Contents
Introduction
microRNAs contribute to pancreatic cancer invasion
and metastasis
microRNAs and epithelial to mesenchymal
transition
microRNAs and cancer stem cells or tumor initiating
cells
microRNAs and tumor-associated signaling
pathways
microRNAs and epigenetic alterations
Conclusion
Introduction
Pancreatic cancer is the fourth most common cause of
cancer-related death with 43,140 new diagnoses and 36,800 deaths
during 2010 in the US (1).
Surgical resection is currently the only curative option for
pancreatic cancer, however, due to unusual biological property,
high invasive ability and early metastatic feature, only ∼15 to 20%
of tumors can be surgically removed when diagnosed. Even after
radical resection, recurrence and metastasis occur within 1–2 years
in the majority of cases and the prognosis remains poor (2). Therefore, there is a growing need to
deeply understand the mechanisms of the invasive and metastatic
profile of pancreatic cancer, which may ultimately lead to an
improvement in the prognosis of this fatal disease.
microRNAs (miRNAs) are a class of natural small
non-coding RNAs that target protein-coding mRNAs at the
post-transcriptional level (3,4).
Wightman and coworkers discovered the first miRNA in
Caenorhabditis elegants in 1993 (5), and in 2000 the second miRNA, let-7,
was found in the same species (6).
Abnormal expression of miRNAs is fundamental to the development and
progression of various cancers based on their involvement in basic
cellular functions. The link between miRNAs and cancer was firstly
demonstrated in 2002 by Calin et al (7), who found that miR-15 and miR-16 were
involved in the pathogenesis of chronic lymphocytic leukemia.
Subsequently, the critical role of miRNA in cancer initiation and
progression has been reported in various cancers including
pancreatic cancer (8–10). It was also demonstrated that there
is a significantly different miRNA expression profile in metastatic
carcinomas compared to non-metastatic tumors, and these
metastasis-related miRNAs also significantly correlate with the
survival of patients (11).
Therefore, interference with the expression of these miRNAs may
affect tumor metastasis and improve prognosis. In this review, we
summarize several important miRNAs in pancreatic cancer
progression, highlighting recent advances in elucidating the role
of miRNAs in pancreatic cancer invasion and metastasis.
microRNAs contribute to pancreatic cancer
invasion and metastasis
The role of miRNAs in the development of tumor
invasion and metastasis was not discovered until 2007. Ma et
al (12) reported that miR-10b
initiated breast cancer invasion and metastasis. Subsequently, the
important role of miRNA in cancer invasion and metastasis in
various human malignant tumors, including liver, prostate, lung and
colorectal cancers has been reported (13–17).
In recent years, various miRNAs have also been found to play a
significant role in pancreatic cancer invasion and metastasis.
miR-21, which is strongly overexpressed in pancreatic cancer, has
been proven to be an ‘oncogene’ miRNA and its high expression was
found to contribute to poor overall survival and chemotherapy
resistance (18,19). It has also been reported that
miR-21 modulates pancreatic cancer cell growth and invasion.
Pancreatic cancer cells transfected with miR-21 precursor were
found to exhibit apparently increased cancer cell proliferation and
invasion. Conversely, inhibition of miR-21 has an opposite outcome.
Furthermore, the level of metastasis-related genes, matrix
metalloproteinase-2 and -9, and vascular endothelial growth factor
were positively correlated with miR-21 expression, suggesting that
MMP-2, MMP-9 and VEGF are ‘indirect’ target genes of miR-21
(20).
miR-146a has been found to be silent in multiple
human cancers, and restoration of its expression can reduce the
meta-static potential of cancer cells through suppression of IRAK-1
and subsequent inhibition of NF-κB activity (15,21,22),
which is activated in a number of human cancers and involved in
promoting tumor development, invasion and metastasis (23,24).
In pancreatic cancer, miR-146a is also downregulated, and
re-expression of miR-146a inhibits the invasive capacity of
pancreatic cancer cells. Notably, a number of studies have
indicated that the natural dietary compounds 3,3′-diindolyl-methane
(DIM) or isoflavone may upregulate miR-146a and inhibit cancer cell
invasion. Further mechanistic analysis suggests that miR-146a
regulates a set of genes and suppresses cancer cell invasion and
migration through targeting EGFR and IRAK-1 (25). Another study also revealed that
miR-146b-5p was significantly downregulated in human pancreatic
cancer cells, and overexpressed miR-146b-5p notably reduced the
abilities of invasion and migration of MIAPaCa-2 pancreatic cancer
cells by targeting MMP-16 (26).
In addition, miR-27a is abnormally upregulated in
pancreatic adenocarcinoma and inhibition of miR-27a was found to
suppress the growth, colon formation and migration of pancreatic
cancer cells by targeting Sprouty2, which is an antagonist of the
RAS/MAPK signaling pathway in cancers (27). More recently, Yu et al
(28) reported that re-expression
of miR-200c appeared to be associated with upregulation of
E-cadherin, a gene known to be involved in inhibiting the invasion
of pancreatic cancer cells. Wang et al (29) identified that miR-520h
downregulated ABCG2 in pancreatic cancer cells to inhibit
migration, invasion and side population, and indicated that it
could be a potential therapeutical target for pancreatic cancer.
Preis et al (30) found
that miR-10b was overexpressed in pancreatic ductal adenocarcinoma
tissues and lower levels of miR-10b were linked with improved
response to neoadjuvant therapy, delayed time to metastasis and
increased survival. Srivastava et al (31) reported that the level of miR-150
was significantly lower in pancreatic tumors compared with matched
normal pancreas tissue. Furthermore, ectopic expression of miR-150
significantly inhibited pancreatic cancer cell invasion and
migration as well as tumor growth by suppressing the MUC4 gene
in vitro. Other miRNAs have also been reported to play an
important role in pancreatic cancer cell invasion and migration,
such as miR-17-5p, miR-29a and miR-20a (32–36).
Results from these studies may provide an opportunity to carry out
miRNA-based therapeutic intervention for pancreatic cancer
metastasis (Table I).
 | Table I.Pancreatic cancer invasion and
metastasis-related miRNAs. |
Table I.
