Introduction
Atherosclerosis (As) is a chronic progressive
endothelial injury- and inflammation-caused vascular intimal
lesion. The plaque ruptures after shedding embolus and subsequent
thrombosis is the main cause of acute ischemic stroke of heart head
blood-vessel, and plaque rupture almost occur on the basis of
vulnerable plaques (1,2). Therefore, vulnerable plaque
stabilization is the precondition of prevention and treatment of
atherosclerosis. Vulnerable plaque refers to those which are
unstable and thrombophilia plaques (3,4). In the
partial mind plaques, with soft texture, larger lipid necrotic
core, thin fibrous cap, the collagen content and the production of
collagen fibers, smooth muscle cells (SMCs) decreased
significantly, including a large number of inflammatory cell
(macrophages, T cells and mast cells) infiltration. The fibrous cap
bears most of the stress, and leads to thrombosis in fat nuclear
separation in the blood and plaques. The thinner fibrous cap is
associated with greater surrounding pressure, and is thus easier to
break (5,6).
The integrity of the fibrous cap and fracture
resistance mainly depends on the extracellular matrix (ECM)
constantly reshaping. ECM content, thickness and strength are very
important to prevent plaque rupture. ECM synthesis and degradation
maintains a dynamic balance, and matrix metalloproteinase (MMP) and
tissue inhibitors of metalloproteinase (TIMP) play important roles
in maintaining this balance. MMP is a set of homologous enzymes
dependent on Ca+ and Zn+ neutral protease.
MMPs degrade ECM to thin fibrous caps. TIMP are polygenes encoding
the protein family, with endogenous MMP serving as a specific
inhibitory factor (7–9). The TIMP inhibition for MMP activity can
reduce the harm to the plaque. At present, we have found four
TIMPs, including TIMP-1, −2, −3 and −4 (7). TIMP1 mainly relates to atherosclerosis
(8,9). Transforming growth factor-β (TGF-β) is
a protein peptide with a variety of functions, widely in
organization, TGF-β1 had highest proportion (>90%) and the
strongest activity in cells, and is secreted in the blood vessel
walls (such as SMCs, endothelial cells and macrophages) of cells.
TGF-β can promote ECM synthesis and secretion, adjust MMP content
and activity, and stimulate TIMP synthesis to play a key role in
keeping the balance between atheromatous plaque inflammation and
fibrosis. TGF-β is an important factor for originating protection
of atherosclerosis, as well as maintain the plaque stability
(10,11). As a result, we assumed ‘protective
factor’ TGF-β1 and restraining factor TIMP1 to be imported in
vulnerable plaque inflammatory cells, to stabilize ECM to achieve
stable plaques.
Ultrasound-targeted microbubble destruction (UTMD)
is a rapid development non-viral vector-mediated gene transfection
technique utilized because of its safety and ultrasonic irradiation
target tissue specificity. UTMD has been successfully applied in a
variety of tissue and organ experimental research (12,13). In
the present study, we combined the UTMD and recombinant
adeno-associated virus (rAAV), to achieve a high safety and
transfection, and gene expression (14). In animal models, when the target
therapy of TGF-β2, combined with RNAi intervention technology, and
hyperplasia of fundus change in PVR models were significantly
suppressed, the treatment effect was significantly improved
(15).
At present, UTMD is mainly directed towards
myocardial ischemia (13).
Therefore, in the present study, on the basis of previous research,
using IRES sequence, we determined whether ultrasound-targeted
microbubble destruction (UTMD) combined with dual targeting of
transforming growth factor (TGF)-β1 and tissue inhibitors of
metalloproteinase (TIMP) 1 recombinant adeno-associated virus
(rAAV) can stabilize atherosclerotic vulnerable plaques. The
results showed that TGF-β1 and TIMP1 are crucial in the regulation
of atherosclerotic plaques.
