Introduction
Breast cancer is the most common solid tumor found
in women, and is the main cause of mortality due to cancer
(1). According to the American
Cancer Society's estimation, over 40,000 patients in the US
diagnosed with breast cancer succumbed to the disease in 2005.
Presently, the treatment of breast cancer includes surgery and
radiation, sometimes supported by adjuvant chemotherapy or hormone
therapy (2). Although major
advances have been made in understanding the pathogenesis of this
disease, therapeutic problems such as the unselective sacrificing
of normal vs. tumor cells persists.
Arsenic trioxide (As2O3) is an
arsenic compound found in nature and has been used as a medicinal
agent for more than 2,400 years for conditions ranging from
infectious diseases to cancer (3).
In the 1970s, researchers at Harbin Medical University discovered
its ability to cure acute promyelocytic leukemia (APL) (4,5).
Since then, our studies and those of other research groups have
demonstrated that As2O3 also inhibits many
solid tumors, including gastric carcinoma and lung cancer. For
example, As2O3 was found to induce apoptosis
through a reactive oxygen species-dependent pathway and the loss of
mitochondrial membrane potential in HeLa cells (6). In human gastric cancer MGC-803 cells,
As2O3 was also found to inhibit cell growth
and to induce cell apoptosis (7).
Similar findings were observed in esophageal carcinoma (8), neuroblastoma (9), prostate and ovarian carcinoma
(10), and breast cancer (11,12)
cells. However, the molecular mechanisms underlying the
As2O3-promoted apoptosis of solid tumor cells
remain unknown.
HERG belongs to the family of voltage-gated
potassium channels ether-a-go-go (EAG), and mutations in this gene
can cause long Q-T syndrome-2 (LQT2) in humans (13). We previously found that
As2O3 prolonged the QT interval and regulated
several ion channels in the guinea pig heart (14). In particular,
As2O3 was reported to downregulate the
protein expression of cardiac potassium channel HERG and to
decrease IKr in guinea pig ventricular myocytes (15). The reduced trafficking of HERG
channels to the cell surface in patients treated with
As2O3 contributed to the induction of QT
prolongation and torsade de pointes (16). Notably, HERG expression was noted
in a variety of tumor cell lines of varied histogenesis, but was
absent from the healthy cells from which the respective tumor cells
were derived (17–19). In our previous study, we showed
that HERG expression facilitates tumor cell proliferation caused by
tumor necrosis factor (TNF) ligand (TNF-α). Cisapride, a specific
blocker for the human HERG, channel was shown to exhibit
therapeutic effects on gastric cancer by inhibiting the growth of
gastric cancer cells through the regulation of the cell cycle and
the induction of apoptosis. Similarly, silencing of HERG protein
expression by siRNA technology was found to decrease HERG currents
and to inhibit proliferation, invasion and tumorigenicity, and to
induce the apoptosis of gastric cancer cells by inhibiting their
entry into the S phase from the G1 phase (20). These findings prompted us to
hypothesize that the HERG channel may be involved in the regulation
of cell death by As2O3 in breast cancer. The
present study was designed to investigate the anti-cancer effect of
As2O3 on human breast cancer MCF-7 cells, and
to examine the role of the HERG channel in this process.
Materials and methods
Materials
Dulbecco's minimal essential medium (DMEM), foetal
bovine serum (FBS), penicillin, streptomycin and other cell culture
reagents were obtained from Gibco (Grand Island, NY, USA). The
terminal deoxynucleotidyl transferase-mediated dUTP nick end
labeling (TUNEL) detection kit was purchased from Roche (Penzberg,
Germany). Apoptosis-FITC was from Bao Sai Company (Beijing, China).
Anti-HERG and anti-glyceraldehyde-3-phosphate dehydrogenase (GAPDH)
antibodies were purchased from Santa Cruz Biotechnology (USA). The
CaspACE™ assay system, a fluorometric detection kit, was obtained
from Promega (USA). As2O3 was from Yida
(Harbin, China).
Cell line and cell culture
The MCF-7 cell line was provided by Dr Wang Zhiguo
of the Montreal Heart Institute, Canada. The cells were maintained
in DMEM supplemented with 10% FBS and 1% penicillin and
streptomycin in a humidified atmosphere with 5% CO2 at
37°C. Cells were passaged regularly and subcultured to ∼80%
confluence before conducting the experimental procedures.
Cell proliferation assay
Cell proliferation was assessed using the MTT assay.
