Introduction
Apoptosis is a mode of cell death in which single
cells are eliminated in the midst of living tissue. The term
derives from a Greek word that is used for the dropping off of
leaves from trees. It is characterized by structural changes that
appear with marked fidelity in cells of widely different lineage,
and presumably represent a pleiotropic effector response (1). Of the different mechanisms of cell
death, apoptosis has been proposed to explain the cell loss
observed in numerous neurological disorders, including Alzheimer’s
disease, vascular dementia (VaD), Parkinson’s disease, Huntington’s
disease, amyotrophic lateral sclerosis and stroke (2). VaD is the second most common type of
dementia following Alzheimer’s disease. When the blood supply to
the brain is reduced by a blocked or diseased vascular system, VaD
occurs and leads to a progressive decline in memory and cognitive
function (3). It is possible to
induce chronic cerebral hypoperfusion by permanent bilateral
occlusion of the common carotid arteries in rats, resulting in
significant white matter lesions, learning and memory impairment
(4), and hippocampal neuronal
damage (5). Clinical evidence
supports the hypothesis that chronic cerebral hypoperfusion is
associated with cognitive decline in aging and in neurodegenerative
disorders (6). Thus, permanent
bilateral occlusion of the common carotid arteries in rats provides
a model useful for understanding the pathophysiology of chronic
cerebrovascular hypoperfusion and for screening drugs with
potential therapeutic value for VaD (7).
Resveratrol, a polyphenolic compound in juice and
wine, was first reported in 1939 by the Japanese researcher, Dr
Michio Takaoka (8). It possesses a
wide range of biological effects including anti-apoptotic (9,10),
anti-oxidative (11),
anti-inflammatory (12) and
anti-carcinogenic properties (13). Resveratrol has been reported to
exhibit neuroprotective effects in the models of numerous diseases,
such as cerebral ischemia (14,15),
kainic acid-induced excitotoxicity (16), Huntington’s disease (17), Parkinson’s disease (18) and Alzheimer’s disease (19). However, data evaluating the effects
of resveratrol in VaD are lacking. In the present study, the
permanent, bilateral common carotid artery occlusion rat model was
used to attain information on the effects of resveratrol on
VaD.
Materials and methods
Animals
Wistar rats (provided by the Henan Laboratory Animal
Research Center, Zhengzhou, China), aged 12–14 months, weighing
300–400 g and of unlimited gender, were used in the study. The rats
were housed in standard cages with liquid and food available ad
libitum, at a mean ± standard deviation (SD) constant
temperature of 22±2°C, humidity of 55±5% and under an artificial
reversed 12-h light-dark cycle with the light off at 7.00 a.m. All
procedures were conducted in accordance with the Guidance
Suggestions for the Care and Use of Laboratory Animals, formulated
by the Ministry of Science and Technology of the People’s Republic
of China (20).
Methods
Experimental design
The rats were randomly divided into four groups
(n=20/group) as follows: Group A, normal control group in which the
rats were subjected to sham surgery; group B, VaD model group in
which the rats underwent VaD-modeling surgery; group C, resveratrol
control group in which the rats were subjected to sham surgery and
treated with resveratrol; group D, treatment group in which the
rats underwent VaD-modeling surgery and were treated with
resveratrol. The models of VaD were established by permanent
bilateral occlusion of the common carotid arteries in groups B and
D. The bilateral common carotid arteries were isolated but not
ligated in groups A and C. The rats of groups C and D received a
daily oral dose of 25 mg/kg resveratrol (obtained from
Sigma-Aldrich, St. Louis, MO, USA) starting from 8 weeks after the
surgery until 12 weeks after the surgery; and the rats of groups A
and C received the same volume of ethanol. The study was approved
by the Committee on Ethics of Life Sciences of the First Affiliated
Hospital of Zhengzhou University (Zhengzhou, China).
Bilateral common carotid
occlusion
The bilateral common carotid arteries of the rats
were occluded as previously described by Ni et al (4). The rats were anesthetized with a 10%
chloral hydrate (0.3 ml/100 g; Sigma-Aldrich) intraperitoneal
injection. To prevent respiratory distress, the rats were also
administered atropine sulfate (0.1 mg/kg, intramuscularly; Polfa
Warszawa S.A., Warsaw, Poland). A midline incision was made to
expose the bilateral common carotid arteries. The common carotid
arteries were carefully separated from the surrounding tissues,
including the vagus nerve, and ligated with Ethicon Coated Vicryl
(polyglactin 910) plus antibacterial absorbable surgical suture
(size 3-0; Johnson & Johnson Medical Ltd., Wokingham, UK), ~1
cm inferior to the origin of the external carotid artery. The
control rats were subjected to the same surgical procedure without
occlusion of the arteries.
