Introduction
Cancer cachexia occurs in the majority of cancer
patients before death, and it is responsible for the death of 22%
of cancer patients (1).
Abnormalities associated with cancer cachexia include anorexia,
weight loss, muscle loss and atrophy, anemia and alterations in
carbohydrate, lipid and protein metabolism (2,3). The
degree of cachexia is inversely correlated with the survival time
of the patient and is always indicative of a poor patient prognosis
(4–6). Perhaps one of the most relevant
characteristics of cachexia is that of asthenia, which reflects the
significant muscle wasting that occurs in the cachectic cancer
patient (7). Deplection of lean
body mass is one of the main consequences of cachexia which
involves not only skeletal muscle but also affects cardiac
proteins, resulting in alterations in heart performance. In
addition to the increased muscle protein degradation found during
cancer growth, the presence of the tumor also induces an increased
rate of DNA fragmentation in skeletal muscle in both rats and mice
(8).
β2-adrenergic agonists are potent muscle
growth promoters in many animal species (9,10).
Treatment with β2-adrenergic agonists results in
skeletal muscle hypertrophy (11–14),
while they cause a reduction in the body fat content (15,16).
Formoterol, one of these compounds, is a highly potent
β2-adrenoceptor-selective agonist which combines the
clinical advantages of rapid onset of action with duration of
action. This compound is currently in use in humans for the
treatment of bronchospasm associated with asthma. In vitro,
formoterol is a potent airway smooth muscle relaxant with high
efficacy and high affinity and selectivity for the
β2-adrenoceptor (17).
Moreover, formoterol relaxes bronchial smooth muscle and also
provides important clinical benefits in symptomatic patients with
chronic obstructive pulmonary disease (18).
Previous studies carried out in our laboratory
demonstrated that formoterol treatment in tumor-bearing animals
resulted in an amelioration of muscle loss through different
mechanisms that include muscle apoptosis and proteolysis (19). In light of these findings, the aim
of the present investigation was to determine the influence of the
cachectic state on the physical performance of rats, and to assess
whether the β2-adrenergic agonist formoterol is
associated with an improvement in physiological parameters such as
grip force and total physical activity.
Materials and methods
Animals
Male Wistar rats (Interfauna, Barcelona, Spain), 5
weeks of age, were used in the different experiments. The animals
were maintained at 22±2°C under a regular light-dark cycle (lights
on from 08:00 a.m. to 08:00 p.m.) and had free access to food and
water. The food intake was measured daily. All animal manipulations
were carried out in accordance with the European Commission
guidelines for the use of laboratory animals.
Tumor inoculation
Rats were divided into two groups: controls and
tumor-bearing hosts. The latter received an intraperitoneal
inoculum of 108 Yoshida ascites AH-130 hepatoma cells
obtained from cells exhibiting exponential growth as previously
described (20). Both groups were
further divided into treated and untreated groups, the former being
administered a daily subcutaneous (s.c.) dose of formoterol [0.3
mg/kg body weight (bw) dissolved in physiological saline solution],
and the latter a corresponding volume of solvent. On day 7 after
tumor transplantation, the animals were weighed and anesthetized
with an intraperitoneal injection (i.p.) of ketamine/xylazine
mixture (3:1) (Imalgene® and Rompun®,
respectively). Each tumor was harvested from the peritoneal cavity,
and the volume and cellularity were evaluated. Tissues were rapidly
excised, weighed and frozen in liquid nitrogen.
Total physical activity
Total physical activity was determined for 7 days in
the control and tumor-bearing animals (non-treated and treated
rats) using activity sensors (IR Actimeter System and Actitrak
software from Panlab, Barcelona, Spain) that translate individual
changes in an infrared pattern caused by movements of the animals
into arbitrary activity counts. Data were collected for a total
period of 24 h. In order to carry out the measurements, animals
remained in their home cage, and a frame containing an infrared
beam system was placed on the outside of the cage. This minimized
stress to the animals.
Grip force assessement
Skeletal muscular strength in rats was quantified by
the grip-strength test as previously described (21,22).