Pancreatic cancer invasion and
metastasis-related miRNAs.
| miRNA |
Up/downregulated | Potential
target | Materials | Reference |
|---|
| miR-21 | Up | MMP-2, MMP-9,
VEGF | PANC-1, AsPC-1,
CFPAC-1, SUIT-2 | (20) |
| miR-146a | Down | EGFR, NF-κB, MTA-2,
IRAK-1 | Colo357,
PANC-1 | (25) |
| miR-146b-5p | Down | MMP16 | MIA-PaCa and
FFPE | (26) |
| miR-27a | Up | Spry2 | PANC-1, MIA-PaCa-2,
FFPE | (27) |
| miR-200c | Down | E-cadherin | FFPE, SUIT-2, KP-3,
PANC-1 | (28) |
| miR-520h | Up | ABCG2 | PANC-1 | (29) |
| miR-10b | Up | nc | FFPE, EUS-FNA
sample | (30) |
| miR-150 | Down | MUC4 | Panc10.05, HPAF,
Colo357 | (31) |
| miR-17-5P | Up | nc | FFPE, SUIT-2,
KP-2 | (32) |
| miR-29a | Down | nc | PANC-1 | (33) |
| miR-20a | Uown | Stat3 | PANC-1, BxPC-3 | (34) |
| miR-126 | Down | ADAM9 | PANC-1, ASPC-1,
FFPE, FNA | (35) |
| miR-26a | Down | HMGA1 | PANC-1, Sw1990,
nude mice | (36) |
| miR-96 | Down | KRAS | MIA-PaCa-2, PANC-1,
FFPE, nude mice | (78) |
microRNAs and epithelial to mesenchymal
transition
The epithelial to mesenchymal transition (EMT) is a
process by which epithelial cells lose their polarity and are
converted to a mesenchymal phenotype, which is regarded as a
critical event in morphogenetic changes during embryonic
development, wound healing and malignant tumor progression
(37,38). This process is accompanied by
detachment of cells from each other and subsequent increased cell
movement and dissemination. Increasing evidence shows that aberrant
activation of EMT is a trigger of malignant tumor invasion and
metastasis in various human cancers (39–41).
The transcriptional repressor zinc-finger E-box binding homeobox 1
(ZEB1), a crucial inducer of EMT, was recently shown to promote
cancer invasion and metastasis in vitro and in vivo
(42). Furthermore, members of the
miR-200 family (miR-200a, miR-200b, miR-200c, miR-141) induce
epithelial differentiation through direct targeting of ZEB1 and
ZEB2 (43) whereas ZEB1 was
reported to directly suppress the transcription of two members of
the miR-200a family (miR-200c and miR-141). Therefore, several
researchers suggest that there is a feedforward loop between the
miR-200 family and ZEB1, which promotes EMT and invasion in cancer
cells (44).
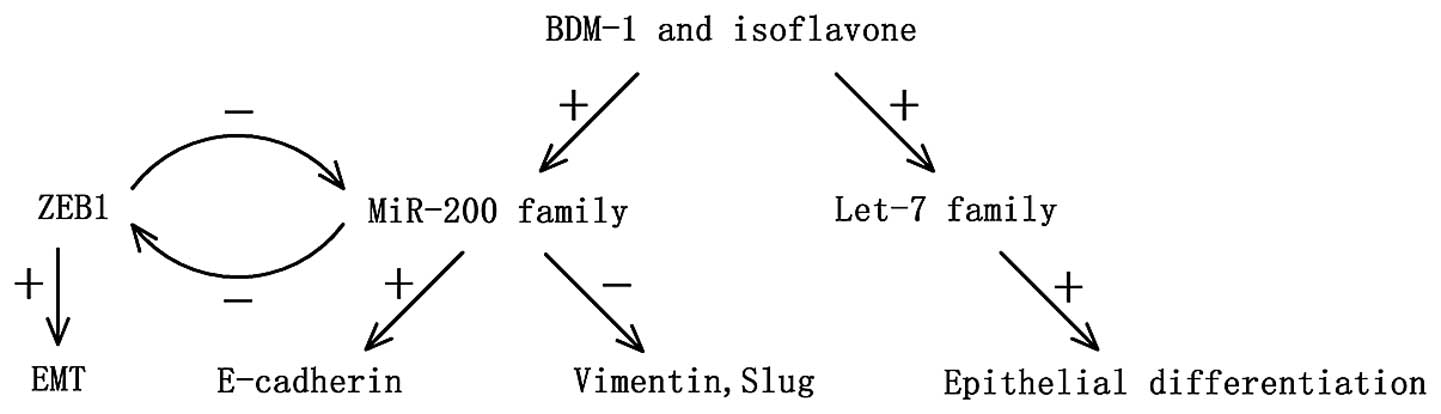
Notably, this feedforward loop also exists in
pancreatic cancer. Wellner et al (45) reported that expression of miR-200c
and miR-203 is low in pancreatic cancer, while ZEB1 is
overexpressed. In vitro, silencing of ZEB1 by RNA
interference in PANC-1 and MIA-PaCa-2 cell lines resulted in an
epithelial transition. In orthotopic xenograft models, injection of
ZEB1-knockdown cell clones resulted in smaller primary tumors and
less invasion and distant metastasis than the control group. In
another study, the level of the miR-200 family was found to be
significantly low in gemcitabine-resistant cancer cells, which
showed a typical EMT characteristic, and re-expression of miR-200
could be essential for the reversal of the EMT phenotype, and thus
inhibit cancer cell invasion and metastasis through increasing the
epithelial marker E-cadherin expression and suppressing the
expression of ZEB1, slug and vimentin. Furthermore, the natural
agent B-DIM and isoflavone may also significantly upregulate the
expression of let-7, which is known as a tumor-suppressor miRNA
(46,47), consequently inhibiting pancreatic
cancer progression by reversing EMT characteristics (48) (Fig.
1).