Morris water maze test
The spatial learning and memory performance of the
rats was measured using the Morris water maze task (provided by
Chinese Academy of Medical Sciences, Beijing, China). The test was
administered by an operator blinded to the group conditions. The
Morris water maze consisted of a painted circular pool (120 cm in
diameter and 30 cm in depth) in which the rats were trained to
escape from the water by swimming to a hidden platform (9 cm in
diameter) 1.5 cm beneath the surface, the location of which was
only identifiable using distal extra-maze cues attached to the room
walls. The water was maintained at 22°C and made opaque with
titanium dioxide throughout all training and testing. The pool was
divided into four quadrants: North (Target), south (Opposite), east
(Adjacent 1) and west (Adjacent 2). The experiments were recorded
using a camera connected to a video recorder and a computerized
tracking system (Shanghai Jiliang Software Technology Co. Ltd.,
Shanghai, China).
The Morris water maze test procedure was conducted
as previously reported (21). The
first 4 days were the reference memory test phase, which consisted
of 16 training trials: 4 training trials per day for 4 training
days with an inter-trial interval of 30–40 min. At the beginning of
each trial, the rat was placed into one of the four quadrants
facing the wall. Although the starting point was randomly selected,
the protocol was fixed at the beginning of each trial and was
maintained throughout all trials. Each rat was allowed 180 sec to
locate and mount the platform; 30 sec after the rat mounted the
platform, it was removed, placed in a holding cage and warmed with
a heat lamp. The rats that failed to locate and mount the platform
within 180 sec were gently guided to the platform and required to
remain there for 30 sec prior to being transferred to the holding
cage. A video camera mounted above the pool was used to track the
rats. The amount of time spent locating and mounting the platform
(escape latency) and the swimming pathway prior to locating the
platform (escape distance) were calculated from the recorded videos
using Morris water maze software (Shanghai Jiliang Software
Technology Co. Ltd., Shanghai, China). On the fifth day, a probe
test was performed in which the platform was removed. The rats were
placed into the water in one of the two quadrants adjacent to the
platform (Adjacent 1 and Adjacent 2 quadrants) and were allowed to
swim freely for 120 sec. The percentage of time spent and the
swimming distance percentage in the target quadrant were
recorded.
Collection and preservation of brain
tissues
The rats were anesthetized with diethyl ether and
then perfused with phosphate buffer saline (pH 7.4). The brain of
each rat was immediately removed from the skull, and the
hippocampus was dissected on ice. All brain tissues were stored at
−80°C until biochemical analysis.
Western blot analysis
Western blot analysis was performed using the
hippocampus of each rat, which had been dissected and stored at
−80°C. The brain tissues were homogenized with lysis buffer [10 mM
