Introduction
In the field of breast cancer research, focus has
gradually changed from the clinical parameters and pathological
index levels to the biology and gene expression levels of
molecules, including certain cell signal transduction pathways that
regulate cell cycle development (1,2). The Wnt
signaling transduction pathway is closely associated with the
pathogenesis of breast cancer. β-catenin is a multifunctional
protein and a key carcinogenic regulator of the Wnt signaling
transduction pathway. β-catenin may accumulate aberrantly in the
cytoplasm due to its degradation barrier, and may then be
transferred into the nucleus when it reaches a certain
concentration. Through the interaction with T-cell factor/lymphoid
enhancer factor (TCF/LEF) and coactivators, β-catenin is able to
activate a number of downstream target genes, such as cyclin D1,
which results in uncontrolled cell proliferation and
differentiation, and ultimately tumorigenesis (3). Traditionally, the breast cancer
estrogen receptor (ER) refers to the ER-α. A previous study has
indicated that the positive expression rate of cyclin D1 was
significantly increased in breast cancer tissue with positive
expression of ER-α compared with that with negative expression of
ER-α (4). ER-mediated signaling may
activate β-catenin, and the Wnt signaling transduction pathway is
subsequently activated and the expression of cyclin D1 increased in
breast cancer tissue (5). These
results suggest that the ER-mediated signaling pathway may be
associated with the Wnt signaling pathway. However, in 1996, Kuiper
et al identified an additional breast cancer cell estrogen
receptor subtype, namely ER-β (6).
Long-term exposure to estrogen may lead to breast tumorigenesis.
Previous studies have indicated that ER-β is differentially
expressed in breast cancer tissues (7,8), which
may be associated with tumorigenesis and the development of breast
cancer.
In the present study, the expression levels of
β-catenin, cyclin D1 and ER-β were detected using
immunohistochemistry. Subsequently, the associations among the
expression levels of β-catenin, cyclin D1 and ER-β were assessed.
In addition, the effects of Wnt signaling transduction pathway and
the positive expression of ER-β on the survival times of breast
cancer patients were further analyzed.
Materials and methods
Patient data
Paraffin-embedded breast cancer tissues were
collected from 226 patients with pathologically confirmed breast
cancer. All the patients were diagnosed and treated in the First
Affiliated Hospital of Xinjiang Medical University (Ürümqi, China)
between January 2000 and December 2010. The patients exhibited
infiltrating ductal carcinoma, with the clinical stages varying
between stage 0 and stage II. The clinical characteristics of the
patients are presented in Table I,
and the follow-up period for the patients ranged between 2 and 12
years, during which 15 patients were censored. Patients were
censored if they succumbed to other causes, or were lost to
follow-up at the time of last contact or prior to the study
cut-off.
 | Table I.Clinical data of the patients
(n=226). |
Table I.
Clinical data of the patients
(n=226).
| Clinical feature | Cases, n (%) |
|---|
| Age (years) |
|
| ≤50 | 163 (72.1) |
|
>50 | 63 (27.9) |
| Menstruation |
|
|
Menopause | 114 (50.4) |
|
Non-menopause | 112 (49.6) |
| Tumor diameter
(cm) |
|
| ≤2 | 141 (62.4) |
|
>2–3 | 85 (37.6) |
| Histological
grade |
|
| Grade
I | 56 (24.8) |
| Grade
II | 120 (53.1) |
| Grade
III | 50 (22.1) |
| Lymph node
metastasis |
|
Negative | 169 (74.8) |
|
Positive | 57 (25.2) |
| Tumor stage |
| Stage
0-I | 117 (51.8) |
| Stage
II | 109 (48.2) |
Prior written and informed consent was obtained from
each patient and the study was approved by the Ethics Review Board
of Xinjiang Medical University.
Reagents
An anti-human β-catenin monoclonal antibody (1:50;
ZM0422) and an anti-human cyclin D1 monoclonal antibody (1:50;
ZA0101) were purchased from Beijing Zhongshan Golden Bridge
Biotechnology Co., Ltd. (Beijing, China). An anti-human ER-β
polyclonal antibody (1:60; BY-02101) was obtained from Shanghai
Yueyan Biological Technology, Co., Ltd. (Shanghai, China), while
horseradish peroxidase (HRP)-conjugated anti-rabbit and anti-mouse
IgG secondary antibodies were purchased from Santa Cruz
Biotechnology, Inc. (sc-47047; Santa Cruz, CA, USA). A streptavidin
peroxidase (SP) immunohistochemical hypersensitivity kit and
3,3′-diaminobenzidine (DAB) reagents were obtained from Fuzhou
Maixin Biotechnology Co., Ltd. (Fuzhou, China).
