1. Introduction
Metastasis is the most important biological
characteristic of malignant tumors and is currently the main cause
of cancer-associated mortality. Cancer invasion and metastasis are
two essential steps in the complex tumor development process and
result from a series of biological changes in cancer and stromal
cells (1). Firstly, cancer cells
acquire the movement characteristics of epithelial mesenchymal
cells, which is accompanied by a decrease in intercellular
adhesion. During this period, the extracellular matrix adhesion
abilities of cancer cells are decreased. Therefore, cancer cells
depart from the primary cancer tumor and are transported through
membranes easily. The cancer cells then enter the vascular or
lymphatic vessels. Finally, the new proliferated high-colony cells
are established in the distant sites (2). In previous years, studies have found
that the Wnt signaling pathway is closely associated with other
signaling cellular elements, including adhesion molecules,
proteases and angiogenic factors, during the cancer metastasis
process (3,4).
2. Wnt5A is involved in noncanonical Wnt
signaling
WNT genes encode a family of 38–43 kDa
glycoproteins, which are growth factors typically secreted by
cells. Wnt proteins were first identified in mammals as the
proto-oncogenic integration site for the mouse mammary tumor virus.
These proteins include a hydrophobic signal peptide, highly
conserved cysteine residues and N-linked glycosylation sites, with
the absence of additional transmembrane domains (5). There are common characteristics in
the primary structure of the Wnt family proteins, including a
signal sequence for secretion, glycosylation sites essential for
activity and a cysteine-rich domain that is involved in protein
folding. These proteins associate with cell surfaces and the
extracellular matrix, and a number are known to bind to the
Frizzled family of receptors (Fzd) that resemble typical G
protein-coupled receptors (6). To
date, at least 19 Wnt members have been shown to be present in
human and mice, with functions contributing to the regulation of a
wide range of cellular processes, particularly in development. Wnt
proteins have also been implicated in tumor formation. The WNT5A
gene was first identified in mice, with multiple transcripts
encoding a cysteine-rich protein consisting of 379 amino acids,
whose temporal expression correlated with the spatial patterning
and morphogenesis (7). Clark et
al (8) reported the cloning
and sequencing of several overlapping cDNA molecules that encoded
the 4.1 kb human homologue of Wnt5a. The mature human Wnt5a protein
contains 343 residues and has >93% homology to the reported
sequences of other Wnt5a proteins (>99% homologous to mouse
Wnt5a). Using a combination of Southern blotting, polymerase chain
reaction amplification and in situ hybridization, the human
WNT5A gene was mapped to chromosome 3p14-p21 (9).
Wnt proteins activate a number of signaling
pathways, which can be divided into two general categories; the
canonical β-catenin pathway and the noncanonical β-catenin
independent pathways. In the canonical pathway, a Wnt protein (such
as Wnt1, Wnt3a and Wnt7a) binds to the receptors and triggers a
cascade that is mediated by dishevelled proteins, inhibiting
glycogen synthase kinase-3β (GSK-3β) activity. Inactivation of
GSK-3β results in the hypophosphorylation of β-catenin, which
escaped from the complex that formed with adenomatous polyposis
coli (APC) and Axin, and degraded by the ubiquitin/proteasome
system. The accumulated free β-catenin enters the nucleus,
activates the Tcf/Lef transcription factors and in turn triggers
the transcription of a set of target genes, ultimately leading to
the regulation of cell proliferation and cell apoptosis, as well as
cell transformation (10,11).
Wnt5a is a representative ligand that activates the
noncanonical Wnt signaling pathways, which are defined as being
independent from the β-catenin pathway. Noncanonical Wnt cascades
are diverse and in a number of cases, hard to define. The pathways
are classified into the following categories for clarity and
simplicity: i) Wnt/planar cell polarity signaling; ii) Wnt-cyclic
guanosine monophosphate/Ca2+ signaling; iii) Wnt-RAP1
signaling; iv) Wnt-receptor tyrosine kinase-like orphan receptor 2
(ROR2) signaling; v) Wnt-protein kinase A signaling; vi)
Wnt-GSK-3-microtubule signaling; vii) Wnt-atypical protein kinase C
(PKC) signaling; viii) Wnt-receptor-like tryosine kinase signaling;
and ix) Wnt-mammalian target of rapamycin signaling (12). These classifications are not rigid
since the pathways overlap and intersect with one another and are
evolving. Wnt5a is involved in various cellular functions by
activating multiple signaling pathways. However, the role of Wnt5a
in cancer metastasis appears to be more complex. Nevertheless, a
previous study (13) indicated
that Wnt5a plays a key role in malignant progression, although
whether Wnt5a exhibits a tumor metastasis-suppressing effect or a
promoting effect remains unclear.
