Introduction
Asthma is a disease that features chronic bronchial
inflammation, exaggerated bronchial response and airway obstruction
(1). Immune tolerance has been
demonstrated to be mediated by regulatory T (Treg) cells, defined
as a sub-population of CD4+ T cells that mediates the
immune reaction in the lungs through the prohibition of the
proliferation and activities of effecting factors secreted by other
T cells (2). T helper 17 (Th17)
cells consist of a definitive lineage of pro-inflammatory Th cells,
which serve a major role in autoallergic diseases (3). Experimental data have suggested an
association between the pathology induced by Th17 cells and the
regulatory role of Treg cells (4).
Murine models have demonstrated that Th17 cells induce autoimmune
response by promoting tissue inflammation and activating the innate
immune system (5). Treg cells, a
subgroup of T lymphocytes with features of immune repression on
effector T cells, can be recognized by the expression of the high
affinity α-chain receptor of interleukin (IL)-2, namely CD25.
However, CD25 is also an upregulated marker of the activated
CD4+ T cells, and is thus considered as a non-exclusive
marker. In addition, further distinction of CD4+
CD25high Treg cells from other T cells can be achieved
based on the selective expression of the forkhead box P3 (FOXP3)
transcription factor, since this expression has a central role in
the growth and function of Treg cells (6). Although the underlying mechanism
through which the proliferation of other T cells is suppressed by
Treg cells has not been elucidated, previous evidence has revealed
that Treg cells are involved in the prevention of autoimmune
response and the control of pulmonary and gastric inflammation
in vivo (7,8).
Increasing evidence has demonstrated that Th17 and
Treg cells are essential T cell subsets and are critical in
maintaining the homeostasis of the immune system. The native T cell
precursor pool responsible for the generation of Treg cells can
also generate CD4+ Th17 cells that may secrete IL-17
cytokines (9). In previous animal
models, numerous inflammation cytokines were found to be involved
in the differentiation of T cells, which are responsible for the
activation of adaptive immune response. The potential role of IL-6,
transforming growth factor (TGF)-β, IL-10 and IL-23 in the
promotion of human Th17 cell differentiation has been revealed in a
previous study (10), although the
exact cytokine combination requires further verification. It has
been suggested that IL-6 or IL-23 are indispensable for the ability
of activated T cells to secrete IL-17, while IL-23 along with TGF-β
are essential for Th17 cell differentiation from primitive T cells
(11,12). Cytokine levels in have been shown to
orchestrate the development of these cells in a temporal and
spatial manner (13–15). Th17 cells are potent activators of
neutrophils to enable the eradication of pathogens (16,17).
Although Th17 cells are considered to be biologically crucial in
eradicating extracellular pathogens, the abnormal expression of
IL-17a by these cells is considered as a factor participating in
the pathogenesis of various airway inflammatory disorders. For
instance, elevated levels of IL-17a have been identified in
rheumatoid arthritis, multiple sclerosis and systemic lupus
erythematosus (18). IL-10 is
upregulated in the lungs of asthma patients, indicating that an
immune response driven by Th17 cells contributes to the
pathogenesis of asthma (19). The
balance between Th17 and Treg cells may be necessary to maintain
the immune homeostasis (20);
however, no previous studies have analyzed this balance in
individual asthma patients to date.
The aim of the present study was to examine whether
the imbalance of Th17 and Treg cells had an important role in
asthma patients. A stringent flow cytometry-based staining and
gating strategy was developed to accurately enumerate the Treg and
Th17 cells using FOXP3 and IL-17a, respectively, as the defining
markers. Each cell type was investigated individually, and was
compared in the same subgroup of patients simultaneously. It was
hypothesized that the imbalance observed in cell types may be
caused by the expression of FOXP3+ and pro-inflammatory
cytokines. Therefore, the expression levels of IL-6, IL-10, IL-23
and TGF-β were detected in the bronchoalveolar lavage fluid (BALF)
specimens of asthma patients and controls. The present study
focused on the regulation of the Treg cells ratio on the cytokine
environment in an inflammatory condition.
Patients and methods
Patients
Asthma patients treated at the Department of
Respiratory Medicine of the Shandong Provincial Hospital (Jinan,
China) between March 2014 and September 2014 were retrospectively
analyzed in the present study. Informed consent was acquired from
all patients prior to sample collection. Ethical approval for the
present study was granted by the Ethical Committee of Wuxi People's
Hospital of Nanjing Medical University, and was performed according
to the National Statement on Ethical Conduct in Research Involving
Humans (1999) issued by the National Health and Medical Research
Council of China. In total, 25 allergy asthma patients (female, 11;
male, 14) and 18 control individuals (female, 7; male, 11) were
enrolled into the study, and blood samples were collected. All
asthma patients did not present clinical exacerbation at the time
of blood sample collection, based on the evaluation of clinical
symptoms, signs and serum C-reactive protein levels (<10 mg/l).