The grip-strength device (Panlab-Harvard Apparatus) comprised a
triangular pull bar connected to an isometric force transducer
(dynamometer). In brief, the grip strength meter was positioned
horizontally, and the rats were held by the tail and lowered
towards the device. The animals were allowed to grasp the
triangular pull bar and were then pulled backwards in a horizontal
plane. The force applied to the bar just before the grip was lost
was recorded as the peak tension. At least three measurements were
taken per rat at baseline and on test days, and the results were
averaged for analysis. This force was measured in grams.
Statistical analysis
Statistical analysis of the data was performed by
means of the Student’s t-test.
Results and Discussion
Implantation of the tumors resulted in a significant
decrease in food intake (26%) of the rats (Table I). This was not reversed upon
formoterol treatment, repudiating any possible implication of the
β2-agonist in the reversal of cancer-induced
anorexia.
 | Table I.Food intake, body weight and muscle
weight in tumor-bearing rats. |
Table I.
Food intake, body weight and muscle
weight in tumor-bearing rats.
| C | C+F | T | T+F |
|---|
| Food intake | 106±2.0 | 112±2.5 | 78±4.4a | 85±2.1a |
| Body weight | | | | |
| Initial | 128±2.4 | 122±3.5 | 127±4.4 | 125±2.2 |
| Final | 164±4.3 | 159±3.5 | 123±7.2a | 126±2.8a |
| Difference | 36±2.2 | 37±0.9 | −4.1±4.8a | 1.6±2.0a |
| Muscle weight | | | | |
| Gastrocnemius | 663±16 | 725±20d | 545±8.5a |
627±9.6a.f |
| Tibialis | 215±5.7 | 238±3.8e | 188±3.6b |
208±4.8a.d |
| EDL | 51±1.9 | 60±2.5d | 41±2.0c | 48±1.3b |
| Diaphragm | 279±14 | 362±14f | 117±11a |
131±7.3a.d |
| Heart | 412±16 | 493±14e | 318±8.1a |
367±8.8a.e |
| Carcass weight | 90±1.8 | 91±2.5 | 72±1.1a | 78±0.7a.f |
| Tumor cell
content | - | - | 3638±179 | 3752±558 |
Seven days after tumor inoculation, a clear decrease
in body weight associated with a significant decrease in muscle
weight was noted (Table I). The
decrease in body weight was attenuated by formoterol treatment; in
fact, formoterol treatment resulted in significant increases in
muscle weight in the tumor-bearing rats (Table I). This effect was observed in the
gastrocnemius, tibialis and extensor digitorum longus (EDL) muscles
and also in the heart. Similar results were previously described by
our research group (19,23). Indeed, formoterol and other
β2-agonists such as clenbuterol were found to be
effective in ameliorating muscle weight loss during wasting
(19,23,24).
At the biochemical level, the mechanisms underlying
the effects of the β2-agonist are complex. Formoterol
was found to decrease protein degradation in skeletal muscle by
inhibiting the ubiquitin-proteasome pathway (19). In addition, formoterol was found to
decrease the enhanced apoptosis observed in skeletal muscle during
cancer cachexia (19). Thirdly, at
least in vitro formoterol increased protein synthesis in
skeletal muscle (19). Notably,
these effects of the β2-agonist appear to be associated
with an increased muscle regeneration capacity (25).
In spite of these previously demonstrated positive
effects of the β2-agonist at the biochemical level, no
measures of physical performance associated with formoterol
treatment during cancer cachexia have been reported. Therefore, the
aim of the present investigation was to assess whether formoterol,
in addition to improving physical and biochemical parameters in an
experimental model of cancer cachexia, also affects various factors
involved in improving quality of life such as total physical
activity and muscle force. In fact, previous investigations with
β2-agonists and muscle strength have lead to
controversial results. Lanigan et al assessed limb muscle
strength and endurance following administration of
β2-agonists and found no beneficial effects on muscle
performance (26). Conversely,
Signorile et al reported that, at least in patients with
muscular atrophy following spinal cord injury,
β2-adrenergic agonist treatment resulted in an
improvement in muscle strength (27).