3. Metastasis-promoting activity of Wnt5a
and the underlying mechanisms
WNT5A has been identified as an oncogene in numerous
types of tumors. Kurayoshi et al (14) detected the expression of Wnt5a in
237 cases of primary gastric cancer via immunohistochemistry. The
results revealed that the expression of Wnt5a was upregulated in
30% (71/237) of patients with gastric cancer, which positively
correlated with the T grade (depth of invasion) and N grade (degree
of lymph node metastasis). The positive rate of Wnt5a expression in
stage III/IV gastric cancer tissues (49.5%) was significantly
higher than that in stage I/II gastric cancers (13.8%). These
results indicated that the upregulation of Wnt5a may be associated
with cancer infiltration and lymph node metastasis. In addition,
the positivity of Wnt5a expression correlated with advanced stages
and poor prognosis of gastric cancer. The authors further
demonstrated that Wnt5a had the ability to stimulate cell migration
and invasion in gastric cancer cells. Cell migration, membrane
ruffling and turnover of paxillin were suppressed in WNT5A
knockdown cells. Wnt5a activated focal adhesion kinase and small
GTP-binding protein Rac, both of which are known to play a role in
cell migration. Laminin γ is also required for liver metastasis of
gastric cancer cells in vivo. A recent study showed that an
anti-Wnt5a polyclonal antibody (pAb5a-5) inhibited the activation
of Rac1 and the expression of laminin γ2, which decreased
Wnt5a-dependent internalization of the receptor, thereby
suppressing the metastasis of gastric cancer cells (15). In addition, Wnt5a promoted the
migration of gastric cancer cells via the phosphatidylinositol
3-kinase/protein kinase B/GSK-3β/RhoA signaling pathway (16). Therefore, the expression of Wnt5a
in gastric cancer cells may be critical for the migration and
invasion of cancer cells from primary regions. Thus, the expression
of Wnt5a correlates with the aggressiveness of the gastric
cancer.
Gene expression profiling has indicated that WNT5A
may be a marker of aggressive in melanomas (17). Immunohistochemical analyses
revealed that Wnt5a staining was negative in the majority of benign
tumors and was heterogeneously distributed among primary melanomas,
while the majority of metastatic tumors were stained strongly
positive for Wnt5a. Wnt5a overexpression correlates significantly
with the survival and the development of metastases in melanomas.
Da Forno et al (18)
detected the expression levels of Wnt5a and p16ink4a in 59 cases of
primary melanoma and their matched metastatic tissues. With tumor
progression, the expression of Wnt5a in the cytoplasm gradually
increased, however, the expression of p16ink4a was reduced,
indicating that overexpression of Wnt5a in the cytoplasm was
positively correlated with the progress of the melanoma, as well as
poor prognosis. Recently, Grossmann et al (19) identified a novel mechanism of
action for Wnt5a, by which Wnt5a binds to the cellular Fzd 4-LRP6
receptor complex and activates ADP-ribosylation factor 6, thus,
inducing the disruption of the N-cadherin and β-catenin complex and
resulting in the enhancement of melanoma invasion and
metastasis.
The homeodomain transcription factor, CUTL1,
functions as a target of transforming growth factor-β and an
important mediator via its effects on cell migration and
invasiveness (20). In pancreatic
carcinomas, Wnt5a is upregulated by CUTL1 on an mRNA and protein
level. Increased activity of a luciferase construct containing the
putative Wnt5a promoter upon CUTL1 overexpression indicates that
CUTL1 transcriptionally activates Wnt5a. In addition, Wnt5a is a
crucial downstream mediator of tumor cell migration, invasion and
proliferation induced by CUTL1. Wnt5a is upregulated early during
pancreatic carcinogenesis in pancreatic intraepithelial neoplasias
lesions and in invasive pancreatic adenocarcinomas, as compared
with normal pancreatic tissues. The effects of Wnt5a are also
accompanied with epithelial-mesenchymal transition (EMT). These
results indicate that WNT5A is an important target of CUTL1 and
functions as a novel mediator of invasiveness and tumor progression
in pancreatic cancer (21).
Certain studies (22–23)
have clarified the association between Wnt5a and tumor metastasis
from the aspects of cell polarization and directional movements,
since a shared feature of a number of noncanonical Wnt pathways is
the targeting of the cytoskeleton, with important implications for
cancer metastasis. Witze et al (24) investigated the mechanism of how
Wnt5a polarizes the cytoskeleton to promote directional motility in
cultured melanoma cells. In response to Wnt5a, several proteins,
including a melanoma cell adhesion molecule, a Fzd receptor and
cytoskeletal components (action microfilaments and a nonmuscle
myosin, which are together called actomysin), become polarized in
overlapping bands within the trailing edge of the migrating cells.
