Introduction
Retinopathy of prematurity (ROP) affects premature
infants and is a major cause of blindness and visual impairement
despite continous improvements in neonatal care (1). There are ~250,000 very low birth weight
(VLBW) infants born in China every year. Among these VLBW, ~30,000
will develop ROP, and ~10,000 receive ophthalmological surgery
(2). As reported in the
International Classification of Retinopathy of Prematurity
(3,4)
and a multi-center clinical trial on retinopathy of prematurity
(ROP) (5–9), current treatments for ROP mainly
include cryotherapy and laser photocoagulation for threshold ROP,
as well as sclera buckling and vitrectomy at a later stage.
Although these procedures do significantly reduce the long-term
adverse outcomes of ROP, such as blindness and retinal detachment,
they may cause permanent damage to the retinal structure and are
associated with life-long eye problems, including visual field
defects and refractive error (10–13).
Researchers have, therefore, increasingly focused on preventing ROP
progression by inhibiting angiogenesis based on the pathogenesis of
ROP (14–23). Two distinct phases have been
identified: Phase I, involving delayed retinal vascular growth
after premature birth; and phase II, concerning uncontrolled
proliferative growth of retinal blood vessels. Such approaches have
been proven feasible in animal experiments; however, angiogenesis
inhibitors may also inhibit normal vascular development and hinder
normal ocular anatomical and functional development (24,25).
These problems remain unresolved. Furthermore, neither surgical
procedures nor angiogenesis inhibitors are capable of preventing
the occurrence of ROP, since they are measures taken after the
development of ocular lesions.
Estrogen (E2), or 17β-estradiol, can act
on vascular cells and neurons and plays an important role in
retinal vascular development. Estrogen receptor (ER)-mediated
E2 is particularly important in normal retinal vascular
development and the pathogenesis of ROP (26–32).
Preterm infants are prone to develop ROP, possibly due to the
unique metabolism of E2 in these infants. Early
E2 replacement administered prior to the occurrence of
abnormal vascular lesions may, therefore, promote the development
of normal blood vessels and thereby improve the prognosis for ROP.
This provides a novel idea for preclinical and clinical trials and
may help validate the role of E2 and ER in normal
retinal vascular development and the pathogenesis of ROP from the
perspective of treatment.
Materials and methods
Experimental animals
A total of 120 healthy 7-day-old C57BL/6J mice of
either gender were selected. The mice were not weaned and were
raised with lactating female rats. These were clean-grade animals
that were provided by the Department of Anatomy, Histology and
Embryology of Fudan University Shanghai Medical College (Shanghai,
China). The study protocol was approved by the university Animal
Care and Use Committee and conformed to international standards for
the humane treatment of experimental animals.
Materials and instruments
The E2 used in the study was
estra-1,3,5(10)-triene-3,17β-diol
(Sigma-Aldrich, St. Louis, MO, USA). The molecular formula was
C18H24O2 and the molecular weight
was 272.4 Da. We customized a 50×40×25-cm glass container with
three round holes in the lid. Of the three holes, one was for
inserting an air inlet pipe connecting the oxygen cylinder and the
nitrogen cylinder, one was an air outlet hole and the remaining
hole was connected to an oxygen analyzer. The bottom of the
container was covered in soda lime to keep the container dry. A
common mouse cage was placed in the container. The room temperature
was maintained at 23±2°C and room luminosity did not exceed 300 Lux
with illumination for 12 h a day.
Experimental methods
Grouping
In the study group, ROP was induced in C57BL/6 mice
by exposing postnatal 7-day-old (P7) mice to 75% oxygen (hyperoxia)
for 5 days, followed by 5 days in normal room air. The mice in the
control group were raised in room air for 10 days. The mice in the
control and hyperoxia groups received an intraperitoneal injection
of either E2 or normal saline (NS) once per day and were
further grouped according to the administered agent, dose and
dosing time, as shown in Table
I.
 | Table I.Experimental grouping for
E2 therapy. |
Table I.
Experimental grouping for
E2 therapy.
| Group | Number of mice | Drug | Dose | Normal air or
oxygen | Time of
injection |
|---|
| 1 | 6 | E2 | 0.5 µg/0.05 ml | Normal air | P7-16 |
| 2 | 18 | E2 | 0.5 µg/0.05 ml | Oxygen
inhalation | P7-11 |
| 3 | 6 | E2 | 0.5 µg/0.05 ml | Oxygen
inhalation | P7-16 |
| 4 | 6 | E2 | 0.5 µg/0.05 ml | Oxygen
inhalation | P12-16 |
| 5 | 6 | E2 | 1.0 µg/0.05 ml | Normal air | P7-16 |
| 6 | 18 | E2 | 1.0 µg/0.05 ml | Oxygen
inhalation | P7-11 |
| 7 | 6 | E2 | 1.0 µg/0.05 ml | Oxygen
inhalation | P7-16 |
| 8 | 6 | E2 | 1.0 µg/0.05 ml | Oxygen
inhalation | P12-16 |
| 9 | 6 | E2 | 1.5 µg/0.05 ml | Normal air | P7-16 |
| 10 | 18 | E2 | 1.5 µg/0.05 ml | Oxygen
inhalation | P7-11 |
| 11 | 6 | E2 | 1.5 µg/0.05 ml | Oxygen
inhalation | P7-16 |
| 12 | 6 | E2 | 1.5 µg/0.05 ml | Oxygen
inhalation | P12-16 |
| 13 | 6 | NS | 0.05 ml | Normal air | P7-16 |
| 14 | 6 | NS | 0.05 ml | Oxygen
inhalation | P7-16 |
Flat mounting
The flat-mounted retinas were prepared as follows.