In summary, the expression of various miRNAs may
affect EMT characteristics, whereas the EMT process is necessary
for the invasion and metastasis of pancreatic cancer. Therefore,
identification of the aberrant expression of these miRNAs may
provide insight concerning important mechanisms that contribute to
pancreatic cancer development and progression.
microRNAs and cancer stem cells or
tumor-initiating cells
Cancer stem cells (CSCs) or tumor-initiating cells
are a small population of cells within a tumor that initiate
proliferation, possess the capacity for self-renewal and ensure
multilineage differentiation. The presence of CSCs is considered as
the origination of various malignant biological behaviors in human
cancers, especially involved in cancer cell migration, invasion and
metastasis (49). CSCs were
initially discovered in myeloid leukemia cells by Bonnet and Dick
(50) in 1997. Subsequently, the
existence of CSCs was verified in various cancers, including
breast, colorectal and brain cancer (51–53).
In 2007, a small subset of highly tumorigenic cancer cells with the
cell surface markers CD44, CD24, epithelial-specific antigen (ESA)
triple-positive expression, was found in human pancreatic cancer
cells (54), which was proven to
possess the characteristics of CSCs. Meanwhile, a subpopulation of
CD133+, CXCR4+ pancreatic cancer stem cells
was verified and confirmed to be essential for tumor metastasis
(55). Furthermore,
CD133+ pancreatic cancer cells were also found to be
related with increased cell proliferation, migration and invasion
(56). All of these findings
indicate that the presence of CSCs is responsible for the high
invasive and metastatic phenotype of pancreatic cancer.
As discussed above, there is a feedforward loop
between the miR-200 family and ZEB1 that promotes EMT and invasion
in pancreatic cancer cells. It has also been reported that ZEB1
promotes EMT activation and maintains the stemness of cancer cells
by suppressing the miR-200 family and miR-203, which is also known
as a stemness-inhibiting miRNA, consequently promoting pancreatic
cancer invasion and metastasis. In addition, the miR-200 family and
miR-203 were found to suppress the expression of ‘stem cell
factors’ such as Sox2, Kif4 and the polycomb repressor Bmi-1 in
cancer cells (45). These data
suggest that the miR-200 family and miR-203 may prevent pancreatic
cancer invasion and metastasis not only by reversion of epithelial
to mesenchymal transition, but also by suppressing cancer stem
cells.
DCAMKL-1, a microtubule-associated kinase expressed
in post-mitotic neurons, is upregulated in human pancreatic cancer
and was identified as a putative cancer stem cell marker (57). Silencing of DCAMK-1 in pancreatic
cancer cells resulted in suppression of snail, slug, twist and
upregulation of miR-200a, which inhibits EMT by repressing the
transcription factor ZEB1 and ZEB2 with subsequent rescue of
E-cadherin, a marker of epithelial lineage. It was also suggested
that DCAMKL-1 regulates epithelial-mesenchymal transition through a
miR-200a-dependent mechanism, contributing to pancreatic cancer
development, invasion and metastasis. In addition, it was found
that DCAMKL-1 knockdown resulted in downregulation of c-Myc and
KRAS through a let-7a miRNA-dependent mechanism, and inhibition of
the Notch-1 pathway by upregulation of miR-144 (58). These results suggest that miRNAs
may modulate tumor stem cell properties by regulating their target
or related genes.
miR-34 was identified as a p53 target and a
tumor-suppressor miRNA (59,60).
It has been reported that miR-34 is involved in pancreatic cancer
stem cell self-renewal and differentiation. In addition,
restoration of miR-34 significantly inhibited the p53-mutant human
pancreatic cancer cell growth and invasion and finally led to an
87% reduction in the CD133+/CD44+
tumor-initiating cell population. Furthermore, miR-34 was found to
inhibit its target genes Notch and Bcl-2, ‘stem cell genes’ to
achieve a reduction in cancer stem cells (61). A similar function of miR-34 has
also been reported in gastric cancer (62). In addition, it was reported that
the curcumin analogue CDF may attenuate EZH2 and other cancer stem
cell marker genes, such as Nanog, CD44 and EpCAM through
upregulation of tumor-suppressive miRNAs (miR-26a, miR-101,
miR-146a, miR-200b and c, let-7a, b, c and d), thus decreasing cell
growth, clonogenicity, cell migration and the elimination of CSCs
(63).
It is possible that there is a unique miRNA
signature responsible for the maintenance and enrichment of cancer
stem cells. These stem cell-specific miRNAs may influence
pancreatic cancer invasion and metastasis by regulating the
‘stemness’ and also may provide a potential therapeutic target for
pancreatic cancer metastasis (Table
II).
| Table II.miRNAs and pancreatic cancer stem
cell markers. |
Table II.
miRNAs and pancreatic cancer stem
cell markers.
| miRNA |
Up/downregulated | Related stem cell
markers | Materials | Reference |
|---|
| miR-200 family and
miR-203 | Down | ZEB1 | MIA-PaCa-2, BxPC-3,
PANC-1 | (45) |
| miR-200a, let-7 and
miR-144 | Down | DCAMKL-1 | AsPC-1, BxPC-3 | (58) |
| miR-34 | Down | CD133/CD44 | MIA-PaCa-2,
BxPC-3 | (61) |
| miR-101 | Down | EZH2, EpCAM, Nanog,
Sox2, Oct4 | MIA-PaCa-2 | (63) |
microRNAs and tumor-associated signaling
pathways
It is well known that various signaling pathways
such as Notch, hedgehog and Wnt/β-catenin are dysregulated in human
carcinomas including pancreatic cancer consequently contributing to
tumor development and progression. It has also been shown that
several miRNAs appear to possess various essential functions in
cancer invasion and metastasis through regulating important
signaling pathways. Therefore, the interaction between miRNAs and
tumor progression-related signaling pathways in human cancer must
be elucidated in detail.