Subjects in the control group were all non-allergic healthy
volunteers. The blood sample was collected and stored in the EDTA
tubes. Patients were included if they fulfilled the following
criteria: i) Positive diagnosis (ii) no traumatic and inflammatory
treatment; and iii) no surgical intervention. Patients exhibited
mild (n=11), moderate (n=10) and severe (n=4) severe asthma.
Flow cytometry analysis
Peripheral blood mononuclear cells (PBMCs) were
separated via density gradient centrifugation in lymphocyte
separation medium (Ficoll-Paque Plus; GE Healthcare, Chalfont, UK)
at 600 × g for 20 min at 4°C. In order to recognize the Treg cells,
1×106 PBMCs were surface stained with anti-CD4
monoclonal antibodies (1:50; 564419) labeled with fluorescein
isothiocynate, anti-CD8 (1:50; RPA-T8) and anti-CD25 antibodies
(1:50; 561399) labeled with phycoerythrin (PE)-Cyanine 5 (Cy5; all
from BD Biosciences, Sydney, Australia). Next, permeabilization in
the FOXP3 fix/perm solution (eBioscience, Inc., San Diego, CA, USA)
was performed, and then intracellular labeling was conducted with
an anti-FOXP3 antibody conjugated with PE (1:50; PCH101;
eBioscience, Inc.), according to the manufacturer's protocol. For
IL-17a assays, PBMCs (2×106) were stimulated for 5 h
using 50 ng/ml phorbol 12-myristate 13-acetate (PMA) and 1 g/ml
ionomycin in the presence of 5 g/ml brefeldin A solution (all from
Sigma-Aldrich; Merck Millipore, Darmstadt, Germany), incubated with
5% CO2 at 37°C. Cells were washed twice in
phosphate-buffered saline (PBS) and surface labeled with
CD3-PE-Cy5, prior to fixing with 4% w/v paraformaldehyde and
permeabilizing with 0.1% w/v saponin (Sigma-Aldrich; Merck
Millipore). Subsequent to permeabilization, all samples were washed
with PBS. Cells were blocked with 5% w/v separated milk powder in
PBS/0.1% w/v saponin for 30 min, followed by intracellular labeling
with anti-IL17a-PE (clone ebio64DEC17; eBioscience, Inc.). Flow
cytometry was conducted with the BD FACScan system (BD
Biosciences), with 300,000–500,000 events gathered in each
experiment and lymphocytes gated based on the properties of their
forward and side light scatter. Stained cells were all analyzed by
a BD LSR II flow cytometer (BD Biosciences), and the flow cytometry
data were analyzed by FlowJo 7.6 software (FlowJo, LLC, Ashland,
OR, USA).
RNA isolation and analysis
RNA from whole blood collected in EDTA tubes was
isolated via an organic extraction technique, using TRIzol LS
(Thermo Fisher Scientific, Inc., Waltham, MA, USA) according to the
manufacturer's protocols. Further purification of DNA was performed
on a QIAamp spin column (Qiagen, Hilden, Germany) and on-column
DNase digestion. mRNA expression levels of FOXP3 and IL-17a in the
blood were determined by reverse transcription-quantitative
polymerase chain reaction (RT-qPCR) with the use of a QuantiTect RT
kit (Qiangen) and a StepOnePlus Real-Time PCR system (Applied
Biosystems; Thermo Fisher Scientific, Inc.). qPCR was performed
with the following thermal cycles: 95°C for 10 min, followed by
95°C for 15 sec, 60°C for 30 sec and 72°C for 30 sec for 40 cycles
using cDNA (2 µl), forward primer (0.4 µl), reverse primer (0.4
µl), SYBR Green solution (10 µl), and ddH2O (7.2 µl).
Specific primers for FOXP3, IL-17a and β-actin used were as
previously reported (21). To
quantify the results, 2−ΔΔCq was calculated for every
sample (22), and the mRNA
expression levels were normalized to β-actin.
IL-6, IL-23, IL-10 and TGF-β
expression levels determined by ELISA
Expression levels of IL-6 (EH2IL62), IL-23 (KHC0231;
both Thermo Fisher Scientific, Inc.), IL-10 (RAB1060;
Sigma-Aldrich; Merck Millipore)and TGF-β (E-EL-H0110c; Elabscience
Bioengineering Co., Ltd., Wuhan, China) were detected in BALF
samples obtained from the patients and control subjects using
respective ELISA kits according to the manufacturer's protocols.