For the three groups with an injection time of P7-11, retinas were
separated and flat-mounted on the slides on days 0, 2 and 5 (i.e.,
P12, 14 and 17), respectively, after the mice were removed from the
oxygen container. Six eyes were enucleated at each time-point from
6 mice. For the remaining groups, 6 eyes were enucleated from 6
mice, and the flat-mounted retinas were prepared on P17 to observe
the profile of retinal vascular development and proliferation. The
specific method of retina flat-mounting and ADPase enzyme
histochemistry is as detailed below. For anesthesia, the mice
received a peritoneal injection of 2,2,2-tribromoethanol (Avertin®;
0.5 ml/15 g; Sigma-Aldrich). For perfusion, the chest cavity of
each mouse was opened and the heart was exposed. Through an
infusion needle inserted in the left ventricle, NS was infused for
2–3 min followed by infusion of 4% paraformaldehyde solution for ~5
min. The eyeballs of the mice were enucleated and fixed in 4%
paraformaldehyde solution. For sampling, the limbus was cut open
circularly at 1 mm posterior to the limbus. The cornea was removed
and the lens was extracted. Four to five radial incisions were made
using the optic papilla as the center. The sclera and choroid were
removed, and the vitreous body and the retinal pigment epithelial
layer were removed by a brush. The samples were then rinsed with
0.05 mol Tris-maleate buffer (pH 7.2; Sigma-Aldrich) five times (15
min each time). Next, the samples were incubated in the reaction
solution at 37°C for 15 min. For color development, the samples
were reacted with 10% (1:10) ammonium sulfide for 5 min. The
results were observed with an optical microscope.
Preparation and observation of paraffin
sections
Six eyes of 6 mice were collected from each group.
The mice were sacrificed by cervical dislocation 5 days after the
removal of the mice from the oxygen container. The eyeballs were
enucleated and directions were marked. For fixation, the enucleated
eyeball was placed in 4% paraformaldehyde solution and fixed for 24
h. Samples were dehydrated with gradient alcohol and made
transparent with xylene. The samples were then embedded in soft
paraffin, hard paraffin I and hard paraffin II for 30 min, 1 h and
1 h, respectively. Serial sections of 6 µm were sliced parallel to
the sagittal plane from the cornea to the optic papilla. Slices
were then flattened in warm water and mounted on their slide. The
samples were dewaxed with xylene and dehydrated with gradient
alcohol. Conventional hematoxylin and eosin staining was then
performed. Finally, an optical microscope was used to observe the
results.
Endothelial cell nucleus count
The endothelial cell nucleus count in the new
retinal blood vessels was performed as follows. Ten pathological
slices were taken intermittently from each eyeball. The interval
between two adjacent sections was 60 µm (10 slices). The
endothelial cell nuclei of the blood vessels that broke through the
internal limiting membrane of the retina were counted by section
and by eyeball. Only the cell nuclei of blood vessels near the
internal limiting membrane, rather than those of blood vessels in
the vitreous cavity not associated with the internal limiting
membrane, were counted.
Statistical analysis
All data were analyzed with the statistical analysis
software SPSS 11.0 for Windows (SPSS, Inc., Chicago, IL, USA).
Analysis of variance was used to compare the number of nuclei in
the endothelial cells of new blood vessels that broke through the
internal limiting membrane among the groups. P<0.05 was
considered to indicate a statistically significant difference.