The Notch signaling pathway has been well-documented
to be involved in the regulation of numerous cellular process,
including cell proliferation, apoptosis, differentiation, invasion
and metastasis (64,65). The Notch pathway has been reported
to play both oncogenic and tumor-suppressor roles in multiple human
cancers (66–68). In pancreatic cancer, the Notch
signaling pathway is frequently deregulated with upregulated
expression of Notch receptors and their ligands, and functions as
an oncogene during tumor growth and progression (69). This is thought to be related to the
absence of miR-34. As discussed above, in
CD44+/CD133+ MIA-PaCa-2 cancer stem cells
with high Notch/Bcl-2 levels, miR-34 restoration significantly
inhibits tumor growth and formation in vitro and in
vivo. It has also been reported that miR-34 is silent in
pancreatic cancer, and restoration of miR-34 expression in
pancreatic cancer cell lines MIA-PaCa-2 and Bxpc-3 downregulates
the expression of Bcl-2 and Notch1/2, subsequently inhibiting
clonogenic cell growth and cell invasion (61). Recently, Nalls et al
(70) reported that upregulation
of miR-34a by 5-Aza-dC and HDAC inhibitor vorinostat (SAHA) in
pancreatic cancer cells suppressed the cell proliferation, cell
cycle progression, cell self-renewal, cell invasion and EMT
process. Moreover, re-expression of miR-34 by SAHA in pancreatic
cancer stem cells inhibited the mRNA expression of all the
components of the Notch pathway, including Notch receptor, Notch-1,
Notch-3, its ligand Jagged 1 and Notch target gene Hes1; thus
miR-34a may inhibit cancer invasion and progression by targeting
the Notch signaling pathway (70).
In addition, overexpression of Notch-1 in AsPC-1 cells leads to
increased cell growth, invasion, clonogenicity, migration,
induction of EMT phenotype and decreased expression of miR-200a,
miR-200c, let7a, let7b and let7c, while re-expression of miR-200b
attenuates the acquisition of the EMT phenotype in large part due
to the downregulation of the expression of Notch-1 in
Notch-1-overexpressed AsPC-1 cells (71).
The Sonic hedgehog (Shh) pathway is aberrantly
activated and plays a decisive role in the development and
progression of multiple human cancers (72). In contrast, the blockage of the
hedgehog pathway could inhibit cancer invasion and metastasis in
vivo (73). Increasing
evidence supports the possibility that there is an inner link
between miRNAs and the hedgehog signaling pathway in human cancers
(74,75). It was reported that certain miRNAs
activate the hedgehog signaling pathway by targeting Smo, Cos2 and
Fu in Drosophila, which are antagonistic components of the
hedgehog pathway (76). Moreover,
Tsuda et al (77) reported
that synthetic Gli-1-miRNA-3548 and its corresponding Duplex-3548
inhibited cell division and stimulated apoptosis of MIA-PaCa-2
cells through targeting of the Gli-1 gene which controls the
pathway of Shh signaling; this suggests that Gli-1 inhibition with
miRNA may be a potentially novel approach to pancreatic cancer
therapy.
Moreover, miRNAs promote pancreatic cancer invasion
and metastasis through the regulation of other tumor associated
signaling pathways including RAS, TGF-β and NF-κB. For example,
ectopic expression of miR-96 inhibited pancreatic cancer invasion
and migration with KRAS down-regulation in vitro and in
vivo, and miR-96 was identified as a core regulator of the
KRAS/AKT pathway (78).
These studies suggest that various miRNAs play key
roles in multiple signal pathways that are responsible for
pancreatic cancer invasion and metastasis, and these miRNAs are
essential for the unique metastatic characteristics of pancreatic
cancer.
microRNAs and epigenetic alterations
Epigenetic modification such as DNA hypermethylation
could have a great impact on the expression of miRNAs, and thus
contribute to cancer development and progression. Frequent DNA
methylation of cytosine-phospho-guanine (-CpG) island regions
adjacent to miR-34a and miR-34b/c, the direct p53 target genes, was
observed in various human malignant tumors, including colorectal,
prostate and pancreatic cancer, which may mediate cancer cell
apoptosis, cell cycle arrest and senescence (79). Epigenetic silencing of miR-107
contributes to MIA-PaCa-2 and PANC-1 cell growth through regulating
the expression of cyclin-dependent kinase 6 (CDK-6), which is a
cyclin-D1-dependent kinase facilitating cell cycle progression by
regulating the activity of tumor-suppressor protein Rb (80). In addition, the epigenetic
inactivation of miRNAs can be reversed by treatment with the DNA
demethylating agent, 5-aza-2,-deoxycytidine (DAC) and the histone
deacetylase inhibitor, trichostatin A (TSA) (81). Thus, it is possible that aberrant
miRNA epigenetic alterations could also play a vital role in
pancreatic cancer invasion and metastasis. Recently, miR-224 and
miR-486 were found to be significantly overexpressed with
epigenetic alterations in highly invasive and metastatic pancreatic
cancer, while protein expression of the cell surface marker CD40, a
member of the tumor necrosis factor family related to anti-tumor
immune responses, was low (82,83).
Thus, miRNA-regulated CD40 expression seems to be essential for
pancreatic cancer invasion and the process of metastasis (84).
EP-300 is a group of proteins that function as
transcriptional coactivators and are also involved in tumor
development and progression (85,86).
By using 16 human PDAC cell lines in a murine orthotopic PDAC
model, Mees et al (87)
reported that epigenetic alterations with upregulation of miR-184,
miR-200b, miR-200c and miR-429, which target EP300, are likely to
cause downregulation of EP300 mRNA and protein expression in highly
metastatic PDAC cell lines. The authors also indicated that miRNAs
may be able to affect metastatic behavior of pancreatic cancer by
modulating the expression of metastasis-related suppressor genes,
such as EP-300 (87).
