Introduction
Fucoidan is a sulfated polysaccharide extracted from
marine brown algae and some echinoderms that has various beneficial
health effects (1–3). There is a growing interest among
producers and consumers in using new functional ingredients, such
as fucoidan, in the diet (1).
Fucoidan has been extensively studied due to it having multiple
biological activities and potential applications, including
anticancer, antiviral, anti-metastasis, anti-lymphangiogenesis,
immune modulation, blood anticoagulant, protection from radiation
damage, tissue engineering and pathogen inhibition effects
(2–6).
Fucoidan may also have beneficial effects on kidney
function. Previous studies have reported that fucoidan may reduce
the metabolic abnormalities of diabetic rats and delay the
progression of diabetic renal complications in a rat model of
streptozoin-induced diabetes (7).
Fucoidan has also been demonstrated to have a renoprotective role
in oxalate-mediated peroxidative injury (8). Furthermore, previous studies have
reported that fucoidan may alleviate the symptoms of chronic kidney
disease in a model of subtotal nephrectomy chronic kidney disease
and have a renoprotective effect on active Heymann nephritis
(9,10).
Although fucoidan has a protective effect against
uric acid nephropathy (UAN) (11),
little is known about the underlying molecular mechanism of this.
In the present study, a rat model of UAN was established using
adenine in order to further investigate the renoprotective role of
fucoidan and its molecular mechanism. Using hematoxylin and eosin
(H&E) and periodic acid-methenamine silver (PAM)-Masson
staining, it was indicated that fucoidan was able to alleviate
symptoms of UAN in a rat model.
Organic cation transporter 2 (OCT2) is expressed
predominantly in the kidney, at the basolateral membrane of S2 and
S3 segments of the proximal tubule (12,13).
OCT2 is a major transporter of various organic cations from the
blood stream into renal epithelial cells and is important for the
cellular uptake of cationic drugs across the basolateral membrane
of proximal tubule cells, including metformin, lamivudine and
cimetidine (14). OCT2 is regulated
by protein kinase A (PKA), phosphoionositide 3-kinase, p56lck
tyrosine kinase and mitogen-activated protein kinase kinase 1 and 2
(15). To gain insight into the
possible roles of PKA and OCT2 in the protective effect of
fucoidan, PKA 2β, phosphorylated (p)-PKA 2β, surface OCT2 and total
OCT2 protein levels were evaluated in renal tissues of a rat model
of UAN in the present study. The results indicated that PKA 2β and
p-PKA 2β levels were significantly elevated in renal tissues of the
fucoidan treatment group compared with the UAN model group,
suggesting that PKA was activated in the kidney tissue of fucoidan
treatment group rats. Notably, the surface OCT2 level was markedly
increased by fucoidan treatment with no change in total OCT2
level.
To further ascertain the role of fucoidan in
regulating PKA and surface OCT2 expression, COS-7 cells ectopically
expressing OCT2 were established. The level of PKA 2β and p-PKA 2β
and surface OCT2 was significantly increased by treatment with
fucoidan compared with the control in OCT2-expressing COS-7 cells.
These observations indicated that fucoidan may activate PKA and
upregulate surface OCT2 in OCT2-expressing COS-7 cells.
Furthermore, it was confirmed that activation of PKA with
8-bromoadenosine 3′,5′-cyclic monophosphate (8-bromo-cAMP)
upregulated surface OCT2 in OCT2-expressing COS-7 cells. In
summary, fucoidan upregulated surface OCT2 via upregulating the
expression of PKA in order to alleviate the symptoms of UAN.
Materials and methods
Rat model of UAN and experimental
procedures
A total of 30 male Sprague Dawley rats (weight,
167.2±1.9 g; age, 6–8 weeks), were randomly assigned into three
experimental groups. The rats were housed in temperature-(22–25°;C)
and humidity-(40–70%) controlled conditions with a 12-h light/dark
cycle and ad libitum access to food and water. All rats were
treated by intragastric administration. Group 2 and 3 rats were
administered with 30 mg/kg adenine once a day for 18 days to
establish a rat model of UAN. Group 1 was allocated as the normal
control group and rats werefed an equivalent amount of
carboxymethylcellulose sodium (CMC-Na; Sigma-Aldrich; Merck KGaA,
Darmstadt, Germany). Rats in Group 2 were then fed with an
equivalent amount of saline buffer (30 mg/kg body weight) for 18
days to establish the disease model. Rats in Group 3 were fed with
a single dose of fucoidan polysaccharide sulfate (400 mg/kg body
weight) to establish the fucoidan treatment group.
Rats were obtained from the Experimental Animal
Center of Sun Yat-sen University (Guangzhou, China). The research
was carried out in accordance with the Guidelines for Human
Treatment of Animals set by the Association of Laboratory Animal
Sciences and the Center for Laboratory Animal Sciences at Sun
Yat-sen University. The study was approved by the Committee of
Biomedical Ethics of Sun Yat-sen University [SYXK (yue):
2007-0081].