Results
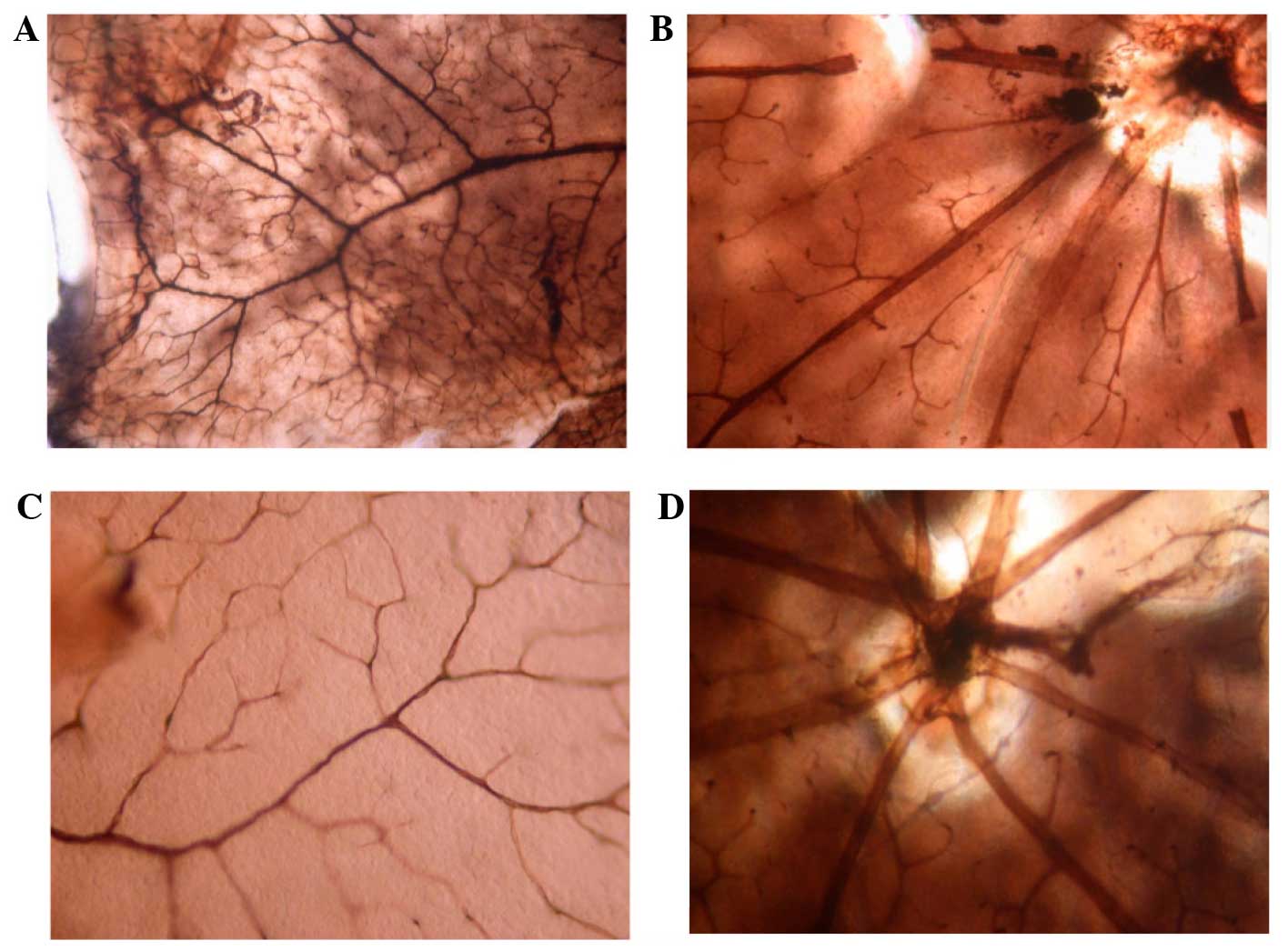
Vascular morphological changes
following E2 treatment
No retinal vascular abnormalities were observed at
P17 in the normal mice that received various doses of E2
or NS between P7 and 16. All blood vessels were mature, and the
larger vessels had more branches with consistently large lumen. The
surrounding small vessels were well developed, forming a vascular
arch at the edge of the retina. The branching structure of the
superficial vascular network and the polygonal mesh-like structure
of the deep vascular network could be clearly noted, and no blood
vessel obstruction was observed (Fig.
1).
Mice receiving oxygen supplement consistently showed
retinal neovascularization following NS injection or injection of
E2 at varying doses during different periods. On P12,
non-perfusion areas at the center of the retina, poorly-branched
large vessels with a narrow lumen and naïve surrounding small
vessels were observed. On P14, neovascularization was found at the
interface between the vascularized zone and the non-vascularized
zone. On P17, the number of new vessels had increased, and the
two-tier vascular network had lost its normal structure, with
accompanying vascular obstruction. The number of blood vessels or
the size of the central non-perfusion area could not be compared
morphologically among the different groups.
