Introduction
Low back pain (LBP), one of the most common
musculoskeletal complaints, has become a serious health and
socioeconomic problem affecting modern society (1,2).
Among numerous pain generators in the lumbar spine, intervertebral
disc (IVD) degeneration is considered to be the most important
contributor to LBP (3,4). IVD degeneration results from the
excessive degradation and impaired synthesis of the extracellular
matrix (ECM) secreted by nucleus pulposus (NP) cells. This
imbalance between catabolic and anabolic activity leads to changes
in the biological and mechanical characteristics of IVDs (5–11).
A number of previous studies have proven that IVD
degeneration is linked to various pro-inflammatory cytokines and
metabolites, including interleukin (IL)-1β, IL-6, tumor necrosis
factor-α (TNF-α), lipopolysaccharide (LPS) and reactive oxygen
species (ROS) (12–17). LPS, an endotoxin which exists in
the outer membrane of Gram-negative bacteria, has been used to
induce inflammatory responses in several pre-clinical models of
inflammation (18,19). There is evidence to indicate that
LPS is a potential inducer of multiple pro-inflammatory cytokines
(IL-1β, IL-6 and TNF-α) through binding to Toll-like receptor
(TLR)-2 and TLR-4 (18,20–24). In cartilaginous tissue, the
mitogen-activated protein kinases (MAPK) pathway, with p38,
extracellular signal-regulated kinase (ERK) and c-Jun N-terminal
kinase (JNK) as major members, is an important downstream
inflammatory pathway. The results from previous studies on
cartilage degeneration have indicated that the MAPK pathway plays a
pivotal role in the inflammatory process (25,26). Current treatment strategies for
LBP are limited to conservative treatment (e.g., physical therapy
and oral analgesics) and surgical intervention (27–29); anti-inflammatory and regenerative
drug treatment has not been applied to clinical practice. Although
few studies have focused on the application of plant-derived
compounds to attenuate IVD degeneration, some anti-inflammatory
candidates may provide us with novel therapeutic strategeis for the
treatment of LBP.
Crocus sativus L. (or saffron), which belongs
to the Iridaceae family, is often used as a culinary spice or an
anodyne or tranquilizer in traditional Chinese medicine. Crocin is
the main constituent which is responsible for the multiple
bioactivities of saffron. Previous studies have demonstrated the
anti-inflammatory effects of crocin which are mediated through the
inhibition of nitric oxide (NO) synthesis in LPS-induced
inflammation (30–32). Crocin has recently been reported
to exert an anti-arthritic effect on cartilaginous tissues,
suggesting that crocin protects cartilage from IL-1β-induced
inflammation by suppressing the expression of matrix
metalloproteinases (MMPs) (33).
Therefore, in this study, we aimed to further
explore the reported anti-inflammatory effects of crocin by
investigating the role of crocin in inflammation induced by LPS in
the NP. In the present study, inflammation-related proteolytic
enzymes and pro-inflammatory factors involved in IVD degeneration
were explored as inflammatory markers. A disc organ culture system,
serving as an ex vivo model of IVD degeneration, was used to
conduct histological and biochemical assays. Furthermore, we
investigated the potential mechanisms responsible for the
anti-inflammatory effects of crocin by analyzing the relevant
signaling pathways.
Materials and methods
Reagents and antibodies
The chemical reagents, including crocin, used in the
present study were purchased from Sigma-Aldrich (St. Louis, MO,
USA) unless stated otherwise. Stock solution of crocin was prepared
in phosphate-buffered saline (PBS) and stored at −20°C. The MAPK
family antibody sampler kit (Cat. no. 9926) and phospho-MAPK family
antibody sampler kit (Cat. no. 9910) were obtained from Cell
Signaling Technology, Inc. (Beverly, MA, USA). Anti-type II
collagen (collagen-II) antibody (ab34712) was purchased from Abcam
(Cambridge, MA, USA).
NP cell isolation and culture
The present study was conducted in strict accordance
with the recommendations in the Guide for the Care and Use of
Laboratory Animals of the National Institutes of Health. The study
protocol was approved by the Animal Care and Experiment Committee
of Shanghai Jiao Tong University School of Medicine, Shanghai,
China. A total of 30 Sprague-Dawley rats (3 months old) were
provided by the Experimental Animal Center of Shanghai Ninth
People's Hospital (Shanghai, China). Lumbar IVDs were harvested
from the rats immediately following sacrifice (by intraperitoneal
injection of excessive chloral hydrate). Under a magnifier, the NP
tissues were carefully removed without adhesion of the annulus
fibrosus and endplate from the IVD. To ensure the purity of the NP
cells, the NP tissues were rinsed with DMEM/F-12 medium many times
until all impurities were removed. The NP tissues were then
dissected into 1-mm3 sections for sufficient digestion.
The NP sections were digested in 0.25% trypsin (Sigma-Aldrich) for
15 min and 0.025% type II collagenase (Sigma-Aldrich) for 4 h at
37°C. Tissues debris was removed from the digestion product using a
70-µM filter. The NP cells were cultured in DMEM/F-12 medium
containing 10% fetal bovine serum (FBS) and 1%
penicillin/streptomycin at 37°C, in a 5% CO2 humidified
incubator. When the primary cells reached a confluence of 80–90%,
they were harvested, replated and allowed to proliferate. The
second-generation cells were used throughout the experiments.
Cytotoxicity assay
Cell viability was measured with a Cell Counting
Kit-8 (CCK-8; Dojindo Laboratories, Kumamoto, Japan) according to
the instructions provided by the manufacturer. Briefly, the rat NP
cells were seeded in 96-well plates at a density of
5×103 cells/well, and then incubated with various
concentrations of crocin (10, 50, 100 and 200 µM) for 24 h.
CCK-8 solution (10 µl) was added to each well and the plates
were incubated for 2 h. The absorbance of each well was measured at
450 nm using a microplate reader (Omega Bio-Tek, Inc., Norcross,
GA, USA) with the culture medium as a blank. Cell viability was
calculated using the following formula: cell viability (%) =
[absorbance (with crocin) - absorbance (blank)]/[absorbance
(without crocin) - absorbance (blank)] ×100.
Cell treatment
The NP cells were seeded in 6-well plates at a
density of 1×105 cells/well. After reaching a confluence
of 80–90%, the cells were treated with or without various
concentrations of crocin (10, 50 and 100 µM) for 2 h,
followed by stimulation with LPS (Sigma-Aldrich; 10 µg/ml)
for 24 h. The cells were harvested for use in an expression assay
of IVD degeneration-related genes. Supernatants were collected for
the evaluation of MMP-3 and MMP-13 expression by enzyme-linked
immunosorbent assay (ELISA). For western blot analysis, the cells
were pre-treated with crocin for 4 h, and total proteins were
extracted 30 min following stimulation with LPS.
Isolation of RNA and reverse
transcription-quantitative polymerase chain reaction (RT-qPCR)
Total RNA was isolated using TRIzol reagent
(Invitrogen, Carlsbad, CA, USA) following the manufacturer's
instructions. Reverse transcription was carried out with 1
µg of total RNA using a reverse transcriptase kit (Takara
Bio, Inc., Otsu, Japan) for first-strand complementary DNA (cDNA)
synthesis. Quantitative (real-time) PCR (qPCR) was conducted with a
SYBR Premix Ex Taq kit (Takara Bio, Inc.) and an ABI 7500
Sequencing Detection System (Applied Biosystems/Life Technologies,
Foster City, CA, USA) according to the manufacturer's instructions.