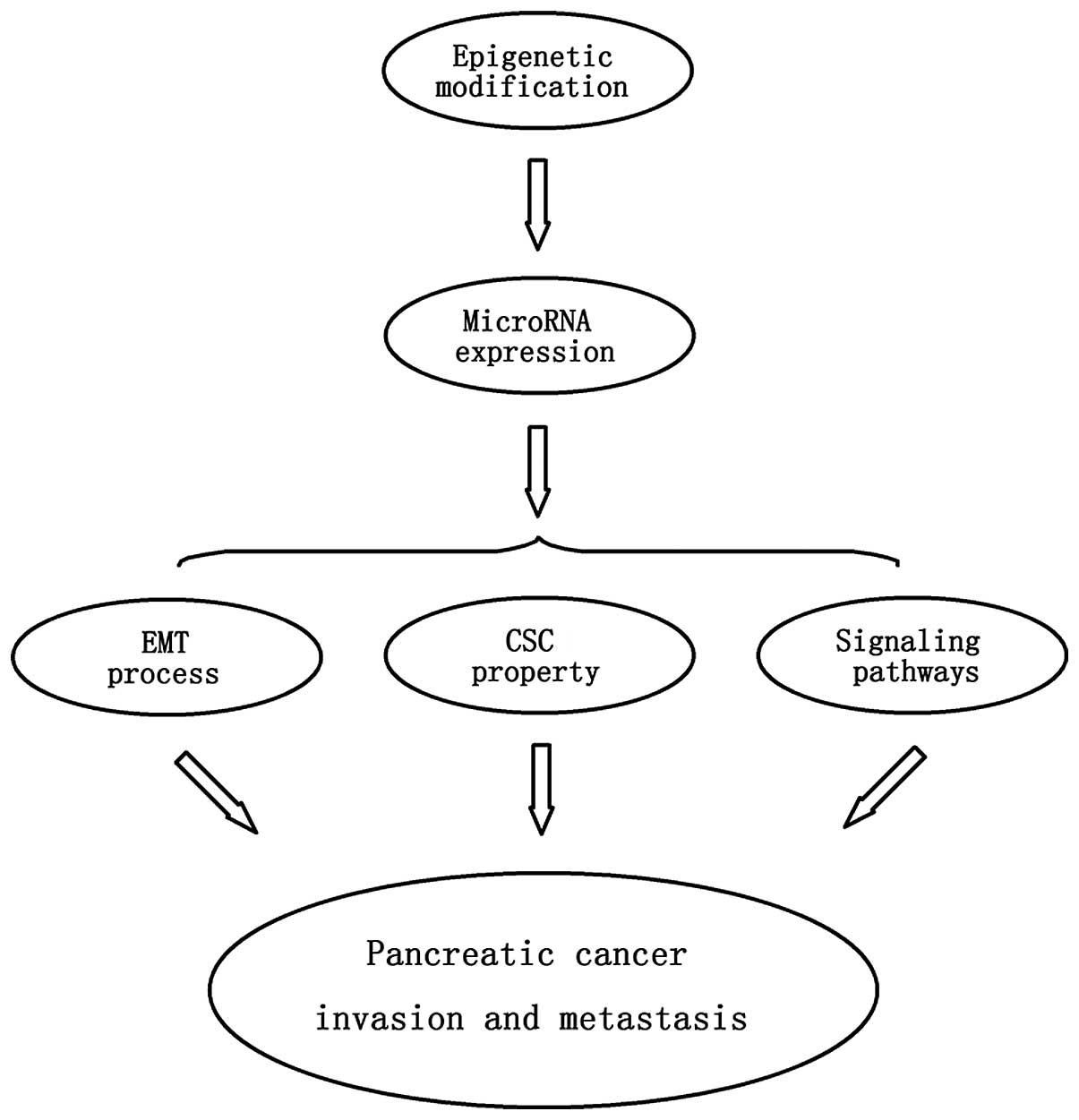
In summary, the expression of various specific
miRNAs functioning as tumor-suppressor genes or oncogenes may be
regulated by epigenetic modifications such as DNA methylation,
finally contributing to pancreatic cancer progression and
metastasis. As previously mentioned, differential expression of
miRNAs may affect the EMT process, cancer stem cell properties and
specific tumor signaling pathways, which may ultimately modulate
cancer invasion and metastasis. Therefore, the epigenetic
modification responsible for abnormal expression of miRNAs in
pancreatic tumors may be a crucial mechanism contributing to cancer
invasion and metastasis (Fig.
2).
Conclusion
Although knowledge concerning the role of miRNA in
the regulation of cancer development and progression has been
greatly advanced since the first discovery of miRNA nearly 18 years
ago, the details of miRNA-specific molecular pathogenesis of
pancreatic cancer still remain to be elucidated. Recent studies
suggest that a variety of miRNAs are frequently dysregulated in
pancreatic cancer and have a crosstalk with various important
biological processes including DNA methylation, the presence of
cancer stem cells, the process of EMT and associated signaling
pathways, which may be crucial in tumorigenesis and metastasis.
Moreover, research concerning the targeting of miRNAs in
vitro and in vivo has demonstrated amazing results in
altering the features of pancreatic cancer invasion and metastasis.
Various related miRNAs affect pancreatic cancer invasion and
metastasis through the regulation of the EMT process, cancer stem
cell properties, and tumor-associated signaling pathways, while
epigenetic modification may be an important mechanism causing
dysregulation of miRNA expression. Therefore, detailed exploration
of miRNA molecular mechanisms not only contributes to the
understanding of the origin of the highly metastatic properties of
pancreatic cancer, but may also provide insight for the improvement
of the prognosis for this fatal disease.
Acknowledgements
This study was supported by National
Natural Science Foundation of China grants (Nos. 81172276 and
81001058).
References
|
1.
|
Jemal A, Siegel R, Xu J and Ward E: Cancer
statistics, 2010. CA Cancer J Clin. 60:277–300. 2010. View Article : Google Scholar
|
|
2.
|
Zuckerman DS and Ryan DP: Adjuvant therapy
for pancreatic cancer: a review. Cancer. 112:243–249. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
3.
|
Esquela-Kerscher A and Slack FJ: Oncomirs
- microRNAs with a role in cancer. Nat Rev Cancer. 6:259–269. 2006.
View Article : Google Scholar
|
|
4.
|
Denli AM, Tops BB, Plasterk RH, Ketting RF
and Hannon GJ: Processing of primary microRNAs by the
microprocessor complex. Nature. 432:231–235. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
5.
|
Wightman B, Ha I and Ruvkun G:
Posttranscriptional regulation of the heterochronic gene lin-14 by
lin-4 mediates temporal pattern formation in C. elegans.
Cell. 75:855–862. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
6.
|
Reinhart BJ, Slack FJ, Basson M, et al:
The 21-nucleotide let-7 RNA regulates developmental timing in
Caenorhabditis elegans. Nature. 403:901–906. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
7.
|
Calin GA, Dumitru CD, Shimizu M, et al:
Frequent deletions and down-regulation of micro-RNA genes miR15 and
miR16 at 13q14 in chronic lymphocytic leukemia. Proc Natl Acad Sci
USA. 99:15524–15529. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
8.
|
Bloomston M, Frankel WL, Petrocca F, et
al: microRNA expression patterns to differentiate pancreatic
adenocarcinoma from normal pancreas and chronic pancreatitis. JAMA.