Immunohistochemical staining
Immunohistochemical staining was conducted according
to the instructions provided with the SP immunohistochemical
hypersensitivity kit, with minor modifications. Briefly, tissues
were fixed in formaldehyde and embedded in paraffin. After dewaxing
and rehydration, sections were incubated with 0.3% hydrogen
peroxide to inactivate endogenous peroxidase activity. Antigen
retrieval was achieved by incubating with sodium citrate (pH 6.0).
After blocking, the sections were incubated with the
anti-β-catenin, anti-cyclin D1 and anti-ER-β primary antibodies at
37°C in the dark for 1 h. Following washing with phosphate-buffered
saline (PBS), the HRP-conjugated IgG secondary antibodies were
added and incubated in the dark for 30 min. Subsequently, the
sections were developed with DAB chromogenic reagent and
counterstained with hematoxylin. Breast cancer tissues with known
positive expression of β-catenin, cyclin D1 and ER-β were used as
positive controls. For a negative control, the primary antibody was
replaced with PBS.
Determination criteria of
expression
Positive expression of β-catenin was observed as
brownish-yellow granular staining. Under normal conditions,
β-catenin expression is located in the cell membrane of cancer
cells. Cells with membranes exhibiting positive expression of
β-catenin of >70% were defined as normal expression cells. Under
abnormal conditions, β-catenin expression is distributed in the
cell cytoplasm and nucleus of cancer cells. Cells that exhibited
positive expression of β-catenin in the cytoplasm or nucleus of
>10% were defined as abnormal expression cells.
Positive expression of cyclin D1 was located in the
nucleus, and the positive cells exhibited a brownish-yellow
granular pigment. A total of 500 cells were counted under a DM LB2
microscope (Leica Microsystems GmbH, Wetzlar, Germany) at a high
magnification. The positive staining rate of the cells was
calculated, and the cells with a positive rate of >10% were
defined as overexpression cells.
ER-β positive expression was shown as brown granules
in the nucleus. In total, five fields were selected at random under
high magnification. Cells exhibiting positive staining were
counted, and the percentage of ER-β positive staining was
calculated as the number of positively stained cells to the total
cell number. An ER-β positive staining rate of ≥10% was defined as
ER-β positive expression.
Statistical analysis. Statistical analysis was
performed using SPSS software, version 13.0 (SPSS, Inc., Chicago,
IL, USA). Differences between groups were analyzed using the
χ2 test, while the Kaplan-Meier method was used for
survival analysis. P<0.05 was considered to indicate a
statistically significant difference.
Results
Expression of β-catenin, cyclin D1 and
ER-β in human breast cancer tissues
In order to determine the expression levels of
β-catenin, cyclin D1 and ER-β, immunohistochemical staining assays
were performed in 226 cases of human breast cancer tissues.
Representative immunohistochemical staining results are shown in
Fig. 1. Positive staining in the
cells was shown as a brownish-yellow granular pigment. Normal
expression of β-catenin in the cell membrane is shown in Fig. 1A. These cells were counted and the
normal β-catenin expression rate was calculated. Cells exhibiting
positive expression of β-catenin in the membrane of >70% were
defined as cells with normal expression. Ectopic expression of
β-catenin in the cytoplasm or the nucleus was considered abnormal
expression (Fig. 1B). After
counting, the abnormal expression rate was calculated, in which
cells with cytoplasm- or nucleus-positive expression of β-catenin
of >10% were defined as abnormal expression cells. A negative
control for cyclin D1 expression and a representative sample with
positive cyclin D1 expression are shown in Fig. 1C and D, respectively. In addition, a
negative control for ER-β expression and a representative sample
with positive ER-β expression are shown in Fig. 1E and F, respectively. Positively
stained cells were counted and the positive expression rate was
calculated. Cells with a positive cyclin D1 expression rate of
>10% were defined as overexpression cells, while cells with a
positive ER-β expression rate of ≥10% were defined as
overexpression cells.