Relative gene expression was measured using the ∆∆Ct method with
β-actin as an internal control. The primer sequences are presented
in Table I.
 | Table ISequences of primers used for
RT-qPCR. |
Table I
Sequences of primers used for
RT-qPCR.
| Gene | Primer sequences
(5′-3′) |
|---|
| MMP-1 | F:
AGCTCATACAGTTTCCCCGT |
| R:
GCCTCAGCTTTTCAGCCATC |
| MMP-3 | F:
TTTGGCCGTCTCTTCCATCC |
| R:
GCATCGATCTTCTGGACGGT |
| MMP-13 | F:
ACCATCCTGTGACTCTTGCG |
| R:
TTCACCCACATCAGGCACTC |
| ADAMTS-4 | F:
ACCGATTACCAGCCTTTGGG |
| R:
CCGACTCCGGATCTCCATTG |
| ADAMTS-5 | F:
CCGAACGAGTTTACGGGGAT |
| R:
TGTGCGTCGCCTAGAACTAC |
| IL-1β | F:
GCACAGTTCCCCAACTGGTA |
| R:
GGAGACTGCCCATTCTCGAC |
| IL-6 | F:
CATTCTGTCTCGAGCCCACC |
| R:
AGTCTCCTCTCCGGACTTGT |
| TNF-α | F:
TCGTAGCAAACCACCAAGCA |
| R:
TCGTAGCAAACCACCAAGCA |
| iNOS | F:
ACACAGTGTCGCTGGTTTGA |
| R:
AGAAACTTCCAGGGGCAAGC |
| TLR-2 | F:
TGGAGGTCTCCAGGTCAAATC |
| R:
TGTTTGCTGTGAGTCCCGAG |
| Aggrecan | F:
CAGATGGCACCCTCCGATAC |
| R:
GACACACCTCGGAAGCAGAA |
| Collagen-II | F:
GGCCAGGATGCCCGAAAATTA |
| R:
ACCCCTCTCTCCCTTGTCAC |
| β-actin | F:
AACCTTCTTGCAGCTCCTCCG |
| R:
CCATACCCACCATCACACCCT |
ELISA
As described above, following pre-treatment with
various concentrations of crocin (0, 10, 50 and 100 µM) for
2 h, the NP cells were stimulated with LPS (10 µg/ml) for 24
h. The supernatants were then collected and centrifuged to remove
cell fragments. The levels of MMP-3 and MMP-13 in the supernatant
were measured using an ELISA kit (R&D Systems, Inc.,
Minneapolis, MN, USA) and a microplate reader (Omega Bio-Tek, Inc.)
according to the manufacturer's instructions.
Western blot analysis
Total protein was extracted using
radio-immunoprecipitation assay (RIPA) lysis buffer containing a
mixture of protease inhibitor, and the protein amount was
determined by the Bradford method. The denatured protein was
separated by sodium dodecyl sulfate-polyacrylamide gel
electrophoresis (SDS-PAGE), transferred onto PVDF membranes
(Millipore Corp., Billerica, MA, USA) and probed with the primary
antibodies against p38, p-p38, ERK, p-ERK, JNK, p-JNK and β-actin.
Following incubation with the appropriate secondary antibodies
conjugated with IRDye 800CW (molecular weight 1162 Da), bands of
interest were detected by exposure using an Odyssey infrared
imaging system (LI-COR Biosciences, Lincoln, NE, USA). Quantitative
analysis of the intensity of bands was done using Adobe Photoshop
CS6 (Adobe Systems Incorporated, San Jose, CA, USA).
Organ culture
An extra 10 Sprague-Dawley rats were euthanized by
an injection of a lethal dose of chloral hydrate. The entire discs
were harvested, including the vertebral endplates, annulus fibrosus
and NP, and then placed into a 12-well plate. The discs were
cultured in DMEM/F-12 medium containing 10% FBS, 1%
insulin-transferrin-selenium (Sigma-Aldrich), 50 µg/ml
ascorbate-2-phosphate and 1% penicillin/streptomycin. The medium
was replaced every day. Additional NaCl was added to the medium to
raise the osmolarity to 410 mOsm/kg, as previously described
(34,35). In different groups, the discs were
cultured for 7 days with or without 10 µg/ml LPS and 100
µM crocin. Moreover, at the end of the culture period, the
discs were treated with 0.75 mg/ml nitroblue tetrazolium (NBT;
Sigma-Aldrich) in fresh complete medium and incubated at 37°C for
24 h.
Histological analysis
Following 7 days of culture, the discs were
harvested, fixed in 4% paraformaldehyde and decalcified in EDTA for
14 days. The decalcified discs were embedded in paraffin for
sectioning. Serial mid-sagittal sections of discs
(5-µm-thick) were obtained to prepare slides. NBT and
4,6-diamino-2-phenyl-indole (DAPI; Sigma-Aldrich) dual staining was
used to evaluate cell viability in the organ cultures as previously
described (36). NBT staining was
imaged under transmitted light illumination using a microscope
(LEICA DM4000 B; Leica Microsystems, Wetzlar, Germany) and DAPI
staining was imaged under ultraviolet illumination (359 nm) using
the same microscope (LEICA DM4000 B; Leica Microsystems). Viable
cells were defined as cells with both DAPI and NBT staining, and
cells with DAPI staining alone were deemed dead. A total of 3
sections of each disc were analyzed and 3 fields (magnification,
×400) of each section were selected to calculate the average. Cell
viability was calculated as follows: cell viability (%) = live
cells in field (NBT stains)/total cells in field (DAPI signal)
×100. Mid-sagittal sections were also stained with
hematoxylin-eosin (H&E) and Safranin O-fast green stain to
assess the degeneration of IVDs. All H&E staining and Safranin
O-fast green staining slides were imaged under transmitted light
illumination using a microscope (LEICA DM4000 B; Leica
Microsystems). All staining images were observed and assessed by an
unblinded investigator, while the evaluation of the content of the
ECM was based on staining intensity in the ECM area. Collagen-II
expression was detected by immunohistochemistry (IHC) using a mouse
monoclonal antibody (ab34712; 1:200; Abcam) and a horseradish
peroxidase-conjugated anti-mouse antibody (K5007; 1:50; Dako,
Glostrup, Denmark), followed by color development with
diaminobenzidine tetrahydrochloride (DAB; Dako). The results of
collagen-II staining were quantified and the integrated optical
density (IOD) was determined using Image Pro-Plus (IPP) version 6.0
software (Media Cybernetics, Inc., Bethesda, MD, USA).
Proteoglycan content assay
Following 7 days of culture, NP tissue was isolated
from each disc and then digested with papain at 37°C for 48 h
(37). The proteoglycan content
of the papain digests was determined by the dimethylmethylene blue
(DMMB) assay as previously described (38). The DMMB assay was standardized by
the dsDNA content of the NP tissue, which was determined by an
assay of the total dsDNA content using the Quant-iT™
PicoGreen® dsDNA Assay kit (Invitrogen).
Statistical analysis
All experiments were repeated at least 3 times. The
results are expressed as the means ± standard deviation (SD).
Statistical analysis was performed with a one-way analysis of
variance (ANOVA) for multiple comparisons using SPSS 19.0 software
(IBM, Corp., Armonk, NY, USA). A value of P<0.05 was considered
to indicate a statistically significant difference.