Polarized bands of actomyosin contract to lift the trailing edge,
with cell process extensions at the leading edge, and then promote
migration toward a chemokine source. Wnt5a functions through the
small guanosine triphosphatase (GTPase), RhoB, which controls the
movement of intracellular vesicles called endosomes. The authors
also observed that an enrichment of multivesicular bodies existed
in the polarized cell edge, and polarization required the Rab4
GTPase (known to regulate multivesicular body formation) and
dynamin (known to control the formation of endosomes at the cell
surface). In conclusion, Wnt5a polarizes cells by promoting the
recycling of membrane components to specific surface sites. The
concurrent or subsequent recruitment of actomyosin and other
factors to these sites defines the trailing edge of the cell, thus,
the direction of migration. However, the mechanisms underlying the
coupling of membrane polarization to the cytoplasm and the
targeting of various proteins to the same sites remain unclear. In
addition, it remains unknown how Wnt5a signaling is interfaced with
the graded chemokines to ensure the polarization occurs at the cell
surface distal to the chemokine source remains unknown (25).
An early study of gene expression profiling found
that Wnt5a/PKC signaling was associated with aggressive melanoma
behavior (17). The Weeraratna
group (26) used microarray
analysis to demonstrate that Wnt5a directly promoted cell invasion
and motility via the inhibition of metastasis-associated gene
expression (KISS and CD44) and the stimulation of EMT (a key step
in metastasis) in a PKC-dependent manner. Nomachi et al
(27) found that ROR2, an orphan
receptor belonging to the ROR family of receptor tyrosine kinases,
played critical roles in Wnt5a-induced cell migration by regulating
the formation of lamellipodia and the reorientation of the
microtubule-organizing center (MTOC). ROR2 possesses an
extracellular cysteine-rich domain that resembles the Wnt-binding
sites of the Fzd proteins, thus, it is possible that ROR2 interacts
with members of the Wnt family (28). RORs play critical roles in
enhancing cell invasion and migration in multiple cancer types
(29). Wnt5a stimulation induces
the activation of c-Jun N-terminal kinase (JNK) at the wound edge
in a ROR2-dependent manner, and inhibiting JNK activity abrogates
Wnt5a-induced lamellipodia formation and MTOC reorientation.
Additionally, the association of ROR2 with the actin-binding
protein filamin A is required for Wnt5a-induced JNK activation and
polarized cell migration. Thus, JNK may play roles in cell
migration and invasion by phosphorylating the focal
adhesion-associated protein, paxillin (30). Further study revealed that
Wnt5a-induced JNK activation and MTOC reorientation can be
suppressed by inhibiting PKC (31). Therefore, Wnt5a exhibits
metastasis-promoting activities in several types of cancer through
different signaling pathways (Fig.
1). These novel findings demonstrate the diversity of the
noncanonical signaling pathways, as well as further the
understanding of tumor metastasis.
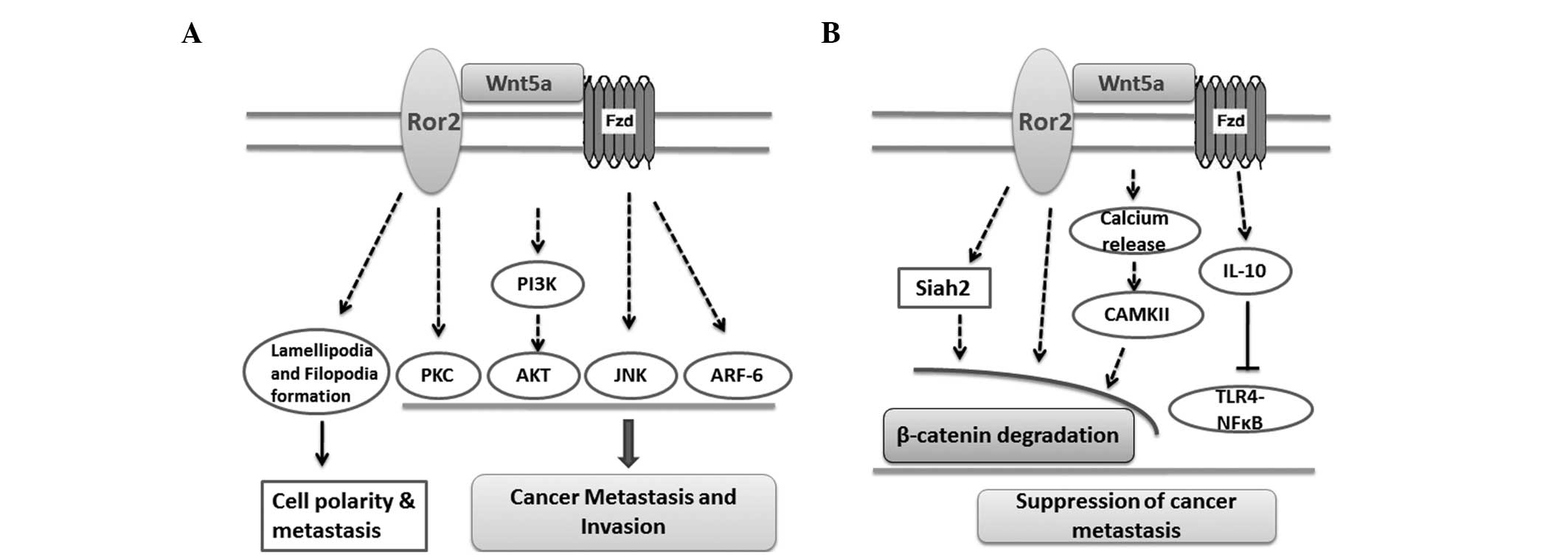 | Figure 1Roles of Wnt5a in cancer metastasis-
(A) promoting and (B) suppressive processes. Fzd, frizzled; PI3K,
phosphatidylinositol 3-kinase; PKC, protein kinase C; AKT, protein
kinase B; JNK, c-Jun N-terminal kinase; ARF-6, ADP-ribosylation
factor 6; IL-10, interleukin-10; CAMKII,
Ca2+/calmodulin-dependent protein kinase II; TLR4,
Toll-like receptor 4; NFκB, nuclear factor κB; ROR2, receptor
tyrosine kinase-like orphan receptor 2. |
4. Metastasis-suppressive activity of Wnt5a
and the underlying mechanisms
In addition to the tumor metastasis promoting roles
of Wnt5a, there is evidence indicating that increased Wnt5a
expression is important for the suppression of metastasis in a
variety of cancer types, including colon, hematopoietic, breast and
thyroid cancers, as well as esophageal squamous cell carcinoma. In
the canonical Wnt signaling pathway, accumulation of β-catenin
activates the transcription of specific genes in tumor progression