All samples were measured in duplication. Optical densities were
determined at 450 nm using a microplate reader.
Statistical analysis
All analyses were performed with the SPSS
statistical software for Windows (version 10.1.4; SPSS, Inc.,
Chicago, IL, USA). Data are expressed as the mean ± standard error.
In paired comparison, Student t test and Wilcoxon rank-sum
(Mann-Whitney U) test were used for normally or non-normally
distributed data, respectively. One-way analysis of variance and
Kruskal-Wallis tests were used in the comparison of multiple
groups. For all the analyses, two-sided P-values of <0.05 were
considered to indicate statistically significant differences.
Results
Patients and sampling
Blood samples were collected from 25 asthma patients
(female, 11; male, 14) and 18 controls without lung diseases
(female, 7; male, 11), with mean ages of 38.7±3.1 and 45.8±2.8
years, respectively. In addition, BALF samples were collected from
15 allergy asthma patients and 10 controls, with mean ages of
30.9±7.63 and 46.8±15.1 years, respectively. There were no
significant differences in age, gender and other clinical data
among the patients and healthy subjects.
Reduced number of CD4+
CD25bright FOXP3+ Treg cells in the
peripheral blood of asthma patients
In the present study, the highest intensity of
CD25+ staining was ranked as the brightest 0.5%
CD4+ CD25+cells for all patient samples. As
shown in Fig. 1, the flow cytometric
analysis demonstrated that patients with asthma generally exhibited
a statistically decreased absolute number of Treg cells in each ml
of peripheral blood, compared with the controls. The mean number of
Treg cells/ml was found to be significantly lower in asthma
patients (5.56±3.3×103/ml) in comparison with the
healthy controls (8.98±4.3×103/ml; P=0.0093). In
addition, the proportion of CD4+ CD25bright
FOXP3+ Treg cells among PBMCs was 0.012–0.51% in asthma
patients and 0.121–0.55% in the control group (data not shown).
Elevated number of Th17 cells in the
peripheral blood of asthma patients
The count of Th17 cells in the blood was assessed by
flow cytometry in order to determine whether it was altered in
asthma patients compared with the controls. As the stimulation of
PMA/ionomycin can lead to decreased human CD4 expression,
CD3+CD8− was selected as a marker for
CD4+ Th17 cells, according to a previous study (23). CD3+CD8− gating