Results
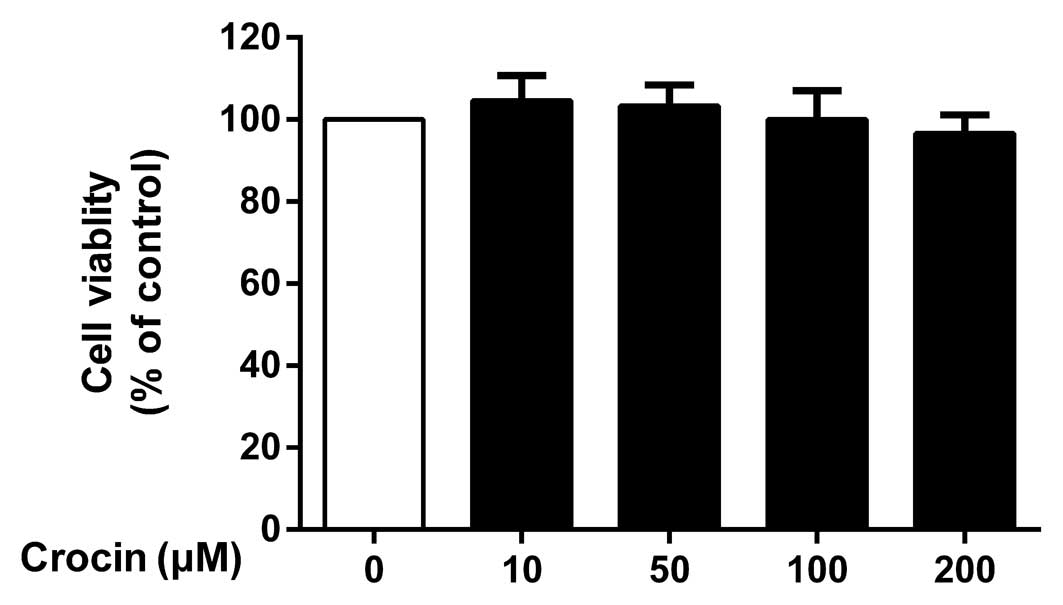
Potential cytotoxicity of crocin
To determine the potential cytotoxicity of crocin,
the NP cells were incubated with various concentrations (10, 50,
100 and 200 µM) of crocin for 24 h prior to performing a
CCK-8 assay. Crocin did not exhibit any significant cytotoxic
effects at concentrations of 10–200 µM (P>0.05; Fig. 1).
Crocin inhibits the LPS-induced increase
in the mRNA expression of catabolic enzymes and pro-inflammatory
factors in NP cells
As shown by RT-qPCR, the upregulated mRNA expression
of MMPs (MMP-1, MMP-3 and MMP-13) and a disintegrin-like and
metalloprotease (reprolysin type) with thrombospondin type 1 motif
(ADAMTS; ADAMTS-4 and ADAMTS-5) induced by LPS was significantly
inhibited by crocin in a concentration-dependent manner (Fig. 2A–E). However, the lowest
concentration (10 µM) of crocin used did not significantly
inhibit the increase in the expression of ADAMTS-4 and ADAMTS-5
induced by LPS (Fig. 2D and E).
Similarly, LPS markedly increased the mRNA expression of
pro-inflammatory and oxidative stress factors [IL-1β, TNF-α, IL-6
and inducible NO synthase (iNOS)]; this increase was significantly
inhibited by pre-treatment with crocin in a concentration-dependent
manner (Fig. 2F–I). Moreover,
crocin partly reversed the LPS-induced decrease in the mRNA
expression of aggrecan and collagen-II at the concentration of 50
and 100 µM (Fig. 2J and
K). Notably, the overexpression of TLR-2 induced by LPS was
also significantly suppressed by pre-treatment with crocin
(Fig. 2L).
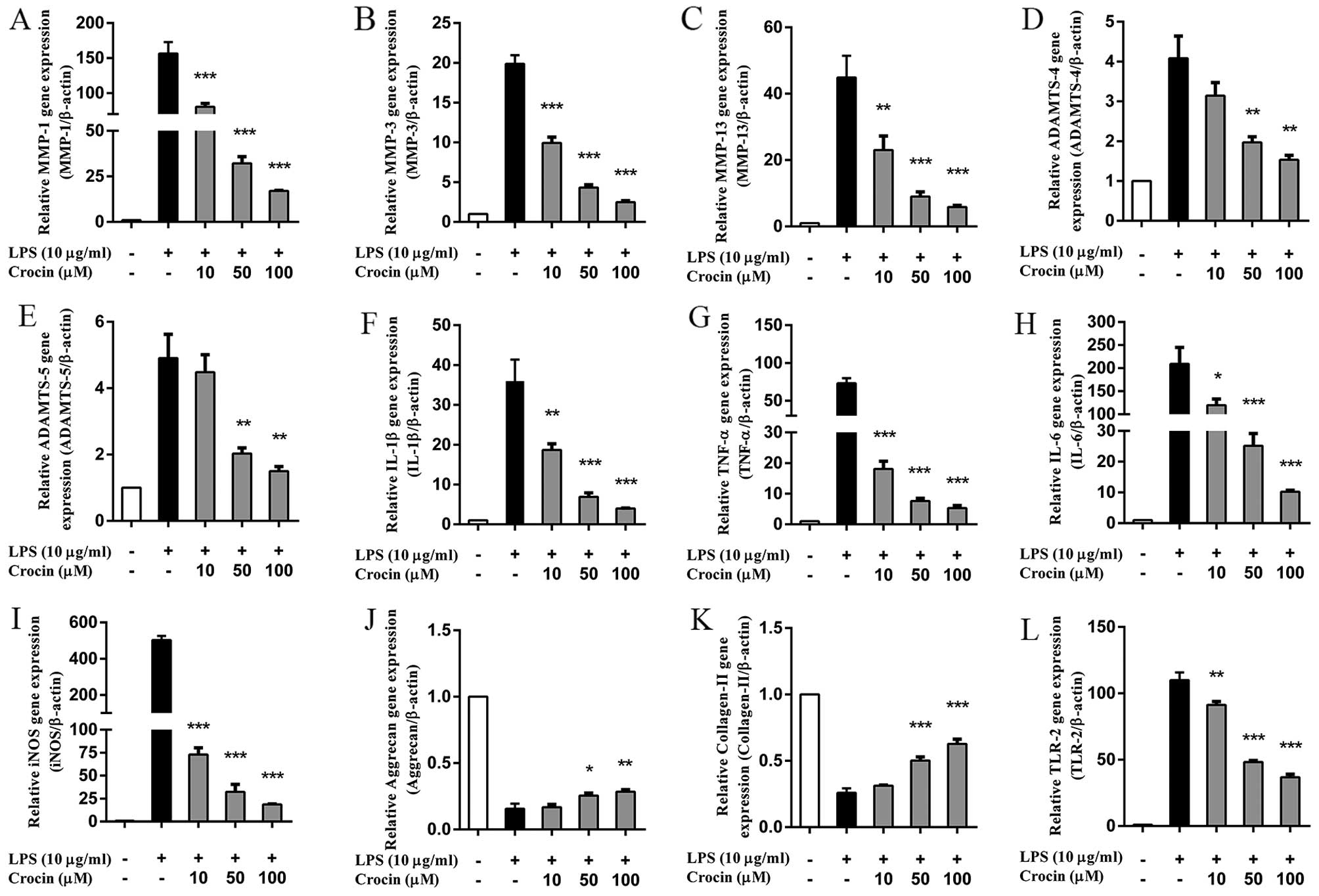 | Figure 2Effects of crocin on the
lipopolysaccharide (LPS)-induced increase in the mRNA expression of
intervertebral disc (IVD) degeneration-related genes. (A-L) The
relative mRNA expression of (A) MMP-1, (B) MMP-3, (C) MMP-13, (D)