229:1901–1908. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
9.
|
Lee JW, Choi CH, Choi JJ, et al: Altered
microRNA expression in cervical carcinomas. Clin Cancer Res.
14:2535–2542. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
10.
|
Iorio MV, Ferracin M, Liu CG, et al:
MicroRNA gene expression deregulation in human breast cancer.
Cancer Res. 65:7065–7070. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
11.
|
Budhu A, Jia HL, Forgues M, et al:
Identification of metastasis-related microRNAs in hepatocellular
carcinoma. Hepatology. 47:897–907. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
12.
|
Ma L, Teruya-Feldstein J and Weinberg RA:
Tumour invasion and metastasis initiated by microRNA-10b in breast
cancer. Nature. 449:682–688. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
13.
|
Meng F, Henson R, Wehbe-Janek H, Ghoshal
K, Jacob ST and Patel T: MicroRNA-21 regulates expression of the
PTEN tumor suppressor gene in human hepatocellular cancer.
Gastroenterology. 133:647–658. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
14.
|
Asangani IA, Rasheed SA, Nikolova DA, et
al: MicroRNA-21 (miR-21) post-transcriptionally downregulates tumor
suppressor Pdcd4 and stimulates invasion, intravasation and
metastasis in colorectal cancer. Oncogene. 27:2128–2136. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
15.
|
Lin SL, Chiang A, Chang D and Ying SY:
Loss of mir-146a function in hormone-refractory prostate cancer.
RNA. 14:417–424. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
16.
|
Crawford M, Brawner E, Batte K, et al:
MicroRNA-126 inhibits invasion in non-small cell lung carcinoma
cell lines. Biochem Biophys Res Commun. 373:607–612. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
17.
|
Huang Q, Gumireddy K, Schrier M, et al:
The microRNAs miR-373 and miR-520c promote tumour invasion and
metastasis. Nat Cell Biol. 10:202–210. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
18.
|
Hwang JH, Voortman J, Giovannetti E, et
al: Identification of microRNA-21 as a biomarker for
chemoresistance and clinical outcome following adjuvant therapy in
resectable pancreatic cancer. PloS One. 5:e106302010. View Article : Google Scholar : PubMed/NCBI
|
|
19.
|
Giovannetti E, Funel N, Peters GJ, et al:
MicroRNA-21 in pancreatic cancer: correlation with clinical outcome
and pharmacologic aspects underlying its role in the modulation of
gemcitabine activity. Cancer Res. 70:4528–4538. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
20.
|
Moriyama T, Ohuchida K, Mizumoto K, et al:
MicroRNA-21 modulates biological functions of pancreatic cancer
cells including their proliferation, invasion, and chemoresistance.
Mol Cancer Ther. 8:1067–1074. 2009. View Article : Google Scholar
|
|
21.
|
Bhaumik D, Scott GK, Schokrpur S, Patil
CK, Campisi J and Benz CC: Expression of microRNA-146 suppresses
NF-kappaB activity with reduction of metastatic potential in breast
cancer cells. Oncogene. 27:5643–5647. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
22.
|
Kogo R, Mimori K, Tanaka F, Komune S and
Mori M: Clinical significance of miR-146a in gastric cancer cases.
Clin Cancer Res. 17:4277–4284. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
23.
|
Fujioka S, Sclabas GM, Schmidt C, et al:
Function of nuclear factor kappaB in pancreatic cancer metastasis.
Clin Cancer Res. 9:346–354. 2003.PubMed/NCBI
|
|
24.
|
Karin M: Nuclear factor-kappaB in cancer
development and progression. Nature. 441:431–436. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
25.
|
Li Y, Vandenboom TG II, Wang Z, et al:
miR-146a suppresses invasion of pancreatic cancer cells. Cancer
Res. 70:1486–1495. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
26.
|
Lin F, Wang X, Jie Z, et al: Inhibitory
effects of miR-146b-5p on cell migration and invasion of pancreatic
cancer by targeting MMP16. J Huazhong Univ Sci Technolog Med Sci.
31:509–514. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
27.
|
Ma Y, Yu S, Zhao W, Lu Z and Chen J:
miR-27a regulates the growth, colony formation and migration of
pancreatic cancer cells by targeting Sprouty2. Cancer Lett.
298:150–158. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
28.
|
Yu J, Ohuchida K, Mizumoto K, et al:
MicroRNA, hsa-miR-200c, is an independent prognostic factor in
pancreatic cancer and its upregulation inhibits pancreatic cancer
invasion but increases cell proliferation. Mol Cancer. 9:1692010.
View Article : Google Scholar : PubMed/NCBI
|
|
29.
|
Wang F, Xue X, Wei J, et al: hsa-miR-520h
downregulates ABCG2 in pancreatic cancer cells to inhibit
migration, invasion, and side populations. Br J Cancer.
103:567–574. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
30.
|
Preis M, Gardner TB, Gordon SR, et al:
MicroRNA-10b expression correlates with response to neoadjuvant
therapy and survival in pancreatic ductal adenocarcinoma. Clin
Cancer Res. 17:5812–5821. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
31.
|
Srivastava SK, Bhardwaj A, Singh S, et al:
MicroRNA-150 directly targets MUC4 and suppresses growth and
malignant behavior of pancreatic cancer cells. Carcinogenesis.
32:1832–1839. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
32.
|
Yu J, Ohuchida K, Mizumoto K, Fujita H,
Nakata K and Tanaka M: MicroRNA miR-17-5p is overexpressed in
pancreatic cancer, associated with a poor prognosis, and involved
in cancer cell proliferation and invasion. Cancer Biol Ther.
10:748–757. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
33.
|
Muniyappa MK, Dowling P, Henry M, et al:
MiRNA-29a regulates the expression of numerous proteins and reduces
the invasiveness and proliferation of human carcinoma cell lines.