Chemicals and reagents
Fucoidan polysaccharide sulfate was diluted with 10%
dimethyl sulfoxide in ethanol to a final concentration of 50 mg/ml.
Fucoidan was purchased from South Product Co., Ltd. (Uruma, Japan).
Adenine tablets were purchased from Amresco, LLC (Solon, OH, USA)
and diluted with 0.15% CMC-Nato a final concentration of 3%. Cell
Surface Protein Isolation kit (K295) was purchased from BioVision,
Inc., (Milpitas, CA, USA). Sulfo-NHS-SS-biotin and Avidin Agarose
beads were purchased from Thermo Fisher Scientific, Inc. (Waltham,
MA, USA). COS-7 cells were obtained from the Cell Bank of the
Chinese Academy of Sciences (Shanghai, China) and were cultured in
Dulbecco's modified Eagle's medium supplemented with 10% fetal
bovine serum (both Gibco; Thermo Fisher Scientific, Inc.) and 100
U/ml penicillin and 100 U/ml streptomycin (Amresco, LLC). The cells
were cultured in a 5% CO2 atmosphere at 37°C.
8-bromo-cAMP (Sigma-Aldrich; Merck KGaA) was used asaPKA activator.
OCT2-expressing COS-7 cells were treated with PKA activator
8-bromo-cAMP (1 µmol/l) or phosphate-buffered saline (PBS) for 12 h
at 37°C.
Tissue surface protein extraction/cell
surface biotinylation and western blot analysis
After treatment for 18 days, rats were euthanized by
CO2 inhalation, and the renal tissues from rats in
different groups were frozen and stored at −20°C immediately after
dissection. Renal tissues were mechanically dissociated and
homogenized according to the manufacturer's instructions for the
surface protein extraction kit. For COS-7 cell surface
biotinylation, after treatment with fucoidan (500 µg/ml) or control
salinefor 24 h, cells were washed and incubated with PBS
supplemented with 0.5 mg/ml sulfo-NHS-SS biotin for 40 min at 4°C,
and excess biotin was quenched with 50 mM Tris-PBS buffer for 20
min at 4°C. Subsequently, cells were collected, lysed in
radioimmunoprecipitation buffer from BioVision, Inc., and subjected
to streptavidin-agarose beads at 4°C for a further 3 h. The protein
samples (30 µg) obtained were boiled, subjected to 12% SDS-PAGE and
transferred to polyvinylidene difluoride membranes (EMD Millipore,
Billerica, MA, USA). The membranes were blocked with Tris-buffered
saline-Tween-20 containing 2% bovine serum albumin (Sigma-Aldrich;
Merck KGaA) at room temperature for 2 h, and the membranes were
then incubated with specific antibodies against OCT2 (sc-233; Santa
Cruz Biotechnology, Inc., Dallas, TX, USA; 1:1,500), PKA 2β
(sc-365615; Santa Cruz Biotechnology, Inc; 1:1,500), p-PKA 2β
(sc-293036; Santa Cruz Biotechnology, Inc.; 1:1,000), GAPDH
(ab8245; Abcam, Cambridge, UK; 1:8,000). Horseradish
peroxidase-conjugated goat anti-rabbit secondary antibodies (20162;
ProMab Biotechnologies, Inc., Richmond, CA, USA; 1:1,000) were
used. For incubation, all antibodies were incubated at 4°C for 12
h. The membranes were washed inTBST (Sigma-Aldrich; Merck KGaA) at
room temperature four times between the incubations with the
primary and secondary antibodies. Protein bands were detected using
an enhanced chemiluminescence reaction (Biological Industries,
Kibbutz Beit Haemek, Israel). The intensity of each band was
quantified using Quantity One software v4.62 (Bio-Rad Laboratories,
Inc., Hercules, CA, USA). All experiments were repeated three
times.
H&E staining of renal tissues
Renal tissues of rats were removed, fixed with 4%
paraformaldehyde in PBS at room temperature for 2 h, decalcified
for 10 days at room temperature with EDTA and embedded in paraffin
for histological analysis. Kidney sections (5 µm thick) were
stained with hematoxylin for 2 min, then eosin for 30 sec. Images
of the kidney from multiple sections in each rat were observed
under a light microscope with identical parameters (magnification,
resolution and light intensity).
PAM-Masson staining
In brief, rat kidneys were harvested, cut along the
short axis at the maximum area of the whole kidney, fixed in
Carnoy's solution (Beyotime Institute of Biotechnology, Haimen,
China) at 37°C for 2 h, embedded in paraffin, sectioned (1 µm
thick), and stained with PAM. Images of the kidney from multiple
sections in each rat were observed under a light microscope with
identical parameters (magnification, resolution and light
intensity).
Immunohistochemistry assay for
PKA
Immunohistochemistry was performed by National
Engineering Center for Biochip at Shanghai (China), as described
previously (16,17). Immunohistochemistry was carried out
with antibodies specific for PKA using anti-rat PKA (sc-365615;
Santa Cruz Biotechnology, Inc.; 1:300) in blocking buffer and
incubated overnight at 4°C. Negative control sections were
incubated with PBS instead of primary antibodies.