Briefly, cells were treated with various concentrations of
As2O3 for 24 h. Then, 15 μl of MTT reagent
was added to each well. After 4 h of incubation at 37°C, the
supernatants were discarded and the crystals were dissolved in
dimethyl sulfoxide (DMSO). The absorbance was measured at 490
nm.
Fluorescence microscopy measurements
For the detection of apoptosis, cells were stained
with acridine orange/ethidium bromide (AO/EB). The fluorescent dye
AO readily enters either intact cells or cells with damaged
membranes and stains them green. EB, which is impermeable to cells
with preserved membranes, stains cells red. These dyes were used to
detect apoptotic and necrotic cells. For the AO/EB procedure, cells
were harvested with 10 μl of prepared AO/EB working solution (100
μg/ml AO and 100 μg/ml of EB) in phosphate-buffered saline (PBS)
for 5 min and examined under a fluorescence microscope (Eclipse
TE300, Nikon, Japan).
Terminal deoxynucleotidyl
transferase-mediated dUTP nick end labeling (TUNEL) assay
DNA fragmentation of individual cells was detected
in situ by TUNEL with the In Situ Cell Death Detection kit,
Fluorescein. Cells grown on cover-slips were washed with PBS
containing (in mM) NaCl 137.0, KCl 2.7,
Na2HPO4 4.3, KH2PO4 1.4
(pH 7.4), and were fixed in 4% paraformaldehyde solution for 1 h at
4°C. The cells were permeabilized in solution containing 0.1%
Triton X-100 for 2 min on ice, followed by incubation in freshly
prepared TUNEL reaction mixture for 1 h at 37°C in the dark. The
coverslips were then washed with PBS and mounted on slides with
anti-fading solution. TUNEL staining was analyzed using
fluorescence microscopy (Olympus, Tokyo, Japan).
Flow cytometric analysis of apoptotic
progression
Quantitative assessment of apoptosis was conducted
using an Annexin V assay kit. MCF-7 cells were centrifuged at 1000
x g for 10 min at 4°C after trypsinization, then washed with
ice-cold PBS twice. The pellet was resuspended in ice-cold binding
buffer provided in the kit. Subsequently, 10 μl Annexin V-FITC and
5 μl propidium iodide (PI) were added to the cell suspension, which
was maintained on ice in the dark for 15 min. The samples were then
assessed for viable (Annexin V−/PI−), early
apoptotic (Annexin V+/PI−), late apoptotic
(Annexin V+/PI+) and necrotic (Annexin
V−/PI+) cells using a flow cytometer
(Fc500MDL; Beckman Coulter, USA).
Caspase-3 activity assay
Caspase-3 activity was measured using the CaspACE™
assay system, fluorometric kit, according to the manufacturer's
instructions. Briefly, the cells were lysed and the supernatant was
used for the assay. The fluorogenic substrates for caspase-3 were
labeled with fluorochrome 7-amino-methylcoumarin (AMC). AMC was
released from these substrates upon cleavage by caspase-3. Enzyme
activity was determined by monitoring the fluorescence produced by
free AMC using the GloMax™ 20/20n luminometer (Promega)
at 360/460 nm.
Western blot analysis
The immunoblotting procedures were as follows: cells
were incubated at 37°C in DMEM in the presence or absence of
As2O3. After treatment for 24 h, the adherent
cells were scraped off from the culture flask in ice-cold PBS and
centifuged at 2,500 x g for 10 min. RIPA buffer (Beyotime,
Shanghai) containing (in mM) Tris 50.0 (pH 7.4), NaCl 150.0, 1%
NP-40, 0.5% sodium deoxycholate, 0.1% SDS, sodium orthovanadate,
sodium fluoride, EDTA and leupeptin was added to the pellets, and
the cells were homogenized on ice for 45 min. The cells were then
centrifuged at 13,500 x g for 30 min, and the cleared lysates were
used for immunoblotting. Protein concentration was determined
according to the Bradford method (Sigma) using BSA as a standard.
Denatured protein was separated using 10% sodium dodecyl sulfate
polyacrylamide gel electrophoresis (SDS-PAGE), transferred to a
PVDF membrane (Stratagene) and blocked in 5% nonfat milk overnight.
The next day, the membrane was incubated with primary antibodies
against GAPDH (1:1000 dilution) and the specific polyclonal rabbit
anti-HERG antibody (dilution 1:200). Goat anti-rabbit Alexa Fluor
700 (1:4000 dilution, Molecular Probes) was used as a secondary
antibody. The Odyssey infrared fluorescent scanning system (LICOR)
was used to detect the protein bands. The intensity of the band was
determined by densitometry using Odyssey v1.2 software.