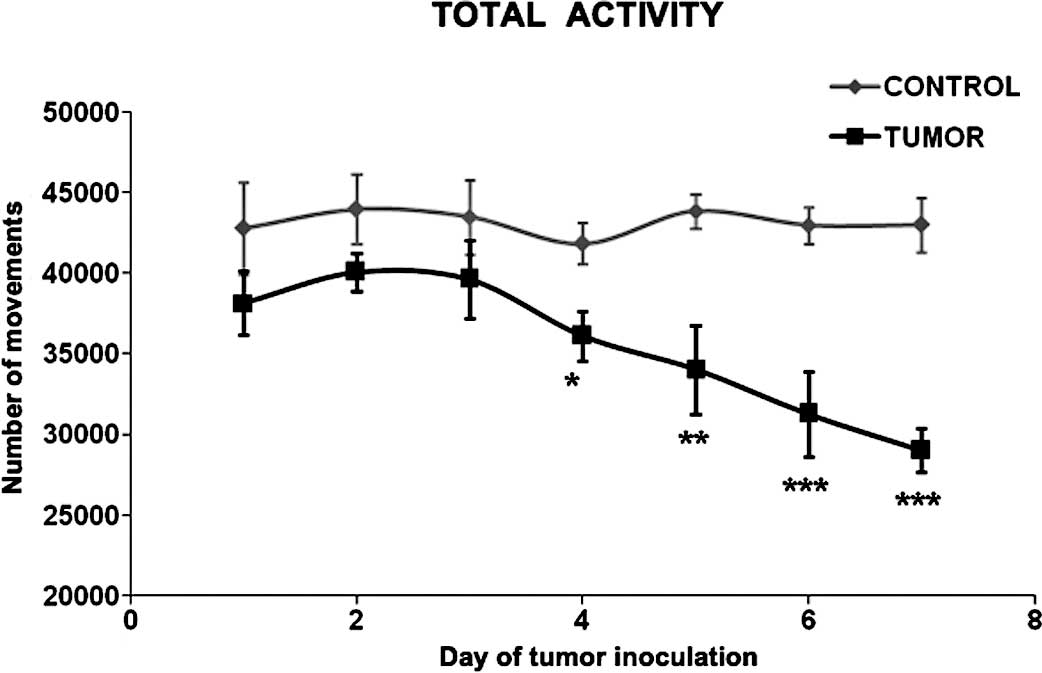
In the present study, tumor burden significantly
affected the total physical activity in the rats bearing the
Yoshida AH-130 ascites hepatoma cell tumors (Fig. 1). As early as 4 days after tumor
implantation – at which point body and muscle weight loss are
already apparent (28) – a
significant decrease in physical activity was observed. The
decrease continued up until day 7 after tumor inoculation. Similar
results have been previously reported using the same tumor model
(29). Tumor burden causes a
reduction in total physical activity through the activation of
muscle wasting either via the release of tumor factors (30) or alternatively through changes in
circulating and tissular cytokines or cytokine receptors (31,32).
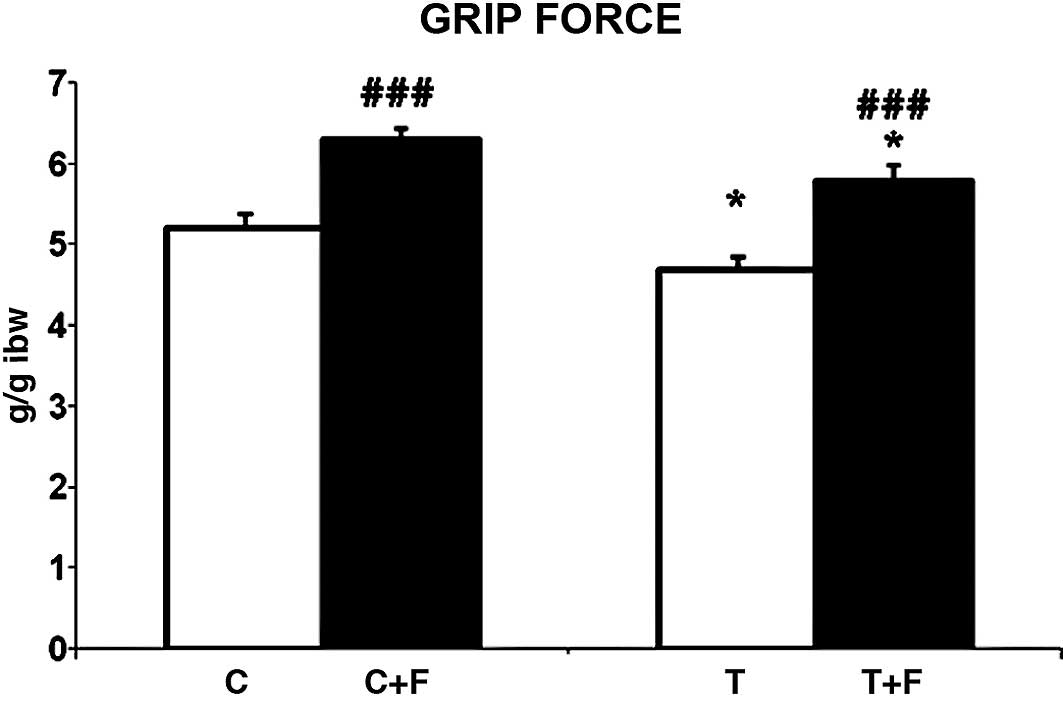
We demonstrated that formoterol treatment