OCT2-expressing plasmid construction
and transfection
Green fluorescent protein (GFP)-OCT2 and GFP-C
(control) vector plasmids were purchased from OriGene Technologies,
Inc., (Rockville, MD, USA) and transfected into COS-7 cells using a
Lipofectamine® 2000 (Invitrogen; Thermo Fisher
Scientific, Inc.) reagent, according to the manufacturer's
protocol.
Terminal
deoxynucleotidyl-transferase-mediated dUTP nick end labelling
(TUNEL) assay
Frozen renal tissue sections (3 µm thick) were cut
and mounted on 3-aminopropyltriethoxysilane-treated glass slides.
The TUNEL protocol was performed using the In Situ Cell
Death Detection kit (Sigma-Aldrich; Merck KGaA), according to the
manufacturer's instructions. Sections were dried overnight, fixed
in 1% formal dehydeat room temperature for 2 h, washed with PBS
four times for 5 min each, permeabilized with 0.2% Triton X-100 at
4°C for 10 min, and re-washed with PBS before application of TUNEL
reagents. Images were observed under a light microscope with
identical parameters (magnification, resolution and light
intensity).
Statistical analysis
All data were presented as the mean ± standard error
of the mean of at least three independent experiments. Student's
t-test was used to evaluate differences between two groups with
GraphPad Prism v5.01 (GraphPad Software, Inc., La Jolla, CA, USA).
P<0.05 was considered to indicate a statistically significant
result.
Results
Fucoidan alleviates the symptoms of
UAN in a rat model of UAN
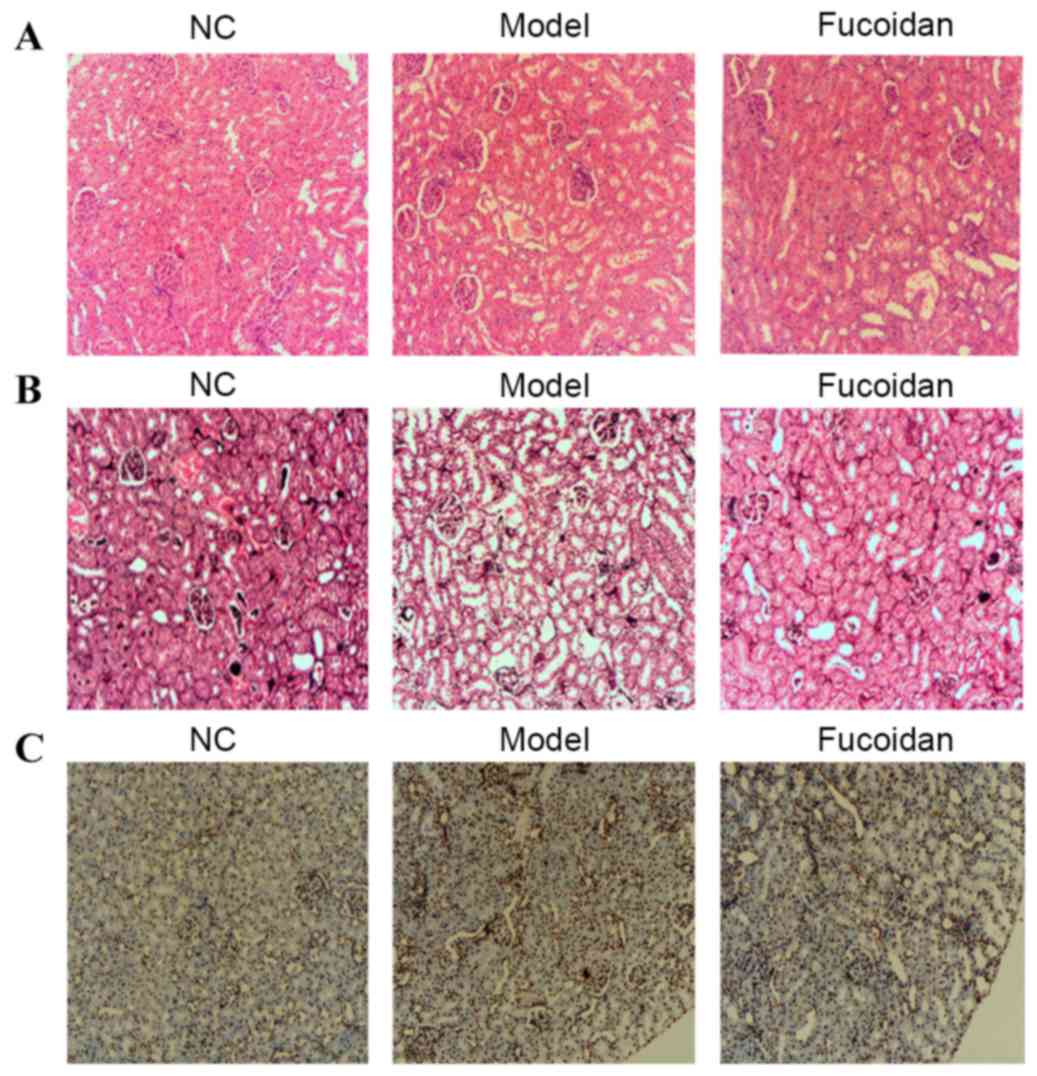
An experimental model of UAN was induced using
uricopoiesis promoter adenine. Histopathological analysis of renal
tissues in each group was performed. H&E staining of kidney
tissue indicated that the control group maintained normal
glomerular size, integrity and demonstrated no notable inflammatory
response (Fig. 1A). In the UAN model
group, cellular infiltrate in the tubules, atrophic glomeruli,
tubular ectasia, granuloma hyperplasia focal fibrosis and
accumulated urate crystals were observed. However, treatment with
fucoidan decreased the deposition of urate crystals and
inflammatory cellular influx in the tubule compared with the model
group. Furthermore, tubular ectasia and interstitial fibrosis were
improved by fucoidan treatment, suggesting that kidney damage was
attenuated by fucoidan. The renal tissues were also processed and
stained with PAM-Masson for examination under a light microscope.