Statistical analysis
Data are expressed as the mean ± standard error of
the mean (SEM), with the exception of the 50% inhibitory
concentration (IC50). Statistical differences were
analyzed by the Student's t-test. A two-tailed p<0.05 was
considered statistically significant.
Results
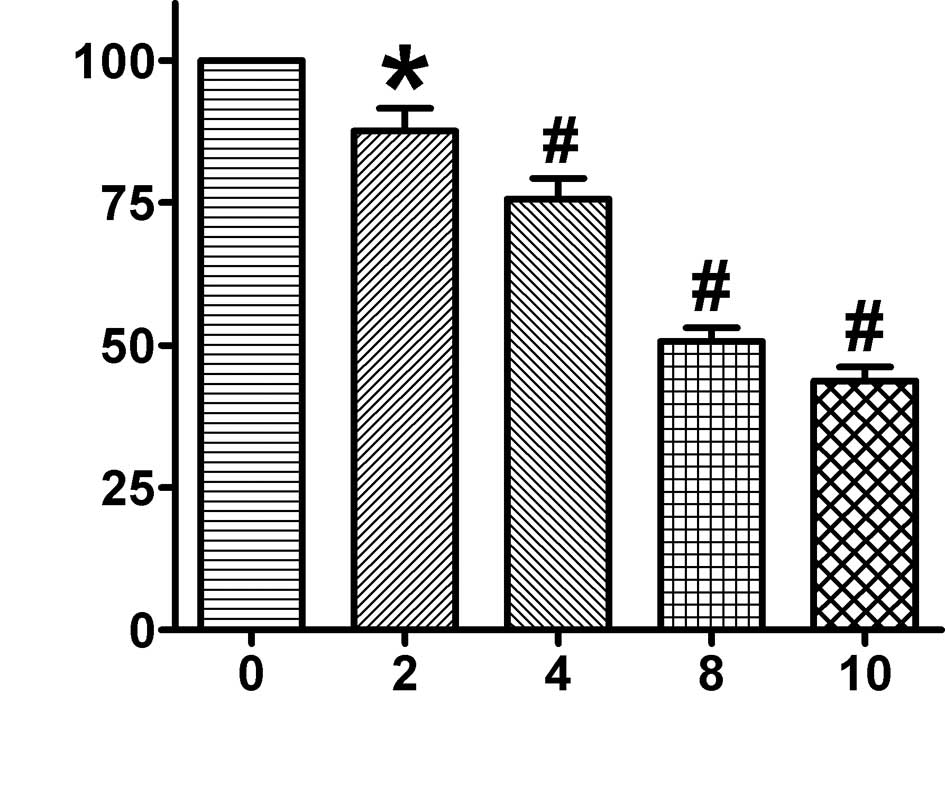
As2O3 reduced the
cell viability of MCF-7 cells
To detect cell viability, the
mitochondrial-dependent reduction of MTT to formazan was measured.
The cells were incubated with 2, 4, 8 and 10 μM
As2O3 for 24 h. As shown in Fig. 1, cell viability was markedly
inhibited with increased As2O3 concentrations
in a dose-dependent manner. As2O3 (2 and 4
μM) treatment of MCF-7 cells resulted in 87.63±4.90 and 75.67±4.46%
reduced cell viability, respectively, whereas 8 and 10 μM
As2O3 resulted in only 50.70±2.84 and
42.41±2.66% reduced cell viability, respectively, after a 24-h
treatment compared to the controls. The IC50 value of
the inhibition of MCF-7 cells by As2O3 was
8.2 μM upon treatment for 24 h.
As2O3 induced the
apoptosis of MCF-7 cells
Since cell viability was decreased by
As2O3, we hypothesized that apoptosis might
be involved in this process. Several assays were used to determine
whether As2O3 induced cellular apoptosis in
the MCF-7 cells. Apoptotic morphological changes in the nuclear
chromatin of cells were detected by AO/EB staining. The cells
treated with As2O3 showed typical apoptotic
morphology, which included condensed nuclei, membrane blebbing and
the formation of apoptotic bodies (Fig. 2B). By contrast, control cells
showed intact nuclear architecture (Fig. 2A). The number of apoptotic cells
was increased by 33.47±2.30% in the
As2O3-treated group compared to the control
group (Fig. 2C).
