Introduction
Cardiopulmonary bypass (CPB) is a necessary
procedure during open-heart surgery, and its use in clinical
applications has been expanding. However, CPB may induce
postoperative pulmonary dysfunction (1,2). During
CPB, oxygen is supplied to the lung only through the bronchus,
thus, ischemia and hypoxia may occur in the lung. Therefore, the
lung is often reperfused, resulting in anoxia-reoxygenation
(A-R)-induced acute injury (3). A-R
has been reported to induce inflammation and enhance the cellular
permeability of pulmonary vascular endothelial cells, which then
led to pneumonedema, inducing hyoxemia, acute respiratory distress
syndrome and even mortality (4).
Thus, inhibiting inflammation and cellular permeability may improve
the prognosis following CPB (5).
Pre-B-cell colony-enhancing factor was first
identified and cloned from peripheral blood lymphocytes in 1994 by
Samal et al (6).
Subsequently, this protein was also detected in adipose cells, and
named visfatin or nicotinamide phosphoribosyltransferase (NAMPT)
(7). Studies have demonstrated that
NAMPT serves important roles in lung injury following CPB (8,9). NAMPT
is often highly expressed during acute lung injury, and exhibits a
close correlation with the levels of inflammatory factors,
including tumor necrosis factor-α (TNF-α), interleukin-1β (IL-1β)
and IL-6 (8). In addition, a
previous report has also identified that CPB promoted the release
of inflammatory factors and enhanced the cellular permeability of
pulmonary vascular endothelial cells, while upregulation of NAMPT
contributed to A-R-induced cellular permeability enhancement
(9), suggesting that NAMPT may
facilitate CPB-triggered inflammation and the changes in cellular
permeability. However, the mechanism through which NAMPT regulates
these cell responses remains unclear.
Mitogen-activated protein kinase (MAPK) signaling
serves important roles in cell inflammation, apoptosis,
proliferation and differentiation (10). MAPK signaling includes three
pathways: Extracellular signal-regulated kinase (ERK), c-Jun
N-terminal kinase (JNK) and p38 MAPK pathways. Furthermore, MAPK
signaling is activated by post-translational phosphorylation
mediated by various kinases (11,12). It
has been demonstrated that MAPK signaling was activated in the rat
lung tissue following CPB (13).
However, this signaling was blocked following MAPK signaling
inhibitor treatment, and simultaneously, the release of
inflammatory factors and infiltration were restrained (14). Therefore, MAPK signaling may
participate in CPB-induced lung injury, although whether NAMPT
regulates CPB-triggered inflammation and cellular permeability
enhancement of lung tissue through MAPK signaling remains
unknown.
In the present study, an A-R model was established
using human umbilical vein endothelial cells (HUVECs) to
investigate the regulation of MAPK signaling by NAMPT during
CPB-induced A-R. The present data demonstrated that NAMPT was
upregulated by A-R, and subsequently induced the release of
inflammatory factors and the expression levels of cellular
permeability-associated proteins. Additionally, calcium and MAPK
signaling were activated; however, NAMPT knockdown blocked
A-R-induced MAPK signaling and the cell responses, including
inflammation and cellular permeability enhancement. The present
study also indicated that NAMPT may promote A-R-induced
inflammation and cellular permeability enhancement through
activating MAPK signaling.
Materials and methods
Chemicals
Chemicals were purchased commercially as follows:
Bicinchoninic acid (BCA) kit, enhanced chemiluminescence (ECL)
reagent, Dulbecco's modified Eagle's medium (DMEM) and fetal bovine
serum (FBS) were obtained from GE Healthcare Life Sciences
(Hyclone; Logan, UT, USA); horseradish peroxidase (HRP)-labeled
goat anti-Rabbit IgG (H+L) (cat. no. A0208) and glyceraldehyde
phosphate dehydrogenase (GAPDH) mouse monoclonal antibodies (cat.
no. AF0006) were purchased from Beyotime Institute of Biotechnology
(Shanghai, China); enzyme-linked immunosorbent assay (ELISA) kits
for TNF-α (cat. no. MBS2502004), IL-1β (cat. no. MBS175901) and
IL-6 (cat. no. MBS021993; all from MyBioSource, Inc., San Diego,
CA, USA); rabbit polyclonal antibodies against myosin light-chain
(MLC; cat. no. ab79935), phosphorylated (p)-MLC (cat. no. ab2480),
vascular endothelial (VE)-cadherin (cat. no. ab33168),
p-VE-cadherin (cat. no. ab119785), β-catenin (cat. no. ab6302),
focal adhesion kinase (FAK; cat. no. ab61113), p-FAK (cat. no.
ab39967), paxillin (cat. no. ab2264), p38 MAPK (cat. no. ab197348),
p-p38 (cat. no. ab47363), ERK (cat. no. ab17942) and p-ERK (cat.
no. ab24157) were from Abcam (Cambridge, MA, USA); and
Lipofectamine 2000 was from Thermo Fisher Scientific, Inc.