Tris pH 7.4, 100 mM NaCl, 1 mM ethylenediamine-N,N,N′,
N′-tetraacetic acid (EDTA), 1 mM
ethyleneglycol-bis(2-aminoethyl)-N,N,N′, N′-tetraacetic acid
(EGTA), 1 mM NaF, 20 mM Na4P2O7, 2
mM Na3VO4, 0.1% sodium dodecyl sulfate (SDS),
0.5% sodium deoxycholate, 1% Triton-X 100, 10% glycerol, 1 mM
phenylmethylsulfonyl fluoride (PMSF; made from a 0.3 M stock in
dimethylsulfoxide), 60 μg/ml aprotinin, 10 μg/ml leupeptin, and 1
μg/ml pepstatin] for 30 min. The soluble fraction was obtained by
centrifugation at 2,500 × g for 20 min at 4°C. The concentration of
the protein was determined using a BCA assay (Pierce Biotechnology,
Inc., Rockford, IL, USA). The western blotting procedure was
conducted as previously reported (22). Equal amounts of protein (20 μg)
were boiled at 100°C for 10 min in loading buffer (Fermentas,
Beijing, China) and were separated in 8–10% SDS-polyacrylamide gel,
and the resolved proteins were electrotransferred to polyvinylidene
difluoride membranes (Bio-Rad, Hercules, CA, USA). Subsequently,
the membranes were blocked with 5% non-fat milk in TBST (10 mM
Tris-HCl pH 8.0, 150 mM NaCl and 0.2% Tween-20) for 1 h at room
temperature and incubated with the appropriate primary antibody
[1:200 Bax and Bcl-2; Santa Cruz Biotechnology, Inc., Santa Cruz,
CA, USA; 1:5,000 β-actin, Sigma-Aldrich; and 1:1,000 cleaved
caspase-3 and poly(ADP-ribose) polymerase (PARP), Cell Signaling
Technology Inc., Beverly, MA, USA] at 4°C overnight. The membranes
were then washed twice with TBST and probed with the corresponding
secondary antibodies conjugated with horseradish peroxidase (HRP)
(anti-mouse/rabbit-HRP was used at a dilution of 1:5,000) at room
temperature for 1 h. The membranes were challenged with Vectastain
ABC agent (Vector Laboratories, Burlingame, CA, USA). After
washing, the signals were developed with a ECL Advance Western
Blotting Detection kit (Amersham Pharmacia Biotech,
Buckinghamshire, UK). The blots were stripped and reprobed with
anti-β-actin as a loading control. The band intensities were
quantified by densitometric analyses using an AxioCam digital
camera and the KS400 photo analysis system, version 3.0 (Carl
Zeiss, Oberkochen, Germany).
Statistical analysis
Data are expressed as the mean ± SD and were
analyzed using SPSS statistical software, version 16.0 (SPSS, Inc.,
Chicago, IL, USA). Each procedure was performed in duplicate in
three to five independent experiments. Statistical analyses were
performed using one-way analysis of variance followed by two-tailed
Student’s t-test, and statistical significance was assumed at
P<0.05.
Results
Effects of resveratrol on memory
impairment in VaD rat models
The Morris water maze was used to test the spatial
learning and memory performance of the rats by measuring the escape
latency (the amount of time spent locating and mounting the
platform in the water maze) and escape distance (the swimming
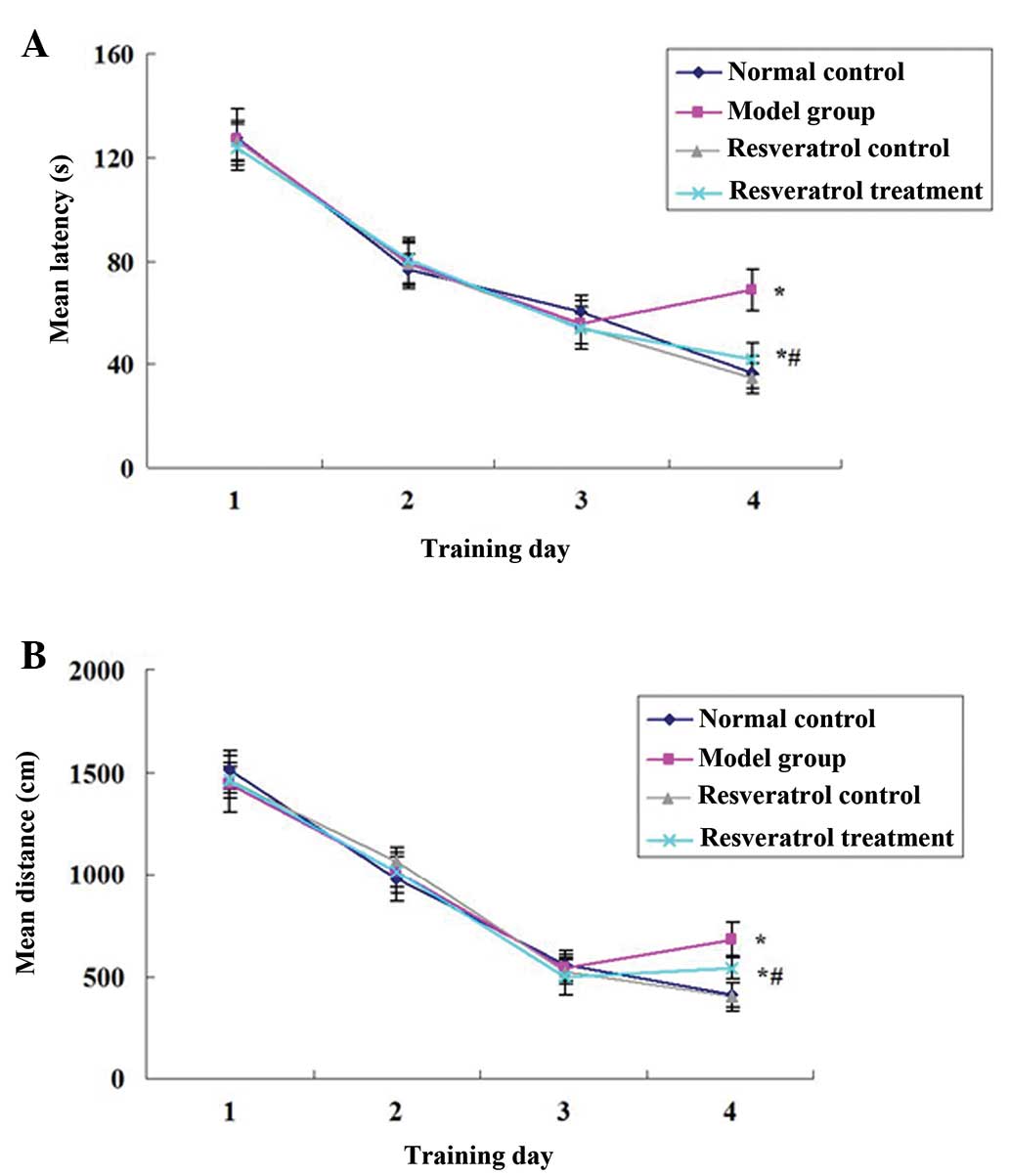
pathway prior to locating the platform in the water maze). In the
first three days, no differences were identified in the escape