ADAMTS-4, (E) ADAMTS-5, (F) IL-1β, (G) TNF-α, (H) IL-6, (I) iNOS,
(J) aggrecan, (K) collagen-II and (L) TLR-2 was evaluated by
RT-qPCR. Following treatment with various concentrations of crocin
(10, 50 and 100 µM) for 2 h, nucleus pulposus (NP) cells
were stimulated with LPS (10 µg/ml) for 24 h and then
harvested for us in RT-qPCR. Results are expressed as the means ±
SD. *P<0.05, **P<0.01,
***P<0.001, compared to the LPS-stimulated group. |
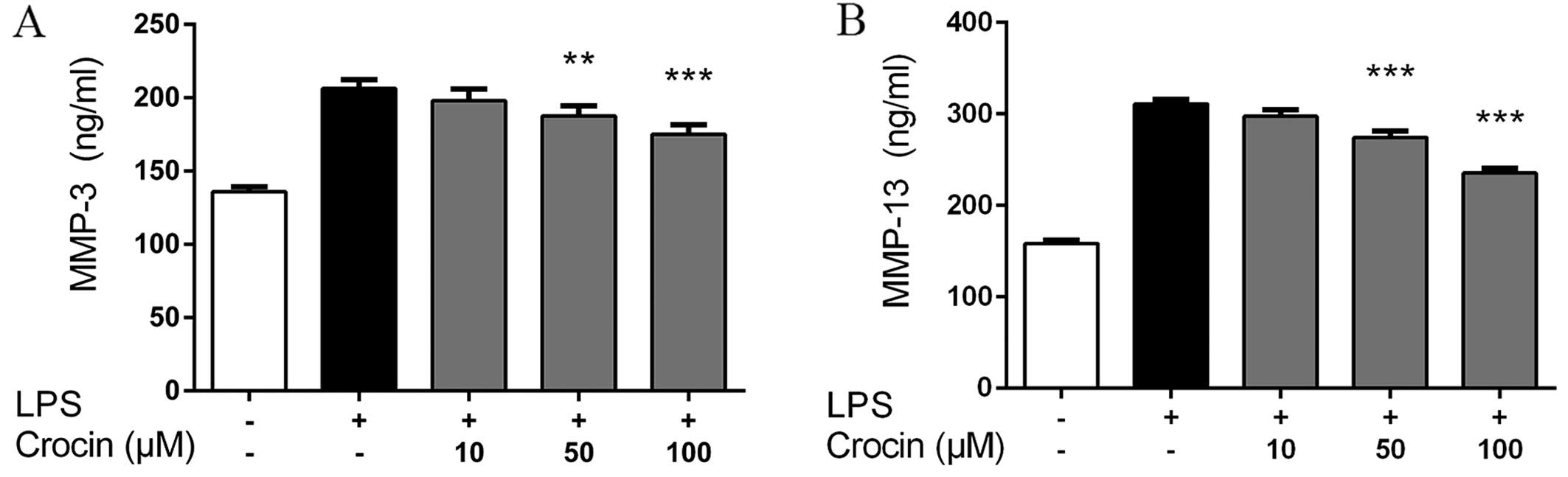
Crocin inhibits the LPS-induced increase
in the protein expression of MMP-3 and MMP-13 in NP cells
Consistent with the results obtained for the mRNA
expression profiles, the results obtained by ELISA revealed that
stimulation with LPS markedly increased the protein expression of
MMP-3 and MMP-13 in the NP cells. Compared with LPS-stimulated
group, treatment with crocin at 50 and 100 µM significantly
inhibited the LPS-induced increase in the protein expression of
MMP-3 and MMP-13. No signfiicant effects were observed following
pretreatment with crocin at the concentration of 10 µM
(Fig. 3).
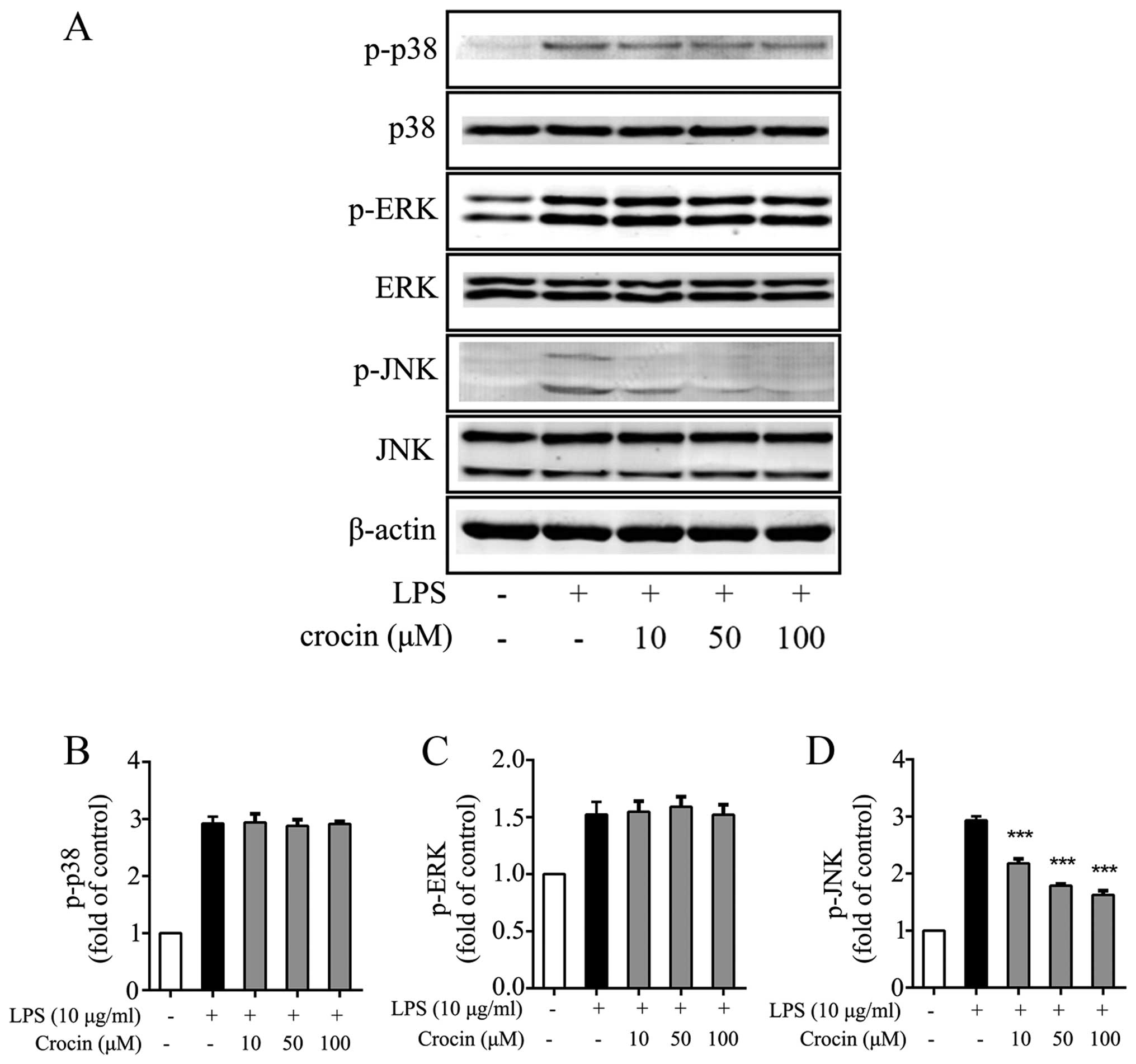
Crocin inhibits the activation of JNK
induced by LPS
Growing evidence supports the involvement of the
MAPK signaling pathway in the inflammation and degeneration of IVDs
(39–41). Hence, the ability of crocin to
inhibit the activation of the MAPK signaling pathway in NP cells
was investigated by western blot analysis. As expected, LPS
significantly induced the phosphorylation of p38, ERK and JNK
(Fig. 4). The phosphorylation of
JNK was potently attenuated by treatment with crocin (Fig. 4A and D), while the levels of p-p38
and p-ERK were not obviously affected (Fig. 4A–C). The results of western blot
analysis demonstrated that crocin inhibited the LPS-induced
activation of the MAPK signaling pathway through JNK, and not p38
or ERK1/2.