(Waltham, MA, USA).
Cell culture, establishment of A-R
model and small interfering RNA (siRNA) transfection
HUVECs (cat. no. CRL1730) were purchased from
American Type Culture Collection (ATCC, Manassas, VA, USA), and
maintained in DMEM supplemented with 10% (v/v) FBS. The A-R model
was established as described in a previous study (9). Briefly, HUVECs in the logarithmic
growth phase were arranged into the sterilized hypoxic box. The box
was filled with 5% CO2 and 94% N2, and the
oxygen concentration decreased to 1% within 30 min. After 20-h
hypoxic treatment, the cells were maintained in a normal incubator
with 5% CO2 at 37°C to establish the A-R model.
After an additional 9 h, A-R-treated HUVECs were
used in subsequent siRNA transfection. For NAMPT knockdown, NAMPT
siRNA and negative control siRNA were synthesized by Invitrogen;
Thermo Fisher Scientific, Inc. according to the reported sequences:
NAMPT siRNA, 5′-CCACCCAACACAAGCAAAGUUUAUU-3′; and negative control
siRNA, 5′-UUCUCCGAACGUGUCACGUTT-3′. These siRNAs were then
transfected into the A-R-treated HUVECs in FBS-free DMEM by using
Lipofectamine 2000 following the manufacturer's protocol
(Invitrogen; Thermo Fisher Scientific, Inc.). After 6-h
transfection, the cells were cultured in DMEM with 10% FBS for an
additional 48 h.
Detection of the TNF-α, IL-1β and IL-6
content by ELISA and of the Ca2+ levels by
Fura-2-acetoxymethyl ester (Fura-2-AM)
The four treatment groups included the normal
control cells (without A-R induction), A-R model cells, A-R cells
transfected with negative control siRNA and A-R cells transfected
with NAMPT siRNA transfection. Following treatment, the cells were
centrifuged at 1,000 × g at 4°C for 5 min to harvest the
supernatant. According to the ELISA kit instructions, the contents
of TNF-α, IL-1β and IL-6 in each group were detected based on three
independent replicates using a microplate reader (Infinite F50;
Tecan Trading AG, Männedorf, Switzerland).
For detection of Ca2+ levels, the cells
were treated with 1 µM Fura-2-AM (Invitrogen; Thermo Fisher
Scientific, Inc.) and 2 mM calcium chloride (Sigma-Aldrich; Merck
KGaA, Darmstadt, Germany) for 30 min at 37°C. Subsequently, cells
were washed three times with phosphate-buffered saline, and
fluorescence was detected at 555 nm using a laser scan confocal
microscope (Carl Zeiss LSM 700; Carl Zeiss, Thornwood, NY,
USA).
Western blot analysis of protein
expression levels
The cells were centrifuged at 1,000 × g at 4°C for 5
min, and the precipitates were lysed in radioimmunoprecipitation
assay buffer (150 mM NaCl, 1% NP-40, 0.5% sodium deoxycholate, 0.1%
SDS, 50 mM Tris, pH 8.0, 5.0 mM EDTA, pH 8.0, 0.5 mM dithiothreitol
and 1 mM phenylmethylsulfonyl fluoride) containing protease
inhibitors for 30 min. The cell lysates were then centrifuged at
12,000 × g at 4°C for 10 min, and protein concentrations in the
supernatants were detected using the BCA kit according to the
manufacturer's instructions. Next, the proteins were treated with
SDS sample buffer in boiling water for 10 min, then separated using
12.5% SDS-polyacrylamide gel electrophoresis and transferred to a
nitrocellulose membrane. The membranes were incubated with blocking
buffer (5% bovine serum albumin (Sigma-Aldrich; Merck KGaA), 50 mM
Tris, 150 mM NaCl, 0.1% Tween-20) for 1 h at room temperature, and
subsequently incubated with the corresponding primary antibodies
against MLC, p-MLC, VE-cadherin, p-VE-cadherin, β-catenin, FAK,
p-FAK, paxillin, p38 MAPK, p-p38, ERK and p-ERK at a 1:1,000
dilution at 4°C overnight. After being washed with wash buffer (50
mM Tris, 150 mM NaCl, 0.1% Tween-20) three times for 5 min each
time, the membranes were incubated with the HRP-labeled goat
anti-rabbit IgG (H+L) secondary antibody at room temperature for 2
h. Following treatment with the ECL regent, the target protein
bands were observed by Molecular Image ChemiDoc XRS system (Bio-Rad
Laboratories, Inc., Hercules, CA, USA). The protein expression was
quantitatively analyzed by Quantity One software (version 4.62;
Bio-Rad Laboratories, Inc.).