The PAM-Masson results revealed that renal interstitial fibrosis
was less prominent among renal tissues from the fucoidan treatment
group compared with the model group (Fig. 1B). In addition, the fucoidan
treatment group demonstrated a lower proportion of apoptotic nuclei
in the kidney compared with the model group, as assessed by TUNEL
staining (Fig. 1C). These results
suggested that fucoidan was able to alleviate the symptoms of UAN
in a rat model.
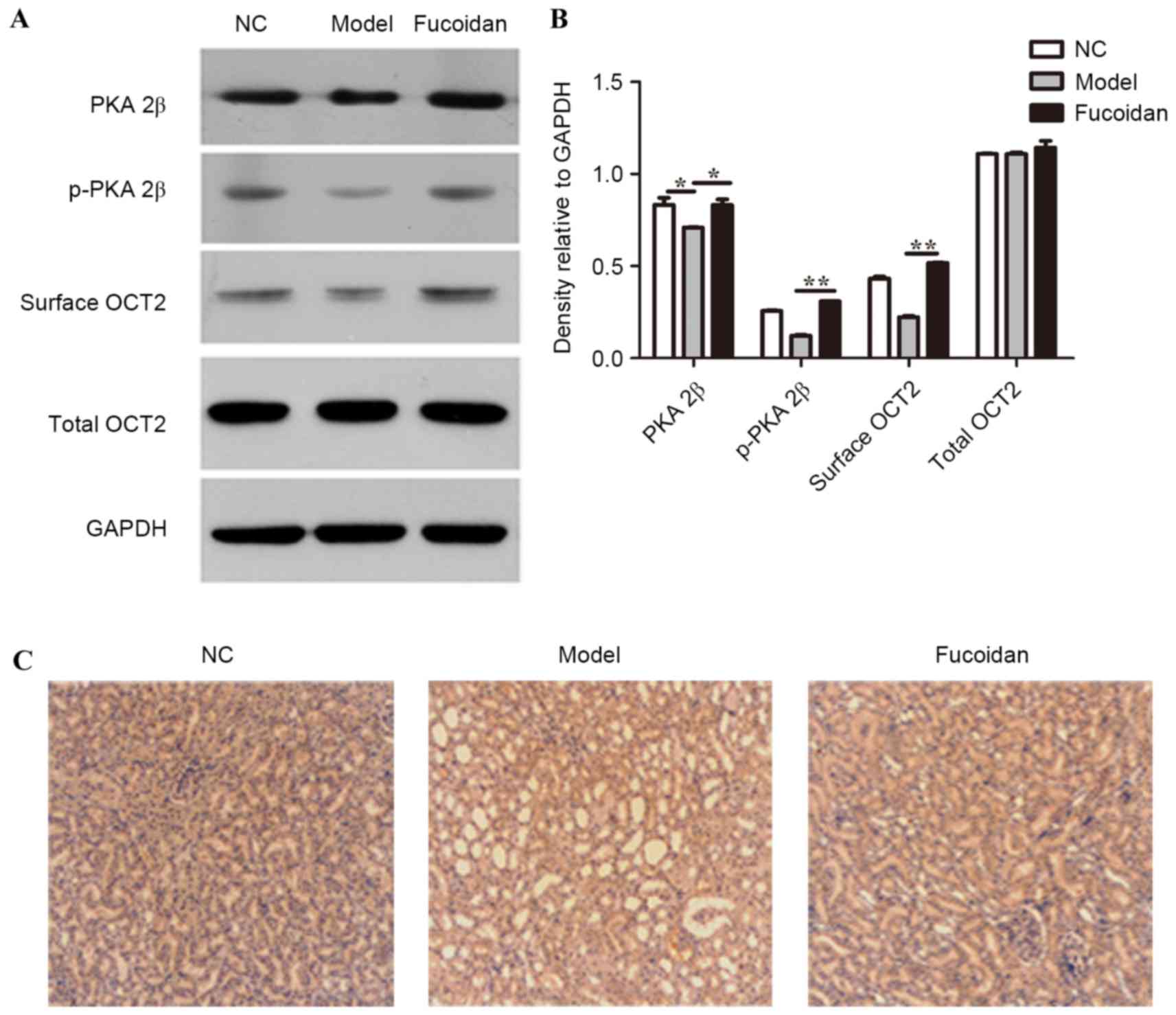
Fucoidan activates PKA and upregulates
surface OCT2 in renal tissues of a rat model of UAN
Given that fucoidan alleviated the symptoms of UAN,
further study to investigate the specific molecular mechanism was
required. Thus, the protein levels of PKA 2β, p-PKA 2β, surface
OCT2 and total OCT2 were evaluated in renal tissues of the
different groups. As demonstrated in Fig. 2A and B, PKA 2β and p-PKA 2β protein
expression levels were significantly elevated in the renal tissues
of the fucoidan treatment group compared with the model group
(P<0.05 and P<0.01, respectively), suggesting that increased
expression of PKA occurred in kidney tissue of fucoidan treatment
group rats. The upregulation of PKA protein expression was further
demonstrated by immunohistochemistry staining of PKA in rat renal
tissues (Fig. 2C). The level of
surface OCT2 protein was significantly increased by fucoidan
treatment compared with the model group (P<0.01), with no
significant change in total OCT2 level (Fig. 2A and B).