latency (Fig. 1A; P>0.05) and
escape distance (Fig. 1B;
P>0.05) of any of the groups. On the fourth day, the escape
latency and escape distance of the resveratrol control group were
not significantly different compared with those of the normal
control group (P>0.05), those of the model group were
significantly higher than those of the normal control group
(P<0.05), and all of the changes were partly attenuated by the
resveratrol treatment. The escape latency and escape distance of
the rats in the resveratrol treatment group were significantly
shorter than those of the rats in the model group (P<0.05;
Fig. 1).
Following the water maze training test, a probe test
was performed to analyze the maintenance of memory, in which the
platform was removed and the percentage of time spent and of the
swimming distance in the target quadrant were recorded. During the
probe test, the percentage of time spent (Fig. 2A) and of the swimming distance
(Fig. 2B) in the target quadrant
of the resveratrol control group were not significantly different
compared with those of the normal control group (P>0.05), and
those of the model group rats were significantly shorter than those
of the normal control group rats (P<0.05), and the changes were
partly attenuated by the resveratrol treatment. The percentage of
time spent (Fig. 2A) and of the
swimming distance (Fig. 2B) of the
rats in the resveratrol treatment group were significantly longer
than those of the rats in the model group (P<0.05; Fig. 2).
Effects of resveratrol on Bax/Bcl-2 in
the brains of VaD rat models
Western blot analysis of the hippocampus lysates was
performed using Bax and Bcl-2 antibodies. It was identified that
the expression levels of Bcl-2 and Bax in the resveratrol control
group were not significantly different compared with those of the
normal control group (P>0.05). The expression levels of Bax were
significantly increased, the expression levels of Bcl-2 were
significantly reduced and the ratio of Bax/Bcl-2 was significantly
increased in the model group rats compared with those of the normal
control group (P<0.05). By contrast, the expression levels of
Bax were significantly reduced, the expression levels of Bcl-2 were
significantly increased and the Bax/Bcl-2 ratio was significantly
decreased in the resveratrol treatment group rats compared with
those of the model group rats (P<0.05; Fig. 3).
Effects of resveratrol on the expression
of cleaved caspase-3 in the brains of VaD rat models
Western blot analysis of the hippocampus lysates was
performed using a cleaved caspase-3 antibody. It was identified
that the expression of cleaved caspase-3 in the resveratrol control
group was not significantly different compared with that of the
normal control group (P>0.05), and the expression levels of
cleaved caspase-3 were significantly increased in the model group
rats compared with those of the normal control group rats
(P<0.05). By contrast, the expression levels of cleaved
caspase-3 were significantly reduced in the resveratrol treatment
group rats compared with those of the model group rats (P<0.05;
Fig. 4).