Materials and methods
Animals
To establish the carotid atherosclerotic vulnerable
plaque rabbit model, 18 New Zealand healthy white male rabbits,
weight 2.3–2.9 kg, 4–5 months of age were provided by the Naval
Medical Research Institute. Starting from week 1, experimental
rabbits were given high fat (1% cholesterol) feed for up to 10
weeks. After 2 weeks, the experimental rabbits were anesthetized
with 3% pentobarbital 30 mg/ml/kg after ear marginal vein
anesthesia. A proximal left thigh line was inserted into the left
femoral artery to separate the skin and subcutaneous tissue, and
femoral of 2 cm was isolated. Ligation telecentric with surgical
suture near the heart filled a femoral artery to block blood flow,
and a ‘V’ type small mouth artery wall was cut. In retrograde
manner, a balloon catheter (diluted with 1:15 heparin saline
infiltration) was inserted to thoracic aorta (about 20 cm) using
thread, and a 20-ml syringe, a 10-ml air injection was used to fill
the balloon, after which the syringe was gradually pulled back
until the common iliac artery (about 10-cm scale) allowing for
insertion of the catheter. The syringe was pulled once again
ensuring the intimal injury, prior to removing the catheter.
Ligation followed and skin and subcutaneous tissue were obtained
prior to washing the wound with penicillin sodium. Four weeks after
confirmation of the model, the model group was injected with
saline, the UTMD vector group was injected with UTMD vector and the
UTMD rAAV group was injected with UTMD targeted rAAV. The injection
was given once per week, for up to 10 weeks.
The animal experiments were approved by the Ethics
Committee of Shanghai General Hospital Affiliated to Nanjing
Medical University.
Hematoxylin and eosin (H&E)
staining
The right common carotid artery was selected in the
rabbits midway through embedding 1 cm. Each group of blood vessels
was placed in the same wax block (3 replicates of thickness 3–4 µm
at 10 µm intervals). After H&E staining, the tissues were
observed at ×10 magnification.
THP-1 macrophage cell induction
THP-1 cells were purchased from ATCC. Mononuclear
THP-1 cells were cultured in RPMI-1640 medium containing 10%
heat-inactivated fetal bovine serum, 10 µ/ml penicillin and 10
µg/ml of streptomycin, and incubated at 37°C and 5% CO2
prior to maintaining the cell number at 1×108/flask. The
subculture THP-1 cells were diluted at 1×106/ml,
vaccinated in 35-mm Petri dishes containing 100 ng/ml phorbol
esters (PMA), 0.3% BSA serum-free RPMI-1640 medium, and stimulated
for 48 h to induce differentiation. We used light microscope
(Olympus, Tokyo, Japan) to observe the morphology, and for the
identification of differentiation to macrophages.
RT-PCR mRNA expression
Total RNA was extracted using Total RNA kit (Toyobo,
Osaka, Japan). SYBR-Green RT-PCR was performed and GAPDH mRNA was
used to normalize the RNA inputs. The primers were produced by
Shanghai Sangon Biological (Shanghai, China). RT-PCR was used to
detect the target gene expression TGF-β1 and TIMP1. Total RNA was
extracted for RNA reverse transcription cDNA.
Immunofluorescence
After transfection for 24 h, the cells in each group
washed three times and fixed in 4% paraformaldehyde for 15 min in
PBS, permeabilized with 0.1% Triton X-100, and blocked with 1% BSA
in PBS for 1 h. Cell nuclei were dyed with DAPI (Vector
Laboratories, Burlingame, CA, USA). After washing, the cells were
visualized using a confocal laser microscope system (Nikon, Tokyo,
Japan).
Transfection efficiency and cell
apoptosis detection
Each cell group was transfected for 48 h and then
digested with serum free medium suspension cells. According to
Annexin V-FITC apoptosis kit manual introduction, flow cytometry
was used to test transfection and cell apoptosis.
Western blotting
Cells were collected in a lysis buffer, the protein
quantification was determined by BCA protein kit (Thermo Fisher
Scientific, Waltham, MA, USA) and equal amounts of proteins were
separated by 12% SDS-PAGE as described elsewhere (16). GAPDH was chosen as an internal
control.