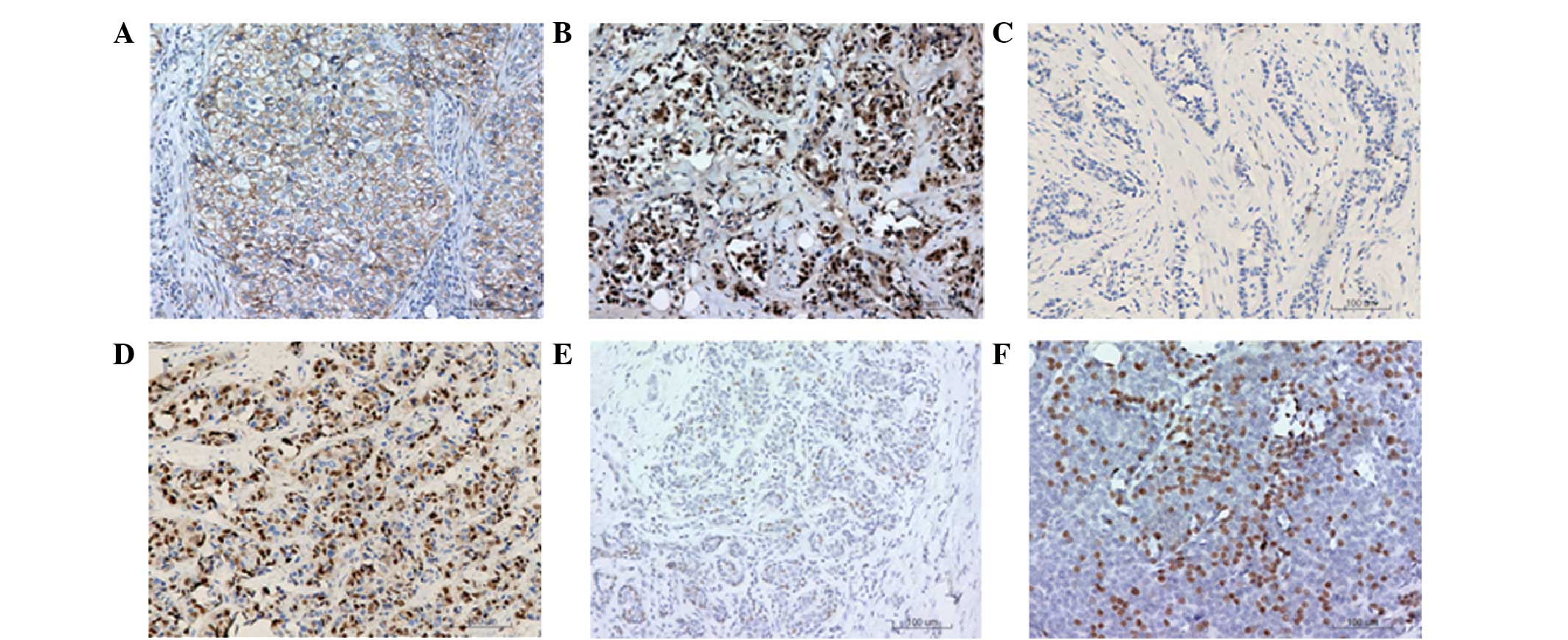 | Figure 1.Immunohistochemical staining of
β-catenin, cyclin D1 and estrogen receptor (ER)-β in human breast
cancer tissues. Representative results are shown, where cells
exhibiting a brownish-yellow granular pigment are positively
stained. (A) Normal expression of β-catenin in the cell membranes
of breast cancer tissues (magnification, ×400). (B) Abnormal
expression of β-catenin in the cell cytoplasm and nuclei of breast
cancer tissues (magnification, ×400). (C) Negative control for
cyclin D1 (magnification, ×200), where the primary antibody
(anti-cyclin D1) was replaced with phosphate-buffered saline (PBS).
(D) Cyclin D1 positive expression in breast cancer cell nuclei
(magnification, ×200). (E) Negative control for ER-β
(magnification, ×200), where the primary antibody (anti-ER-β) was
replaced with PBS. (F) ER-β positive expression in breast cancer
tissue (magnification, ×200). |
Breast cancer tissues with abnormal
β-catenin expression exhibit a higher cyclin D1 positive expression
rate
To assess the association between β-catenin and
cyclin D1 expression, the cases with abnormal β-catenin expression
and positive cyclin D1 expression were calculated. Subsequently,
the cyclin D1 positive expression rate in β-catenin normal and
abnormal expression tissues was analyzed (Table I). In total, 56 cases exhibited
normal β-catenin expression and 170 cases exhibited abnormal
β-catenin expression, with an abnormal β-catenin expression rate of
75.2%. Furthermore, a total of 52 cases were cyclin D1-negative and
174 cases exhibited positive cyclin D1 expression, with a positive
cyclin D1 expression rate of 77.0%. The cyclin D1 positive
expression rate in the normal β-catenin expression tissues was
50.0% (28/56), while in the abnormal β-catenin expression tissues,
the cyclin D1 positive expression rate was 85.9% (146/170). The
cyclin D1 positive expression rate in the abnormal β-catenin
expression tissues was higher compared with the normal β-catenin
expression tissues, and the difference was statistically
significant (P<0.05; Table II).