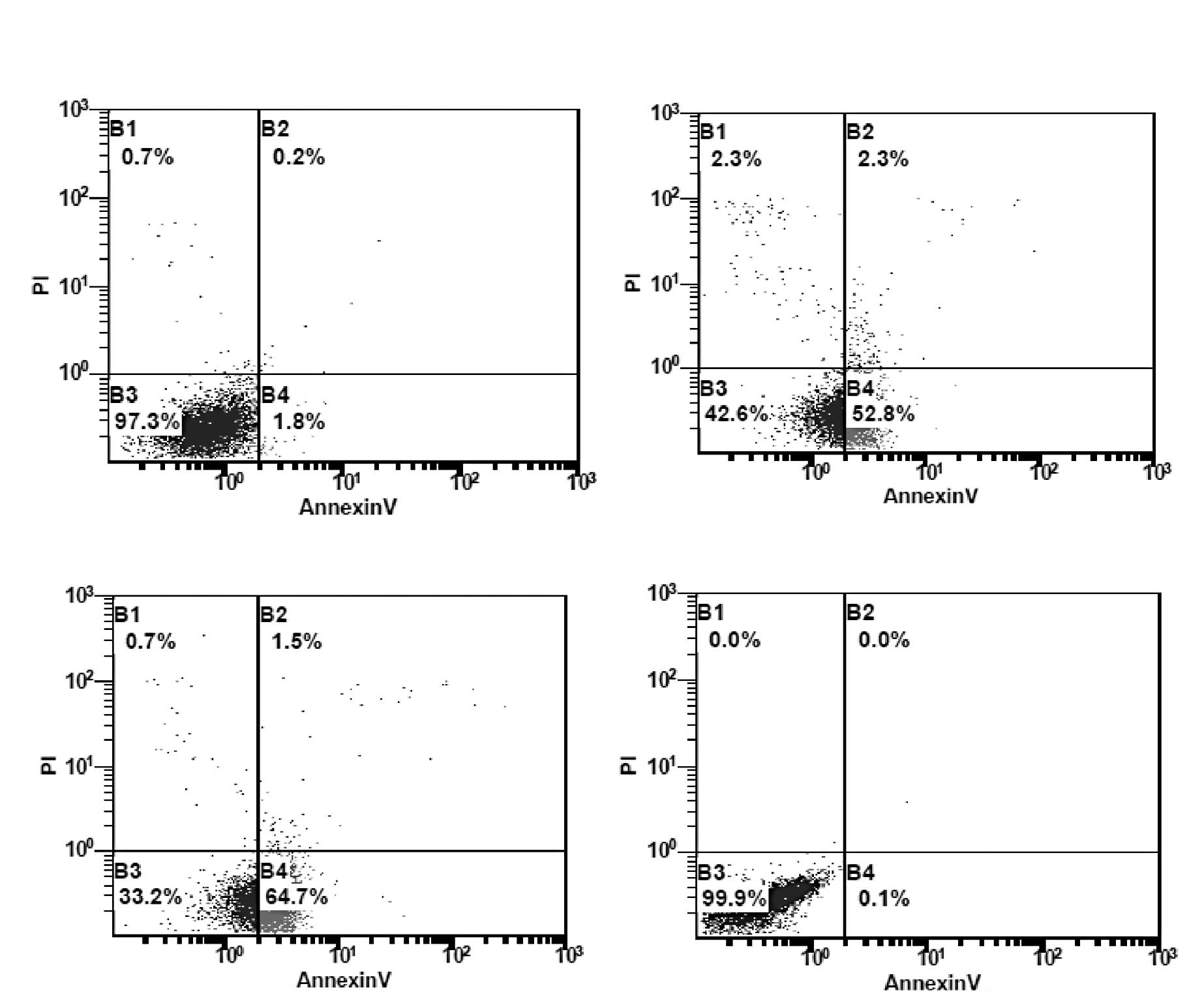
For the quantification of
As2O3-induced apoptotic death, the number of
apoptotic and necrotic cells was measured by flow cytometry with
the Annexin V/PI assay. Apoptotic cells were detected by Annexin V
binding to phospholipid phosphatidylserine (PS), which was
translocated from the inner to the outer leaflet of the plasma
membrane of the apoptotic cells. The cells were treated with 8 or
16 μM As2O3 for 24 h. As shown in Fig. 3, treatment with all concentrations
of As2O3 resulted in a statistically
significant increase in the number of early apoptotic cells. At 8
μM As2O3, the median values for Annexin
V−/PI− (normal) the Annexin
V+/PI− (early apoptotic) cells were
52.13±6.14 and 45.10±5.19%, respectively. With increasing
concentration, the number of normal cells decreased while the
number of early apoptotic cells increased. When the cells were
treated with 16 μM As2O3 for 24 h, the median
values of normal and early apoptotic cells were 39.60±5.28 and
57.43±5.13%, respectively.