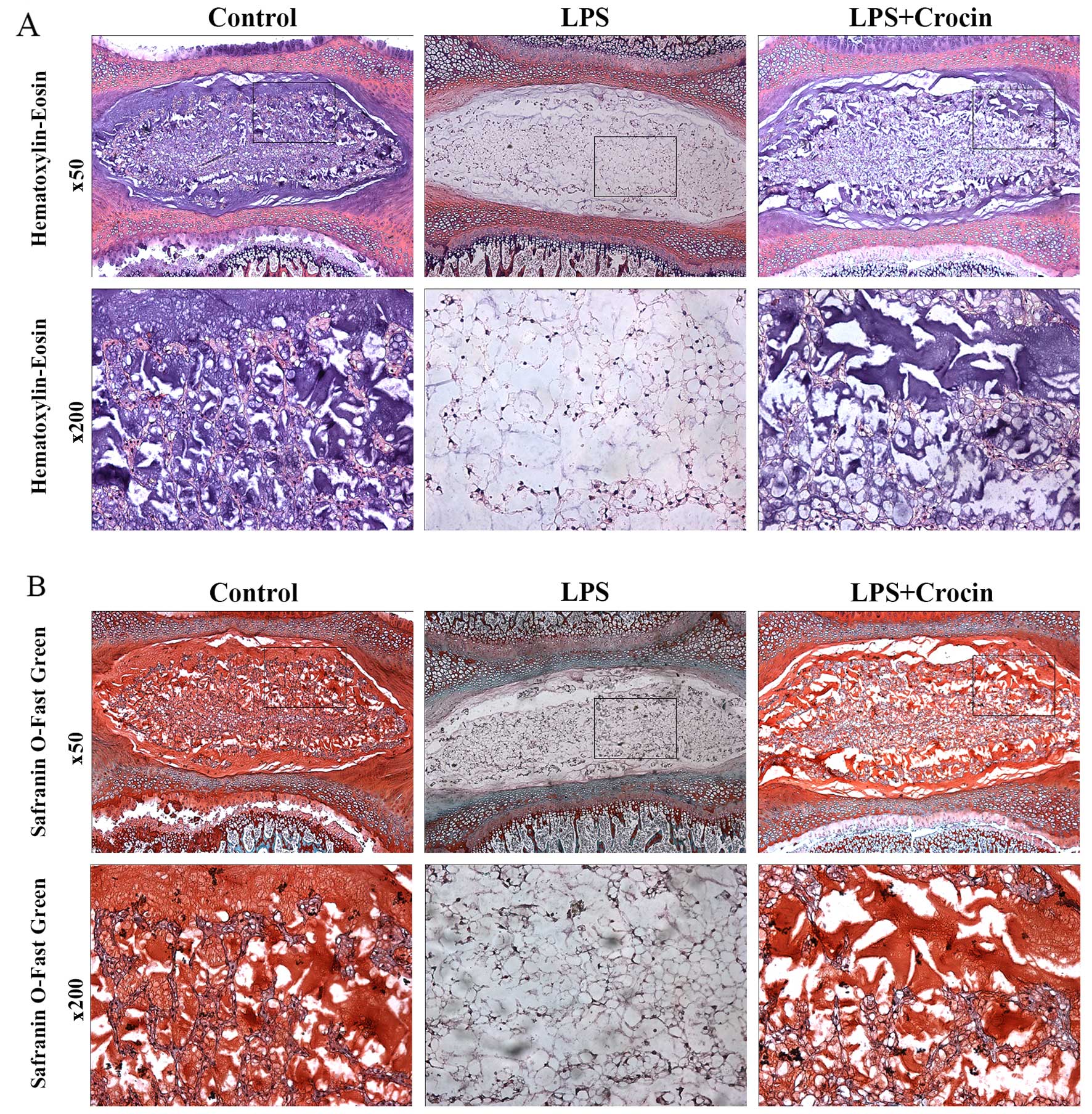
Crocin inhibits the LPS-induced
degeneration of rat IVDs in organ culture
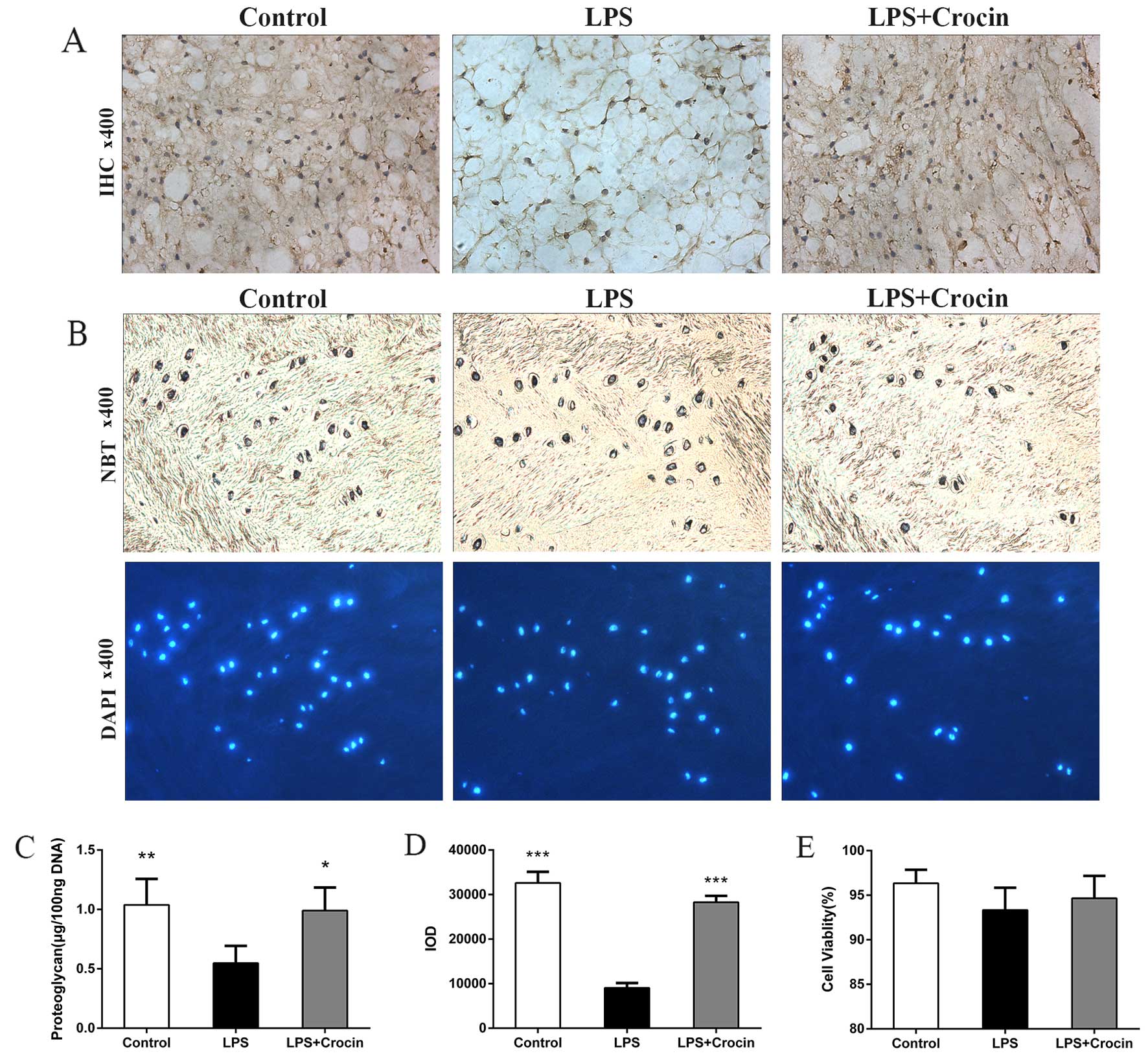
By contrast, as shown in H&E staining images,
treatment with LPS led to decreased staining in the ECM area, with
only NP cells and skeleton-like ECM left, which indicates the
depletion of ECM components and disruption of disc matrix structure
(Fig. 5A). In Safranin O-fast
green staining images, LPS reduced the Safranin O staining
intensity of NP, revealing the dereased content of proteoglycan
(Fig. 5B). Treatment with crocin
attenuated LPS-induced decreased staining of ECM in H&E
staining and Safranin O-fast green staining, maintaining the
integrity of the disc matrix structure (Fig. 5A and B). A DMMB assay was used to
quantify the proteoglycan content in the NP tissue in the cultured
discs. In accordance with the results obtained from histological
analysis, the proteoglycan content in the NP tissue was less in the
LPS group compared with the control and the LPS + crocin groups
(Fig. 6C). Ex vivo
experiments on collagen-II, the other important component of the
ECM, revealed similar results. By performing immunohistochemical
qualitative and quantitative assays, we found that crocin
significantly prevented the loss in the collagen-II content in the
NP tissue from LPS-induced degradation (Fig. 6A and D). These histological
changes demonstrated that crocin significantly inhibited the
LPS-induced catabolism of the disc matrix, thereby protecting the
disc from degeneration. The results of NBT/DAPI dual staining
revealed that cell viability in the discs from all groups,
following 7 days of culture, was >90% with no significant
differences observed among the groups (Fig. 6B and E), which confirms the
reliability of the organ culture condition and excludes the ex
vivo toxicity of crocin.