was performed in PBMCs to identify IL-17+ cells by
distinguishing Th17 cells from the cluster of T cells in PBMCs. The
results indicated that the percentage of Th17 cells in the sample
of PBMCs in asthma patients was 0.36–1.25%, while this percentage
was 0.10–0.49% in the control group (data not shown). As shown in
Fig. 2, the number of Th17 cells per
ml was significantly higher in the blood of asthma patients when
compared with the control group (16.45±7.0×103/ml vs.
7.44±4.60×103/ml, respectively; P=0.007).
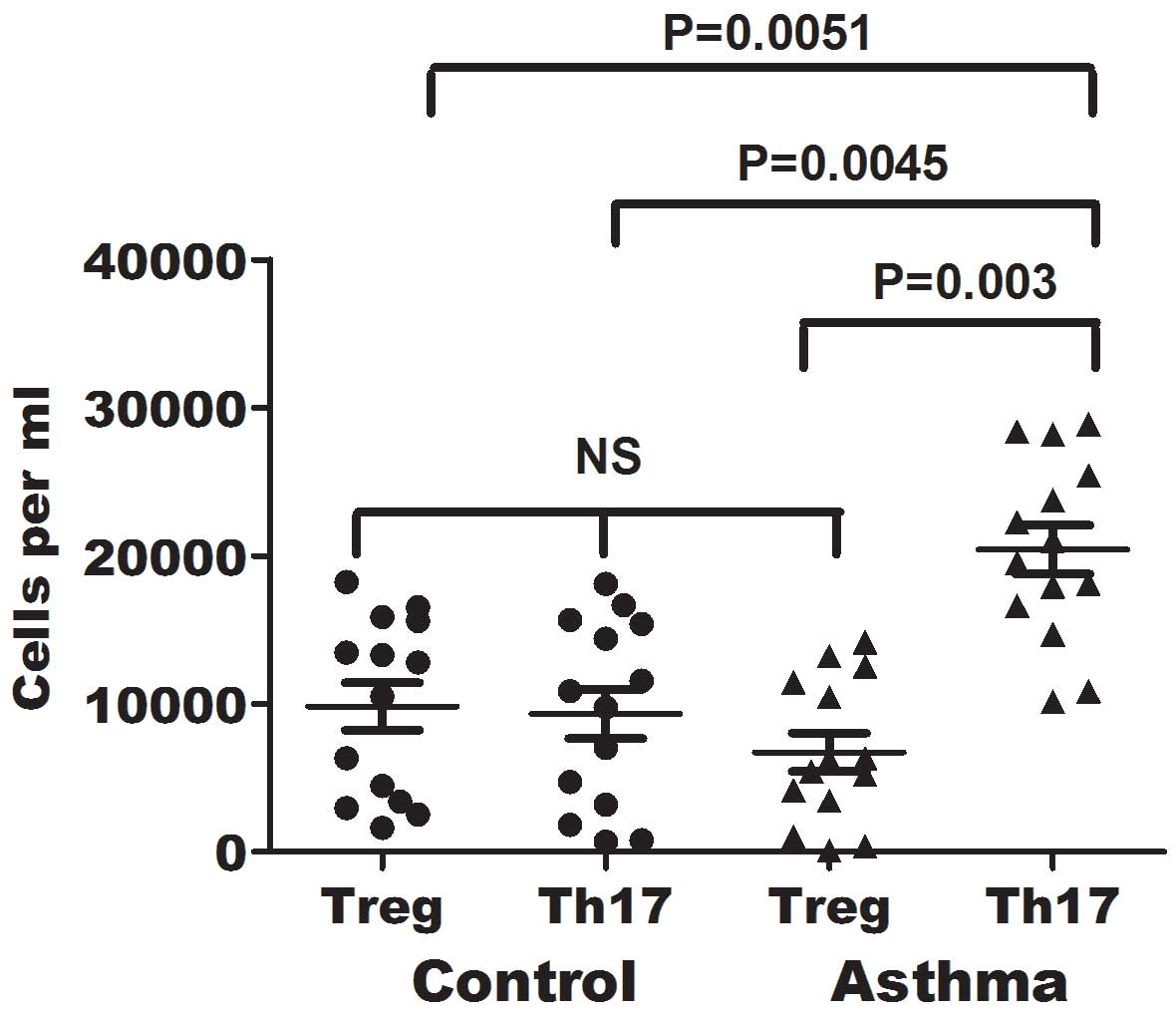
Imbalance between Treg and Th17 cells
in the blood of asthma patients
As shown earlier, the number of Treg cells was
decreased, while that of Th17 cells was elevated in asthma
patients. This association was further investigated to determine
the association of Treg and Th17 cell counts in the asthma patients
and corresponding control individuals. The number of Treg and Th17
cells in the peripheral blood specimens of asthma patients was
directly measured simultaneously for comparison, and the balance of
Treg and Th17 cells was assessed in 14 controls and 14 asthma
patients. As shown in Fig. 3, the
ratio of Th17/Treg was significantly elevated in patients with
asthma in comparison with that in healthy individuals (P=0.003),
whereas the numbers of Treg and Th17 cells were comparable in the
control group. Thus, an imbalance between Treg and Th17 cells was
observed in the peripheral blood of asthma patients (Fig. 3), due to the observed reduction in
the number of Treg cells and increase in the number of Th17 cells.
No significant differences in the number of Treg cells were
detected between the asthma and control groups. However, a
significant increase in the number of Th17 cells was noted in
asthma patients, when compared with healthy control subjects
(P=0.0045).
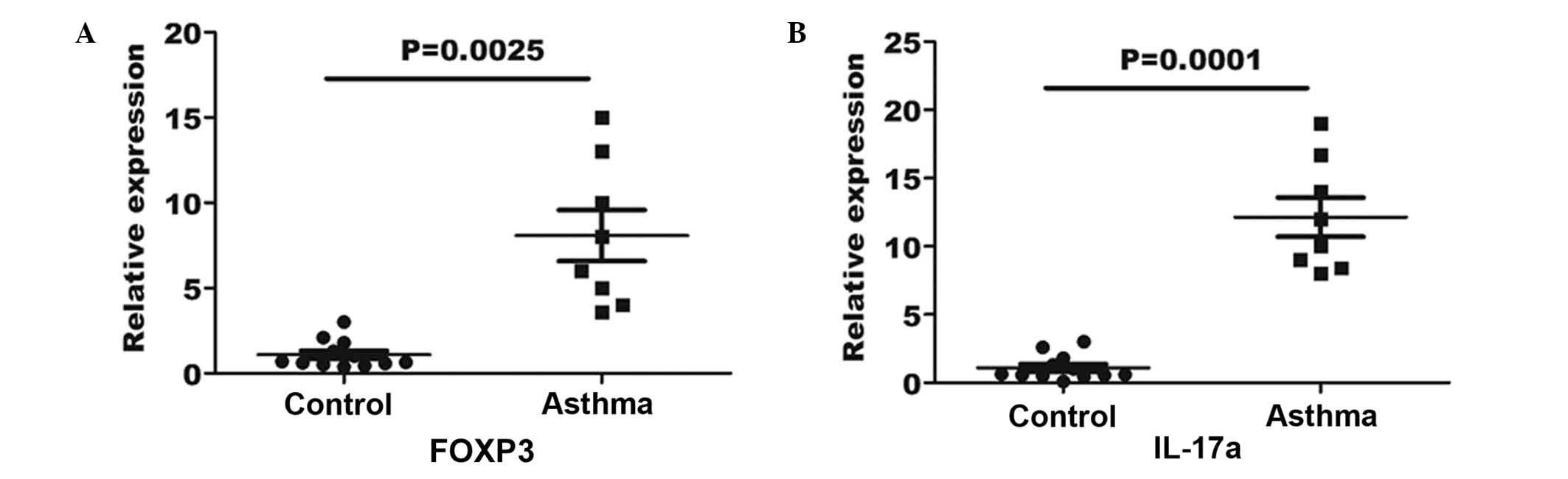
Upregulated gene expression levels of
FOXP3 and IL-17a in the peripheral blood of asthma patients
In order to verify the Treg and Th17 cell numbers in
the asthma patients and healthy controls, RT-qPCR was performed for
the measurement of FOXP3 and IL-17a gene expression in the blood
specimens. An 8-fold elevation in FOXP3 gene expression was
observed in the asthma blood samples in comparison with the
expression in the controls (P=0.0025; Fig. 4A). Furthermore, a 12-fold elevation
in IL-17a expression was determined in asthma patients in