(32). Wnt5a is able to antagonize
the canonical Wnt signaling pathway, promote the degradation of
β-catenin and inhibit tumor invasion and metastasis (33). During esophageal squamous cell
carcinoma pathogenesis, Wnt5a is frequently silenced by promoter
methylation and antagonizes the Wnt/β-catenin pathway to exhibit
tumor suppressor properties (34).
Kremenevskaja et al (35)
reported that Wnt5a increased the release of intracellular calcium
ions via the Ca2+/calmodulin-dependent protein kinase II
(CAMKII) signaling pathway to promote β-catenin phosphorylation and
accelerate degradation, thereby inhibiting the translocation and
expression of oncogenes, including c-myc. In colonic epithelial
cancer cells with truncated APC, activation of the calcium-sensing
receptor (CaSR) by ionized Ca2+ ions resulted in the
upregulation of WNT5A transcripts and the increase in Wnt5a protein
secretion. The secreted Wnt5a protein then activated the orphan
tyrosine kinase, ROR2, or an uncharacterized Fzd receptor in an
autocrine manner to increase the mRNA and protein expression levels
of the E-ubiquitin ligase, SIAH2. SIAH2 then mediated growth of
β-catenin degradation and reduced β-catenin signaling. When
full-length APC was overexpressed in these cells, Ca2+
stimulation of the CaSR did not upregulate Wnt5a or increase SIAH2
protein expression (36). These
observations indicated that Wnt5a secretion stimulated by CaSR
activation may inhibit Wnt signaling in an autocrine manner.
Specific studies showed that the inhibitory effect of Wnt5a was not
dependent on GSK-3β (37).
Jöhnsson and Andersson (38)
transfected non-WNT5A expressing MCF-7 breast cancer cells with a
mammalian vector carrying WNT5A cDNA. The production of Wnt5a
protein in MCF-7 cells was accompanied by an increased ability of
discoidin domain receptor 1 (DDR1) phosphorylation and by the
improvement of the cancer cell phenotype. DDR1 not only functions
as an adhesion receptor, but also increases the number of collagen
matrix cells and connections, which can trigger cell signaling and
cytoskeleton rearrangement. The study also demonstrated that Wnt5a
protein participated in the regulation of adhesion and migration
via collagen, and was necessary for the collagen-induced activation
of DDR1 in mammary epithelial cells. Dejmek et al (39) observed loss or downregulation of
Wnt5a expression in 50% of Dukes’ B tumors. Wnt5a negativity was a
strong predictor of an adverse outcome, with a relative mortality
risk of 3.007 (95% confidence interval, 1.336–6.769; P=0.008) after
five years in Wnt5a-negative patients, while the median survival
time following diagnosis was 109.1 months for patients with
Wnt5a-positive primary tumors. The expression of Wnt5a was
decreased in the highly metastatic SW620 human colon cancer cell
line as compared with the nonmetastatic SW480 human colon cancer
cell line. Furthermore, silencing WNT5A in the highly invasive
human colon cancer cell line may contribute to transcriptional
regulation by histone modification (40). Additionally, the expression of
recombinant Wnt5a significantly reduced the migratory capacity of
SW480 cells, the cells that do not express Wnt5a protein possess
the ability of high-invasive and metastasis as well as poorly
differentiated. However, equivalent treatment did not significantly
alter the migration in the Wnt5a-expressing Caco-2 colon cancer
cell line. Since inactivating the APC mutation in SW480 cells
mimics constant activation of canonical Wnt signaling, and the
expression of Wnt5a protein is concurrently absent in SW480 cells,
it was hypothesized that there may be a certain degree of
antagonism between noncanonical Wnt5a expression and canonical Wnt
signaling. These observations indicate that the expression of Wnt5a
in primary Dukes’ B colon cancer tissue constitutes a good
prognostic marker for longer survival. Ying et al (41) also reported the tumor-suppressive
activity of Wnt5a. The promoter region of WNT5A is a typical CpG
island, which is susceptible to deactivation by methylation in
hematological malignancies (34).