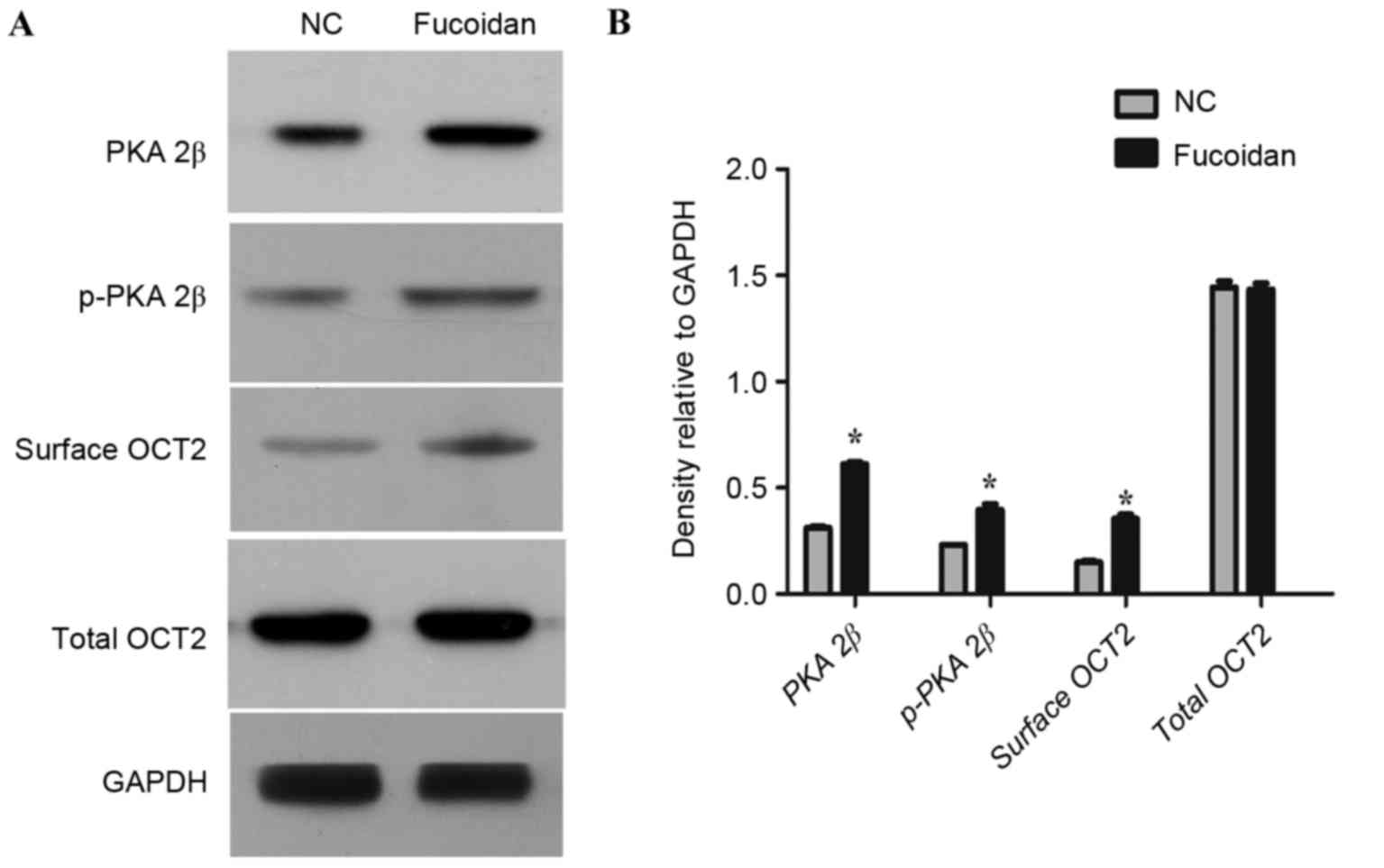
Surface OCT2 is upregulated with PKA
upregulation by fucoidan in OCT2-expressing COS-7 cells
To further ascertain the role of fucoidan in
regulating PKA and surface OCT2 expression, COS-7 cells ectopically
expressing OCT2 were established. COS-7 cells ectopically
expressing OCT2 were treated with fucoidan (500 µg/ml) or PBS for
24 h. A significant increase in the proportion of surface OCT2
protein was induced by fucoidan treatment compared with the control
(P<0.05; Fig. 3), with no
significant difference in total OCT2 level between the two groups.