Typical apoptotic nuclear condensation is regarded
as the morphological marker of apoptosis. When DNA strands are
cleaved or nicked by nucleases, a large number of 3′-hydroxyl ends
are exposed. The TUNEL assay was performed to detect cells
containing massive DNA fragmentation, a hallmark of late apoptosis.
As shown in Fig. 4,
Tunnel-positive cells were seldom observed in the control MCF-7
cells, whereas in the As2O3-treated MCF-7
cells, the number of Tunnel-positive cells significantly increased
by 29.52±2.83%. These obvious morphologic changes of apoptosis were
markedly induced by treatment with 8 μM
As2O3.
As2O3 induced
apoptosis through activation of caspase-3
Activation of caspase-3 is important in the
initiation of apoptosis in diverse biological processes. In order
to investigate apoptotic signaling, the activities of caspase-3
were examined. As shown in Fig. 5,
when the cells cultured to ∼80% confluence were exposed to 8 μM
As2O3 for 6 h, caspase-3 activity increased
markedly. The activity of caspase-3 increased 55.67±0.12% as
compared to the control group.
Expression of HERG in MCF-7 cells treated
with As2O3
HERG channel was reported to have oncogenic
properties. The distribution is restricted in normal tissue and
becomes ubiquitous in tumor cells. To examine whether HERG is
involved in the cell growth and cell death induced by
As2O3 in the MCF-7 cell line, we used Western
blot analysis to detect the expression of HERG protein. The cells
seeded in culture medium to 80% confluence were treated with
As2O3 at 0 and 8 μM, respectively. As shown
in Fig. 6, the expression of HERG
decreased by 41±0.11% according to the level of non-treated
cells.
Discussion
In the heart, HERG encodes the rapid delayed
rectifier K+ current and undergoes marked developmental
changes, predominating in the fetal heart and dissipating in the
adult (21,22). Most strikingly, HERG is abundantly
expressed in a variety of tumor cells but is not present in the
corresponding healthy cells implicating HERG in the regulation of
tumor cell proliferation. The present study was carried out to
investigate whether As2O3 may induce an
anticancer effect in MCF-7 cells and whether the HERG channel
protein is involved in this process. Our results revealed that i)
As2O3 induced apoptosis in MCF-7 cells and
ii) expression of HERG decreased in the apoptotic process.
Recent studies have shown that
As2O3 is effective in the inhibition of solid
tumors and showed efficacy in a pilot treatment of APL patients
(23). In our study, the
cytotoxicity of As2O3 against human breast
cancer MCF-7 cells was assessed using several parameters. A series
of concentrations of As2O3 ranging from 1 to
100 μM was tested in preliminary experiments. MTT assay showed that
the cell viability was significantly inhibited in a dose-dependent
manner by As2O3. The IC50 was 8.2
μM at 24 h. The concentration of 8 μM As2O3 was chosen
for subsequent experiments on the induction of apoptosis in MCF-7
cells.
Apoptosis, defined as programmed cell death or cell
suicide, is believed to be an important mechanism and a target for
treating APL cell lines and other solid human tumors (24). In our study, apoptosis occurred in
MCF-7 cells upon treatment for over 24 h at a concentration of 8 μM
As2O3. Morphological changes are important
features of cells undergoing apoptosis and are readily observed by
microscopy. The images of AO/EB staining showed specific apoptotic
morphological changes after treatment with
As2O3 for 24 h.
To further confirm the induction of apoptosis by
As2O3, flow cytometric analysis of Annexin
V/PI-stained cells was carried out. Apoptotic cells were detected
by Annexin V binding to phospholipid phosphatidylserine (PS), whose
externalization was observed in the majority of treated cells.
Meanwhile, TUNEL assay identified internucleosomal DNA
fragmentation in apoptotic cells by attachment of a fluorescent
indicator to the ends of fragmented DNA suggesting that decreased
cell viability was due to As2O3-induced cell
apoptosis.
Several action mechanisms involved in
As2O3-induced apoptosis of MCF-7 cells have
been identified, including signal-regulated kinase (ERK), p38 and
c-Jun N-terminal kinase (JNK), p53, Bcl-2 (11,12).