By analyzing the epigenetic modification of the
WNT5A gene in colon cells from normal colon tissues, colon cancer
tissues and colorectal cancer (CRC) cell lines, Ying et al
found that WNT5A is silenced in the majority of CRC cell lines due
to promoter methylation (41).
However, Wnt5a is expressed in the majority of normal tissues,
including the colon, and is unmethylated in normal colon epithelial
cells. WNT5A expression was reactivated by pharmacological or
genetic demethylation, indicating that methylation directly
mediates the silencing. Similarly, WNT5A methylation was frequently
detected in CRC tumors (14/29, 48%), but only occasionally in
paired normal colon tissues (2/15, 13%; P=0.025). Ectopic
expression of Wnt5a, but not its nonfunctional short-isoform with
the Wnt domain deleted, in CRC cells resulted in the substantial
inhibition of tumor cell clonogenicity, which is associated with
downregulated intracellular β-catenin protein levels and a
concomitant decrease in β-catenin activity. Consistent with the
results from previous studies, transfection of WNT5A in the FTC-133
thyroid tumor cell line was able to reduce proliferation,
migration, invasiveness and clonogenicity in the cells (35). These effects of Wnt5a
overexpression are mediated through an increase in intracellular
Ca2+ release and β-catenin phosphorylation via CaMKII
pathways, which result in membranous β-catenin translocation and
c-myc oncogene suppression (42).
Therefore, Wnt5a serves as an antagonist to the canonical Wnt
signaling pathway, exhibiting tumor suppressor activity in
differentiated thyroid carcinomas.
Analysis of human primary leukemia cases identified
that a deletion of the WNT5A gene and/or loss of Wnt5a expression
was present in the majority of the patient samples. Wnt5a
hemizygous mice developed myeloid leukemia and B-cell lymphoma,
that are common in clonal origin, and exhibited a loss of Wnt5a
function in tumor tissues. Wnt5a signaling through the noncanonical
Wnt/Ca2+ pathway suppresses cyclin D1 expression and
negatively regulates B cell proliferation in a cell-autonomous
manner (43). Ying et al
(44) revealed hypermethylation in
the promoter of WNT5A in six acute lymphoblastic cell lines (TOM-1,
NALM-20, MY, LOUCY, JURKAT and TANOUE), as well as in 43% (132/307)
of the acute lymphoblastic leukemia cases. Hypermethylation was
found to be associated with the upregulation of cylin D1. As
aforementioned, Wnt5a functions as an effector of tumor metastasis
suppressive actions via various pathways or by epigenetic changes
in its promoter (Fig. 1).
5. Contradictory effects of Wnt5a in breast
cancer metastasis
Contradictory effects of Wnt5a have been
demonstrated in breast cancer. There is increasing evidence that
Wnt5a is a tumor suppressor in breast cancer. Jönsson et al
(45) identified the loss of Wnt5a
expression in 59 out of 96 cases of invasive ductal breast cancer,
which was associated with high-histological grade, high-mitotic
index and negative expression of the estrogen and progesterone
receptors. In addition, 78% of the 32 relapse cases exhibited a
lack of Wnt5a expression, whereas only 35% of the 51 non-recurrent
cases lost Wnt5a expression, and that recurrence-free survival was
obviously shorter in the Wnt5a-negative group. Based on the results
indicating the inhibition metastasis ability of Wnt5a, FOXY-5, a
Wnt5a peptide agonist was developed and tested. Recombinant Wnt5a
and FOXY-5 suppressed cell migration and invasion in vitro.
While in vivo, the metastasis of breast cancer cells that
had been injected into the mice fat pad was also inhibited by
intraperitoneal injection of FOXY-5 into the mice (46,47).