significantly improved grip force in the tumor-bearing rats (23%)
(Fig. 2). This correlated with an
increase in muscle weight as shown in Table I. Therefore, the
β2-agonist clearly acts at the biochemical level, and
its action is reflected in a physiological parameter, grip force,
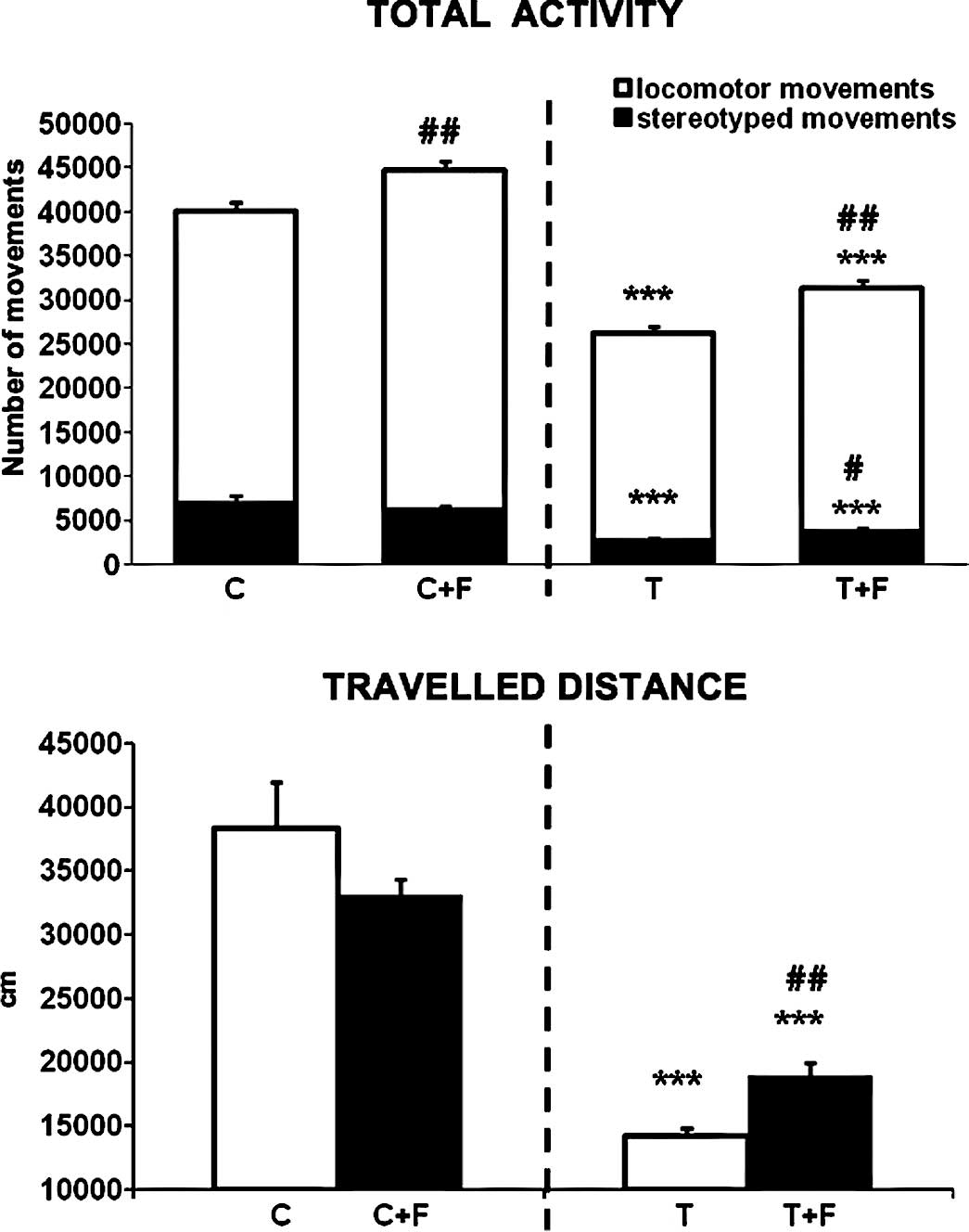
in this case. Notably, formoterol also improved the physical
performance of the animals. Total physical activity and total
distance travelled by the rats were significantly increased by
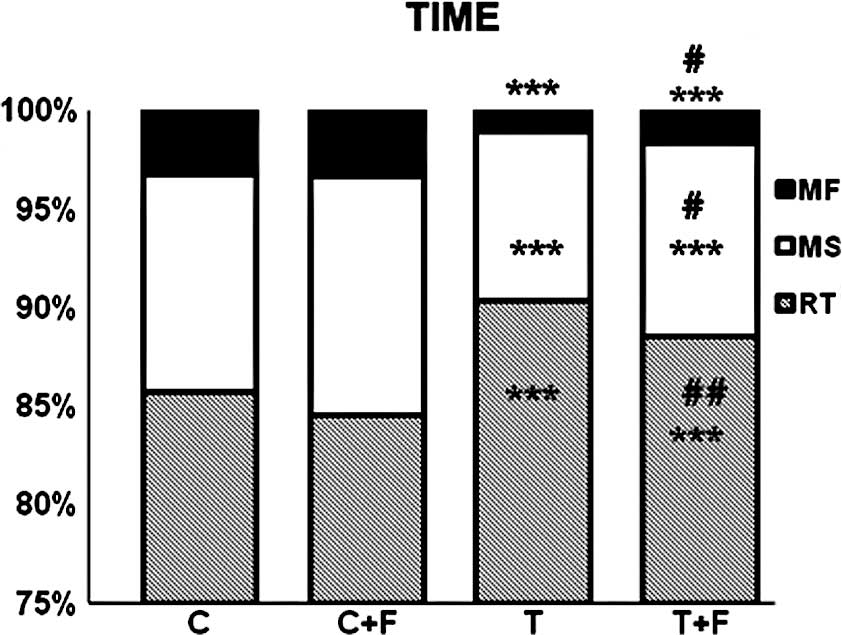
treatment with formoterol (19 and 33% respectively) (Fig. 3). Moreover, resting time, which was
increased in the tumor-bearing rats, was decreased by formoterol
treatment. Conversely, slow and fast movement times, which
decreased in the tumor-bearing rats, increased in the
formoterol-treated rats (Fig.
4).
Collectively, the results presented here allow us to
conclude that the treatment of tumor-bearing animals with the
β2-agonist formoterol clearly resulted in an improvement
in both muscle force and total physical performance. This, together
with previous results obtained by our research group (19), clearly indicate that formoterol may
be a good candidate drug for the treatment of muscle wasting
associated with cancer cachexia. Further preclinical studies are
therefore warranted.
Acknowledgements
This study was supported by grants
from the Ministerio de Ciencia y Tecnología (SAF-02284-2008). The
authors would like to thank Industriale Chimica s.r.l. (Saronno,
Italy), which kindly provided micronized formoterol fumarate. Dr
Roberto Serpe was supported by grant CRP1_296 from the Regione
Autonoma della Sardegna by PO Sardegna FSE 2007-2013 (L.R.7/2007)
titled “Promotion of Scientific and Technological Research in
Sardinia”, Italy.
