mills JR, Robert F and
Pelletier J: c-Myc and eIF4F are components of a feedforward loop
that links transcription and translation. Cancer Res. 68:5326–5334.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Lin CJ, Malina A and Pelletier J: c-Myc
and eIF4F constitute a feedforward loop that regulates cell growth:
Implications for anticancer therapy. Cancer Res. 69:7491–7494.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Cohen EE and Rosner MR: MicroRNA-regulated
feed forward loop network. Cell Cycle. 8:2477–2478. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Cohen EE, Zhu H, Lingen MW, Martin LE, Kuo
WL, Choi EA, Kocherginsky M, Parker JS, Chung CH and Rosner MR: A
feed-forward loop involving protein kinase Calpha and microRNAs
regulates tumor cell cycle. Cancer Res. 69:65–74. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Rokavec M, Wu W and Luo J-L: IL6-mediated
suppression of miR-200c directs constitutive activation of
inflammatory signaling circuit driving transformation and
tumorigenesis. Mol Cell. 45:777–789. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Alexiou P, Maragkakis M, Papadopoulos GL,
Reczko M and Hatzigeorgiou AG: Lost in translation: An assessment
and perspective for computational microRNA target identification.
Bioinformatics. 25:3049–3055. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Reyes-Herrera PH and Ficarra E: One decade
of development and evolution of microRNA target prediction
algorithms. Genomics Proteomics Bioinformatics. 10:254–263. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Mendes ND, Freitas AT and Sagot MF:
Current tools for the identification of miRNA genes and their
targets. Nucleic Acids Res. 37:2419–2433. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Nazarov PV, Reinsbach SE, Muller A, Nicot
N, Philippidou D, Vallar L and Kreis S: Interplay of microRNAs,
transcription factors and target genes: Linking dynamic expression
changes to function. Nucleic Acids Res. 41:2817–2831. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Dubchak I, Munoz M, Poliakov A, Salomonis
N, Minovitsky S, Bodmer R and Zambon AC: Whole-Genome rVISTA: A
tool to determine enrichment of transcription factor binding sites
in gene promoters from transcriptomic data. Bioinformatics.
29:2059–2061. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Krystkowiak I, Lenart J, Debski K, Kuterba
P, Petas M, Kaminska B and Dabrowski M: Nencki Genomics Database -
Ensembl funcgen enhanced with intersections, user data and
genome-wide TFBS motifs. Database (Oxford). 2013:bat0692013.
View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Jiang C, Xuan Z, Zhao F and Zhang MQ:
TRED: A transcriptional regulatory element database, new entries
and other development. Nucleic Acids Res. 35:(Database). D137–D140.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Yamashita R, Sugano S, Suzuki Y and Nakai
K: DBTSS: DataBase of Transcriptional Start Sites progress report
in 2012. Nucleic Acids Res. 40:(D1). D150–D154. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Kolchanov NA, Podkolodnaia OA, Anan'ko EA,
Ignat'eva EV, Podkolodnyĭ NL, Merkulov VM, Stepanenko IL,
Pozdniakov MA, Belova OE, Grigorovich DA, et al: Regulation of
eukaryotic gene transcription: Description in the TRRD database.
Mol Biol (Mosk). 35:934–942. 2001.(In Russian). PubMed/NCBI
|
|
62
|
Robertson G, Hirst M, Bainbridge M,
Bilenky M, Zhao Y, Zeng T, Euskirchen G, Bernier B, Varhol R,
Delaney A, et al: Genome-wide profiles of STAT1 DNA association
using chromatin immunoprecipitation and massively parallel
sequencing. Nat Methods. 4:651–657. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Perez-Pinera P, Ousterout DG, Brunger JM,
Farin AM, Glass KA, Guilak F, Crawford GE, Hartemink AJ and
Gersbach CA: Synergistic and tunable human gene activation by
combinations of synthetic transcription factors. Nat Methods.
10:239–242. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Hesselberth JR, Chen X, Zhang Z, Sabo PJ,
Sandstrom R, Reynolds AP, Thurman RE, Neph S, Kuehn MS, Noble WS,
et al: Global mapping of protein-DNA interactions in vivo by
digital genomic footprinting. Nat Methods. 6:283–289. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Hellman LM and Fried MG: Electrophoretic
mobility shift assay (EMSA) for detecting protein-nucleic acid
interactions. Nat Protoc. 2:1849–1861. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
66
|
Fernandez PC, Frank SR, Wang L, Schroeder
M, Liu S, Greene J, Cocito A and Amati B: Genomic targets of the
human c-Myc protein. Genes Dev. 17:1115–1129. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
67
|
Wang J, Lu M, Qiu C and Cui Q: TransmiR: A
transcription factor-microRNA regulation database. Nucleic Acids
Res. 38:(Database). D119–D122. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
68
|
Yan Z, Shah PK, Amin SB, Samur MK, Huang
N, Wang X, Misra V, Ji H, Gabuzda D and Li C: Integrative analysis
of gene and miRNA expression profiles with transcription
factor-miRNA feed-forward loops identifies regulators in human
cancers. Nucleic Acids Res. 40:e1352012. View Article : Google Scholar : PubMed/NCBI
|
|
69
|
Poliseno L, Salmena L, Zhang J, Carver B,
Haveman WJ and Pandolfi PP: A coding-independent function of gene
and pseudogene mRNAs regulates tumour biology. Nature.
465:1033–1038. 2010. View Article : Google Scholar : PubMed/NCBI
|