Statistical analysis
All the data were collected from three independent
replicates and analyzed with SPSS version 17.0 software package
(SPSS, Inc., Chicago, IL, USA). Statistically significant
differences were analyzed by Student's t-test and considered when
P<0.05.
Results
NAMPT knockdown by siRNA transfection
inhibits A-R-induced release of inflammatory factors in HUVECs
In the present study, an A-R model was established
in HUVECs in order to investigate the mechanisms underlying
A-R-induced inflammation. Compared with the normal control, A-R
clearly upregulated NAMPT protein expression (Fig. 1A). Following transfection of the
cells with NAMPT siRNA, NAMPT expression was markedly
downregulated, even under A-R treatment, as compared with the
negative control siRNA transfection group (Fig. 1A). Next, the contents of three
inflammatory factors in the culture medium of HUVECs were detected
using ELISA subsequent to NAMPT knockdown. As shown in Fig. 1B-D, TNF-α, IL-1β and IL-6 contents
were significantly increased following A-R treatment along with the
NAMPT upregulation (P<0.01). However, NAMPT knockdown by siRNA
transfection significantly inhibited the A-R-upregulated
inflammatory factor contents (P<0.01; Fig. 1B-D). Therefore, the results suggest
that A-R induces inflammation in HUVECs by upregulating NAMPT
expression.
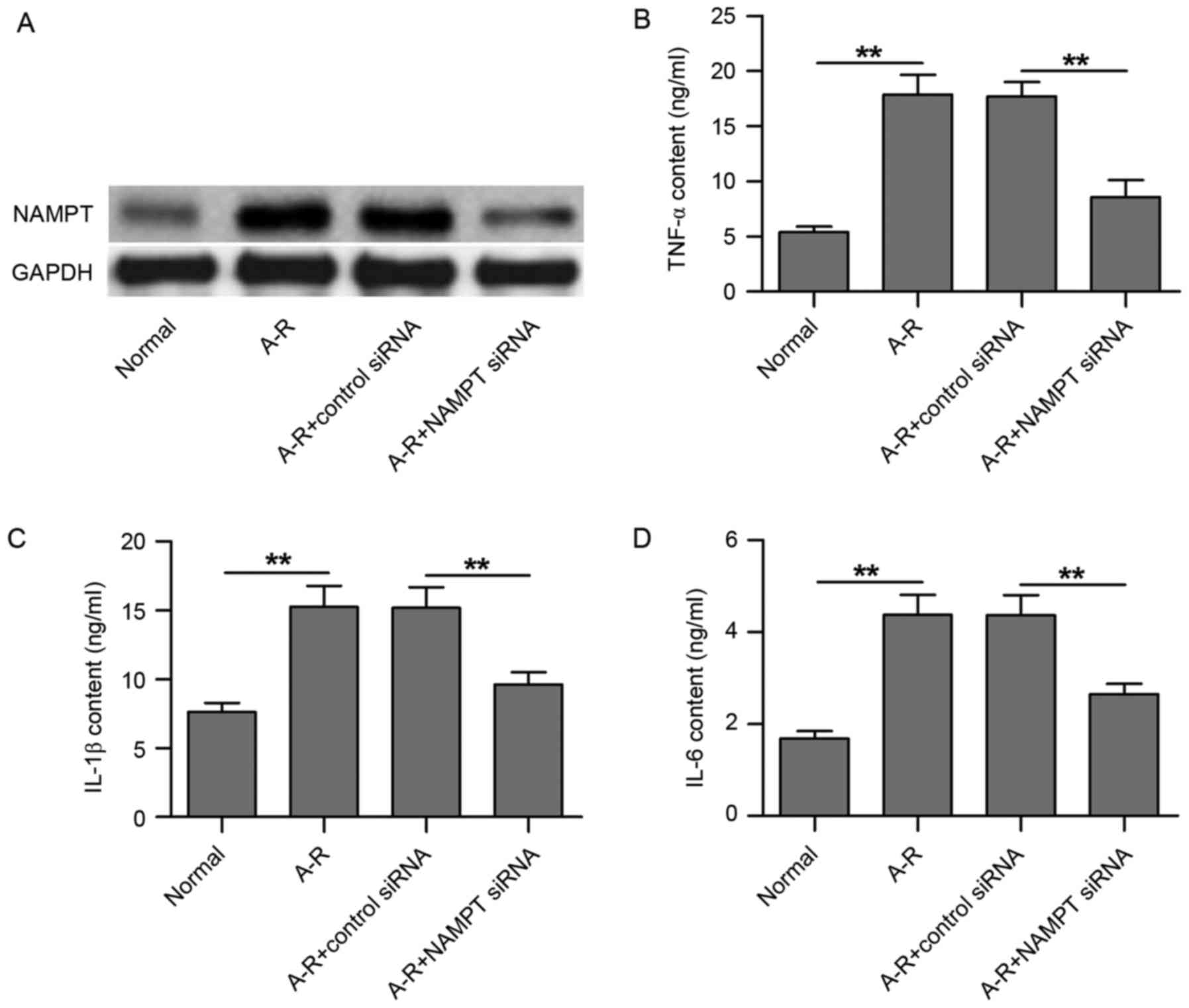 | Figure 1.NAMPT knockdown by siRNA transfection
inhibits A-R-induced release of inflammatory factors in HUVECs. (A)
Western blots demonstrating the expression of NAMPT in the four
groups of treated cells, including normal cells, A-R-induced cells,
A-R-induced cells transfected with negative control siRNA, and
A-R-induced cells transfected with NAMPT siRNA. Enzyme-linked
immunosorbent assay was used to analyze the contents of (B) TNF-α,
(C) IL-1β and (D) IL-6 in the four treated cell groups. Normal and