The suppression phenotype induced by Wnt5a is associated with the
induction of interleukin-10 and the inhibition of the Toll-like
receptor 4-nuclear factor κB signaling pathway (48). However, conflicting results remain.
Lejeune et al (49)
reported that the mRNA expression level of WNT5A was upregulated by
ten and four times in benign and invasive breast cancer,
respectively, as compared with normal breast tissues. In addition,
the tumor tissue was Wnt5a negative in 10 cases of the 17 ductal
invasive breast cancer patients. There is increasing evidence
indicating that interactions between tumor cells and the stromal
compartment are critical for malignant progression. Wnt5a appears
to promote malignant progression in breast cancer in a
microenvironment-dependent manner. Overexpression of Wnt5a has been
demonstrated not only in tumor cells, but also in the tumor stroma,
particularly tumor-associated macrophages. Wnt5a is detectable in
macrophages from metastatic lymph nodes, and Wnt5a-positive
macrophages are detected specifically at the invasive front of
lymph node metastases. In vitro experiments that co-cultured
MCF-7 breast cancer cells and macrophages resulted in the
upregulation of Wnt5a in the macrophages. In the context of breast
cancer, Wnt5a induces invasiveness of cancer cells and the
production of matrix metalloproteinase-7 and tumor necrosis
factor-α in macrophages. Thus, Wnt5a is essential for
macrophage-induced invasiveness in breast cancer. The function of
Wnt5a as a suppressor or as a promoter of malignant progression is
determined not only by specific intracellular signaling pathways,
but also by intercellular interactions among various cell types
(50,51).
6. Conclusion
Although the results discussed appear conflicting,
it is conceivable that the roles of Wnt5a in cancer metastasis.
Wnt5a plays a variety of roles in different types of tumors,
depending on the binding receptors, downstream effectors, exogenous
inhibitors, tumor microenvironments and the extracellular matrix
(39), particularly the
cell/tissue-tropic contexts (52).
To further clarify the role of Wnt5a in tumor metastasis, a number
of questions require investigation. It is necessary to reveal the
detailed regulatory mechanisms of Wnt5a on cross-talking between
the canonical and noncanonical pathways. Further investigation of
Wnt5a and the noncanonical Wnt signaling pathways may contribute to
revealing the occurrence, development and metastasis of tumors,
which may provide novel targets for the diagnosis and treatment of
cancer.
Acknowledgements
The study was supported by grants from the National
Natural Science Foundation of China (nos. 81000946 and 81270359)
and the Youth Project of the Educational Department of Hunan
Province (no. 10B090), and supported by the Construct Program of
the Key Discipline in Hunan province.
References
|
1
|
Duffy MJ, McGowan PM and Gallagher WM:
Cancer invasion and metastasis: changing views. J Pathol.
214:283–293. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Larue L and Bellacosa A:
Epithelial-mesenchymal transition in development and cancer: role
of phosphatidylinositol 3′ kinase/AKT pathways. Oncogene.
24:7443–7454. 2005.
|
|
3
|
Valkenburg KC, Steensma MR, Williams BO
and Zhong Z: Skeletal metastasis: treatments, mouse models, and the
Wnt signaling. Chin J Cancer. 32:380–396. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Zerlin M, Julius MA and Kitajewski J:
Wnt/Frizzled signaling in angiogenesis. Angiogenesis. 11:63–69.
2008. View Article : Google Scholar
|
|
5
|
Nusse R, Brown A, Papkoff J, et al: A new
nomenclature for int-1 and related genes: the Wnt gene family.
Cell. 64:2311991. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Yang-Snyder J, Miller JR, Brown JD, Lai CJ
and Moon RT: A frizzled homolog functions in a vertebrate Wnt
signaling pathway. Curr Biol. 6:1302–1306. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Wong GT, Gavin BJ and McMahon AP:
Differential transformation of mammary epithelial cells by Wnt
genes. Mol Cell Biol. 4:6278–6286. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Clark CC, Cohen I, Eichstetter I, et al:
Molecular cloning of the human proto-oncogene Wnt-5A and mapping of
the gene (WNT5A) to chromosome 3p14-p21. Genomics. 18:249–260.
1993. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Danielson KG, Pillarisetti J, Cohen IR, et
al: Characterization of the complete genomic structure of the human
WNT-5A gene, functional analysis of its promoter, chromosomal
mapping, and expression in early human embryogenesis. J Biol Chem.
270:31225–31234. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
DasGupta R, Kaykas A, Moon RT and Perrimon
N: Functional genomic analysis of the Wnt-wingless signaling
pathway. Science. 308:826–833. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Nemeth MJ, Topol L, Anderson SM, Yang Y
and Bodine DM: Wnt5a inhibits canonical Wnt signaling in
hematopoietic stem cells and enhances repopulation. Proc Natl Acad
Sci USA. 104:15436–15441. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Semenov MV, Habas R, Macdonald BT and He
X: SnapShot: Noncanonical Wnt signalingpathways. Cell.