comparison with the controls (P=0.0001; Fig. 4B).
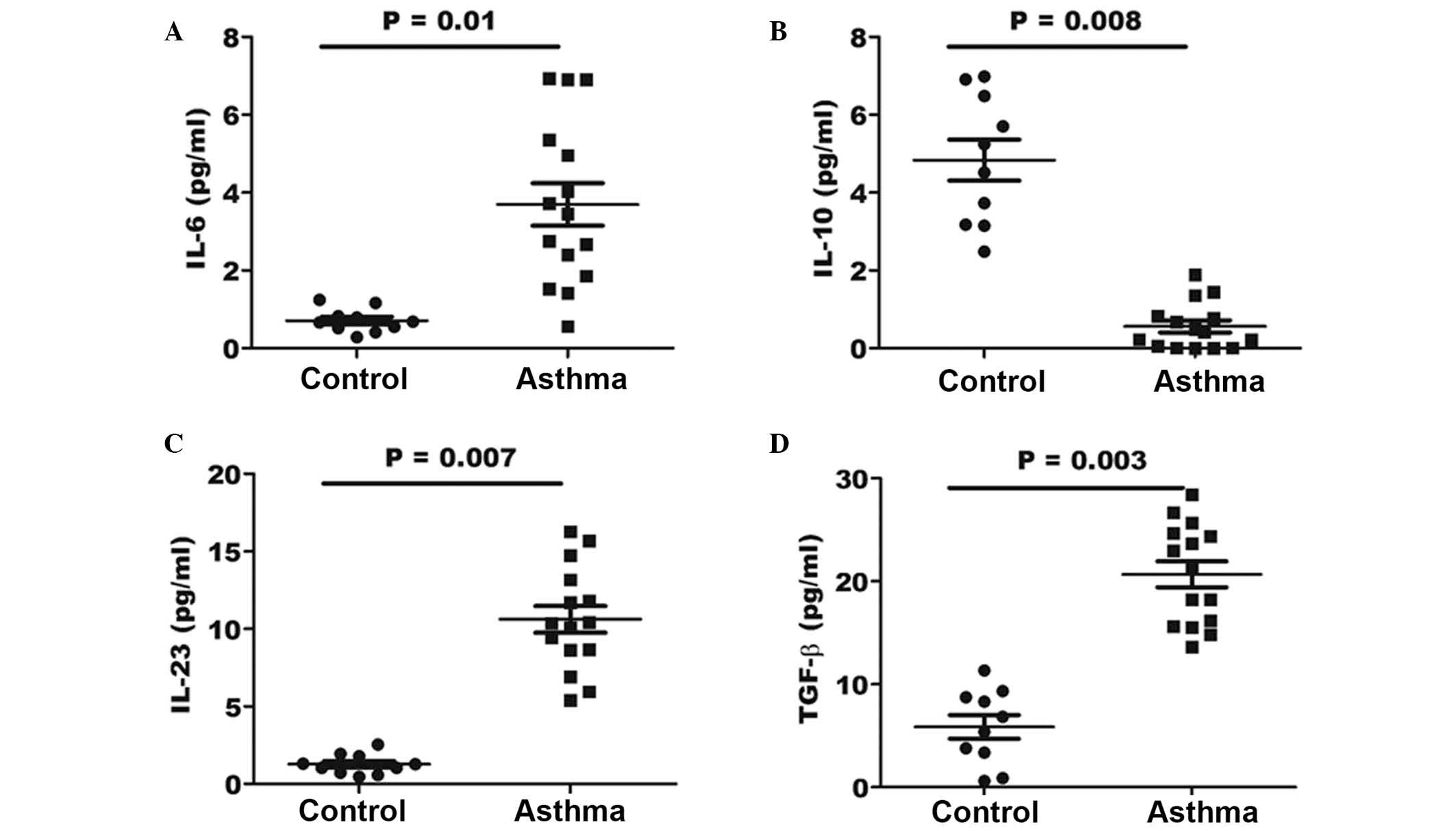
Increased levels of Th17-associated
cytokines within the BALF of asthma patients
In order to determine whether the effect of
cytokines in the lungs of asthma patients may be beneficial for the
generation of pathological Th17 cells, the expression levels of
various associated cytokines were investigated by ELISA. The
concentration of the IL-6, IL-10, IL-23 and TGF-β cytokines was
determined in the BALF specimens of asthma and control patients. As
shown in Fig. 5, the results
indicated a significantly elevated concentration of IL-6, IL-23 and
TGF-β in the BALF of asthma patients (P=0.01, 0,007 and 0.003,
respectively) when compared with that in healthy controls. However,
the expression of IL-10 in the BALF samples of asthma patients was
significantly lower when compared with that of control individuals
(P=0.008).
Discussion
The present study reported that CD4+
CD25high T cells are able to suppress T-cell responses.
Immunotherapy with peptides targeting CD4 T cell epitopes has been
under extensive investigation in animal models for the prevention
and amelioration of inflammatory responses with antigen
specificity, and is therefore currently the focus of clinical
trials in the allergy and autoimmunity areas (24). Asthma is characterized by a decline
in Treg cells along with an elevation in Th17 cells in the
peripheral blood of patients in disease remission (25). In the different pathological stages
of asthma, an increase in the expression levels of FOXP3 and
IL-17a, which are alternative markers of Treg and Th17
cells, respectively, has been observed in the peripheral blood
(26). The data reported in the
present study indicated significantly increased expression levels
of FOXP3 and IL-17a in asthma patients, suggesting
that Treg may be activated and released into the peripheral blood
in order to suppress the pro-inflammatory Th17 immune responses. In
addition, the current results identified a strong correlation
between the amount of FOXP3 transcript and the T cell
number. However, although Treg cells isolated from the peripheral
blood of asthma patients were demonstrated to be functionally
suppressive in vitro, their ability to regulate Th17
proliferation and its effector activity may be limited in
vivo due to the levels of pro-inflammatory cytokines.
For the inflammatory cytokines participating in the