A significantly elevated PKA 2β and p-PKA 2β level was observed in
OCT2-expressing COS-7 cells treated with fucoidan compared with the
control (both P<0.05; Fig. 3).
These results demonstrated that fucoidan may increase the
expression of PKA and upregulate surface OCT2 in OCT2-expressing
COS-7 cells.
PKA upregulates surface OCT2 in OCT-2
expressing COS-7 cells
Previous studies have reported that PKA regulates
OCT2-mediated organic cation transport (18,19). The
increased expression of PKA by forskolin leads to stimulation of
4-[4-(dimethylamino) styryl]-N-methylpyridinium uptake in
IHKE-1 cells (20). To study the
relationship between PKA expression and surface OCT2 upregulation
in COS-7 cells, surface, intracellular and total OCT2 protein
levels were evaluated in OCT2-expressing COS-7 cells treated with
PKA activator 8-bromo-cAMP. OCT2-expressing COS-7 cells were
treated with PKA activator, 8-bromo-cAMP (1 µmol/l) or PBS. As
demonstrated in Fig. 4, in the
8-bromo-cAMP-treated COS-7 cells, the surface OCT2 protein level
was increased significantly compared with the control (P<0.05),
while intracellular OCT2 level decreased significantly (P<0.01),
with no significant change in total OCT2. The findings presented in
the present study indicate that fucoidan may upregulate surface
OCT2 via PKA to alleviate the symptoms of UAN.
Discussion
UAN is one of the most common and extensive
metabolic diseases worldwide, and is a key risk factor for the
development of uric acid nephrolithiasis, gouty arthritis, renal
diseases and cardiovascular diseases, particularly hypertension
(21–23). The typical characteristic of UAN is
high serum uric acid level, resulting in the accumulation of urate
crystals in the joints and kidneys (24,25). Due
to the fact that acid is the insoluble, circulating end product of
purine nucleotide metabolism in humans, the urate crystals deposit
in the collecting duct (23,26). This induces acute kidney injury,
resulting in decreased filtering capacity of the glomerulus and
renal blood flow, toxic or obstructive injury to the renal tubule,
edema and tubular interstitial inflammation (26).
The multiple biological activities of fucoidan have
been widely studied. Its anticancer properties have been indicated
by its ability to induce apoptosis and macrophage-induced tumor
cell death, block the interactions between cancer cells and the
basement membrane and inhibit angiogenesis by interfering with the
binding of vascular endothelial growth factor and basic fibroblast
growth factor (6,27). The anticoagulant activity of fucoidan
from Fucus evanescens was revealed in vitro and in
vivo (5). In addition, fucoidan
has been indicated to have a potent antiviral effect against herpes
simplex virus types 1 and 2, as well as human cytomegalovirus
(28). The gastroprotective effect
of fucoidan against aspirin-inducedulceration in rats demonstrated
the immunomodulatory effect of fucoidan (29). Fucoidan also suppresses
hypoxia-induced lymphangiogenesis and lymphatic metastasis in
murine hepatocarcinoma by suppressing hypoxia-inducible
factor-1α/vascular endothelial growth factor C (30).
Consistent with previous studies, the results of the
present study indicated that fucoidan was able to alleviate the
symptoms of UAN in a rat model. H&E staining revealed cellular
infiltrate in the tubules, atrophic glomeruli, tubular ectasia,
granuloma hyperplasia focal fibrosis and accumulated urate crystals
in the tubules of renal tissues from rats with UAN, suggesting that
the rat model of UAN was established successfully. The deposition
of urate crystals and inflammatory cellular influx in the tubules
were decreased by fucoidan administration. The proportion of kidney
apoptosis was decreased by fucoidan treatment compared with the
model group, as evaluated by a TUNEL staining assay. The protein
levels of PKA 2β, p-PKA 2β, surface OCT2 and total OCT2 were
evaluated in renal tissues. It was indicated that both the PKA 2β
and p-PKA 2β level were significantly elevated in renal tissues of
the fucoidan treatment group compared with the UAN model group.
These results suggested that increased expression of PKA occurred
in the kidney tissue of fucoidan treatment group rats. The
upregulated protein level of PKA was further demonstrated by
immunohistochemical staining of PKA in rat renal tissues.
Furthermore, the surface OCT2 level was significantly increased by
fucoidan treatment compared with the model group, with no
significant change in total OCT2 level.