Effects of resveratrol on the expression
of cleaved PARP in the brains of VaD rat models
Western blot analysis of the hippocampus lysates was
performed using a cleaved PARP antibody. It was identified that the
expression of cleaved PARP in the resveratrol control group was not
significantly different compared with that of the normal control
group (P>0.05), and the expression levels of cleaved PARP were
significantly increased in the model group rats compared with those
of the normal control group rats (P<0.05). By contrast, the
expression levels of cleaved PARP were significantly reduced in the
resveratrol treatment group rats compared with those of the model
group rats (P<0.05; Fig.
5).
Discussion
Coinciding with population aging and improved
survival from cardiovascular diseases and stroke, VaD is more
frequent and is likely to affect an increasing number of patients
in the future (23).
Etiopathogenic mechanisms leading to VaD include oxidative stress,
cytotoxicity of reactive oxygen species, mitochondrial dysfunction
and apoptosis (24,25). Resveratrol has been proposed as a
major constituent of the polyphenol fraction to which the health
benefits of red wine consumption are attributed. In vivo and
in vitro studies have shown that resveratrol exhibits
neuroprotective effects in the models of numerous diseases. In the
present study, the permanent, bilateral common carotid artery
occlusion rat model was used to study the effects of resveratrol on
VaD. Using a Morris water maze test, it was identified that the
escape latency and escape distance (which are the time and distance
travelled to reach the platform in the water maze) of the model
group rats were significantly higher than those of the normal
control group rats (P<0.05; Fig.
1). A probe test also showed that the percentage of time spent
(Fig. 2A) and of the swimming
distance (Fig. 2B) in the target
quadrant of the model group rats were significantly shorter than
those of the normal control group rats (P<0.05; Fig. 2). All of these changes were partly
attenuated by the resveratrol treatment (P<0.05; Fig. 1 and 2) which suggests that resveratrol
ameliorates the memory impairment in VaDmodel rats.
Apoptosis is a tightly regulated process, which
involves changes in the expression of a distinct set of genes. Two
of the major genes responsible for regulating apoptotic cell death
are Bcl-2 and Bax. Bcl-2 is a key member of the anti-apoptotic
Bcl-2 family that is crucial in regulating mitochondrial-mediated
apoptotic cell death. Overexpression of Bcl-2 has been demonstrated
to protect neuronal cells from neurotoxic insult (26). By contrast, Bax belongs to the
pro-survival subfamily, which promotes apoptosis by translocation
into the mitochondrial membrane and facilitating cytochrome
c release, to propagate downstream apoptotic events
(27). An elevated intracellular
ratio of Bax to Bcl-2 occurs during increased apoptotic cell death
(28,29). In the present study, it was
demonstrated that the expression levels of Bax were significantly
increased, the expression levels of Bcl-2 were significantly
reduced and the ratio of Bax/Bcl-2 was significantly increased in
the model group rats compared with those of the normal control
group rats (P<0.05; Fig. 3). By
contrast, the expression levels of Bax were significantly reduced,
the expression levels of Bcl-2 were significantly increased and the
ratio of Bax/Bcl-2 was significantly decreased in the resveratrol
treatment group rats compared with those of the model group rats
(P<0.05; Fig. 3).
Caspases are a family of cysteine proteases and are
critical mediators of cell apoptosis. Caspases are important in the
apoptotic process by two distinct pathways: The death receptor
pathway and the mitochondrial pathway (30). Whichever pathway is involved,
caspase-3 acts as an apoptotic executor. Caspase-3 activates DNA
fragmentation factor, which in turn activates endonucleases to
cleave nuclear DNA, and ultimately leads to cell death (31). Activation of caspase-3 appears to
be a key event in the execution of the apoptotic cascade in
numerous central nervous system diseases, such as Alzheimer’s
disease and Down’s syndrome. In the present study, it was also
identified that the expression levels of cleaved caspase-3 were
significantly increased in the model group rats compared with those
of the normal control group rats (P<0.05). By contrast, the
expression levels of cleaved caspase-3 were significantly reduced
in the resveratrol treatment group rats compared with those of the
model group rats (P<0.05; Fig.
4).