maintenance the inflammatory response, prolonged exposure of Treg
cells to various cytokines (particularly IL-10 and TGF-β)may impair
their suppressive properties, as well as prevent their conversion
into Th17 cells (27). This process
may account for the reduced in Treg cell count reported in the
blood samples of asthma patients in the present study, which was
accompanied by an increase in the number of Th17 cells. The
imbalance of circulating Treg and Th17 cells in the blood of asthma
patients may account for the relapsing and remitting nature of
airway inflammation, and methods that aim to restore and maintain
this balance may provide an effective therapeutic strategy.
In mice, it has been reported that TGF-β together
with IL-6 promote the generation of Th17 cells, with TGF-β being a
key cytokine for the initiation of Th17 expression and IL-6 acting
as a crucial co-factor for Th17 differentiation; in addition, IL-23
enhances the expansion of Th17 cells (10). However, the results of the majority
of previous studies demonstrated that a range of additional
cytokines are necessary and able to induce Th17 differentiation in
humans, including combinations of IL-13, IL-6, IL-21, IL-10 and
TGF-β (28,29). The results revealed that there was no
evident difference in IL-10 cytokine expression between the control
and asthma adults; however, IL-10 is known to serve an important
role in the pathogenesis of asthma (30). An alternative pathway of Th17
differentiation involves IL-23 and TGF-β, and may be a mechanism
counteracting the resolution of inflammation that is promoted by
TGF-β in asthma. In the present study, the level of IL-6 was found
to be significantly higher in the peripheral blood of asthma
patients, compared with the controls. Notably, the expression of
IL-17a was closely correlated with IL-6 expression in asthma
patients in a previous study (31),
suggesting that these cytokines are involved in driving Th17
production in asthma patients. The protective role of TGF-β in
asthma patients may also be important, since elevated TGF-β has
been implicated in the differentiation of Treg cells, although
these patients concurrently experienced a low FOXP3 level in the
peripheral blood, suggesting that low levels of Treg cells were
present (32). Thus, TGF-β may only
have a protective effect in the absence of pro-inflammatory
cytokines and may promote Th17 cell growth in the presence of
IL-6.