Statistical analysis
Data were presented as the mean ± standard
deviation. Statistical analysis was carried out using Prism 5.0
(Prism; GraphPad Software, Inc., La Jolla, CA, USA). One-way
analysis of variance and post hoc comparisons were used for the
statistical evaluation. P <0.05 was considered to indicate a
statistically significant difference.
Results
Protein expression with H&E
staining and western blotting
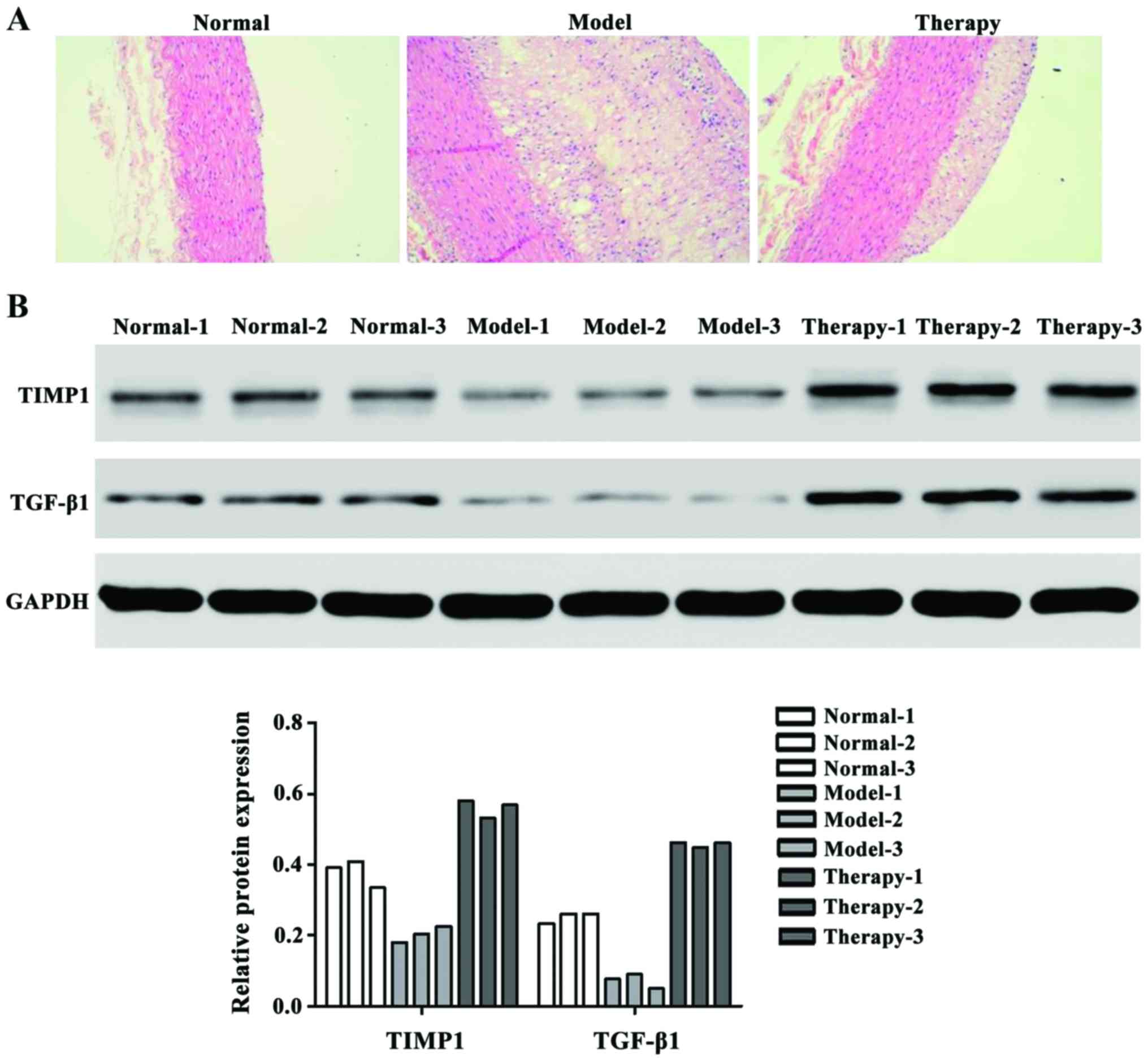
H&E staining results showed that after treating
UTMD with TGF-β1 and TIMP1 rAAV, the therapy group showed
significantly stabilized atherosclerosis vulnerable plaques
(Fig. 1A). We also examined the
protein expression level of TGF-β1 and TIMP1 factors that have
higher expression in therapy group than the model group (Fig. 1B).