Here, we investigated caspase-3 and the expression of HERG in
As2O3-induced apoptosis of MCF-7 cells.
Caspase-3 is a major executioner protease,
responsible for initiating the apoptotic process (25). The expression of caspase-3 and poly
ADP-ribose polymerase (PARP) which is the specific cleavage of its
downstream substrates was observed in the breast carcinoma cell
line MCF-7 by As2O3 treatment (26). Thus, caspase-3 present in the MCF-7
cells along with this enzyme contributed to the apoptotic signaling
process. Our data demonstrated that As2O3
exposure significantly increased the level of caspase-3 and it was
caspase-3 that mediated this apoptosis.
HERG belongs to an evolutionarily conserved
multigenic family of voltage-gated K+ channels, the
eag (ether a-go-go) family. It is expressed in many
tumor cell lines of different histogenesis but is not present in
the corresponding normal cells, which has highlighted the tight
association between HERG and cancer (27–29).
It is reported that the HERG gene and HERG protein are
expressed with high frequency in primary human endometrial cancers,
as compared to its absence in normal and hyperplastic endometrium
(18). A similar expression
pattern was observed in leukemia where almost all of the primary
leukemia cells and K562 leukemia cell lines expressed HERG mRNA,
while no expression was detected in normal bone marrow cells.
Moreover, inhibition of the HERG channel can reduce leukemia cell
proliferation by affecting the G1/S transition phase of the cell
cycle, while not affecting the growth of cells which do not express
HERG channels (30). HERG was also
expressed at the protein and mRNA levels in MDA-MB-435S melanoma
cells. Blockade of HERG channels and downregulation of HERG by
siRNA can both induce an antiproliferative effect on melanoma cell
lines (31). The same association
was observed in human neuroblastoma SH-SY5Y cell lines. It was
found that silencing of the HERG gene by shRNA suppressed
the cellular growth rate, inhibited cell viability and reduced
colony formation (32).
Accumulating evidence indicates that the HERG channel promotes
tumor cell proliferation. Therefore, inhibition of HERG channel
functions or downregulation of HERG channel expression should
inhibit tumorigenesis. The results of the present study
demonstrated that the expression of HERG decreased as a result of
the As2O3-induced antiproliferative effect.
We speculate that inhibition of HERG contributed to the anticancer
effect of As2O3 in MCF-7 cells. A recent
study demonstrated that HERG is physically linked to β1 integrins
and thereby modulates adhesion-dependent signaling (33). Integrins are known to mediate
numerous signaling pathways that are involved in cell
proliferation, migration, differentiation and anti-apoptotic
functions. The finding of the association between HERG protein and
integrins may provide new clues for further study. The HERG channel
has also been related to tumor cell invasion and neoangiogenesis
apart from its activity in cell proliferation (34–36).
Shao et al found that inhibition of HERG protein expression
reduced the invasiveness of gastric cancer cells (37). A similar finding was also reported
in colon cancer, but the mechanism was not clear (38). Moreover, blocking of the HERG
channel significantly impaired VEGF secretion in HERG-expressing
glioblastoma cells (39).
In conclusion, As2O3 induced
apoptosis in MCF-7 cells through the activation of caspase-3 and
downregulation of HERG protein. Because of the specific expression
variation of HERG protein in tumor cells, the HERG channel is
endowed with therapeutic potential.
Acknowledgements
The present study was supported in
part by the National Basic Research Program of China [973 Program,
2007CB512000 (2007CB512006)], the Scientific Research Fund of
Heilongjiang Provincial Education Department (no. 11531216), and
the Scientific Research Fund of Heilongjiang Provincial Health
Department (no. 2007-481).
References
|
1.
|
Sledge GW Jr: Breast cancer as a world
challenge. Clin Breast Cancer. 5:112004. View Article : Google Scholar : PubMed/NCBI
|
|
2.
|
Bange J, Zwick E and Ullrich A: Molecular
targets for breast cancer therapy and prevention. Nat Med.