Thus, breast cancer tissues with abnormal β-catenin expression
exhibited an elevated cyclin D1 positive expression rate.
| Table II.Association between β-catenin and
cyclin D1 expression. |
Table II.
Association between β-catenin and
cyclin D1 expression.
| β-catenin
expression | Cases (n) | Cyclin D1 negative, n
(%) | Cyclin D1 positive, n
(%) | χ2 | P-value |
|---|
| Normal | 56 | 28 (50.0) | 28
(50.0) |
|
|
| Abnormal | 170 | 24 (14.1) | 146 (85.9) | 30.616 | <0.001 |
| Total cases | 226 | 52 (23.0) | 174 (77.0) |
|
|
Breast cancer tissues with abnormal
β-catenin expression exhibit a higher ER-β positive expression
rate
To analyze the association between ER-β expression
and β-catenin expression, the number of cases with ER-β positive
expression was calculated. Subsequently, the ER-β positive
expression rate in the β-catenin normal and abnormal expression
tissues was compared (Table III).
ER-β expression was positive in 98 breast cancer tissue samples and
negative in 128 cases, with a positive expression rate of 43.4%. In
the normal β-catenin expression tissues, the ER-β positive
expression rate was 35.7% (20/56), while in the abnormal β-catenin
expression tissues, the rate was 45.9% (78/170). The difference
between these two groups was statistically significant (P<0.05).
Therefore, higher ER-β positive expression levels in breast cancer
tissues were associated with abnormal β-catenin expression.
| Table III.Association between β-catenin and ER-β
expression. |
Table III.
Association between β-catenin and ER-β
expression.
| β-catenin
expression | Cases (n) | ER-β negative, n
(%) | ER-β positive, n
(%) | χ2 | P-value |
|---|
| Normal | 56 | 36
(64.3) | 20 (35.7) |
|
|
| Abnormal | 170 | 92
(54.1) | 78 (45.9) | 1.773 | 0.183 |
| Total cases | 226 | 128 (56.6) | 98 (43.4) |
|
|
Breast cancer tissues with cyclin D1
positive expression exhibit a higher ER-β positive expression
rate
The association between cyclin D1 expression and
ER-β expression was determined by calculating the positive
expression rate of ER-β in cyclin D1 positive expression tissues.
As shown in Table IV, the ER-β
positive expression rate in the cyclin D1 negative expression group
was 26.9% (14/52), while in the cyclin D1 positive expression
group, the rate was 48.4% (84/174). The ER-β positive expression
rate in the cyclin D1 positive expression tissues was significantly
higher compared with the cyclin D1 negative expression tissues
(P<0.05), indicating that breast cancer tissues with cyclin D1
positive expression possess a higher ER-β positive expression rate
compared with cyclin D1 negative expression.
| Table IV.Association between cyclin D1 and
ER-β expression. |
Table IV.
Association between cyclin D1 and
ER-β expression.
| Cyclin D1
expression | Cases (n) | ER-β negative, n
(%) | ER-β positive, n
(%) | χ2 | P-value |
|---|
| Negative | 52 | 38
(73.1) | 14 (26.9) |
|
|
| Positive | 174 | 90
(51.7) | 84 (48.4) | 7.432 | 0.006 |
| Total cases | 226 | 128 (56.6) | 98 (43.4) |
|
|
Patients with normal β-catenin
expression exhibit an extended survival time
To determine the effect of β-catenin expression on
survival, the tumor-free survival times were analyzed using the
Kaplan-Meier method. The resulting survival curves are presented in
Fig. 2. The median survival time of
patients with normal β-catenin expression was 11.6 years, while
that of patients with abnormal β-catenin expression was 10.5 years.