Mononuclear cell morphological
observation
THP-1 cells were observed under an inverted
microscope had equal size small round cells, surface was smooth
with adherent growth (Fig. 2A). PMA
was used for 48 h induction to produce macrophages, and found that
some cells formed circular to polygon shape under the inverted
microscope. Additionally, the cells showed stick walls, strong
adhesion, the volumes increased and a few revealed long fusiform
shapes (Fig. 2B).
Transfection THP-1 mRNA and protein
expression level
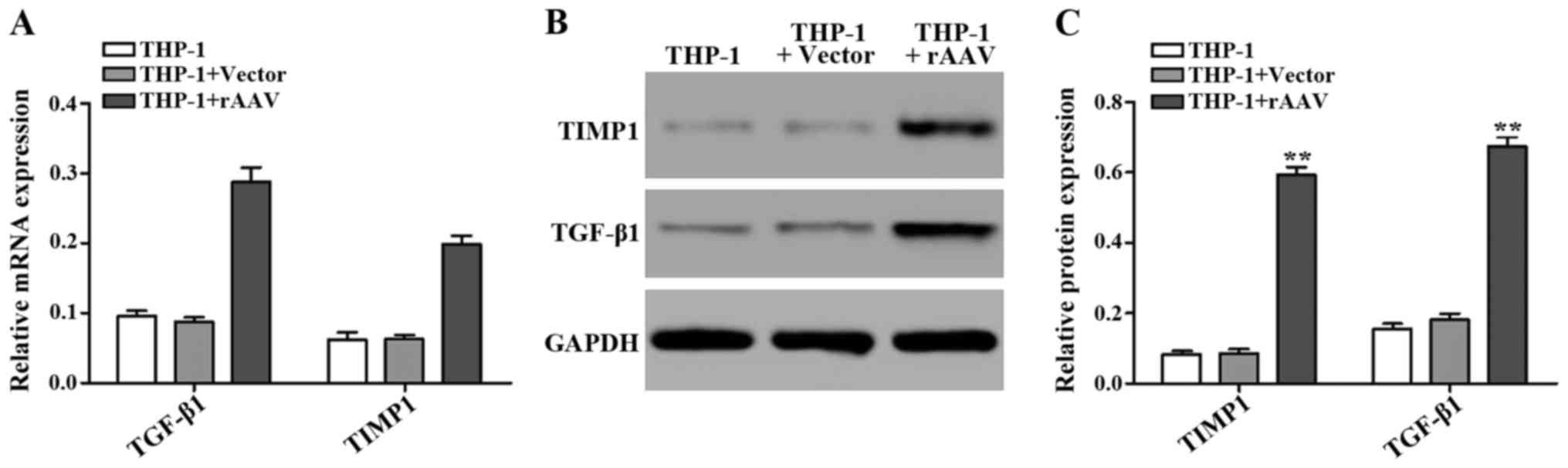
After using UTMD-transfected genes, we tested the
TGF-β1 and TIMP1 mRNA and protein expression level. The GAPDH was
used as an internal control to quantify mRNA expression (Fig. 3).