negative control siRNA-transfected cells were used as the controls
corresponding to the A-R-induced cells and A-R+NAMPT
siRNA-transfected cells, respectively. Values are expressed as the
mean ± standard deviation based on three independent replicates.
Statistical significance was calculated with the Student's t-test.
**P<0.01. NAMPT, nicotinamide phosphoribosyltransferase; A-R,
anoxia-reoxygenation; siRNA, small interfering RNA; TNF-α, tumor
necrosis factor-α; IL, interleukin. |
NAMPT knockdown blocks A-R-induced
expression of cellular permeability-associated proteins and
Ca2+ increase
A previous study has demonstrated that the cellular
permeability of endothelial cells is mediated by intercellular
adherens and the junction between cell and extracellular matrix
(15). VE-cadherin interacts with
β-catenin to form intercellular adherens junctions, and is then
connected to actin cytoskeleton to regulate cellular permeability
(16). In addition, FAK interacts
with paxillin and vinculin to form cell-extracellular matrix
adhesions, and regulates cellular permeability through actin
cytoskeleton (16). Calcium
signaling activated MLC kinase (MLCK) to phosphorylate MLC and
promote the connection between MLC and actin for the regulation of
cell contraction (17). Therefore,
calcium signaling is able to regulate intercellular adherens
junctions and cell-extracellular matrix adhesions for cellular
permeability changes through adjusting actin. Previous studies
observed that NAMPT induced the enhancement of cellular
permeability of pulmonary vascular endothelial cell during
A-R-induced acute lung injury (18,19), and
the aforementioned results of the present study suggested that
NAMPT was upregulated in HUVECs under A-R treatment. Therefore, the
current study attempted to clarify whether NAMPT serves a role in
the A-R-induced cellular permeability enhancement of HUVECs.