Significant evidence indicates that caspase-3 is
partially or totally responsible for the proteolytic cleavage of
numerous key proteins, including PARP (32). PARP is a nuclear DNA-binding
protein of 113 kDa that is constitutively expressed in eukaryotes
and that comprises up to 1% of the total nuclear proteins (33). PARP is important for cell
viability, and cleavage of PARP facilitates cellular disassembly
and serves as a marker of cells undergoing apoptosis (34). In the present study, it was
identified that the expression levels of cleaved PARP were
significantly increased in the model group rats compared with those
of the normal control group (P<0.05). By contrast, the
expression levels of cleaved PARP were significantly reduced in the
resveratrol treatment group rats compared with those of the model
group rats (P<0.05; Fig.
5).
In conclusion, in the present study it was
demonstrated that resveratrol was able to inhibit memory impairment
in a VaD rat model. This effect was associated with attenuation of
the increased expression levels of Bax, cleaved caspase-3 and
cleaved PARP and decreased expression levels of Bcl-2 that were
induced in the VaD model. These results confirm the neuroprotective
effects of resveratrol on VaD and provide a novel insight into the
neuroprotective effects of resveratrol and its possible therapeutic
role in VaD.
References
|
1
|
Wyllie AH: Apoptosis (the 1992 Frank Rose
Memorial Lecture). Br J Cancer. 67:205–208. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Favaloro B, Allocati N, Graziano V, et al:
Role of apoptosis in disease. Aging (Albany NY). 4:330–349.
2012.
|
|
3
|
Jellinger KA: The enigma of vascular
cognitive disorder and vascular dementia. Acta Neuropathol.
113:349–388. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Ni J, Ohta H, Matsumoto K and Watanabe H:
Progressive cognitive impairment following chronic cerebral
hypoperfusion induced by permanent occlusion of bilateral carotid
arteries in rats. Brain Res. 653:231–236. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Kim SK, Cho KO and Kim SY: White matter
damage and hippocampal neurodegeneration induced by permanent
bilateral occlusion of common carotid artery in the rat: Comparison
between Wistar and Sprague-Dawley strain. Korean J Physiol
Pharmacol. 12:89–94. 2008. View Article : Google Scholar
|
|
6
|
Cechetti F, Worm PV, Pereira LO, et al:
The modified 2VO ischemia protocol causes cognitive impairment
similar to that induced by the standard method, but with a better
survival rate. Braz J Med Biol Res. 43:1178–1183. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Gong X, Ma M, Fan X, et al:
Down-regulation of IGF-1/IGF-1R in hippocampus of rats with
vascular dementia. Neurosci Lett. 513:20–24. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Takaoka M: Resveratrol, a new phenolic
compound from Veratrum grandiflorum. Nippon Kagaku Kaishi.
60:1090e1001939.
|
|
9
|
Shen M, Wu RX, Zhao L, et al: Resveratrol
attenuates ischemia/reperfusion injury in neonatal cardiomyocytes
and its underlying mechanism. PLoS One. 7:e512232012. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Ku CR, Lee HJ, Kim SK, et al: Resveratrol
prevents streptozotocin-induced diabetes by inhibiting the
apoptosis of pancreatic β-cell and the cleavage of poly
(ADP-ribose) polymerase. Endocr J. 59:103–109. 2012.PubMed/NCBI
|
|
11
|
Robb EL, Page MM, Wiens BE and Stuart JA:
Molecular mechanisms of oxidative stress resistance induced by
resveratrol: Specific and progressive induction of MnSOD. Biochem
Biophys Res Commun. 367:406–412. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Bo S, Ciccone G, Castiglione A, et al:
Anti-inflammatory and antioxidant effects of resveratrol in healthy
smokers a randomized, double-blind, placebo-controlled, cross-over
trial. Curr Med Chem. 20:1323–1331. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Athar M, Back JH, Kopelovich L, et al:
Multiple molecular targets of resveratrol: Anti-carcinogenic
mechanisms. Arch Biochem Biophys. 486:95–102. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Simão F, Matté A, Breier AC, et al:
Resveratrol prevents global cerebral ischemia-induced decrease in
lipid content. Neurol Res. 35:59–64. 2013.PubMed/NCBI
|
|
15
|
Li Z, Pang L, Fang F, et al: Resveratrol
attenuates brain damage in a rat model of focal cerebral ischemia
via up-regulation of hippocampal Bcl-2. Brain Res. 1450:116–124.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Wu Z, Xu Q, Zhang L, et al: Protective
effect of resveratrol against kainate-induced temporal lobe
epilepsy in rats. Neurochem Res. 34:1393–1400. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Pasinetti GM, Wang J, Marambaud P, et al:
Neuroprotective and metabolic effects of resveratrol: therapeutic
implications for Huntington’s disease and other neurodegenerative
disorders. Exp Neurol. 232:1–6. 2011.PubMed/NCBI
|
|
18
|
Khan MM, Ahmad A, Ishrat T, et al:
Resveratrol attenuates 6-hydroxydopamine-induced oxidative damage
and dopamine depletion in rat model of Parkinson’s disease. Brain
Res. 1328:139–151. 2010.PubMed/NCBI
|
|
19
|
Li F, Gong Q, Dong H and Shi J:
Resveratrol, a neuroprotective supplement for Alzheimer’s disease.