In conclusion, the present study revealed that
asthma is characterized by an imbalance between Th17 effector cells
and Treg cells, with elevated numbers of Th17 cells and increased
levels of proinflammatory cytokines that promote Th17 growth. The
observed deficit in Treg cells in the asthma patients may impair
the ability of the immune system to limit excessive pathogenic
Th17-driven immune responses in the lungs. Accumulating data has
indicated that the imbalance between the counts of Treg and Th17
cells is a characteristic feature of inflammatory disorders
(33). Therefore, re-establishing
the balance by upregulating the number of Treg cells in asthma
patients and by specifically targeting Th17 cells in a disease
environment may be an effective therapeutic strategy for
asthma.
References
|
1
|
Anderson GP: Endotyping asthma: New
insights into key pathogenic mechanisms in a complex, heterogeneous
disease. Lancet. 372:1107–1119. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Tucci M, Stucci S, Strippoli S and
Silvestris F: Cytokine overproduction, T-cell activation, and
defective T-regulatory functions promote nephritis in systemic
lupus erythematosus. J Biomed Biotechnol. 2010:4571462010.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Ristich V, Liang S, Zhang W, Wu J and
Horuzsko A: Tolerization of dendritic cells by HLA-G. Eur J
Immunol. 35:1133–1142. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Hong M, Zheng J, Ding ZY, Chen JH, Yu L,
Niu Y, Hua YQ and Wang LL: Imbalance between Th17 and Treg cells
may play an important role in the development of chronic
unpredictable mild stress-induced depression in mice.
Neuroimmunomodulation. 20:39–50. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
O'Connor RA, Taams LS and Anderton SM:
Translational mini-review series on Th17 cells: CD4+ T
helper cells: Functional plasticity and differential sensitivity to
regulatory T cell-mediated regulation. Clin Exp Immunol.
159:137–147. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Miyara M, Yoshioka Y, Kitoh A, Shima T,
Wing K, Niwa A, Parizot C, Taflin C, Heike T, Valeyre D, et al:
Functional delineation and differentiation dynamics of human
CD4+ T cells expressing the FoxP3 transcription factor.
Immunity. 30:899–911. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Schreiber TH, Wolf D, Tsai MS, Chirinos J,
Deyev VV, Gonzalez L, Malek TR, Levy RB and Podack ER: Therapeutic
Treg expansion in mice by TNFRSF25 prevents allergic lung
inflammation. J Clin Invest. 120:3629–3640. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Wing K, Onishi Y, Prieto-Martin P,
Yamaguchi T, Miyara M, Fehervari Z, Nomura T and Sakaguchi S:
CTLA-4 control over FOXP3+ regulatory T cell function.
Science. 322:271–275. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Chauhan SK, El Annan J, Ecoiffier T, Goyal
S, Zhang Q, Saban DR and Dana R: Autoimmunity in dry eye is due to
resistance of Th17 to Treg suppression. J Immunol. 182:1247–1252.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Sadhu S, Khaitan BK, Joshi B, Sengupta U,
Nautiyal AK and Mitra DK: Reciprocity between regulatory T Cells
and Th17 cells: Relevance to polarized immunity in leprosy. PLoS
Negl Trop Dis. 10:e00043382016. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Maddur MS, Sharma M, Hegde P,
Lacroix-Desmazes S, Kaveri SV and Bayry J: Inhibitory effect of
IVIG on IL-17 production by Th17 cells is independent of anti-IL-17
antibodies in the immunoglobulin preparations. J Clin Immunol. 33
Suppl 1:S62–S66. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Su Z, Shotorbani SS, Jiang X, Ma R, Shen
H, Kong F and Xu H: A method of experimental rheumatoid arthritis
induction using collagen type II isolated from chicken sternal
cartilage. Mol Med Rep. 8:113–117. 2013.PubMed/NCBI
|
|
13
|
Volpe E, Servant N, Zollinger R, Bogiatzi
SI, Hupé P, Barillot E and Soumelis V: A critical function for
transforming growth factor-beta, interleukin 23 and proinflammatory
cytokines in driving and modulating human TH-17 responses. Nat
Immunol. 9:650–657. 2008. View
Article : Google Scholar : PubMed/NCBI
|
|
14
|
Bettelli E, Korn T, Oukka M and Kuchroo
VK: Induction and effector functions of T(H)17 cells. Nature.