References
|
1.
|
Warren S: The immediate causes of death in
cancer. Am J Med Sci. 184:610–615. 1932. View Article : Google Scholar
|
|
2.
|
Argiles JM, Alvarez B and Lopez-Soriano
FJ: The metabolic basis of cancer cachexia. Med Res Rev.
17:477–498. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
3.
|
Argiles JM and Lopez-Soriano FJ: Why do
cancer cells have such a high glycolytic rate? Med Hypotheses.
32:151–155. 1990. View Article : Google Scholar : PubMed/NCBI
|
|
4.
|
Harvey KB, Bothe A Jr and Blackburn GL:
Nutritional assessment and patient outcome during oncological
therapy. Cancer. 43:2065–2069. 1979. View Article : Google Scholar : PubMed/NCBI
|
|
5.
|
Nixon DW, Heymsfield SB, Cohen AE, et al:
Protein-calorie undernutrition in hospitalized cancer patients. Am
J Med. 68:683–690. 1980. View Article : Google Scholar : PubMed/NCBI
|
|
6.
|
DeWys W: Management of cancer cachexia.
Semin Oncol. 12:452–460. 1985.
|
|
7.
|
Argiles JM, Garcia-Martinez C, Llovera M
and Lopez-Soriano FJ: The role of cytokines in muscle wasting: its
relation with cancer cachexia. Med Res Rev. 12:637–652. 1992.
View Article : Google Scholar : PubMed/NCBI
|
|
8.
|
Van Royen M, Carbo N, Busquets S, et al:
DNA fragmentation occurs in skeletal muscle during tumor growth: a
link with cancer cachexia? Biochem Biophys Res Commun. 270:533–537.
2000.PubMed/NCBI
|
|
9.
|
Kim YS and Sainz RD: Beta-adrenergic
agonists and hypertrophy of skeletal muscles. Life Sci. 50:397–407.
1992. View Article : Google Scholar : PubMed/NCBI
|
|
10.
|
Stock MJ and Rothwell NJ: Effects of
beta-adrenergic agonists on metabolism and body composition.
Control and Manipulation of Animal Growth. Buttery PJ, Hayes NB and
Lindsay DB: Butterworths; London: pp. 249–257. 1985
|
|
11.
|
Agbenyega ET and Wareham AC: Effect of
clenbuterol on skeletal muscle atrophy in mice induced by the
glucocorticoid dexamethasone. Comp Biochem Physiol Comp Physiol.
102:141–145. 1992. View Article : Google Scholar : PubMed/NCBI
|
|
12.
|
Rajab P, Fox J, Riaz S, Tomlinson D, Ball
D and Greenhaff PL: Skeletal muscle myosin heavy chain isoforms and
energy metabolism after clenbuterol treatment in the rat. Am J
Physiol Regul Integr Comp Physiol. 279:R1076–R1081. 2000.PubMed/NCBI
|
|
13.
|
Hinkle RT, Hodge KM, Cody DB, Sheldon RJ,
Kobilka BK and Isfort RJ: Skeletal muscle hypertrophy and
anti-atrophy effects of clenbuterol are mediated by the
beta2-adrenergic receptor. Muscle Nerve. 25:729–734. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
14.
|
Wineski LE, von Deutsch DA, Abukhalaf IK,
Pitts SA, Potter DE and Paulsen DF: Muscle-specific effects of
hindlimb suspension and clenbuterol in mature male rats. Cells
Tissues Organs. 171:188–198. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
15.
|
Yang YT and McElligott MA: Multiple
actions of beta-adrenergic agonists on skeletal muscle and adipose
tissue. Biochem J. 261:1–10. 1989.PubMed/NCBI
|
|
16.
|
Mersmann HJ: Overview of the effects of
beta-adrenergic receptor agonists on animal growth including
mechanisms of action. J Anim Sci. 76:160–172. 1998.PubMed/NCBI
|
|
17.
|
Anderson GP: Pharmacology of formoterol:
an innovative bronchodilator. Agents Actions Suppl. 34:97–115.