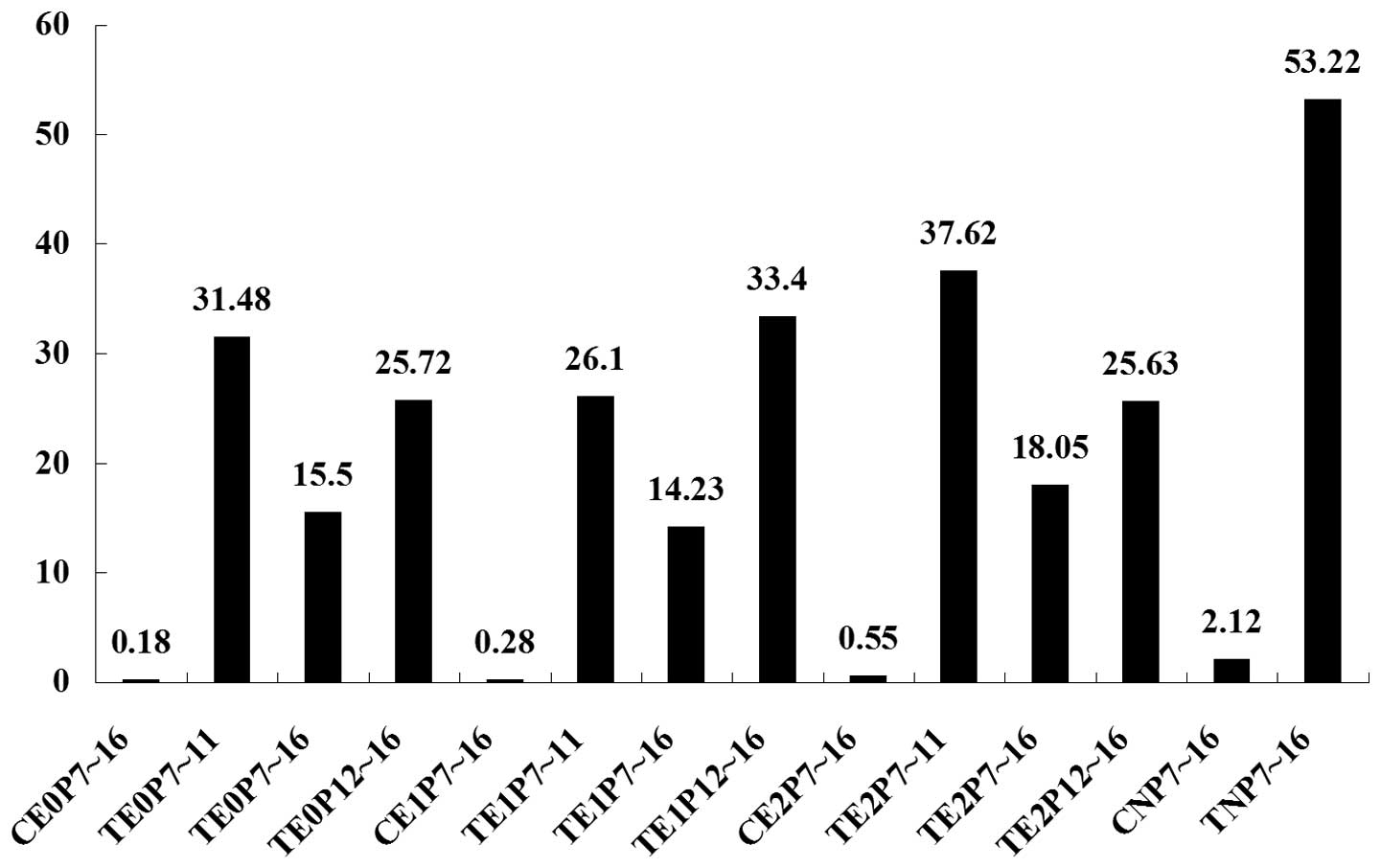
Quantification of angiogenesis
following treatment
The effect of E2 treatment on
angiogenesis was quantified by counting the endothelial cell nuclei
of the blood vessels that broke through the internal limiting
membrane of the retina. It was found that, following both
E2 and NS injection, the number of nuclei in the normal
mice was low, and pair-wise comparison revealed no significant
difference among the four groups (P>0.05). By contrast, a highly
significant difference was found in the number of nuclei between
the normal mice and the mice receiving oxygen inhalation
(P<0.0001). The number of nuclei decreased in the mice receiving
oxygen inhalation following injection of E2 at all doses
and time-points. Following NS injection, the number of nuclei in
the mice receiving oxygen inhalation was significantly higher than
that in the remaining groups (P<0.0001; Table II and Fig. 2).
| Table II.Pair-wise comparison of the results
of the mean (P-value). |
Table II.
Pair-wise comparison of the results
of the mean (P-value).
|
| Group 1 | Group 2 | Group 3 | Group 4 | Group 5 | Group 6 | Group 7 | Group 8 | Group 9 | Group 10 | Group 11 | Group 12 | Group 13 |
|---|
| Group 2 | <0.001 | – | – | – | – | – | – | – | – | – | – | – | – |
| Group 3 | <0.001 | <0.001 | – | – | – | – | – | – | – | – | – | – | – |
| Group 4 | <0.001 |
0.052 |
0.001 | – | – | – | – | – | – | – | – | – | – |
| Group 5 |
0.973 | <0.001 | <0.001 | <0.001 | – | – | – | – | – | – | – | – | – |
| Group 6 | <0.001 |
0.071 | <0.001 |
0.897 | <0.001 | – | – | – | – | – | – | – | – |
| Group 7 | <0.001 | <0.001 |
0.669 | <0.001 | <0.001 | <0.001 | – | – | – | – | – | – | – |
| Group 8 | <0.001 |
0.517 | <0.001 |
0.010 | <0.001 |
0.014 | <0.001 | – | – | – | – | – | – |
| Group 9 |
0.901 | <0.001 | <0.001 | <0.001 |
0.928 | <0.001 | <0.001 | <0.001 | – | – | – | – | – |
| Group 10 | <0.001 |
0.039 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 |
0.155 | <0.001 | – | – | – | – |
| Group 11 | <0.001 | <0.001 |
0.389 |
0.010 | <0.001 |
0.007 |
0.198 | <0.001 | <0.001 | <0.001 | – | – | – |
| Group 12 | <0.001 |
0.048 |
0.001 |
0.978 | <0.001 |
0.875 | <0.001 |
0.009 | <0.001 | <0.001 |
0.011 | – | – |
| Group 13 |
0.514 | <0.001 | <0.001 | <0.001 |
0.536 | <0.001 | <0.001 | <0.001 |
0.597 | <0.001 | <0.001 | <0.001 | – |
| Group 14 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 |
The number of nuclei differed significantly among
the different dose groups due to the difference in the time-period
of drug administration. The number of nuclei was significantly
reduced in the three groups in which the drug was administered
during P7-16, when compared with the number of nuclei in the groups
in which the drug was administered during the two other
time-periods (P<0.05). The effect of different doses
administered during the same time-period was also compared by
counting the number of endothelial cell nuclei in the newly formed
blood vessels. For P7-11, the doses could be ranked as follows: 1.0
µg <0.5 µg <1.5 µg. No significant difference in the number
of endothelial cell nuclei was found between the 1.0- and 0.5-µg
groups, but the results for both these groups were significantly
different from the result for the 1.5-µg group (1.0 µg vs. 1.5 µg,
P=0.039; 0.5 µg vs. 1.5 µg, P<0.0001). For P7-16, the order was
1.0 g <0.5 g <1.5 g and no significant difference could be
found among the three groups. For P12-16, the order was 1.5 µg
<0.5 µg <1.0 µg; no significant difference in the number of
endothelial cell nuclei was found between the 1.5- and 0.5-µg
groups, but both were significantly different from the 1.0-µg group
(1.5 µg vs. 1.0 µg, P=0.009; 0.5 µg vs. 1.0 µg, P=0.01).
Discussion
In addition to its effect on the reproductive