453:1051–1057. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
McGeachy MJ, Chen Y, Tato CM, Laurence A,
Joyce-Shaikh B, Blumenschein WM, McClanahan TK, O'Shea JJ and Cua
DJ: The interleukin 23 receptor is essential for the terminal
differentiation of interleukin 17-producing effector T helper cells
in vivo. Nat Immunol. 10:314–324. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Fontao L, Brembilla NC, Masouyé I, Kaya G,
Prins C, Dupin N, Saurat JH, Chizzolini C and Piguet V:
Interleukin-17 expression in neutrophils and Th17 cells in
cutaneous T-cell lymphoma associated with neutrophilic infiltrate
of the skin. Br J Dermatol. 166:687–689. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Kurimoto E, Miyahara N, Kanehiro A, Waseda
K, Taniguchi A, Ikeda G, Koga H, Nishimori H, Tanimoto Y, Kataoka
M, et al: IL-17A is essential to the development of
elastase-induced pulmonary inflammation and emphysema in mice.
Respir Res. 14:52013. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Korn T, Bettelli E, Oukka M and Kuchroo
VK: IL-17 and Th17 cells. Ann Rev Immunol. 27:485–517. 2009.
View Article : Google Scholar
|
|
19
|
Movahedi M, Mahdaviani SA, Rezaei N,
Moradi B, Dorkhosh S and Amirzargar AA: IL-10, TGF-beta, IL-2,
IL-12, and IFN-gamma cytokine gene polymorphisms in asthma. J
Asthma. 45:790–794. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Littman DR and Rudensky AY: Th17 and
regulatory T cells in mediating and restraining inflammation. Cell.
140:845–858. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Eastaff-Leung N, Mabarrack N, Barbour A,
Cummins A and Barry S: FOXP3+ regulatory T cells, Th17
effector cells, and cytokine environment in inflammatory bowel
disease. J Clin Immunol. 30:80–89. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2-ΔΔCt method. Methods. 25:402–408. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Saurer L and Mueller C: T cell-mediated
immunoregulation in the gastrointestinal tract. Allergy.
64:505–519. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Vanderlugt CL and Miller SD: Epitope
spreading in immune-mediated diseases: Implications for
immunotherapy. Nat Rev Immunol. 2:85–95. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Wilke CM, Bishop K, Fox D and Zou W:
Deciphering the role of Th17 cells in human disease. Trends
Immunol. 32:603–611. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Lim HW, Lee J, Hillsamer P and Kim CH:
Human Th17 cells share major trafficking receptors with both
polarized effector T cells and FOXP3+ regulatory T
cells. J Immunol. 180:122–129. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Fantini MC, Rizzo A, Fina D, Caruso R,
Becker C, Neurath MF, Macdonald TT, Pallone F and Monteleone G:
IL-21 regulates experimental colitis by modulating the balance
between Treg and Th17 cells. Eur J Immunol. 37:3155–3163. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Li H, Li D, Sun J and Li Y, Yang W and Li
Y: Autoimmune regulator-overexpressing dendritic cells induce T
helper 1 and T helper 17 cells by upregulating cytokine expression.
Mol Med Rep. 13:565–571. 2016.PubMed/NCBI
|
|
29
|
Geri G, Terrier B, Rosenzwajg M, Wechsler
B, Touzot M, Seilhean D, Tran TA, Bodaghi B, Musset L, Soumelis V,
et al: Critical role of IL-21 in modulating TH17 and regulatory T
cells in Behçet disease. J Allergy Clin Immunol. 128:655–664. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Umetsu DT, McIntire JJ, Akbari O, Macaubas
C and DeKruyff RH: Asthma: An epidemic of dysregulated immunity.
Nat Immunol. 3:715–720. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Charrad R, Berraïes A, Hamdi B, Ammar J,
Hamzaoui K and Hamzaoui A: Anti-inflammatory activity of IL-37 in
asthmatic children: Correlation with inflammatory cytokines TNF-α,
IL-β, IL-6 and IL-17A. Immunobiology. 221:182–187. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Zhou L, Lopes JE, Chong MM, Ivanov II, Min
R, Victora GD, Shen Y, Du J, Rubtsov YP, Rudensky AY, et al:
TGF-beta-induced Foxp3 inhibits T(H)17 cell differentiation by
antagonizing RORgammat function. Nature. 453:236–240. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Yan C, Zhang BB, Hua H, Li B, Zhang B, Yu
Q, Li XY, Liu Y, Pan W, Liu XY, Tang RX and Zheng KY: The dynamics
of Treg/Th17 and the imbalance of Treg/Th17 in Clonorchis
sinensis-infected mice. PLoS One. 10:e01432172015. View Article : Google Scholar : PubMed/NCBI
|