Apoptosis result
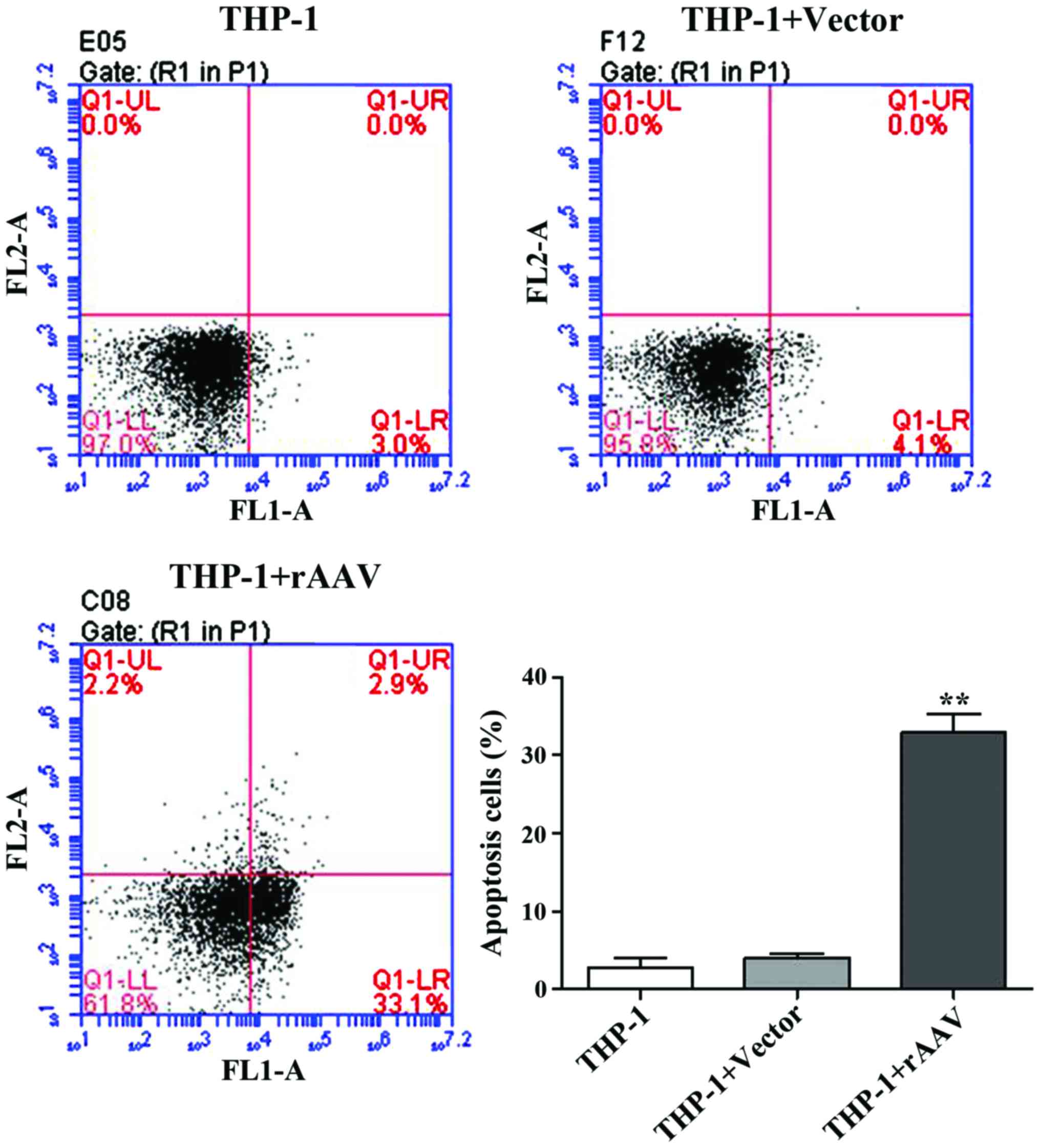
After UTMD treatment, flow cytometry was performed
to examine the cells apoptosis. The results showed that after
transfection with TGF-β1 and TIMP1 rAAV, the apoptotic rates were
significantly induced from 4.1 to 33.1% in the rAAV group (Fig. 4).