OCT2 is critical for organic cation transport,
absorption and excretion of endogenous and exogenous cationic
substances and urinary excretion (31–33).
COS-7 cells ectopically expressing OCT2 were established in the
present study to further evaluate the role of fucoidan in
regulating PKA expression and surface OCT2 expression. It was
indicated that fucoidan was able to increase the expression of PKA
and upregulate surface OCT2 in OCT2-expressing COS-7 cells. The
relationship between PKA and surface OCT2 upregulation in COS-7
cells was also investigated. The increased expression of PKA
induced by 8-bromo-cAMP increased surface OCT2 and decreased
intracellular OCT2 protein levels compared with the control, with
no significant change in total OCT2. Consistent with previous
studies, the upregulation of PKA stimulated OCT2 activity (18,20).
Previous studies have reported that PKA regulates
OCT2 activity in both a heterologous cell system and intact renal
proximal tubules (18–20). The activation of PKA by forskolin
stimulated rabbit OCT2 activity, while inhibition of PKA (by H-89)
reduced transport activity of rabbit OCT2 in CHO-K1 cells (20). However, human OCT2 was reported to be
inhibited by PKA activation and activated by a calmodulin-dependent
signaling pathway in human embryonic kidney HEK-293 cells (34).
In conclusion, the present study identified that
fucoidan upregulated surface OCT2 expression in renal tissues. This
suggests that fucoidan has a protective effect against UAN may have
a potential use in treating UAN. Further in-depth study of the
mechanism by which fucoidan regulates human OCT2 expression and PKA
activation is required.
Acknowledgements
The present study was supported by the Guangdong
Provincial Department of Science and Technology (grant no.
2011B080701007).
References
|
1
|
Cunha L and Grenha A: Sulfated seaweed
polysaccharides as multifunctional materials in drug delivery
applications. Mar Drugs. 14:pii: E42. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
de Souza Rocha MC, Marques CT, Dore Guerra
CM, da Silva Ferreira FR, Rocha Oliveira HA and Leite EL:
Antioxidant activities of sulfated polysaccharides from brown and
red seaweeds. J Appl Phycol. 19:153–160. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Fitton JH, Stringer DN and Karpiniec SS:
Therapies from Fucoidan: An update. Mar Drugs. 13:5920–5946. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Hayashi S, Itoh A, Isoda K, Kondoh M,
Kawase M and Yagi K: Fucoidan partly prevents CCl4-induced liver
fibrosis. Eur J Pharmacol. 580:380–384. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Kuznetsova TA, Besednova NN, Mamaev AN,
Momot AP, Shevchenko NM and Zvyagintseva TN: Anticoagulant activity
of fucoidan from brown algae Fucus evanescens of the Okhotsk Sea.
Bull Exp Biol Med. 136:471–473. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Koyanagi S, Tanigawa N, Nakagawa H, Soeda
S and Shimeno H: Oversulfation of fucoidan enhances its
anti-angiogenic and antitumor activities. Biochem Pharmacol.
65:173–179. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Wang J, Liu H, Li N, Zhang Q and Zhang H:
The protective effect of fucoidan in rats with
streptozotocin-induced diabetic nephropathy. Mar Drugs.
12:3292–3306. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Veena CK, Josephine A, Preetha SP,
Varalakshmi P and Sundarapandiyan R: Renal peroxidative changes
mediated by oxalate: The protective role of fucoidan. Life Sci.
79:1789–1795. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Zhang Q, Li N, Zhao T, Qi H, Xu Z and Li
Z: Fucoidan inhibits the development of proteinuria in active
Heymann nephritis. Phytother Res. 19:50–53. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Zhang Q, Li Z, Xu Z, Niu X and Zhang H:
Effects of fucoidan on chronic renal failure in rats. Planta Med.
69:537–541. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Veena CK, Josephine A, Preetha SP and
Varalakshmi P: Physico-chemical alterations of urine in
experimental hyperoxaluria: A biochemical approach with fucoidan. J
Pharm Pharmacol. 59:419–427. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Okuda M, Saito H, Urakami Y, Takano M and
Inui K: cDNA cloning and functional expression of a novel rat
kidney organic cation transporter, OCT2. Biochem Biophys Res
Commun. 224:500–507. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Karbach U, Kricke J, Meyer-Wentrup F,
Gorboulev V, Volk C, Loffing-Cueni D, Kaissling B, Bachmann S and
Koepsell H: Localization of organic cation transporters OCT1 and
OCT2 in rat kidney. Am J Physiol Renal Physiol. 279:F679–F687.
2000.PubMed/NCBI
|
|
14
|
Muller F and Fromm MF:
Transporter-mediated drug-drug interactions. Pharmacogenomics.