Curr Pharm Des. 18:27–33. 2012.
|
|
20
|
Guidance suggestions for the care and use
of laboratory animals. The Ministry of Science and Technology of
the People’s Republic of China; Beijing: 2006
|
|
21
|
Ma XR, Sun ZK, Liu YR, et al: Resveratrol
improves cognition and reduces oxidative stress in rats with
vascular dementia. Neural Regen Res. 8:2050–2059. 2013.PubMed/NCBI
|
|
22
|
Sun ZK, Yang HQ, Pan J, et al: Protective
effects of erythropoietin on tau phosphorylation induced by
beta-amyloid. J Neurosci Res. 86:3018–3027. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Levine DA and Langa KM: Vascular cognitive
impairment: disease mechanisms and therapeutic implications.
Neurotherapeutics. 8:361–373. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Bennett S, Grant MM and Aldred S:
Oxidative stress in vascular dementia and Alzheimer’s disease: a
common pathology. J Alzheimers Dis. 17:245–257. 2009.
|
|
25
|
Wang J, Zhang HY and Tang XC: Cholinergic
deficiency involved in vascular dementia: possible mechanism and
strategy of treatment. Acta Pharmacol Sin. 30:879–888. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Gustafsson AB and Gottlieb RA: Bcl-2
family members and apoptosis, taken to heart. Am J Physiol Cell
Physiol. 292:C45–C51. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Wolter KG, Hsu YT, Smith CL, et al:
Movement of Bax from the cytosol to mitochondria during apoptosis.
J Cell Biol. 139:1281–1292. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Clementi ME, Pezzotti M, Orsini F, et al:
Alzheimer’s amyloid beta-peptide (1-42) induces cell death in human
neuroblastoma via bax/bcl-2 ratio increase: an intriguing role for
methionine 35. Biochem Biophys Res Commun. 342:206–213. 2006.
|
|
29
|
Sun ZK, Yang HQ, Wang ZQ, et al:
Erythropoietin prevents PC12 cells from beta-amyloid-induced
apoptosis via PI3K/Akt pathway. Transl Neurodegener. 1:72012.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Grütter MG: Caspases: key players in
programmed cell death. Curr Opin Struct Biol. 10:649–655. 2000.
|
|
31
|
Lee MK, Kang SJ, Poncz M, et al:
Resveratrol protects SH-SY5Y neuroblastoma cells from apoptosis
induced by dopamine. Exp Mol Med. 39:376–384. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Alvarez-Gonzalez R, Spring H, Müller M and
Bürkle A: Selective loss of poly(ADP-ribose) and the 85-kDa
fragment of poly(ADP-ribose) polymerase in nucleoli during
alkylation-induced apoptosis of HeLa cells. J Biol Chem.
274:32122–32126. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Lee DH, Park T and Kim HW: Induction of
apoptosis by disturbing mitochondrial-membrane potential and
cleaving PARP in Jurkat T cells through treatment with
acetoxyscirpenol mycotoxins. Biol Pharm Bull. 29:648–654. 2006.
View Article : Google Scholar
|
|
34
|
Oliver FJ, de la Rubia G, Rolli V, et al:
Importance of poly(ADP-ribose) polymerase and its cleavage in
apoptosis. Lesson from an uncleavable mutant. J Biol Chem.
273:33533–33539. 1998. View Article : Google Scholar : PubMed/NCBI
|