Western blot results
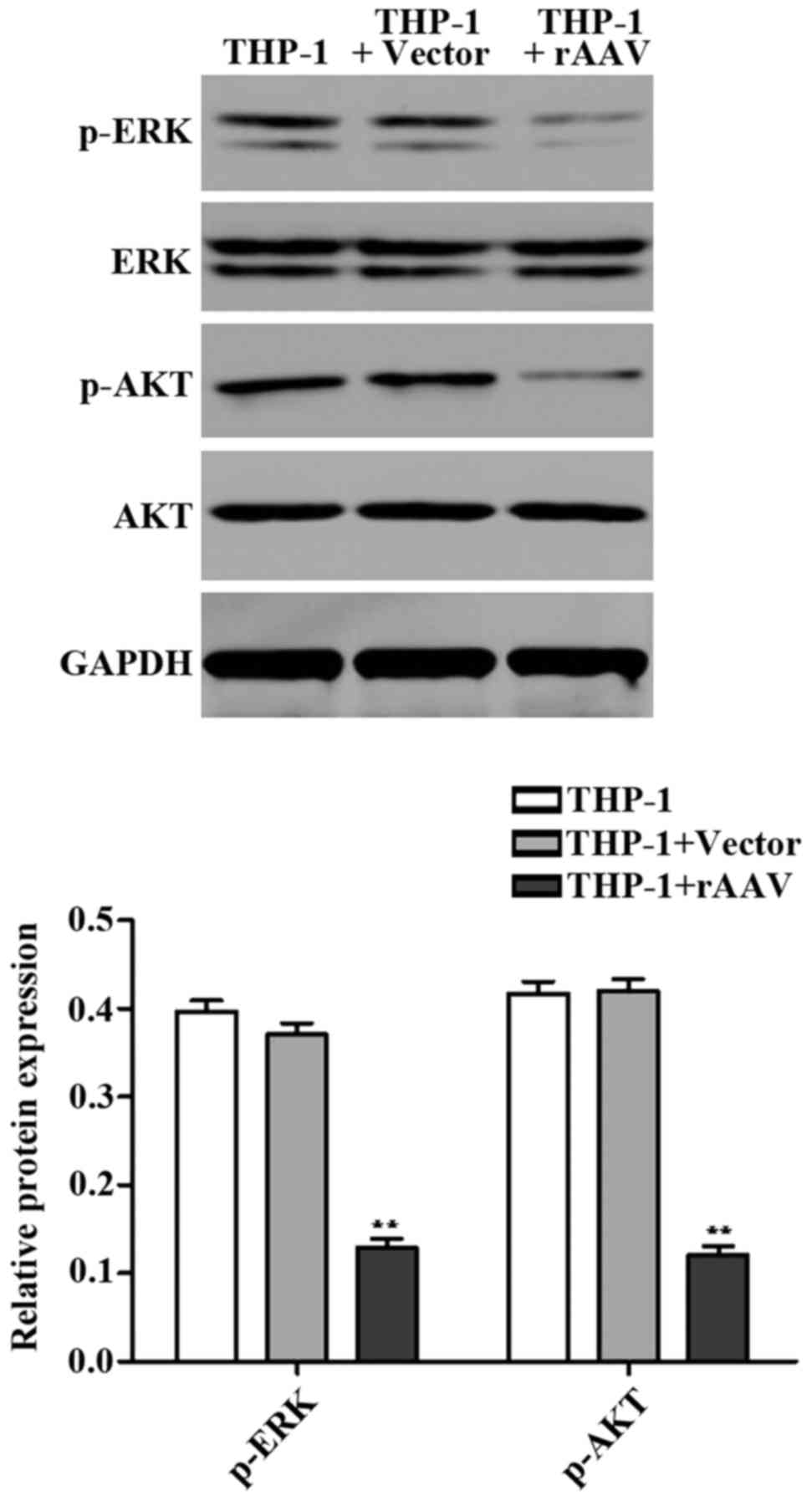
We detected expression of apoptosisrelated proteins
ERK and AKT using western blotting. It was found that the protein
levels of p-ERK and p-AKT in rAAV group were significantly
downregulated compared to the control group (Fig. 5). These data indicated that TGF-β1
and TIMP1 rAAV induced cell apoptosis by regulating ERK and AKT
signaling proteins.