131:13782007. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
McDonald SL and Silver A: The opposing
roles of Wnt-5a in cancer. Br J Cancer. 101:209–214. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Kurayoshi M, Oue N, Yamamoto H, Kishida M,
Inoue A, Asahara T, Yasui W and Kikuchi A: Expression of Wnt-5a is
correlated with aggressiveness of gastric cancer by stimulating
cell migration and invasion. Cancer Res. 66:10439–10448. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Hanaki H, Yamamoto H, Sakane H, et al: An
anti-Wnt5a antibody suppresses metastasis of gastric cancer cells
in vivo by inhibiting receptor-mediated endocytosis. Mol Cancer
Ther. 11:298–307. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Liu J, Zhang Y, Xu R, et al:
PI3K/Akt-dependent phosphorylation of GSK3β and activation of RhoA
regulate Wnt5a-induced gastric cancer cell migration. Cell Signal.
25:447–456. 2013.
|
|
17
|
Bittner M, Meltzer P, Chen Y, et al:
Molecular classification of cutaneous malignant melanoma by gene
expression profiling. Nature. 406:536–540. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Da Forno PD, Pringle JH, Hutchinson P, et
al: WNT5A expression increases during melanoma progression and
correlates with outcome. Clin Cancer Res. 14:5825–5832.
2008.PubMed/NCBI
|
|
19
|
Grossmann AH, Yoo JH, Clancy J, et al: The
small GTPase ARF6 stimulates β-catenin transcriptional activity
during WNT5A-mediated melanoma invasion and metastasis. Sci Signal.
6:ra142013.
|
|
20
|
Michl P, Ramjaun AR, Pardo OE, et al:
CUTL1 is a target of TGF(beta) signaling that enhances cancer cell
motility and invasiveness. Cancer Cell. 7:521–532. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Ripka S, König A, Buchholz M, et al: WNT5A
- target of CUTL1 and potent modulator of tumor cell migration and
invasion in pancreatic cancer. Carcinogenesis. 28:1178–1187. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Schlessinger K, McManus EJ and Hall A:
Cdc42 and noncanonical Wnt signal transduction pathways cooperate
to promote cell polarity. J Cell Biol. 178:355–361. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Kuss P, Kraft K, Stumm J, Ibrahim D,
Vallecillo-Garcia P, Mundlos S and Stricker S: Regulation of cell
polarity in the cartilage growth plate and perichondrium of
metacarpal elements by HOXD13 and WNT5A. Dev Biol. 385:83–93. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Witze ES, Litman ES, Argast GM, Moon RT
and Ahn NG: Wnt5a control of cell polarity and directional movement
by polarized redistribution of adhesion receptors. Science.
320:365–369. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Bowerman B: Cell signaling. Wnt moves
beyond the canon. Science. 320:327–328. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Dissanayake SK, Wade M, Johnson CE, et al:
The Wnt5A/protein kinase C pathway mediates motility in melanoma
cells via the inhibition of metastasis suppressors and initiation
of an epithelial to mesenchymal transition. J Biol Chem.
282:17259–17271. 2007. View Article : Google Scholar
|
|
27
|
Nomachi A, Nishita M, Inaba D, Enomoto M,
Hamasaki M and Minami Y: Receptor tyrosine kinase Ror2 mediates
Wnt5a-induced polarized cell migration by activating c-Jun
N-terminal kinase via actin-binding protein filamin A. J Biol Chem.
283:27973–27981. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Oishi I, Suzuki H, Onishi N, et al: The
receptor tyrosine kinase Ror2 is involved in non-canonical
Wnt5a/JNK signalling pathway. Genes Cells. 8:645–654. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Enomoto M, Hayakawa S, Itsukushima S, et
al: Autonomous regulation of osteosarcoma cell invasiveness by
Wnt5a/Ror2 signaling. Oncogene. 28:3197–3208. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Huang C, Rajfur Z, Borchers C, Schaller MD
and Jacobson K: JNK phosphorylates paxillin and regulates cell
migration. Nature. 424:219–223. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Nomachi A, Nishita M, Inaba D, Enomoto M,
Hamasaki M and Minami Y: Receptor tyrosine kinase Ror2 mediates
Wnt5a-induced polarized cell migration by activating c-Jun
N-terminal kinase via actin-binding protein filamin A. J Biol Chem.