7:548–552. 2001. View
Article : Google Scholar : PubMed/NCBI
|
|
3.
|
Vernhet L, Allain N, Le Vee M, Morel F,
Guillouzo A and Fardel O: Blockage of multidrug
resistance-associated proteins potentiates the inhibitory effects
of arsenic trioxide on CYP1A1 induction by polycyclic aromatic
hydrocarbons. J Pharmacol Exp Ther. 304:145–155. 2003. View Article : Google Scholar
|
|
4.
|
Cyranoski D: Arsenic patent keeps drug for
rare cancer out of reach of many. Nat Med. 13:10052007. View Article : Google Scholar : PubMed/NCBI
|
|
5.
|
Sun HD, Ma L, Hu XC and Zhang TD: Ai-Lin I
treated 32 cases of acute promyelocytic leukemia. Chin J Integr
Chin West Med. 12:170–171. 1992.
|
|
6.
|
Woo SH, Park IC, Park MJ, et al: Arsenic
trioxide induces apoptosis through a reactive oxygen
species-dependent pathway and loss of mitochondrial membrane
potential in HeLa cells. Int J Oncol. 21:57–63. 2002.
|
|
7.
|
Zhang TC, Cao EH, Li JF, Ma W and Qin JF:
Induction of apoptosis and inhibition of human gastric cancer MG803
cell growth by arsenic trioxide. Eur J Cancer. 35:1258–1263. 1999.
View Article : Google Scholar : PubMed/NCBI
|
|
8.
|
Shen ZY, Shen J, Cai WJ, Hong C and Zheng
MH: The alteration of mitochondria is an early event of arsenic
trioxide-induced apoptosis in esophageal carcinoma cells. Int J Mol
Med. 5:155–158. 2000.PubMed/NCBI
|
|
9.
|
Akao Y, Nakagawa Y and Akiyama K: Arsenic
trioxide induces apoptosis in neuroblastoma cell lines through the
activation of caspase 3 in vitro. FEBS Lett. 455:59–62.
1999. View Article : Google Scholar : PubMed/NCBI
|
|
10.
|
Uslu R, Sanli UA, Sezgin C, et al: Arsenic
trioxide-mediated cytotoxicity and apoptosis in prostate and
ovarian carcinoma cell lines. Clin Cancer Res. 6:4957–4964.
2000.PubMed/NCBI
|
|
11.
|
Chow SK, Chan JY and Fung KP: Inhibition
of cell proliferation and the action mechanisms of arsenic trioxide
As2O3 on human breast cancer cells. J Cell
Biochem. 93:173–187. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
12.
|
Ye J, Li A, Liu Q, Wang X and Zhou J:
Inhibition of mitogen-activated protein kinase kinase enhances
apoptosis induced by arsenic trioxide in human breast cancer MCF-7
cells. Clin Exp Pharmacol Physiol. 32:1042–1048. 2005. View Article : Google Scholar
|
|
13.
|
Wang H, Zhang Y, Cao L, et al: HERG
K+ channel, a regulator of tumor cell apoptosis and
proliferation. Cancer Res. 62:4843–4848. 2002.PubMed/NCBI
|
|
14.
|
Sun HL, Chu WF, Dong DL, et al:
Choline-modulated arsenic trioxide-induced prolongation of cardiac
repolarization in Guinea pig. Basic Clin Pharmacol Toxicol.
98:381–388. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
15.
|
Ficker E, Kuryshev YA, Dennis AT, et al:
Mechanisms of arsenic-induced prolongation of cardiac
repolarization. Mol Pharmacol. 66:33–44. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
16.
|
Vandenberg JI, Walker BD and Campbell TJ:
HERG K+ channels: friend and foe. Trends Pharmacol Sci.
22:240–246. 2001.
|
|
17.
|
Bianchi L, Wible B, Arcangeli A, et al:
herg encodes a K+ current highly conserved in tumors of
different histogenesis: a selective advantage for cancer cells?
Cancer Res. 58:815–822. 1998.
|
|
18.
|
Cherubini A, Taddei GL, Crociani O, et al:
HERG potassium channels are more frequently expressed in human
endometrial cancer as compared to non-cancerous endometrium. Br J
Cancer. 83:1722–1729. 2000. View Article : Google Scholar
|
|
19.
|
Meyer R and Heinemann SH: Characterization
of an eag-like potassium channel in human neuroblastoma cells. J
Physiol. 508:49–56. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
20.
|
Shao XD, Wu KC, Hao ZM, Hong L, Zhang J
and Fan DM: The potent inhibitory effects of cisapride, a specific
blocker for human ether-a-go-go-related gene (HERG) channel, on
gastric cancer cells. Cancer Biol Ther. 4:295–301. 2005. View Article : Google Scholar
|
|
21.
|
Wang L, Feng ZP, Kondo CS, Sheldon RS and
Duff HJ: Developmental changes in the delayed rectifier
K+channels in mouse heart. Circ Res. 79:79–85. 1996.