12:1017–1037. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Wilde S, Schlatter E, Koepsell H, Edemir
B, Reuter S, Pavenstadt H, Neugebauer U, Schroter R, Brast S and
Ciarimboli G: Calmodulin-associated post-translational regulation
of rat organic cation transporter 2 in the kidney is gender
dependent. Cell Mol Life Sci. 66:1729–1740. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Wu YR, Qi HJ, Deng DF, Luo YY and Yang SL:
MicroRNA-21 promotes cell proliferation, migration, and resistance
to apoptosis through PTEN/PI3K/AKT signaling pathway in esophageal
cancer. Tumour Biol. 37:12061–12070. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Djordjevic B, Hennessy BT, Li J, Barkoh
BA, Luthra R, Mills GB and Broaddus RR: Clinical assessment of PTEN
loss in endometrial carcinoma: Immunohistochemistry outperforms
gene sequencing. Mod Pathol. 25:699–708. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Zhang Q, Li S, Patterson C and You G:
Lysine 48-linked polyubiquitination of organic anion transporter-1
is essential for its protein kinase C-regulated endocytosis. Mol
Pharmacol. 83:217–224. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Holle SK, Ciarimboli G, Edemir B,
Neugebauer U, Pavenstadt H and Schlatter E: Properties and
regulation of organic cation transport in freshly isolated mouse
proximal tubules analyzed with a fluorescence reader-based method.
Pflugers Arch. 462:359–369. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Soodvilai S, Chatsudthipong A and
Chatsudthipong V: Role of MAPK and PKA in regulation of
rbOCT2-mediated renal organic cation transport. Am J Physiol Renal
Physiol. 293:F21–F27. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Klemp P, Stansfield SA, Castle B and
Robertson MC: Gout is on the increase in New Zealand. Ann Rheum
Dis. 56:22–26. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Arromdee E, Michet CJ, Crowson CS,
O'Fallon WM and Gabriel SE: Epidemiology of gout: Is the incidence
rising? J Rheumatol. 29:2403–2406. 2002.PubMed/NCBI
|
|
23
|
Li Y, Stamler J, Xiao Z, Folsom A, Tao S
and Zhang H: Serum uric acid and its correlates in Chinese adult
populations, urban and rural, of Beijing. The PRC-USA collaborative
study in cardiovascular and cardiopulmonary epidemiology. Int J
Epidemiol. 26:288–296. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Johnson RJ, Kang DH, Feig D, Kivlighn S,
Kanellis J, Watanabe S, Tuttle KR, Rodriguez-Iturbe B,
Herrera-Acosta J and Mazzali M: Is there a pathogenetic role for
uric acid in hypertension and cardiovascular and renal disease?
Hypertension. 41:1183–1190. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Tseng CH: Independent association of uric
acid levels with peripheral arterial disease in Taiwanese patients
with Type 2 diabetes. Diabet Med. 21:724–729. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Shanley PF, Rosen MD, Brezis M, Silva P,
Epstein FH and Rosen S: Topography of focal proximal tubular
necrosis after ischemia with reflow in the rat kidney. Am J Pathol.
122:462–468. 1986.PubMed/NCBI
|
|
27
|
Soeda S, Kozako T, Iwata K and Shimeno H:
Oversulfated fucoidan inhibits the basic fibroblast growth
factor-induced tube formation by human umbilical vein endothelial
cells: Its possible mechanism of action. Biochim Biophys Acta.
1497:127–134. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Lee JB, Hayashi K, Hashimoto M, Nakano T
and Hayashi T: Novel antiviral fucoidan from sporophyll of Undaria
pinnatifida (Mekabu). Chem Pharm Bull (Tokyo). 52:1091–1094. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Raghavendran HR, Srinivasan P and Rekha S:
Immunomodulatory activity of fucoidan against aspirin-induced
gastric mucosal damage in rats. Int Immunopharmacol. 11:157–163.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Teng H, Yang Y, Wei H, Liu Z, Liu Z, Ma Y,
Gao Z, Hou L and Zou X: Fucoidan suppresses Hypoxia-Induced
lymphangiogenesis and lymphatic metastasis in mouse
hepatocarcinoma. Mar Drugs. 13:3514–3530. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Sweet DH, Miller DS and Pritchard JB:
Basolateral localization of organic cation transporter 2 in intact
renal proximal tubules. Am J Physiol Renal Physiol. 279:F826–F834.
2000.PubMed/NCBI
|
|
32
|
Terashita S, Dresser MJ, Zhang L, Gray AT,
Yost SC and Giacomini KM: Molecular cloning and functional
expression of a rabbit renal organic cation transporter. Biochim
Biophys Acta. 1369:1–6. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Grundemann D, Babin-Ebell J, Martel F,
Ording N, Schmidt A and Schomig E: Primary structure and functional
expression of the apical organic cation transporter from kidney
epithelial LLC-PK1 cells. J Biol Chem. 272:10408–10413. 1997.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Cetinkaya I, Ciarimboli G, Yalcinkaya G,
Mehrens T, Velic A, Hirsch JR, Gorboulev V, Koepsell H and
Schlatter E: Regulation of human organic cation transporter hOCT2
by PKA, PI3K, and calmodulin-dependent kinases. Am J Physiol Renal
Physiol. 284:F293–F302. 2003. View Article : Google Scholar : PubMed/NCBI
|