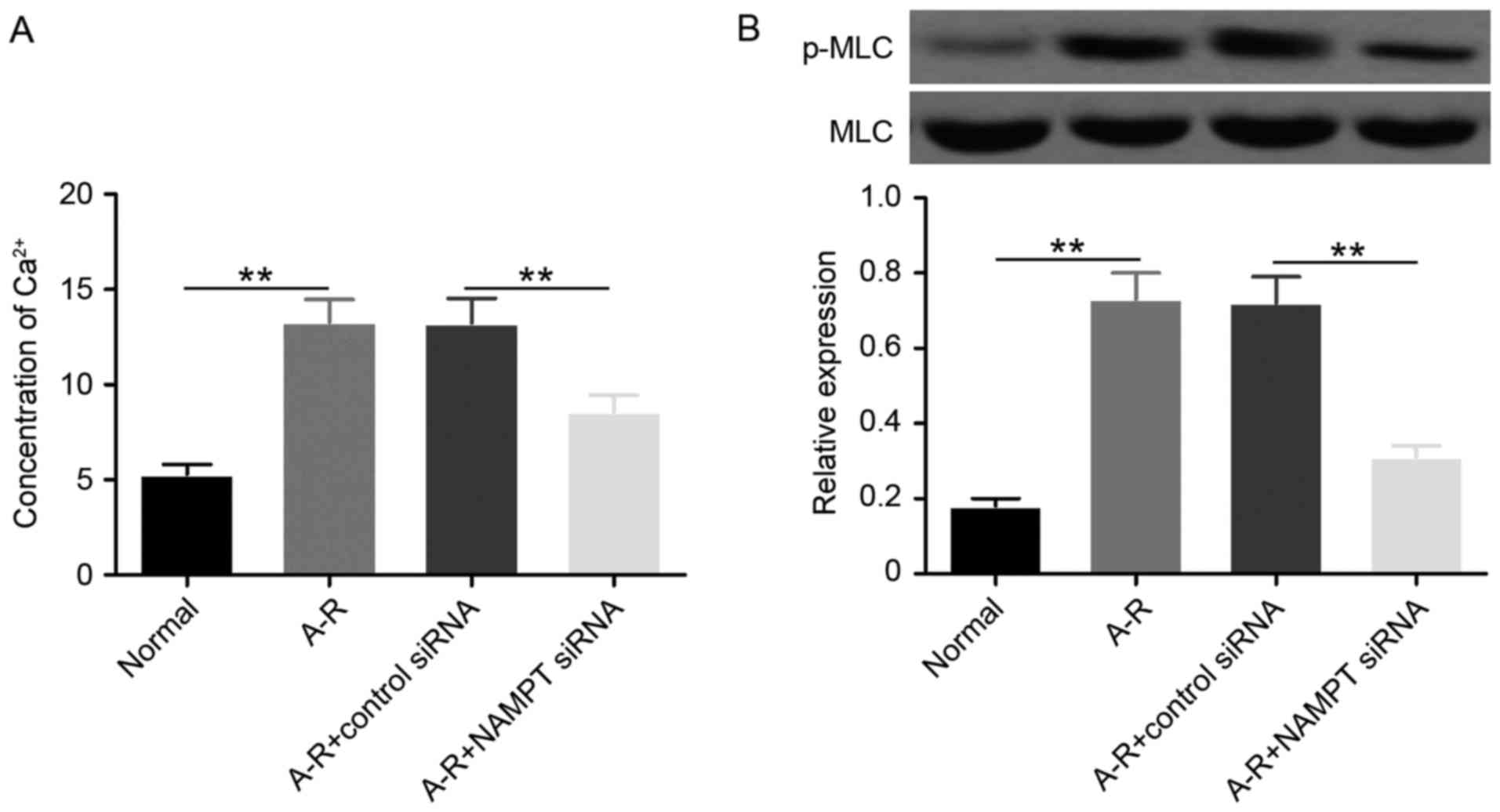
The calcium-mediated phosphorylation of MLC (p-MLC
protein) was first detected to investigate the regulation of actin
cytoskeleton by NAMPT under A-R induction. The data revealed that
A-R significantly upregulated (P<0.01) the Ca2+ level
and the phosphorylation of MLC compared with the normal control
group (Fig. 2). When NAMPT was
depleted by siRNA transfection, the levels of Ca2+ and
p-MLC were significantly downregulated (P<0.01) under A-R
treatment. These results suggested that NAMPT may promote
A-R-induced actin cytoskeleton contraction by activating the
Ca2+-MLC pathway.