organs, E2 acts on the vascular cells and neurons and
plays varying roles under different physiological and pathological
conditions (26,27). E2 may protect the
cardiovascular system against cardiovascular diseases, such as
hypertension and atherosclerosis, by dilating blood vessels and
increasing blood flow (28).
E2 may increase the activity of endothelial cells,
inhibit endothelial cell apoptosis, promote endothelial cell
proliferation, facilitate angiogenesis and the growth of blood
vessels and play an important role in retinal vascular development
(29,30,33).
In vivo, E2 shows its specific effects only
subsequent to binding to ER (31,32).
Basic endothelial cell-derived nitric oxide may dilate blood
vessels and prevent thrombosis by inhibiting platelet aggregation,
a process that may be regulated by ER (34). In a previous study, we found that
ER-mediated E2 plays an important role in normal retinal
vascular development and ROP pathogenesis (35). Maternal E2 levels during
pregnancy may increase by ~100-fold (33,35–39).
Fetal E2 levels increase with gestational age, with the
E2 concentration peaking between 33 and 36 weeks of
pregnancy. E2 levels drop rapidly after birth and may
reach normal neonatal levels at 24 h; therefore, since preterm
infants are delivered early, their E2 levels are lower
than those of full-term infants. E2 levels drop more
rapidly and fluctuate more in preterm infants than those in
full-term neonates, which may explain why such infants are prone to
ROP (36). Theoretically, early
supplementation of exogenous E2 would facilitate
vascular development and thereby prevent severe retinal hypoxia and
subsequent cascade reactions.
In the present study it was found that the number of
endothelial cell nuclei in the newly formed blood vessels was
reduced significantly in all the three groups receiving
E2 treatment compared with those receiving NS
(P<0.0001), suggesting that E2 replacement therapy
did have beneficial effects; however, E2 did not inhibit
angiogenesis completely, and it was difficult to compare
angiogenesis in the flat-mounted retinas among the different groups
based on morphology alone. In the tissue sections in which nuclei
could be counted, the nucleus count was significantly different
from that of the normal mice (P<0.0001), indicating that, to
some extent, E2 plays a role in the development and
progression of ROP. E2 may be involved in this process
by promoting the expression of the VEGF gene (39), resulting in the formation of a large
number of new blood vessels. Miyamoto et al (30) found that retinal angiogenesis was
reduced in ROP mice following intraperitoneal injection of
E2 during P7–11. They hypothesized that E2
supplementation at the hyperoxia stage would partially restore the
expression of the VEGF gene, promoting the growth of retinal
blood vessels and thereby alleviating hyperoxia-induced retinal
injury. Miyamoto et al observed that in the subsequent
hypoxia phase, E2 supplementation at the same dose on
P12-16 also reduced angiogenesis. Furthermore, when the drug was
administered during P7-16, the number of endothelial cell nuclei in
the new blood vessels was the lowest, suggesting that E2
may inhibit angiogenesis via other mechanisms. Transient blocking
of the middle cerebral artery can cause retinal ischemia in adult
female Sprague Dawley rats, and terminal deoxynucleotidyl
transferase-mediated dUTP nick end labeling staining showed a
significantly increased number of apoptotic cells in the ganglion
cell layer at 1 h after ischemia (40). E2 treatment can completely
reverse such apoptosis (40),
suggesting that E2 has neuroprotective effects when the
retina is in a low oxygenation state. This may explain the reduced
angiogenesis following E2 administration during P12-16
and P7-16.