The difference between the patients with normal β-catenin
expression and those with abnormal β-catenin expression was
statistically significant (P=0.049). Therefore, the median survival
time was longer in patients with normal β-catenin expression.
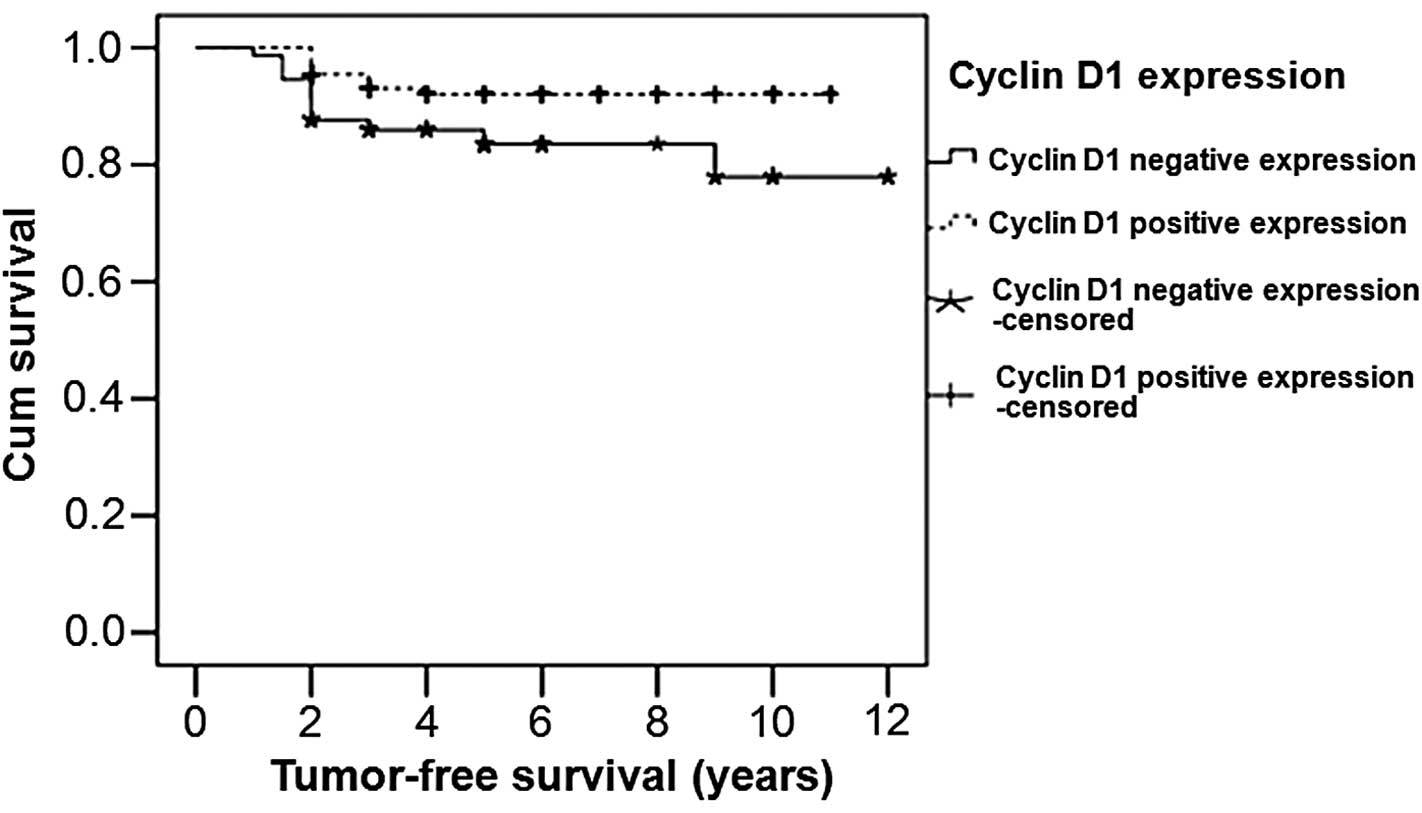
Patients with cyclin D1 positive
expression have longer survival times
For survival analysis in the patients with a
different cyclin D1 expression status, the tumor-free survival time
was analyzed using the Kaplan-Meier method. The survival curves are
shown in Fig. 3. The median survival
time of the cyclin D1 positive expression patients was 10.230
years, which was higher compared with that of the cyclin D1
negative expression patients (10.177 years). A statistically
significant difference was identified between the patients with
positive cyclin D1 expression and those with negative cyclin D1
expression (P=0.026). Therefore, patients with cyclin D1 positive
expression exhibited a longer survival time compared with those
with cyclin D1 negative expression.