Eur J Cancer. 45:3104–3118. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
34.
|
Yan H, Wu J, Liu W, et al: MicroRNA-20a
overexpression inhibited proliferation and metastasis of pancreatic
carcinoma cells. Hum Gene Ther. 21:1723–1734. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
35.
|
Hamada S, Satoh K, Fujibuchi W, et al:
MiR-126 acts as a tumor suppressor in pancreatic cancer cells via
the regulation of ADAM9. Mol Cancer Res. 10:3–10. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
36.
|
Li W, Yuan Y, Huang L, Qiao M and Zhang Y:
Metformin alters the expression profiles of microRNAs in human
pancreatic cancer cells. Diabetes Res Clin Pract. 96:187–195. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
37.
|
Thiery JP, Acloque H, Huang RY and Nieto
MA: Epithelialmesenchymal transitions in development and disease.
Cell. 139:871–890. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
38.
|
Thiery JP and Sleeman JP: Complex networks
orchestrate epithelial-mesenchymal transitions. Nat Rev Mol Cell
Biol. 7:131–142. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
39.
|
Christiansen JJ and Rajasekaran AK:
Reassessing epithelial to mesenchymal transition as a prerequisite
for carcinoma invasion and metastasis. Cancer Res. 66:8319–8326.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
40.
|
Xiong H, Hong J, Du W, et al: Roles of
STAT3 and ZEB1 proteins in E-cadherin down-regulation and human
colorectal cancer epithelial-mesenchymal transition. J Biol Chem.
287:5819–5832. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
41.
|
Maier HJ, Schmidt-Strassburger U, Huber
MA, Wiedemann EM, Beug H and Wirth T: NF-kappaB promotes
epithelialmesenchymal transition, migration and invasion of
pancreatic carcinoma cells. Cancer Lett. 295:214–228. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
42.
|
Spaderna S, Schmalhofer O, Wahlbuhl M, et
al: The transcriptional repressor ZEB1 promotes metastasis and loss
of cell polarity in cancer. Cancer Res. 68:537–544. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
43.
|
Korpal M, Lee ES, Hu G and Kang Y: The
miR-200 family inhibits epithelial-mesenchymal transition and
cancer cell migration by direct targeting of E-cadherin
transcriptional repressors ZEB1 and ZEB2. J Biol Chem.
283:14910–14914. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
44.
|
Burk U, Schubert J, Wellner U, et al: A
reciprocal repression between ZEB1 and members of the miR-200
family promotes EMT and invasion in cancer cells. EMBO Rep.
9:582–589. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
45.
|
Wellner U, Schubert J, Burk UC, et al: The
EMT-activator ZEB1 promotes tumorigenicity by repressing
stemness-inhibiting microRNAs. Nat Cell Biol. 11:1487–1495. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
46.
|
Torrisani J, Bournet B, du Rieu MC, et al:
let-7 microRNA transfer in pancreatic cancer-derived cells inhibits
in vitro cell proliferation but fails to alter tumor progression.
Hum Gene Ther. 20:831–844. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
47.
|
Ali S, Almhanna K, Chen W, Philip PA and
Sarkar FH: Differentially expressed miRNAs in the plasma may
provide a molecular signature for aggressive pancreatic cancer. Am
J Transl Res. 3:28–47. 2010.PubMed/NCBI
|
|
48.
|
Li Y, VandenBoom TG II, Kong D, et al:
Up-regulation of miR-200 and let-7 by natural agents leads to the
reversal of epithelial-to-mesenchymal transition in
gemcitabine-resistant pancreatic cancer cells. Cancer Res.
69:6704–6712. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
49.
|
Wicha MS, Liu S and Dontu G: Cancer stem
cells: an old idea - a paradigm shift. Cancer Res. 66:1883–1890;
discussion. 1895–1896. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
50.
|
Bonnet D and Dick JE: Human acute myeloid
leukemia is organized as a hierarchy that originates from a
primitive hematopoietic cell. Nat Med. 3:730–737. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
51.
|
Singh SK, Hawkins C, Clarke ID, et al:
Identification of human brain tumour initiating cells. Nature.
432:396–401. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
52.
|
Al-Hajj M, Wicha MS, Benito-Hernandez A,
Morrison SJ and Clarke MF: Prospective identification of
tumorigenic breast cancer cells. Proc Natl Acad Scie USA.
100:3983–3988. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
53.
|
O’Brien CA, Pollett A, Gallinger S and
Dick JE: A human colon cancer cell capable of initiating tumour
growth in immunodeficient mice. Nature. 445:106–110.
2007.PubMed/NCBI
|
|
54.
|
Li C, Heidt DG, Dalerba P, et al:
Identification of pancreatic cancer stem cells. Cancer Res.
67:1030–1037. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
55.
|
Hermann PC, Huber SL, Herrler T, et al:
Distinct populations of cancer stem cells determine tumor growth
and metastatic activity in human pancreatic cancer. Cell Stem Cell.
1:313–323. 2007. View Article : Google Scholar
|
|
56.
|
Moriyama T, Ohuchida K, Mizumoto K, et al:
Enhanced cell migration and invasion of CD133+
pancreatic cancer cells cocultured with pancreatic stromal cells.
Cancer. 116:3357–3368. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
57.
|
May R, Sureban SM, Lightfoot SA, et al:
Identification of a novel putative pancreatic stem/progenitor cell
marker DCAMKL-1 in normal mouse pancreas. Am J Physiol Gastrointest
Liver Physiol. 299:G303–310. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
58.
|
Sureban SM, May R, Lightfoot SA, et al:
DCAMKL-1 regulates epithelial-mesenchymal transition in human
pancreatic cells through a miR-200a-dependent mechanism. Cancer
Res. 71:2328–2338. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
59.
|
Tarasov V, Jung P, Verdoodt B, et al:
Differential regulation of microRNAs by p53 revealed by massively
parallel sequencing: miR-34a is a p53 target that induces apoptosis
and G1-arrest. Cell Cycle. 6:1586–1593. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
60.
|
Bommer GT, Gerin I, Feng Y, et al:
p53-mediated activation of miRNA34 candidate tumor-suppressor
genes. Curr Biol. 17:1298–1307. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
61.
|
Ji Q, Hao X, Zhang M, et al: MicroRNA
miR-34 inhibits human pancreatic cancer tumor-initiating cells.