1991.PubMed/NCBI
|
|
18.
|
Mahler DA: The effect of inhaled
beta2-agonists on clinical outcomes in chronic obstructive
pulmonary disease. J Allergy Clin Immunol. 110:S298–S303. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
19.
|
Busquets S, Figueras MT, Fuster G, et al:
Anticachectic effects of formoterol: a drug for potential treatment
of muscle wasting. Cancer Res. 64:6725–6731. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
20.
|
Tessitore L, Costelli P, Bonetti G and
Baccino FM: Cancer cachexia, malnutrition, and tissue protein
turnover in experimental animals. Arch Biochem Biophys. 306:52–58.
1993. View Article : Google Scholar : PubMed/NCBI
|
|
21.
|
Sinis N, Guntinas-Lichius O, Irintchev A,
et al: Manual stimulation of forearm muscles does not improve
recovery of motor function after injury to a mixed peripheral
nerve. Exp Brain Res. 185:469–483. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
22.
|
Zangarelli A, Chanseaume E, Morio B, et
al: Synergistic effects of caloric restriction with maintained
protein intake on skeletal muscle performance in 21-month-old rats:
a mitochondria-mediated pathway. FASEB J. 20:2439–2450. 2006.
View Article : Google Scholar
|
|
23.
|
Fuster G, Busquets S, Ametller E, et al:
Are peroxisome proliferator-activated receptors involved in
skeletal muscle wasting during experimental cancer cachexia? Role
of beta2-adrenergic agonists. Cancer Res. 67:6512–6519. 2007.
View Article : Google Scholar
|
|
24.
|
Costelli P, Garcia-Martinez C, Llovera M,
et al: Muscle protein waste in tumor-bearing rats is effectively
antagonized by a beta 2-adrenergic agonist (clenbuterol). Role of
the ATP-ubiquitin-dependent proteolytic pathway. J Clin Invest.
95:2367–2372. 1995. View Article : Google Scholar
|
|
25.
|
Ametller E, Busquets S, Fuster G, et al:
Formoterol may activate rat muscle regeneration during cancer
cachexia. Insciences J. 1:1–17. 2011. View Article : Google Scholar
|
|
26.
|
Lanigan C, Howes TQ, Borzone G, Vianna LG
and Moxham J: The effects of beta 2-agonists and caffeine on
respiratory and limb muscle performance. Eur Respir J. 6:1192–1196.
1993.PubMed/NCBI
|
|
27.
|
Signorile JF, Banovac K, Gomez M, Flipse
D, Caruso JF and Lowensteyn I: Increased muscle strength in
paralyzed patients after spinal cord injury: effect of beta-2
adrenergic agonist. Arch Phys Med Rehabil. 76:55–58. 1995.
View Article : Google Scholar : PubMed/NCBI
|
|
28.
|
Marzabal M, Garcia-Martinez C, Comas J,
Lopez-Soriano FJ and Argiles JM: A flow cytometric study of the rat
Yoshida AH-130 ascites hepatoma. Cancer Lett. 72:169–173. 1993.
View Article : Google Scholar : PubMed/NCBI
|
|
29.
|
Toledo M, Busquets S, Sirisi S, et al:
Cancer cachexia: physical activity and muscle force in
tumour-bearing rats. Oncol Rep. 25:189–193. 2011.PubMed/NCBI
|
|
30.
|
Argiles JM, Busquets S, Toledo M and
Lopez-Soriano FJ: The role of cytokines in cancer cachexia. Curr
Opin Support Palliat Care. 3:263–268. 2009. View Article : Google Scholar
|
|
31.
|
Llovera M, Garcia-Martinez C,
Lopez-Soriano J, et al: Role of TNF receptor 1 in protein turnover
during cancer cachexia using gene knockout mice. Mol Cell
Endocrinol. 142:183–189. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
32.
|
Argiles JM, Busquets S and Lopez-Soriano
FJ: Cytokines as mediators and targets for cancer cachexia. Cancer
Treat Res. 130:199–217. 2006. View Article : Google Scholar : PubMed/NCBI
|