Discussion
The Wnt signaling pathway is activated in the vast
majority of breast cancer tissues, and has been shown to be
involved in the tumorigenesis of breast tissue (9). In the classical Wnt pathway, β-catenin
serves a key function. In cells that have been activated by Wnt
protein ligands, cytoplasmic β-catenin is prevented from
degradation and stably accumulates in the cytoplasm. The
accumulated β-catenin is subsequently transferred into the nucleus
where it binds with the TCF/LEF, enhancing the transcription of
genes involved in cell proliferation, and thus inducing
carcinogenesis. Cyclin D1 is a crucial target gene in the Wnt
signaling pathway (10). Previous
studies have reported that abnormal expression of β-catenin and
cyclin D1 is associated with breast cancer occurrence and
development (11,12). In addition, clinical studies have
indicated that the β-catenin and cyclin D1 expression rates in
breast cancer tissues were 51–89% and 45–83%, respectively
(13,14). In accordance, the results of the
present study revealed the abnormal expression rate of β-catenin as
75.2% and the positive expression rate of cyclin D1 as 77.0% in the
226 breast cancer samples.
Currently, the association between β-catenin and
cyclin D1 expression in breast cancer tissue remains controversial.
For example, studies by Ozaki et al (15) and Lin et al (12) demonstrated that the abnormal
expression of β-catenin was associated with the overexpression of
cyclin D1. However, Lim and Lee (11) indicated that the abnormal expression
of β-catenin was not associated with cyclin D1 overexpression. Yang
et al (16) analyzed
β-catenin and cyclin D1 expression in 60 breast cancer tissues
using an immunohistochemical method. The authors identified 42
cases with abnormal β-catenin expression, in which 57.1% exhibited
cyclin D1 overexpression, indicating a significant positive
correlation. Thus, it was concluded that abnormal expression of
β-catenin may lead to the occurrence and development of breast
cancer by inducing or activating cyclin D1 overexpression. In the
226 breast cancer samples analyzed in the present study, the cyclin
D1 positive expression rate in the β-catenin abnormal expression
tissues was significantly higher compared with the β-catenin normal
expression tissues. This result indicated that Wnt signaling is
over-activated in breast cancer tissues, resulting in abnormal
expression of β-catenin, which may induce or activate cyclin D1
overexpression, leading to the occurrence and development of breast
cancer.
Lazennec et al (17) observed that ER-β expression levels in
breast cancer tissues were significantly reduced compared with
normal breast tissues, and the loss of ER-β expression resulted in
the occurrence of breast cancer. Numerous studies have demonstrated
that overexpression of cyclin D1 is consistent with ER-α positive
expression (4,18,19).
However, there are a limited number of studies investigating the
association between the Wnt pathway and ER-β expression. Zwijsen
et al (20) reported that
cyclin D1 overexpression in breast cancer tissues was associated
with the expression of the estrogen response element. Cyclin D1 may
perform a similar function to estrogens. Furthermore, the authors
hypothesized that the effects of estrogen on breast tumors may be
exerted via the cyclin D1 pathway, and that ER-α may promote breast
cancer occurrence through the induction of cyclin D1. However, Luo
et al (5) indicated that the
ER pathway was connected with the Wnt signaling pathway through
β-catenin. ER pathway activation may activate β-catenin, which may
further activate the Wnt signaling pathway and result in the
increased expression of cyclin D1. The present results indicated
that the difference in the ER-β positive expression rate between
the normal and abnormal β-catenin expression tissues was
statistically significant. Furthermore, the ER-β positive
expression rate in the cyclin D1 positive expression tissues was
significantly higher compared with the cyclin D1 negative tissues,
indicating that cyclin D1 overexpression may be closely associated
with ER-β positive expression.