Discussion
Crocin, the main functional component in saffron
extracts, has been reported to have various bioactivities in
different tissues (42). However,
to the best of our knowledge, there are no studies focusing on the
effects of crocin on IVD. In the present study, we used LPS to
induce IVD inflammation and demonstrated for the first time, to the
very best of our knowledge, that crocin effectively prevented the
LPS-induced upregulation of catabolic enzymes and pro-inflammatory
factors in the NP in vitro and ex vivo. In addition,
the anti-inflammatory bioactivity of crocin was shown to be
mediated by the suppression of JNK activation.
LPS is a potential inflammatory stimulator and it
has been proven to induce the degeneration of the NP (43–47). Evidence indicates that LPS
stimulates the expression of catabolic enzymes, e.g., MMP-1, MMP-3,
MMP-13, ADAMTS-4 and ADAMTS-5 (44–46). MMPs and ADAMTS degrade aggrecan,
the main proteoglycan in the NP, and collagen-II, resulting in the
breakdown of the disc matrix (48–51). However, our results demonstrated
that crocin markedly suppressed the mRNA expression of MMPs and
ADAMTS induced by LPS in a concentration-dependent manner,
revealing the anti-catabolic effects of croin on IVDs.
Similarly, several pro-inflammatory factors, such as
IL-1β, IL-6, TNF-α, and oxidative stress factors, such as iNOS, are
upregulated by LPS in NP cells (45–47). IL-1β is a well-known
pro-inflammatory factor involved in the overexpression of MMPs and
ADAMTS, and it contributes to the loss of the disc matrix (52–54). TNF-α, another important
inflammation mediator, increases the secretion of NO, prostaglandin
E2 (PGE2) and IL-6 in human NP cells
(55). It has been reported that
IL-6 not only acts as an independent pro-inflammatory factor, but
it also amplifies the effects of IL-1 and TNF-α (56). As regards oxidative stress in the
disc, it has been shown that the levels of iNOS, which help to
produce NO, highly correlate with IVD degeneration (57). In the present study, we
demonstrated that crocin exerts inhibitory effects on the
expression of IL-1β, IL-6, TNF-α and iNOS, which was thought to be
associated with the suppressed expression of MMPs and ADAMTS. In
accordance with a previous study (47), we found that LPS decreases the
expression of aggrecan and collagen-II. As was expected, crocin
suppressed the degradation of the ECM components, and partly
reversed the downregulation of aggrecan and collagen-II induced by
LPS in the NP cells.
Another noteworthy finding of the present study was
that suppressive effects of crocin on TLR-2 expression,
antagonizing the effects of LPS. TLRs play a role in early host
defense against exogenous pathogens (58) and cartilage inflammatory diseases
(59). In particular, TLR-2 has
been found to be involved in matrix degradation and in the
suppression of matrix synthesis in articular cartilage (60). In the present study, TLR-2
expression was significantly increased by LPS stimulation and
inhibited by crocin.
As mentioned above, crocin exerted significant
anti-inflammatory and anti-catabolic effects in NP cells. Thus, the
signaling pathway possibly involved was detected by western blot
analysis. The MAPK signaling pathways are actively involved in
inflammatory responses (25).
Thus, in the present study, the phosphorylation of all 3 MAPKs,
including JNK, p38 and ERK, was found to be induced by LPS.
However, the activation of JNK was significantly suppressed by
crocin, shedding light on a possible mechanism behind its
anti-inflammatory effects. Of note, crocin exerted no obvious
inhibitory effects on the phosphorylation levels of p38 and ERK,
which is possibly due to the multiple roles of p38 and ERK in many
other cellular functions, such as cell cycle progression (61), cell growth and differentiation
(62).
To further cofirm the effects of crocin, an ex
vivo experiment was also performed. LPS was used to induce
inflammation and degeneration of the IVDs ex vivo as
previously described (46).
Previous findings and our results prove that the organ culture
condition adopted in the present study allows LPS and crocin, which
permeate through the endplate and annulus fibrosus, to exert
effects on NP in 7 days of culture (46). Histological and biochemical
assessments demonstrated that LPS effectively induced the depletion
of the ECM components in the NP tissue, including proteoglycan and
collagen-II. However, treatment with crocin significantly
attenuated the adverse effects of LPS on ECM in NP tissue, which
was consistent with our results obtained in vitro.
All of these results indicated that crocin may be a
good choice for the treatment of IVD degeneration in the future,
making the prevention of IVD degeneration a possiblity. However,
relevant animal experiments to mimic the actual IVD degeneration
process and to determine the effects of crocin in vivo are
warranted. Importantly, due to the lack of blood supply in the IVD,
the oral or intravenous administration of crocin may not provide a
satisfactory focal drug concentration which is necessary for the
drug effects. Further studies are required to determine appropriate
administration routes or ideal drug carriers.
In conclusion, the present study demonstrated that
crocin potently inhibited LPS-induced inflammation and catabolism
in rat NP cells by suppressing the activation of JNK. Additionally,
organ culture experiments proved that crocin protected rat IVDs
from ECM depletion. In the future, crocin may prove to be an
important candidate for the treatment of IVD degeneration and make
the early treatment of IVD degenerative diseases a possibility in
clinic practice.
Acknowledgments
The authors appreciate the technical support from
Lei Wang, Xiuqiang Liu, Chuan Jiang and Xiuguo Han. This study was
supported by a grant from the National Natural Science Foundation
of China (81272038).
References
|
1
|
Hart LG, Deyo RA and Cherkin DC: Physician
office visits for low back pain. Frequency, clinical evaluation,
and treatment patterns from a U.S. national survey. Spine.
20:11–19. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Katz JN: Lumbar disc disorders and
low-back pain: socioeconomic factors and consequences. J Bone Joint
Surg Am. 88(Suppl 2): 21–24. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Kuslich SD, Ulstrom CL and Michael CJ: The
tissue origin of low back pain and sciatica: a report of pain
response to tissue stimulation during operations on the lumbar
spine using local anesthesia. Orthop Clin North Am. 22:181–187.