Discussion
This study tested the hypothesis that a specific
TGF-β1 and TIMP1 influence the development of atherosclerotic
plaques. rAAV have some reported biological features such as
non-pathogenic nature, broad host range, ability to infect dividing
and non-dividing cells and long-term expression of foreign genes
(17). UTMD has been shown to be
able to enhance adenoviral transfection efficiency (18). In this study, we used UTMD enhanced
rAAV-mediated gene TGF-β1 and TIMP transfer into models and
cells.
The rabbit model provided direct evidence that the
TGF-β1 and TIMP1 protein levels were significantly upregulated in
the therapy group. Furthermore, H&E staining showed that the
athrosclerosis vulnerable plaques were stabilized in the therapy
group compared to the model group. TGF-β1 is an important cytokine
in the atherogenic process preventing atherosclerotic plaque
formation (19). TIMP1 is
synthesized by most types of connective tissue cells as well as
macrophages. The overexpression of TIMP1 has been reported to
reduce atherosclerotic lesion in ApoE−/− mice (20).
We also demonstrated the UTMD delivered TGF-β1 and
TIMP1 genes by rAAV into THP-1 macrophages. Apoptosis plays an
essential role in different pathological processes including
atherosclerosis, in which it affects all cell types in the
atherosclerotic lesion (21). The
apoptosis rates were significantly induced after UTMD treatment
compared to the control group. ERK is involved in cell death
(22). AKT plays a role in the
development of resistance to apoptosis (23). We showed that rAAV group has
downregulated p-ERK and p-AKT apoptosis-related protein levels
compared to the control group. These studies provided evidence for
the important roles of TGF-β1 and TIMP1 in athrosclerosis
plaques.
As TGF-β1 and TIMP1 have been demonstrated to be
involved in atherosclerotic plaques, we concluded that TGF-β1 and
TIMP1 may therefore beneficially modulate the plaque stability.
References
|
1
|
Anogeianaki A, Angelucci D, Cianchetti E,
D'Alessandro M, Maccauro G, Saggini A, Salini V, Caraffa A, Tete S,
Conti F, et al: Atherosclerosis: a classic inflammatory disease.
Int J Immunopathol Pharmacol. 24:817–825. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Hansson GK: Atherosclerosis - an immune
disease: the Anitschkov Lecture 2007. Atherosclerosis. 202:2–10.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Muller JE, Abela GS, Nesto RW and Tofler
GH: Triggers, acute risk factors and vulnerable plaques: the
lexicon of a new frontier. J Am Coll Cardiol. 23:809–813. 1994.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Fishbein MC: The vulnerable and unstable
atherosclerotic plaque. Cardiovasc Pathol. 19:6–11. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Virmani R, Burke AP, Farb A and Kolodgie
FD: Pathology of the unstable plaque. Prog Cardiovasc Dis.
44:349–356. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Bartels ED, Bang CA and Nielsen LB: Early
atherosclerosis and vascular inflammation in mice with diet-induced
type 2 diabetes. Eur J Clin Invest. 39:190–199. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Massova I, Kotra LP, Fridman R and
Mobashery S: Matrix metalloproteinases: structures, evolution, and
diversification. FASEB J. 12:1075–1095. 1998.PubMed/NCBI
|
|
8
|
Orbe J, Fernandez L, Rodríguez JA, Rábago
G, Belzunce M, Monasterio A, Roncal C and Páramo JA: Different
expression of MMPs/TIMP-1 in human atherosclerotic lesions.
Relation to plaque features and vascular bed. Atherosclerosis.
170:269–276. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Fiotti N, Altamura N, Orlando C, Simi L,
Reimers B, Pascotto P, Zingone B, Pascotto A, Serio M, Guarnieri G,
et al: Metalloproteinases-2, −9 and TIMP-1 expression in stable and
unstable coronary plaques undergoing PCI. Int J Cardiol.