A previous study indicated that β-catenin expression
is increased in breast cancer tissues, and is associated with a
poor prognosis (21). Lin et
al (12) detected the β-catenin
expression levels of 123 breast cancer samples using an
immunohistochemical method, and found that cytoplasmic expression
of β-catenin represented the activated state of β-catenin and that
β-catenin was an independent prognostic factor for breast cancer
survival. Nakopoulou et al (22) examined 141 breast cancer specimens,
and reported that nuclear β-catenin expression correlated with
reduced overall survival (OS) and disease free survival (DFS)
times, while cytoplasmic β-catenin expression was associated with
longer OS and DFS times. In the studies by Lin et al
(12), Lim and Lee (11) and López-Knowles et al
(23), the abnormal expression of
β-catenin in breast cancer tissues was shown to be associated with
a poor prognosis, while β-catenin may be used as an independent
prognostic factor for breast cancer evaluation. However, Chung
et al (24) indicated that
abnormal expression of β-catenin was not associated with prognosis,
and was only associated with a poor prognosis in cases that
presented with abnormal expression of β-catenin and overexpression
of p53 simultaneously. Thus, Chung et al hypothesized that
β-catenin was unable to be used an independent prognostic factor
for breast cancer. In the present study, the median tumor free
survival time of the patients with normal β-catenin expression was
higher compared with the patients with abnormal β-catenin
expression, and the difference was statistically significant.
Furthermore, patients with positive cyclin D1 expression exhibited
a longer survival time compared with those with negative cyclin D1
expression. These results are consistent with those of previous
studies (25–30), and indicate that patients with
positive cyclin D1 expression may expect an improved prognosis.
In conclusion, the results of the present study
indicate that the expression of β-catenin, cyclin D1 and ER-β in
breast cancer tissues are associated with each other, and may serve
crucial functions in the development of breast cancer.
Acknowledgements
This study was supported by grants from the National
Natural Science Foundation of Xinjiang Uygur Autonomous Region (no.
2011211A069) and the National Clinical Key Subject General Surgery
Construction Project.
Abbreviations:
|
DFS
|
disease free survival
|
|
ER
|
estrogen receptor
|
|
OS
|
overall survival
|
|
PBS
|
phosphate-buffered solution
|
|
TCF/LEF
|
T-cell factor/lymphoid enhancer
factor
|
|
HRP
|
horseradish peroxidase
|
|
SP
|
streptavidin peroxidase
|
|
DAB
|
3,3′-diaminobenzidene
|
References
|
1
|
Parkin DM, Bray F, Ferlay J and Pisani P:
Global cancer statistics, 2002. CA Cancer J Clin. 55:74–108. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Kohlberger PD, Breitenecker F, Kaider A,
Lösch A, Gitsch G, Breitenecker G and Kieback DG: Modified
truecolor computer-assisted image analysis versussubjective scoring
of estrogen receptor expression in breast cancer: A comparison.
Anticancer Res. 19:2189–2193. 1999.PubMed/NCBI
|
|
3
|
Kikuchi A: Regulation of beta-catenin
signaling in the Wnt pathway. Biochem Biophys Res Commun.
268:243–248. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Mauro L, Pellegrino M, Giordano F, Ricchio
E, Rizza P, De Amicis F, Catalano S, Bonofiglio D, Panno ML and
Andò S: Estrogen receptor-α drives adiponectin effects on cyclin D1
expression in breast cancer cells. FASEB J. 29:2150–2160. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Luo J, Chen YL and Xu H: Expression of
β-catenin, cyclin D1 and ERα in breast cancer. Shi Yong Yu Fang Yi
Xue. 17:1502–1504. 2010.(In Chinese).
|
|
6
|
Kuiper GG, Enmark E, PeltoHuikko M,
Nilsson S and Gustafsson JA: Cloning of a novel receptor expressed
in rat prostate and ovary. Proc Natl Acad Sci USA. 93:5925–5930.
1996. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Speirs V, Parkes AT, Kerin MJ, Walton DS,
Carleton PJ, Fox JN and Atkin SL: Coexpression of estrogen receptor
α and β: Poor progonostic factors in human breast cancer. Cancer
Res. 59:525–528. 1999.PubMed/NCBI
|
|
8
|
Speirs V, Adams IP, Walton DS and Atkin
SL: Identification of wild-type and exon 5 deletion variants of
estrogen receptor beta in normal human mammary gland. J Clin
Endocrinol Metab. 85:1601–1605. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Howe LR and Brown AM: Wnt signaling and
breast cancer. Cancer Biol Ther. 3:36–41. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Bala S and Peltomäki P: CYCLIN D1 as a
genetic modifier in hereditary non polyposis colorectal cancer.
Cancer Res. 61:6042–6045. 2001.PubMed/NCBI
|
|
11
|
Lim SC and Lee MS: Significance of
E-cadherin/beta-catenin complex and cyclin D1 in breast cancer.