In the present study, the experimental animals were
divided into 14 groups according to dosage and dosing period. The
dosing periods were the same as those in the study by Miyamoto
et al (30), as dosing in
those periods can exactly reflect the effect of dosing
simultaneously or separately in the two phases of ROP development
and progression, determine the treatment window and provide a
preclinical basis for potential clinical application in the future.
Furthermore, elucidating the roles of E2 and its
receptor ER at different stages of ROP pathogenesis may reveal the
optimal mode of treatment. Three dosage groups, 0.5, 1.0 and 1.5
µg, were selected. The doses of 1.0 and 1.5 µg represent the upper
and lower limits of the dosage for E2 replacement
therapy in menopausal women (0.25–0.5 mg/day), respectively, and
were determined by dose conversion between different species
(41). While the two higher doses
are within the physiological range, the lower dose of 0.5 µg is
below the physiological range. It was found that, for each dose,
maximal inhibitory effects were achieved when the drug was
administered during P7-16, which was consistent with the results of
Miyamoto et al (30). The
results support a potential therapeutic benefit of E2 at
the two stages of ROP pathogenesis. The incidence of blindness in
diabetic men aged <45 years is higher than that in diabetic
women of the same age, and the incidence is similar in men and
women aged >45 years, suggesting that E2 may improve
the prognosis of diabetic retinopathy (42). Diabetic retinopathy and ROP have a
similar pathogenesis, which may further support our view that
E2 plays an important role in retinal development and
ROP pathogenesis in premature infants and holds promise as a
potential treatment for ROP.
The present study additionally compared the effects
of different doses of E2 administered at different
time-periods. It was found that the number of nuclei in the
high-dose group (1.5 µg) was greatest when the drug was
administered at P7-11; at this time-period, the difference from the
other two dose groups was highly significant. The results did not
demonstrate the superiority of a high dose. The number of nuclei
did not differ significantly between the 1.0- and 0.5-µg groups
when the drug was administered during P7-16, although the number in
the 1.0-µg hypoxia group was slightly lower. It is therefore
suggested that E2 replacement therapy may achieve the
maximum effect even at a low dose (1.0 µg). E2 has a
long history of clinical application. It has been used widely in
menopausal and postmenopausal women, since it may improve
menopausal symptoms and osteoporosis and provide skin care effects;
however, misuse and abuse of E2, particularly overdose
of E2, may increase the incidence of uterine and breast
cancer (28,43). E2 should therefore be
appropriately used under the supervision of healthcare
professionals and with the close monitoring of blood levels.
A number of clinical trials have evaluated
E2 replacement therapy in premature infants (29,31–33).
Extremely low birth weight neonates with an average gestational age
of 26.6 weeks and a birth weight of 675 g received E2
supplementation at an intravenous dose of 2.3 mg/kg/day. If the
intravenous infusion could not be maintained, transepidermal
administration was used instead. The total duration of treatment
was 6 weeks to maintain the plasma E2 at the
intrauterine level (33). The
neonates were followed-up until 15 months of corrected gestational
age (34). Following drug
administration, the uterine volume, breast, weight, length and head
circumference of the neonates showed a transient increase and the
bone mineral deposition rate trended towards an increase (33,35). The
incidence of chronic lung diseases decreased and complications of
the central nervous system were reduced (33,35).
Although the administered dose was very high and no long-term
follow-up or any multi-center clinical trial is yet available, the
results remain encouraging.
The present study demonstrated that E2 is
important in the normal development of retinal blood vessels and
the pathogenesis of ROP and has evident efficacy in the prevention
and treatment of ROP. Although E2 may not completely
prevent the occurrence of ROP, it may at least alleviate ROP and
thereby reduce the incidence of long-term complications, such as
blindness and refractive errors. This effect of E2 is of
great significance to individuals and society and may provide
significant economic benefits. In addition, due to the special
metabolism of E2 in preterm infants, E2
supplementation not only facilitates retinal development but also
improves the development of several other systems, which may result
in a significant improvement in the prognosis of preterm infants
and their future quality of life. Despite this, a number of steps
are still required to translate these results from animal studies
into eventual clinical application, including rigorous clinical
trials and long-term follow-up studies, in order to determine the
pharmacokinetics, dose, dosing regimen and schedule and assess the
efficacy, safety and side effects.
In conclusion, E2 plays an important role
in retinal vascular development and the pathogenesis of ROP in
mice. E2 replacement therapy may alleviate retinal
disease in neonatal mice, and intraperitoneal injection of
E2 at a dose of 1.0 µg during P7-16 achieved the best
efficacy in mice; however, the long-term efficacy and side effects
of E2 require further study.
Acknowledgements
This study was sponsored by the Foundation for
Doctoral Degrees of Fudan University.
References
|
1
|
Chen J and Smith LE: Retinopathy of
prematurity. Angiogenesis. 10:133–140. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Li Z and Ni WJ: Advances of biological
drugs for retinal neovascularization. Guo Ki Yan Ke Za Zhi.
7:1119–1123. 2007.(In Chinese).
|
|
3
|
No authors listed. An international
classification of retinopathy of prematurity. Pediatrics.