127:350–357. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Bot PT, Grundmann S, Goumans M-J, de
Kleijn D, Moll F, de Boer O, van der Wal AC, van Soest A, de Vries
JP, van Royen N, et al: Forkhead box protein P1 as a downstream
target of transforming growth factor-β induces collagen synthesis
and correlates with a more stable plaque phenotype.
Atherosclerosis. 218:33–43. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Deng H-B, Jiang CQ, Tomlinson B, Liu B,
Lin JM, Wong KS, Cheung BM, Lam TH and Thomas GN: A polymorphism in
transforming growth factor-β1 is associated with carotid plaques
and increased carotid intima-media thickness in older Chinese men:
the Guangzhou Biobank Cohort Study-Cardiovascular Disease
Subcohort. Atherosclerosis. 214:391–396. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Walton CB, Anderson CD, Boulay R and
Shohet RV: Introduction to the ultrasound targeted microbubble
destruction technique. J Vis Exp. 12:29632011.
|
|
13
|
Fujii H, Li S-H, Wu J, Miyagi Y, Yau TM,
Rakowski H, Egashira K, Guo J, Weisel RD and Li R-K: Repeated and
targeted transfer of angiogenic plasmids into the infarcted rat
heart via ultrasound targeted microbubble destruction enhances
cardiac repair. Eur Heart J. 32:2075–2084. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Li HL, Zheng XZ, Wang HP, Li F, Wu Y and
Du LF: Ultrasound-targeted microbubble destruction enhances
AAV-mediated gene transfection in human RPE cells in vitro and rat
retina in vivo. Gene Ther. 16:1146–1153. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Zheng X, Du L, Wang H and Gu Q: A novel
approach to attenuate proliferative vitreoretinopathy using
ultrasound-targeted microbubble destruction and recombinant
adeno-associated virus-mediated RNA interference targeting
transforming growth factor-β2 and platelet-derived growth factor-B.
J Gene Med. 14:339–347. 2012. View
Article : Google Scholar : PubMed/NCBI
|
|
16
|
Padda RS, Gkouvatsos K, Guido M, Mui J,
Vali H and Pantopoulos K: A high-fat diet modulates iron metabolism
but does not promote liver fibrosis in hemochromatotic
Hjv−/− mice. Am J Physiol Gastrointest Liver Physiol.
308:G251–G261. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Daya S and Berns KI: Gene therapy using
adeno-associated virus vectors. Clin Microbiol Rev. 21:583–593.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Howard CM, Forsberg F, Minimo C, Liu JB,
Merton DA and Claudio PP: Ultrasound guided site specific gene
delivery system using adenoviral vectors and commercial ultrasound
contrast agents. J Cell Physiol. 209:413–421. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Mallat Z, Gojova A, Marchiol-Fournigault
C, Esposito B, Kamaté C, Merval R, Fradelizi D and Tedgui A:
Inhibition of transforming growth factor-β signaling accelerates
atherosclerosis and induces an unstable plaque phenotype in mice.
Circ Res. 89:930–934. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Rouis M, Adamy C, Duverger N, Lesnik P,
Horellou P, Moreau M, Emmanuel F, Caillaud JM, Laplaud PM, Dachet
C, et al: Adenovirus-mediated overexpression of tissue inhibitor of
metalloproteinase-1 reduces atherosclerotic lesions in
apolipoprotein E-deficient mice. Circulation. 100:533–540. 1999.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Guevara NV, Chen K-H and Chan L: Apoptosis
in atherosclerosis: pathological and pharmacological implications.
Pharmacol Res. 44:59–71. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Cagnol S and Chambard JC: ERK and cell
death: Mechanisms of ERK-induced cell death - apoptosis, autophagy
and senescence. FEBS J. 277:2–21. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Khwaja A: Akt is more than just a Bad
kinase. Nature. 401:33–34. 1999. View
Article : Google Scholar : PubMed/NCBI
|