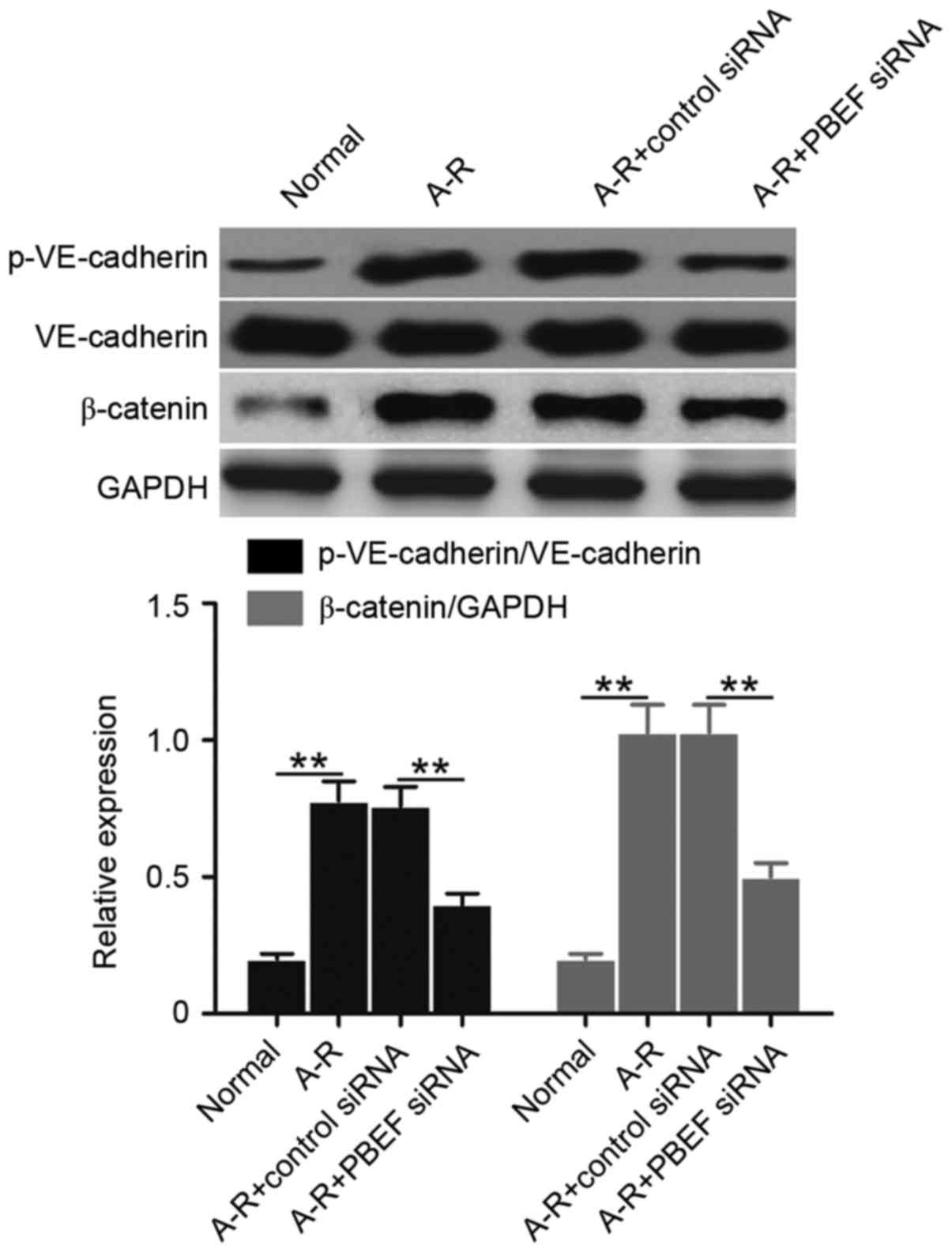
Subsequently, the levels of VE-cadherin
phosphorylation (p-VE-cadherin protein) and β-catenin expression
were examined to determine whether NAMPT is able to regulate the
A-R-induced derangement of intercellular adherens junctions.
Compared with the normal control, A-R significantly upregulated the
levels of p-VE-cadherin and β-catenin (P<0.01; Fig. 3). However, the increased expression
was significantly blocked subsequent to NAMPT knockdown (P<0.01;
Fig. 3). The data indicated that
NAMPT may regulate intercellular adherens junctions during A-R.