283:27973–27981. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Reya T and Clevers H: Wnt signalling in
stem cells and cancer. Nature. 434:843–850. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Katoh M and Katoh M: STAT3-induced WNT5A
signaling loop in embryonic stem cells, adult normal tissues,
chronic persistent inflammation, rheumatoid arthritis and cancer
(Review). Int J Mol Med. 19:273–278. 2007.
|
|
34
|
Li J, Ying J, Fan Y, et al: WNT5A
antagonizes WNT/β-catenin signaling and is frequently silenced by
promoter CpG methylation in esophageal squamous cell carcinoma.
Cancer Biol Ther. 10:617–624. 2010.
|
|
35
|
Kremenevskaja N, von Wasielewski R, Rao
AS, Schöfl C, Andersson T and Brabant G: Wnt-5a has tumor
suppressor activity in thyroid carcinoma. Oncogene. 24:2144–2154.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
MacLeod RJ, Hayes M and Pacheco I: Wnt5a
secretion stimulated by the extracellular calcium-sensing receptor
inhibits defective Wnt signaling in colon cancer cells. Am J
Physiol Gastrointest Liver Physiol. 293:G403–G411. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Topol L, Jiang X, Choi H, Garrett-Beal L,
Carolan PJ and Yang Y: Wnt-5a inhibits the canonical Wnt pathway by
promoting GSK-3-independent beta-catenin degradation. J Cell Biol.
162:899–908. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Jönsson M and Andersson T: Repression of
Wnt-5a impairs DDR1 phosphorylation and modifies adhesion and
migration of mammary cells. J Cell Sci. 114:2043–2053.
2001.PubMed/NCBI
|
|
39
|
Dejmek J, Dejmek A, Säfholm A, Sjölander A
and Andersson T: Wnt-5a protein expression in primary dukes B colon
cancers identifies a subgroup of patients with good prognosis.
Cancer Res. 65:9142–9146. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Li Q and Chen H: Silencing of Wnt5a during
colon cancer metastasis involves histone modifications.
Epigenetics. 7:551–558. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Ying J, Li H, Yu J, Ng KM, Poon FF, Wong
SC, Chan AT, Sung JJ and Tao Q: WNT5A exhibits tumor-suppressive
activity through antagonizing the Wnt/beta-catenin signaling, and
is frequently methylated in colorectal cancer. Clin Cancer Res.
14:55–61. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Kremenevskaja N, von Wasielewski R, Rao
AS, Schöfl C, Andersson T and Brabant G: Wnt-5a has tumor
suppressor activity in thyroid carcinoma. Oncogene. 24:2144–2154.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Liang H, Chen Q, Coles AH, et al: Wnt5a
inhibits B cell proliferation and functions as a tumor suppressor
in hematopoietic tissue. Cancer Cell. 4:349–360. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Ying J, Li H, Chen YW, Srivastava G, Gao Z
and Tao Q: WNT5A is epigenetically silenced in hematologic
malignancies and inhibits leukemia cell growth as a tumor
suppressor. Blood. 110:4130–4132. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Jönsson M, Dejmek J, Bendahl PO and
Andersson T: Loss of Wnt-5a protein is associated with early
relapse in invasive ductal breast carcinomas. Cancer Res.
62:409–416. 2002.PubMed/NCBI
|
|
46
|
Säfholm A, Tuomela J, Rosenkvist J, Dejmek
J, Härkönen P and Andersson T: The Wnt-5a-derived hexapeptide
Foxy-5 inhibits breast cancer metastasis in vivo by targeting cell
motility. Clin Cancer Res. 14:6556–6563. 2008.PubMed/NCBI
|
|
47
|
Serra R, Easter SL, Jiang W and Baxley SE:
Wnt5a as an effector of TGFβ in mammary development and cancer. J
Mammary Gland Biol Neoplasia. 16:157–167. 2011.PubMed/NCBI
|
|
48
|
Bergenfelz C, Medrek C, Ekström E, et al:
Wnt5a induces a tolerogenic phenotype of macrophages in sepsis and
breast cancer patients. J Immunol. 188:5448–5458. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Lejeune S, Huguet EL, Hamby A, Poulsom R
and Harris AL: Wnt5a cloning, expression, and up-regulation in
human primary breast cancers. Clin Cancer Res. 1:215–222.
1995.PubMed/NCBI
|
|
50
|
Kikuchi A and Yamamoto H: Tumor formation
due to abnormalities in the beta-catenin-independent pathway of Wnt
signaling. Cancer Sci. 99:202–208. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Leek RD and Harris AL: Tumor-associated
macrophages in breast cancer. J Mammary Gland Biol Neoplasia.
7:177–189. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Nishita M, Enomoto M, Yamagata K and
Minami Y: Cell/tissue-tropic functions of Wnt5a signaling in normal
and cancer cells. Trends Cell Biol. 20:346–354. 2010. View Article : Google Scholar : PubMed/NCBI
|