74:127–133. 1984.PubMed/NCBI
|
|
4
|
No authors listed. An international
classification of retinopathy of prematurity. II. The
classification of retinal detachment. The International Committee
for the Classification of the Late Stages of Retinopathy of
Prematurity. Arch Ophthalmol. 105:906–912. 1987. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Cryotherapy for Retinopathy of Prematurity
Cooperative Group, . Multicenter trial of cryotherapy for
retinopathy of prematurity. Preliminary results. Arch Ophthalmol.
106:471–479. 1988. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Cryotherapy for Retinopathy of Prematurity
Cooperative Group, . Multicenter trial of cryotherapy for
retinopathy of prematurity. Three-month outcome. Arch Ophthalmol.
108:195–204. 1990. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Cryotherapy for Retinopathy of Prematurity
Cooperative Group, . Multicenter trial of cryotherapy for
retinopathy of prematurity. One-year outcome - structure and
function. Arch Ophthalmol. 108:1408–1416. 1990. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Cryotherapy for Retinopathy of Prematurity
Cooperative Group, . Multicenter trial of cryotherapy for
retinopathy of prematurity. 3 1/2-year outcome - structure and
function. Arch Ophthalmol. 111:339–344. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Cryotherapy for Retinopathy of Prematurity
Cooperative Group, . Multicenter trial of cryotherapy for
retinopathy of prematurity. Snellen visual acuity and structural
outcome at 5 1/2 years after randomization. Arch Ophthalmol.
114:417–424. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Greven CM and Tasman W: Rhegmatogenous
retinal detachment following cryotherapy in retinopathy of
prematurity. Arch Ophthalmol. 107:1017–1018. 1989. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Ben-Sira I, Nissenkorn I, Weinberger D,
Shohat M, Kremer I, Krikler R and Reisner SH: Long-term results of
cryotherapy for active stages of retinopathy of prematurity.
Ophthalmology. 93:1423–1428. 1986. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Quinn GE, Miller DL, Evans JA, Tasman WE,
McNamara JA and Schaffer DB: Measurement of Goldmann visual fields
in older children who received cryotherapy as infants for threshold
retinopathy of prematurity. Arch Ophthalmol. 114:425–428. 1996.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Biglan AW, et al: Progress in ROP
research. Chin J Med. 1:7–13. 1990.
|
|
14
|
Ma DH, Chen JK, Zhang F, Lin KY, Yao JY
and Yu JS: Regulation of corneal angiogenesis in limbal stem cell
deficiency. Prog Retin Eye Res. 25:563–590. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Mintz-Hittner HA, Kennedy KA and Chuang
AZBEAT-ROP Cooperative Group: Efficacy of intravitreal bevacizumab
for stage 3+ retinopathy of prematurity. N Engl J Med. 364:603–615.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Miki K, Miki A, Matsuoka M, Muramatsu D,
Hackett SF and Campochiaro PA: Effects of intraocular ranibizumab
and bevacizumab in transgenic mice expressing human vascular
endothelial growth factor. Ophthalmology. 116:1748–1754. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Yatoh S, Kawakami Y, Imai M, Kozawa T,
Segawa T, Suzuki H, Yamashita K and Okuda Y: Effect of a topically
applied neutralizing antibody against vascular endothelial growth
factor on corneal allograft rejection of rat. Transplantation.
66:1519–1524. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Moshfeghi DM and Berrocal AM: Retinopathy
of prematurity in the time of bevacizumab: Incorporating the
BEAT-ROP results into clinical practice. Ophthalmology.
118:1227–1228. 2011.PubMed/NCBI
|
|
19
|
O'Reilly MS, Boehm T, Shing Y, Fukai N,
Vasios G, Lane WS, Flynn E, Birkhead JR, Olsen BR and Folkman J:
Endostatin: An endogenous inhibitor of angiogenesis and tumor
growth. Cell. 88:277–285. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Suzuma K, Takagi H, Otani A, Oh H and
Honda Y: Expression of thrombospondin-1 in ischemia-induced retinal
neovascularization. Am J Pathol. 154:343–354. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Coughlin SR: Thrombin signaling and
protease-activated receptors. Nature. 407:258–264. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Shafiee A, Penn JS, Krutzsch HC, Inman JK,
Roberts DD and Blake DA: Inhibition of retinal angiogenesis by
peptides derived from thrombospondin-1. Invest Ophthalmol Vis Sci.
41:2378–2388. 2000.PubMed/NCBI
|
|
23
|
Lonchampt M, Pennel L and Duhault J:
Hyperoxia/normoxia-driven retinal angiogenesis in mice: A role for
angiotensin II. Invest Ophthalmol Vis Sci. 42:429–432.
2001.PubMed/NCBI
|
|
24
|
Falavarjani KG and Nguyen QD: Adverse
events and complications associated with intravitreal injection of
anti-VEGF agents: A review of literature. Eye(Lond). 27:787–794.
2013.PubMed/NCBI
|
|
25
|
Hoerster R, Muether P, Dahlke C, Mehler K,
Oberthür A, Kirchof B and Fauser S: Serum concentrations of
vascular endothelial growth factor in an infant treated with
ranibizumab for retinopathy of prematurity. Acta Ophthalmol.