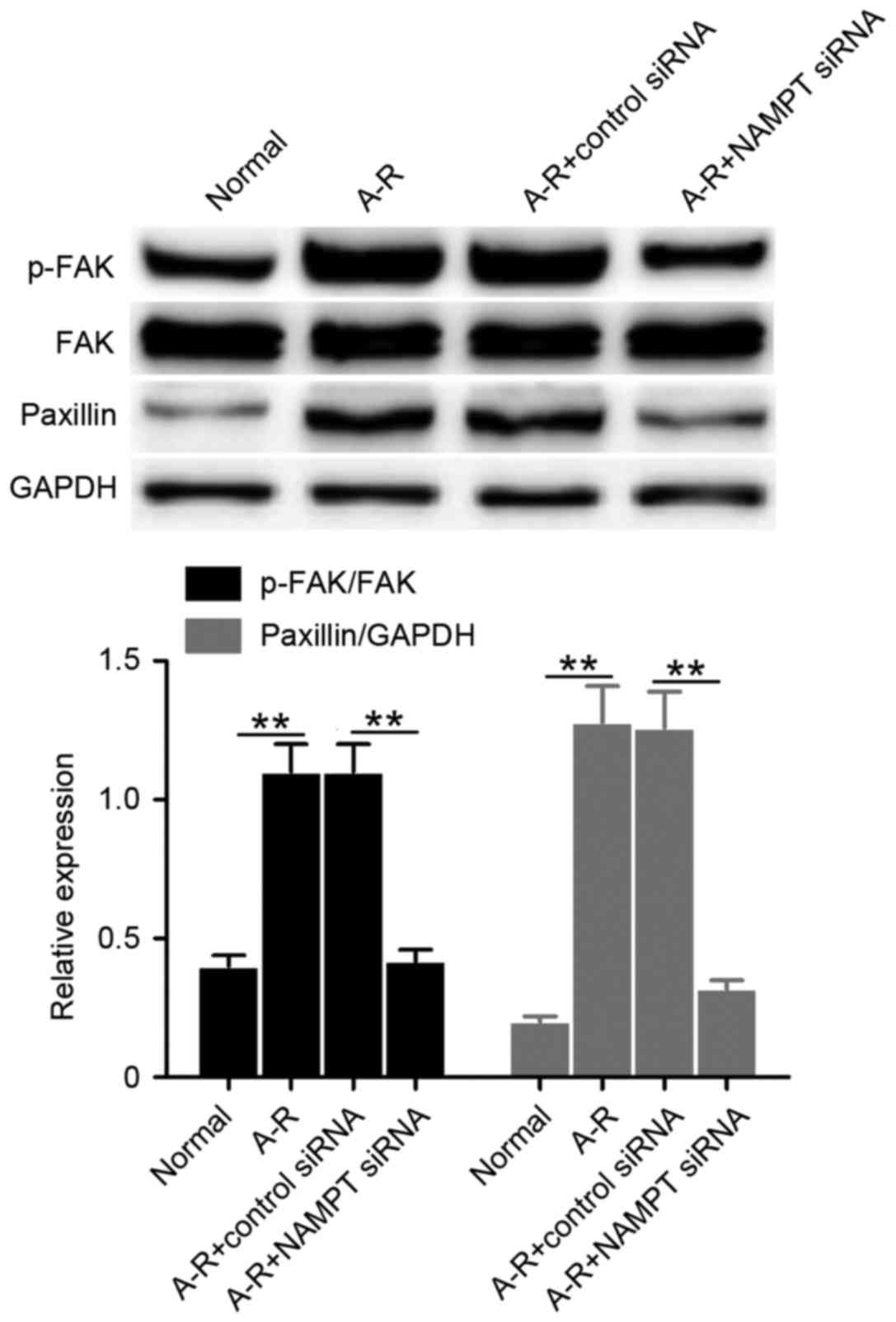
The study also examined the levels of FAK
phosphorylation (p-FAK protein) and paxillin expression to
determine the effect of cell-extracellular matrix adhesions by
NAMPT under A-R treatment. Clearly, the levels of p-FAK and
paxillin significantly increased following A-R induction compared
with the normal control levels (P<0.01; Fig. 4). By contrast, NAMPT knockdown
significantly suppressed this A-R-induced upregulation (P<0.01;
Fig. 4). The results suggested that
NAMPT may regulate cell-extracellular matrix adhesions during A-R.
Collectively, the data indicate that NAMPT may mediate cellular
permeability by affecting the intercellular adherens and the
junction between cell and extracellular matrix.
NAMPT knockdown restrains
A-R-activated MAPK signaling
It has been demonstrated that MAPK signaling is
activated during CPB-triggered acute lung injury, and mediates
cellular inflammation and permeability (20). Thus, the present study tested the
phosphorylation of p38 MAPK and ERK to investigate the regulation
of MAPK signaling by NAMPT under A-R treatment. Compared with the
normal control, p38 and ERK phosphorylation significantly increased
following A-R treatment (P<0.01; Fig.
5). However, siRNA-mediated NAMPT knockdown significantly
decreased the A-R-induced phosphorylation of p38 and ERK
(P<0.01; Fig. 5). These results
suggested that NAMPT positively regulates the A-R-activated MAPK
signaling.