1991.PubMed/NCBI
|
|
4
|
Schwarzer AC, Aprill CN, Derby R, Fortin
J, Kine G and Bogduk N: The relative contributions of the disc and
zygapophyseal joint in chronic low back pain. Spine. 19:801–806.
1994. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Nerlich AG, Schleicher ED and Boos N: 1997
Volvo Award winner in basic science studies. Immunohistologic
markers for age-related changes of human lumbar intervertebral
discs. Spine. 22:2781–2795. 1997. View Article : Google Scholar
|
|
6
|
Boos N, Weissbach S, Rohrbach H, Weiler C,
Spratt KF and Nerlich AG: Classification of age-related changes in
lumbar intervertebral discs: 2002 Volvo Award in basic science.
Spine. 27:2631–2644. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Bibby SR, Jones DA, Lee RB, Yu J and Urban
JPG: The pathophysiology of the intervertebral disc. Joint Bone
Spine. 68:537–542. 2001. View Article : Google Scholar
|
|
8
|
Urban JP and Roberts S: Degeneration of
the intervertebral disc. Arthritis Res Ther. 5:120–130. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Roberts S, Evans H, Trivedi J and Menage
J: Histology and pathology of the human intervertebral disc. J Bone
Joint Surg Am. 88(Suppl 2): 10–14. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Le Maitre CL, Pockert A, Buttle DJ,
Freemont AJ and Hoyland JA: Matrix synthesis and degradation in
human intervertebral disc degeneration. Biochem Soc Trans.
35:652–655. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Zhao CQ, Wang LM, Jiang LS and Dai LY: The
cell biology of intervertebral disc aging and degeneration. Ageing
Res Rev. 6:247–261. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Bachmeier BE, Nerlich AG, Weiler C,
Paesold G, Jochum M and Boos N: Analysis of tissue distribution of
TNF-alpha, TNF-alpha-receptors, and the activating
TNF-alpha-converting enzyme suggests activation of the TNF-alpha
system in the aging intervertebral disc. Ann NY Acad Sci.
1096:44–54. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Shamji MF, Setton LA, Jarvis W, So S, Chen
J, Jing L, Bullock R, Isaacs RE, Brown C and Richardson WJ:
Proinflammatory cytokine expression profile in degenerated and
herniated human intervertebral disc tissues. Arthritis Rheum.
62:1974–1982. 2010.PubMed/NCBI
|
|
14
|
Weiler C, Nerlich AG, Bachmeier BE and
Boos N: Expression and distribution of tumor necrosis factor alpha
in human lumbar intervertebral discs: a study in surgical specimen
and autopsy controls. Spine. 30:44–53; discussion 54.
2005.PubMed/NCBI
|
|
15
|
Burke JG, Watson RW, McCormack D, Dowling
FE, Walsh MG and Fitzpatrick JM: Intervertebral discs which cause
low back pain secrete high levels of proinflammatory mediators. J
Bone Joint Surg Br. 84:196–201. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Liu MH, Sun JS, Tsai SW, Sheu SY and Chen
MH: Icariin protects murine chondrocytes from
lipopolysaccharide-induced inflammatory responses and extracellular
matrix degradation. Nutr Res. 30:57–65. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Kim KW, Chung HN, Ha KY, Lee JS and Kim
YY: Senescence mechanisms of nucleus pulposus chondrocytes in human
intervertebral discs. Spine J. 9:658–666. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Huang QQ and Pope RM: The role of
toll-like receptors in rheumatoid arthritis. Curr Rheumatol Rep.
11:357–364. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Abdollahi-Roodsaz S, Joosten LA, Roelofs
MF, Radstake TR, Matera G, Popa C, van der Meer JW, Netea MG and
van den Berg WB: Inhibition of Toll-like receptor 4 breaks the
inflammatory loop in autoimmune destructive arthritis. Arthritis
Rheum. 56:2957–2967. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Bobacz K, Sunk IG, Hofstaetter JG, Amoyo
L, Toma CD, Akira S, Weichhart T, Saemann M and Smolen JS:
Toll-like receptors and chondrocytes: the
lipopolysaccharide-induced decrease in cartilage matrix synthesis
is dependent on the presence of toll-like receptor 4 and
antagonized by bone morphogenetic protein 7. Arthritis Rheum.
56:1880–1893. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Iacono A, Gómez R, Sperry J, Conde J,
Bianco G, Meli R, Gómez-Reino JJ, Smith AB III and Gualillo O:
Effect of oleocanthal and its derivatives on inflammatory response
induced by lipopolysaccharide in a murine chondrocyte cell line.
Arthritis Rheum. 62:1675–1682. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Pålsson-McDermott EM and O'Neill LA:
Signal transduction by the lipopolysaccharide receptor, Toll-like
receptor-4. Immunology. 113:153–162. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Jasin HE: Bacterial lipopolysaccharides
induce in vitro degradation of cartilage matrix through chondrocyte
activation. J Clin Invest. 72:2014–2019. 1983. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Kittlick PD and Engelmann D: Effect of the
microbial constituents, LPS and BCG, on the glycosaminoglycans of
chondrocyte cultures. Exp Pathol. 42:145–150. 1991. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Berenbaum F: Signaling transduction:
target in osteoarthritis. Curr Opin Rheumatol. 16:616–622. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Avruch J: MAP kinase pathways: the first
twenty years. Biochim Biophys Acta. 1773:1150–1160. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Chung JW, Zeng Y and Wong TK: Drug therapy
for the treatment of chronic nonspecific low back pain: systematic
review and meta-analysis. Pain Physician. 16:E685–E704.
2013.PubMed/NCBI
|
|
28
|
Michaleff ZA, Kamper SJ, Maher CG, Evans
R, Broderick C and Henschke N: Low back pain in children and
adolescents: a systematic review and meta-analysis evaluating the
effectiveness of conservative interventions. Eur Spine J.
23:2046–2058. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Phillips FM, Slosar PJ, Youssef JA,
Andersson G and Papatheofanis F: Lumbar spine fusion for chronic
low back pain due to degenerative disc disease: a systematic
review. Spine (Phila Pa 1976). 38:E409–E422. 2013. View Article : Google Scholar
|
|
30
|
Xu GL, Li G, Ma HP, Zhong H, Liu F and Ao
GZ: Preventive effect of crocin in inflamed animals and in
LPS-challenged RAW 264.7 cells. J Agric Food Chem. 57:8325–8330.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Nam KN, Park YM, Jung HJ, Lee JY, Min BD,
Park SU, Jung WS, Cho KH, Park JH, Kang I, et al: Anti-inflammatory
effects of crocin and crocetin in rat brain microglial cells. Eur J
Pharmacol. 648:110–116. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Kim JH, Park GY, Bang SY, Park SY, Bae SK
and Kim Y: Crocin suppresses LPS-stimulated expression of inducible
nitric oxide synthase by upregulation of heme oxygenase-1 via
calcium/calmodulin-dependent protein kinase 4. Mediators Inflamm.
2014(728709)2014. View Article : Google Scholar
|
|
33
|
Ding Q, Zhong H, Qi Y, Cheng Y, Li W, Yan
S and Wang X: Anti-arthritic effects of crocin in
interleukin-1β-treated articular chondrocytes and cartilage in a
rabbit osteoarthritic model. Inflamm Res. 62:17–25. 2013.
View Article : Google Scholar
|
|
34
|
Risbud MV, Izzo MW, Adams CS, Arnold WW,
Hillibrand AS, Vresilovic EJ, Vaccaro AR, Albert TJ and Shapiro IM:
An organ culture system for the study of the nucleus pulposus:
description of the system and evaluation of the cells. Spine.