91:e74–e75. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Simoncini T, Maffei S, Basta G, Barsacchi
G, Genazzani AR, Liao JK and De Caterina R: Estrogens and
glucocorticoids inhibit endothelial vascular cell adhesion
molecule-1 expression by different transcriptional mechanisms. Circ
Res. 87:19–25. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Ma X, Bi H, Qu Y, Xie X and Li J: The
contrasting effect of estrogen on mRNA expression of VEGF in bovine
retinal vascular endothelial cells under different oxygen
conditions. Graefes Arch Clin Exp Ophthalmol. 249:871–877. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Thompson LP, Pinkas G and Weiner CP:
Chronic 17beta-estradiol replacement increases nitric
oxide-mediated vasodilation of guinea pig coronary
microcirculation. Circulation. 102:445–451. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Suzuma I, Mandai M, Takagi H, Suzuma K,
Otani A, Oh H, Kobayashi K and Honda Y: 17 Beta-estradiol increases
VEGF receptor-2 and promotes DNA synthesis in retinal microvascular
endothelial cells. Invest Ophthalmol Vis Sci. 40:2122–2129.
1999.PubMed/NCBI
|
|
30
|
Miyamoto N, Mandai M, Takagi H, Suzuma I,
Suzuma K, Koyama S, Otani A, Oh H and Honda Y: Contrasting effect
of estrogen on VEGF induction under different oxygen status and its
role in murine ROP. Invest Ophthalmol Vis Sci. 43:2007–2014.
2002.PubMed/NCBI
|
|
31
|
Takeyama J, Suzuke T, Inoue S, Kaneko C,
Nagura H, Harada N and Sasano H: Expression and cellular
localization of estrogen receptors and in the human fetus. J Clin
Endocrinol Metab. 86:2258–2262. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Gruber CJ, Tschugguel W, Schneeberger C
and Huber JC: Production and actions of estrogens. N Engl J Med.
346:340–352. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Huppmann S, Römer S, Altmann R, Obladen M
and Berns M: 17beta-estradiol attenuates hyperoxia-induced
apoptosis in mouse C8-D1A cell line. J Neuro Res. 86:3420–3426.
2008. View Article : Google Scholar
|
|
34
|
Morles DE, McGowan KA, Grant DS,
Maheshwari S, Bhartiya D, Cid MC, Kleinman HK and Schnaper HW:
Molecular and cellular cardiology: Estrogen promotes angiogenic
activity in human umbilical vein endothelial cells in vitro and in
a murine model. Circulation. 91:755–763. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Shi WJ, Chen C, Zhou GM, Hu BY and Xiao
HL: The role of vascular endothelial growth factor, fibroblast
growth factor-2 and estrogen receptor in the pathogenesis of
retinopathy of neonatal mice. Fudan Xue Bao Yi Xue Ban Bian Ji Wei
Yuan Hui. 34:228–232. 2007.(In Chinese).
|
|
36
|
Trotter A, Maier L, Grill HJ, Kohn T,
Heckmann M and Pohlandt F: Effects of postnatal estradiol and
progesterone replacement in extremely preterm infants. J Clin
Endocrinol Metab. 84:4531–4535. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Trotter A, Bokelmann B, Sorgo W,
Bechinger-Kornhuber D, Heinemann H, Schmücker G, Oesterle M,
Köhntop B, Brisch KH and Pohlandt F: Follow-up examination at the
age of 15 months of extremely preterm infants after postnatal
estradiol and progesterone replacement. J Clin Endocrinol Metab.
86:601–603. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Trotter A, Maier L, Kohn T, Böhm W and
Pohlandt F: Growth of the uterus and mammary glands and vaginal
cytologic features in extremely premature infants with postnatal
replacement of estradiol and progesterone. Am J Obstet Gynecol.
186:184–188. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
McLaren J, Prentice A, Charnock-Jones DS,
Millican SA, Müller KH, Sharkey AM and Smith SK: Vascular
endothelial growth factor is produced by peritoneal fluid
macrophages in endometriosis and is regulated by ovarian steroids.
J Clin Invest. 98:482–489. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Kaja S, Yang SH, Wei J, Fujitani K, Liu R,
Brun-Zinkernagel AM, Simpkins JW, Inokuchi K and Koulen P: Estrogen
protects the inner retina from apoptosis and ischemia-induced loss
of Vesl-1 L/Homer 1c immunoreactive synaptic connections. Invest
Ophthalmol Vis Sci. 44:3155–3162. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Xu J and Su H: Body surface area
calculation method for humans and animalsPharmacological
Experiments. Yuanpei Zhang: Beijing Publication Group; Beijing: pp.
2411996
|
|
42
|
Yuen KK and Kahn HA: The association of
femal hormones with blindness from diabetic retinopathy. Am J
Ophthalmol. 81:820–822. 1976. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Porter JC: Hormonal regulation of breast
development and activity. J Invest Dermatol. 63:85–92. 1974.
View Article : Google Scholar : PubMed/NCBI
|