PloS One. 4:e68162009. View Article : Google Scholar : PubMed/NCBI
|
|
62.
|
Ji Q, Hao X, Meng Y, et al: Restoration of
tumor suppressor miR-34 inhibits human p53-mutant gastric cancer
tumorspheres. BMC Cancer. 8:2662008. View Article : Google Scholar : PubMed/NCBI
|
|
63.
|
Bao B, Ali S, Banerjee S, et al: Curcumin
analogue CDF inhibits pancreatic tumor growth by switching on
suppressor microRNAs and attenuating EZH2 expression. Cancer Res.
72:335–345. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
64.
|
Wang Z, Li Y, Banerjee S and Sarkar FH:
Emerging role of Notch in stem cells and cancer. Cancer Lett.
279:8–12. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
65.
|
Demarest RM, Ratti F and Capobianco AJ:
It’s T-ALL about Notch. Oncogene. 27:5082–5091. 2008.
|
|
66.
|
Dotto GP: Notch tumor suppressor function.
Oncogene. 27:5115–5123. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
67.
|
Wang Z, Banerjee S, Li Y, Rahman KM, Zhang
Y and Sarkar FH: Down-regulation of notch-1 inhibits invasion by
inactivation of nuclear factor-kappaB, vascular endothelial growth
factor, and matrix metalloproteinase-9 in pancreatic cancer cells.
Cancer Res. 66:2778–2784. 2006. View Article : Google Scholar
|
|
68.
|
Strizzi L, Hardy KM, Seftor EA, et al:
Development and cancer: at the crossroads of Nodal and Notch
signaling. Cancer Res. 69:7131–7134. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
69.
|
Ristorcelli E and Lombardo D: Targeting
Notch signaling in pancreatic cancer. Expert Opin Ther Targets.
14:541–552. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
70.
|
Nalls D, Tang SN, Rodova M, Srivastava RK
and Shankar S: Targeting epigenetic regulation of miR-34a for
treatment of pancreatic cancer by inhibition of pancreatic cancer
stem cells. PloS One. 6:e240992011. View Article : Google Scholar : PubMed/NCBI
|
|
71.
|
Bao B, Wang Z, Ali S, et al: Notch-1
induces epithelial-mesenchymal transition consistent with cancer
stem cell phenotype in pancreatic cancer cells. Cancer Lett.
307:26–36. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
72.
|
Matthaios D, Zarogoulidis P, Balgouranidou
I, Chatzaki E and Kakolyris S: Molecular pathogenesis of pancreatic
cancer and clinical perspectives. Oncology. 81:259–272. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
73.
|
Feldmann G, Dhara S, Fendrich V, et al:
Blockade of hedgehog signaling inhibits pancreatic cancer invasion
and metastases: a new paradigm for combination therapy in solid
cancers. Cancer Res. 67:2187–2196. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
74.
|
Ferretti E, De Smaele E, Miele E, et al:
Concerted microRNA control of Hedgehog signalling in cerebellar
neuronal progenitor and tumour cells. EMBO J. 27:2616–2627. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
75.
|
Northcott PA, Fernandez LA, Hagan JP, et
al: The miR-17/92 polycistron is up-regulated in sonic
hedgehog-driven medulloblastomas and induced by N-myc in sonic
hedgehog-treated cerebellar neural precursors. Cancer Res.
69:3249–3255. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
76.
|
Friggi-Grelin F, Lavenant-Staccini L and
Therond P: Control of antagonistic components of the hedgehog
signaling pathway by microRNAs in Drosophila. Genetics.
179:429–439. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
77.
|
Tsuda N, Ishiyama S, Li Y, Ioannides CG,
Abbruzzese JL and Chang DZ: Synthetic microRNA designed to target
glioma-associated antigen 1 transcription factor inhibits division
and induces late apoptosis in pancreatic tumor cells. Clin Cancer
Res. 12:6557–6564. 2006. View Article : Google Scholar
|
|
78.
|
Yu S, Lu Z, Liu C, et al: miRNA-96
suppresses KRAS and functions as a tumor suppressor gene in
pancreatic cancer. Cancer Res. 70:6015–6025. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
79.
|
Vogt M, Munding J, Gruner M, et al:
Frequent concomitant inactivation of miR-34a and miR-34b/c by CpG
methylation in colorectal, pancreatic, mammary, ovarian,
urothelial, and renal cell carcinomas and soft tissue sarcomas.
Virchows Arch. 458:313–322. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
80.
|
Tashiro E, Tsuchiya A and Imoto M:
Functions of cyclin D1 as an oncogene and regulation of cyclin D1
expression. Cancer Sci. 98:629–635. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
81.
|
Lee KH, Lotterman C, Karikari C, et al:
Epigenetic silencing of MicroRNA miR-107 regulates cyclin-dependent
kinase 6 expression in pancreatic cancer. Pancreatology. 9:293–301.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
82.
|
Grewal IS and Flavell RA: CD40 and CD154
in cell-mediated immunity. Annu Rev Immunol. 16:111–135. 1998.
View Article : Google Scholar : PubMed/NCBI
|
|
83.
|
Costello RT, Gastaut JA and Olive D: What
is the real role of CD40 in cancer immunotherapy? Immunology Today.
20:488–493. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
84.
|
Mees ST, Mardin WA, Sielker S, et al:
Involvement of CD40 targeting miR-224 and miR-486 on the
progression of pancreatic ductal adenocarcinomas. Ann Surg Oncol.
16:2339–2350. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
85.
|
Kouzarides T: Histone acetylases and
deacetylases in cell proliferation. Curr Opin Genet Dev. 9:40–48.
1999. View Article : Google Scholar
|
|
86.
|
Ida K, Kitabayashi I, Taki T, et al:
Adenoviral E1A-associated protein p300 is involved in acute myeloid
leukemia with t(11;22) (q23;q13). Blood. 90:4699–4704.
1997.PubMed/NCBI
|
|
87.
|
Mees ST, Mardin WA, Wendel C, et al: EP300
- a miRNA-regulated metastasis suppressor gene in ductal
adenocarcinomas of the pancreas. Int J Cancer. 126:114–124. 2010.
View Article : Google Scholar : PubMed/NCBI
|