28:2652–2658; discussion 2658–2659. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Ponnappan RK, Markova DZ, Antonio PJ,
Murray HB, Vaccaro AR, Shapiro IM, Anderson DG, Albert TJ and
Risbud MV: An organ culture system to model early degenerative
changes of the intervertebral disc. Arthritis Res Ther. 13. pp.
R1712011, View
Article : Google Scholar
|
|
36
|
Lim TH, Ramakrishnan PS, Kurriger GL,
Martin JA, Stevens JW, Kim J and Mendoza SA: Rat spinal motion
segment in organ culture: a cell viability study. Spine.
31:1291–1297; discussion 1298. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Chiba K, Andersson GB, Masuda K, Momohara
S, Williams JM and Thonar EJ: A new culture system to study the
metabolism of the intervertebral disc in vitro. Spine.
23:1821–1827; discussion 1828. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Chandrasekhar S, Esterman MA and Hoffman
HA: Microdetermination of proteoglycans and glycosaminoglycans in
the presence of guanidine hydrochloride. Anal Biochem. 161:103–108.
1987. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Studer RK, Aboka AM, Gilbertson LG,
Georgescu H, Sowa G, Vo N and Kang JD: p38 MAPK inhibition in
nucleus pulposus cells: a potential target for treating
intervertebral disc degeneration. Spine (Phila Pa 1976).
32:2827–2833. 2007. View Article : Google Scholar
|
|
40
|
Kim JH, Studer RK, Vo NV, Sowa GA and Kang
JD: p38 MAPK inhibition selectively mitigates inflammatory
mediators and VEGF production in AF cells co-cultured with
activated macrophage-like THP-1 cells. Osteoarthritis Cartilage.
17:1662–1669. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Wuertz K, Vo N, Kletsas D and Boos N:
Inflammatory and catabolic signalling in intervertebral discs: the
roles of NF-κB and MAP kinases. Eur Cell Mater. 23:103–120.
2012.
|
|
42
|
Alavizadeh SH and Hosseinzadeh H:
Bioactivity assessment and toxicity of crocin: A comprehensive
review. Food Chem Toxicol. 64:65–80. 2014. View Article : Google Scholar
|
|
43
|
Aota Y, An HS, Imai Y, Thonar EJ,
Muehleman C and Masuda K: Comparison of cellular response in bovine
intervertebral disc cells and articular chondrocytes: effects of
lipopolysaccharide on proteoglycan metabolism. Cell Tissue Res.
326:787–793. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Ellman MB, Kim JS, An HS, Chen D, Kc R, An
J, Dittakavi T, van Wijnen AJ, Cs-Szabo G, Li X, et al: Toll-like
receptor adaptor signaling molecule MyD88 on intervertebral disk
homeostasis: In vitro, ex vivo studies. Gene. 505:283–290. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Iwata M, Ochi H, Asou Y, Haro H, Aikawa T,
Harada Y, Nezu Y, Yogo T, Tagawa M and Hara Y: Variations in gene
and protein expression in canine chondrodystrophic nucleus pulposus
cells following long-term three-dimensional culture. PLoS One.
8:e631202013. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Kim JS, Ellman MB, Yan D, An HS, Kc R, Li
X, Chen D, Xiao G, Cs-Szabo G, Hoskin DW, et al: Lactoferricin
mediates anti-inflammatory and anti-catabolic effects via
inhibition of IL-1 and LPS activity in the intervertebral disc. J
Cell Physiol. 228:1884–1896. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Rajan NE, Bloom O, Maidhof R, Stetson N,
Sherry B, Levine M and Chahine NO: Toll-Like Receptor 4 (TLR4)
expression and stimulation in a model of intervertebral disc
inflammation and degeneration. Spine. 38:1343–1351. 2013.
View Article : Google Scholar
|
|
48
|
Takaishi H, Kimura T, Dalal S, Okada Y and
D'Armiento J: Joint diseases and matrix metalloproteinases: a role
for MMP-13. Curr Pharm Biotechnol. 9:47–54. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Fosang AJ, Neame PJ, Last K, Hardingham
TE, Murphy G and Hamilton JA: The interglobular domain of cartilage
aggrecan is cleaved by PUMP, gelatinases, and cathepsin B. J Biol
Chem. 267:19470–19474. 1992.PubMed/NCBI
|
|
50
|
Tortorella MD, Burn TC, Pratta MA,
Abbaszade I, Hollis JM, Liu R, Rosenfeld SA, Copeland RA, Decicco
CP, Wynn R, et al: Purification and cloning of aggrecanase-1: a
member of the ADAMTS family of proteins. Science. 284:1664–1666.
1999. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Abbaszade I, Liu RQ, Yang F, Rosenfeld SA,
Ross OH, Link JR, Ellis DM, Tortorella MD, Pratta MA, Hollis JM, et
al: Cloning and characterization of ADAMTS11, an aggrecanase from
the ADAMTS family. J Biol Chem. 274:23443–23450. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Yu ZG, Xu N, Wang WB, Pan SH, Li KS and
Liu JK: Interleukin-1 inhibits Sox9 and collagen type II expression
via nuclear factor-kappaB in the cultured human intervertebral disc
cells. Chin Med J (Engl). 122:2483–2488. 2009.
|
|
53
|
Akyol S, Eraslan BS, Etyemez H, Tanriverdi
T and Hanci M: Catabolic cytokine expressions in patients with
degenerative disc disease. Turk Neurosurg. 20:492–499.
2010.PubMed/NCBI
|
|
54
|
Lee S, Moon CS, Sul D, Lee J, Bae M, Hong
Y, Lee M, Choi S, Derby R, Kim BJ, et al: Comparison of growth
factor and cytokine expression in patients with degenerated disc
disease and herniated nucleus pulposus. Clin Biochem. 42:1504–1511.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Sinclair SM, Shamji MF, Chen J, Jing L,
Richardson WJ, Brown CR, Fitch RD and Setton LA: Attenuation of
inflammatory events in human intervertebral disc cells with a tumor
necrosis factor antagonist. Spine. 36:1190–1196. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Studer RK, Vo N, Sowa G, Ondeck C and Kang
J: Human nucleus pulposus cells react to IL-6: independent actions
and amplification of response to IL-1 and TNF-α. Spine. 36:593–599.
2011. View Article : Google Scholar
|
|
57
|
Furusawa N, Baba H, Miyoshi N, Maezawa Y,
Uchida K, Kokubo Y and Fukuda M: Herniation of cervical
intervertebral disc: immunohistochemical examination and
measurement of nitric oxide production. Spine. 26:1110–1116. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Takeda K and Akira S: Toll-like receptors
in innate immunity. Int Immunol. 17:1–14. 2005. View Article : Google Scholar
|
|
59
|
O'Neill LA and Dinarello CA: The IL-1
receptor/toll-like receptor superfamily: Crucial receptors for
inflammation and host defense. Immunol Today. 21:206–209. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Joosten LA, Koenders MI, Smeets RL,
Heuvelmans-Jacobs M, Helsen MM, Takeda K, Akira S, Lubberts E, van
de Loo FA and van den Berg WB: Toll-like receptor 2 pathway drives
streptococcal cell wall-induced joint inflammation: critical role
of myeloid differentiation factor 88. J Immunol. 171:6145–6153.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Chambard JC, Lefloch R, Pouysségur J and
Lenormand P: ERK implication in cell cycle regulation. Biochim
Biophys Acta. 1773:1299–1310. 2007. View Article : Google Scholar
|
|
62
|
Krishna M and Narang H: The complexity of
mitogen-activated protein kinases (MAPKs) made simple. Cell Mol
Life Sci. 65:3525–3544. 2008. View Article : Google Scholar : PubMed/NCBI
|