View Article : Google Scholar : PubMed/NCBI
|
|
22.
|
Wang L and Duff HJ: Identification and
characteristics of delayed rectifier K+ current in fetal
mouse ventricular myocytes. Am J Physiol. 270:H2088–H2093.
1996.PubMed/NCBI
|
|
23.
|
Liu P and Han ZC: Treatment of acute
promyelocytic leukemia and other hematologic malignancies with
arsenic trioxide: review of clinical and basic studies. Int J
Hematol. 78:32–39. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
24.
|
Han H, Long H, Wang H, Wang J, Zhang Y and
Wang Z: Progressive apoptotic cell death triggered by transient
oxidative insult in H9c2 rat ventricular cells: a novel pattern of
apoptosis and the mechanisms. Am J Physiol Heart Circ Physiol.
286:H2169–H2182. 2004. View Article : Google Scholar
|
|
25.
|
Roy S: Caspase at the heart of the
apoptotic cell death pathway. Chem Res Toxicol. 13:961–962. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
26.
|
Li X, Ding X and Adrian TE: Arsenic
trioxide causes redistribution of cell cycle, caspase activation,
and GADD expression in human colonic, breast, and pancreatic cancer
cells. Cancer Invest. 22:389–400. 2004. View Article : Google Scholar
|
|
27.
|
Crociani O, Guasti L, Balzi M, et al: Cell
cycle-dependent expression of HERG1 and HERG1B isoforms in tumor
cells. J Biol Chem. 278:2947–2955. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
28.
|
Guasti L, Crociani O, Redaelli E, et al:
Identification of a post-translational mechanism for the regulation
of hERG1 K+ channel expression and hERG1 current density
in tumor cells. Mol Cell Biol. 28:5043–5060. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
29.
|
Arcangeli A: Expression and role of hERG
channels in cancer cells. Novartis Found Symp. 266:225–232. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
30.
|
Li H, Liu L, Guo L, et al: HERG
K+ channel expression in
CD34+/CD38−/CD123 (high) cells and primary
leukemia cells and analysis of its regulation in leukemia cells.
Int J Hematol. 87:387–392. 2008.
|
|
31.
|
Afrasiabi E, Hietamäki M, Viitanen T,
Sukumaran P, Bergelin N and Törnquist K: Expression and
significance of HERG (KCNH2) potassium channels in the regulation
of MDA-MB-435S melanoma cell proliferation and migration. Cell
Signal. 22:57–64. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
32.
|
Zhao J, Wei XL, Jia YS and Zheng JQ:
Silencing of herg gene by shRNA inhibits SH-SY5Y cell growth in
vitro and in vivo. Eur J Pharmacol. 579:50–57. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
33.
|
Cherubini A, Hofmann G, Pillozzi S, Guasti
L, Crociani O, Cilia E, et al: Human ether-a-go-go-related gene 1
channels are physically linked to beta1 integrins and modulate
adhesion-dependent signaling. Mol Biol Cell. 16:2972–2983. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
34.
|
Bauer CK, Wulfsen I, Schafer R, Glassmeier
G, Wimmers S and Flitsch J: HERG K(+) currents in human
prolactin-secreting adenoma cells. Pflugers Arch. 445:589–600.
2003.
|
|
35.
|
Gullo F, Ales E, Rosati B, et al: ERG
K+ channel blockade enhances firing and epinephrine
secretion in rat chromaffin cells: the missing link to LQT2-related
sudden death? FASEB J. 17:330–332. 2003.PubMed/NCBI
|
|
36.
|
Rosati B, Marchetti P, Crociani O, et al:
Glucose- and arginine-induced insulin secretion by human pancreatic
beta-cells: the role of HERG K(+) channels in firing and release.
FASEB J. 14:2601–2610. 2000.PubMed/NCBI
|
|
37.
|
Shao XD, Wu KC, Guo XZ, Xie MJ, Zhang J
and Fan DM: Expression and significance of HERG protein in gastric
cancer. Cancer Biol Ther. 7:45–50. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
38.
|
Lastraioli E, Guasti L, Crociani O, et al:
herg1 gene and HERG1 protein are overexpressed in colorectal
cancers and regulate cell invasion of tumor cells. Cancer Res.
64:606–611. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
39.
|
Masi A, Becchetti A, Restano-Cassulini R,
et al: hERG1 channels are overexpressed in glioblastoma multiforme
and modulate VEGF secretion in glioblastoma cell lines. Br J
Cancer. 93:781–792. 2005. View Article : Google Scholar : PubMed/NCBI
|