Oncol Rep. 9:915–928. 2002.PubMed/NCBI
|
|
12
|
Lin SY, Xia W, Wang JC, et al:
Beta-catenin, a novel prognostic marker for breast cancer: Its
roles in cyclin D1 expression and cancer progression. Proc Natl
Acad Sci USA. 97:4262–4266. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Brabletz T, Jung A, Reu S, et al: Variable
beta-catenin expression in colorectal cancers indicates tumor
progression driven by the tumor environment. Proc Natl Acad Sci
USA. 98:10356–10361. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Paola C, Anna P, Marina R, Gnesi E,
Baldini E and Bevilacqua G: Cyclin D1 expression in node positive
(N+) and node-negative (N-) infiltrating human mammary carcinomas.
Int J Cancer. 84:139–144. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Ozaki S, Ikeda S, Ishizaki Y, et al:
Alterations and correlations of the components in the Wnt signaling
pathway and its target genes in breast cancer. Oncol Rep.
14:1437–1443. 2005.PubMed/NCBI
|
|
16
|
Yang JF, Chen SL, Liu ZH and Zhang Y:
Correlation among expression of E-cadherin, beta-catenin, and
cyclin D1 in breast cancers. Ai Zheng. 23:799–802. 2004.(In
Chinese). PubMed/NCBI
|
|
17
|
Lazennec G, Bresson D, Lucas A, Chauveau C
and Vignon F: ER beta inhibits proliferation and invasion of breast
cancer cells. Endocrinology. 142:4120–4130. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
CastroRivera E, Samudio I and Safe S:
Estrogen regulation of cyclin D1 gene expression in ZP-75 breast
cancer cells involves multiple enhancer elements. J Biol Chem.
276:30853–30861. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Ravikumar G and Ananthamurthy A: Cyclin D1
expression in ductal carcinoma of the breast and its correlation
with other prognostic parameters. J Cancer Res Ther. 10:671–675.
2014.PubMed/NCBI
|
|
20
|
Zwijsen R, Wientjens E, Klompmaker R, van
der Sman J, Bernards R and Michalides RJ: CDK-independent
activation of estrogen receptor by cyclin D1. Cell. 88:405–415.
1997. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Musgrove EA: Wnt signaling via the
epidermal growth factor receptor: A role in breast cancer? Breast
Cancer Res. 6:65–68. 2004. View
Article : Google Scholar : PubMed/NCBI
|
|
22
|
Nakopoulou L, Mylona E, Papadaki I,
Kavantzas N, Giannopoulou I, Markaki S and Keramopoulos A: Study of
phospho-beta-catenin subcellular distribution in invasive breast
carcinomas in relation to their phenotype and the clinical outcome.
Mod Pathol. 19:556–563. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
López-Knowles E, Zardawi SJ, McNeil CM, et
al: Cytoplasmic localization of beta-catenin is a marker of poor
outcome in breast cancer patients. Cancer Epidemiol Biomarkers
Prev. 19:301–309. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Chung GG, Zerkowski MP, Ocal IT, et al:
beta-Catenin and p53 analyses of a breast carcinoma tissue
microarray. Cancer. 100:2084–2092. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Gillett C, Smith P, Gregory W, Richards M,
Millis R, Peters G and Barnes D: Cyclin D1 and prognosis in human
breast cancer. Int J Cancer. 69:92–99. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Steeg PS and Zhou Q: Cyclins and breast
cancer. Breast Cancer Res Treat. 52:17–28. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Diana M and Barnes CEG: Cyclin D1 in
breast cancer. Acta Oncology. 24:4211995.
|
|
28
|
Bates S, Parry D, Bonetta L, Vousden K,
Dickson C and Peters G: Absence of cyclin D/CDK complexes in cells
lacking functional retinoblastoma protein. Oncogene. 9:1633–1640.
1994.PubMed/NCBI
|
|
29
|
Quintayo MA, Munro AF, Thomas J, et al:
GSK3β and cyclin D1 expression predicts outcome in early breast
cancer patients. Breast Cancer Res Treat. 136:161–168. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Peurala E, Koivunen P, Haapasaari KM,
Bloigu R and Jukkola-Vuorinen A: The prognostic significance and
value of cyclin D1, CDK4 and p16 in human breast cancer. Breast
Cancer Res. 15:R52013. View
Article : Google Scholar : PubMed/NCBI
|