Discussion
Previous studies revealed that NAMPT serves as a
growth factor and cytokine (18). In
particular, NAMPT has been identified as an inflammatory factor and
a potential novel biomarker in acute lung injury (21). Further research demonstrated that
NAMPT was expressed in lung microvascular endothelium (21). In mice, NAMPT regulated
ventilator-induced lung injury through mediating the release of
inflammatory factors, including IL-6 and TNF-α (22). Previous studies have also observed
that NAMPT expression was upregulated in pulmonary cells, and that
it may contribute to inflammatory factor expression (23). Therefore, NAMPT is considered to
participate in the inflammation during acute lung injury (18). In the present study, an A-R-induced
model was established using HUVECs to investigate how NAMPT
regulates inflammation. It was observed that A-R significantly
induced the expression of NAMPT and the release of TNF-α, IL-1β and
IL-6, suggesting that NAMPT may promote A-R-induced inflammation by
facilitating the release of inflammatory factors. Next, NAMPT
knockdown was performed via siRNA transfection, and it was
demonstrated that the release of these three inflammatory factors
was controlled by the high expression of NAMPT. Thus, these results
indicated that NAMPT promotes A-R-induced inflammation by
facilitating the release of TNF-α, IL-1β and IL-6. Similar results
were also observed in a previous study during rat acute lung
injury, in which NAMPT was highly expressed, and NAMPT inhibitor
decreased TNF-α and IL-1β contents (24). Notably, inhibition of NAMPT was able
to attenuate the pandemic H1N1 2009 virus-induced inflammation in
the lung endothelium, for example, by decreasing the expression
levels of IL-6, IL-8, and TNF-α (25). Therefore, NAMPT may be an important
upstream factor that regulates inflammation.
Cellular permeability alterations in endothelial
cells participate in acute lung injury, while the cellular
permeability in these cells has been demonstrated to be mediated by
intercellular and cell-extracellular matrix adherens (15). VE-cadherin interacts with β-catenin
to form intercellular adherens junctions, and then targets the
actin cytoskeleton to regulate cellular permeability (16). Furthermore, FAK interacts with
paxillin and vinculin to form cell-extracellular matrix adhesions,
and subsequently regulates cellular permeability through the actin
cytoskeleton (16). Calcium
signaling regulates intercellular adherens junctions and
cell-extracellular matrix adhesions for cellular permeability
changes through activating MLCK-mediated MLC phosphorylation.
Phosphorylated MLC then promotes the connection between MLC and
actin to regulate cell contraction (17). In human pulmonary artery endothelial
cells, high expression of NAMPT has been reported to regulate
thrombin-induced alterations of cellular permeability by promoting
Ca2+ entry, MLC phosphorylation and actin polymerization
(26). The present study also
identified that NAMPT regulated the A-R-induced cellular
permeability changes in HUVECs by mediating actin function,
including Ca2+ signaling and MLC phosphorylation.
Furthermore, it was demonstrated that NAMPT regulated VE-cadherin
phosphorylation and β-catenin expression, suggesting the regulation
of intercellular adherens junctions by NAMPT. Meanwhile, NAMPT also
mediated cell-extracellular matrix adhesions by regulating FAK
phosphorylation and paxillin expression. Hence, NAMPT serves a
vital role in the regulation of cellular permeability under A-R
conditions. Combined with previous research, the current study
suggests that NAMPT regulates cellular permeability by mediating
intercellular and cell-extracellular matrix adherens.
NAMPT promotes acute lung injury through regulating
inflammation and cellular permeability, while MAPK signaling may
also be involved in this process (18). MAPK signaling includes the ERK, p38
MAPK and JNK pathways, and serves an important role in cell
inflammation, apoptosis, proliferation and differentiation. MAPK
signaling is activated by post-translational phosphorylation
mediated by various kinases (11,12).
Studies have shown that p38 MAPK and ERK pathways participate in
the inflammation of endothelial cells, while JNK pathway does not
function in this process (27). In
pig lungs following CPB, ERK and p38 MAPK were detected to be
significantly activated (28).
Furthermore, in reperfused rat lungs, the phosphorylation of p38
MAPK and ERK was also upregulated (13). Our previous study demonstrated that
p38 MAPK was activated in rat lungs following CPB and that the
inhibitor of p38 MAPK restrained CPB-triggered inflammation
(14). Therefore, MAPK signaling
serves an important role in acute lung injury and the associated
inflammation. In addition, MAPK signaling also regulates the
cellular permeability of pulmonary endothelial cells (29,30).
Phosphorylated p38 MAPK was able to activate heat shock protein 27
in order to regulate the cellular permeability of rat pulmonary
microvascular endothelial cells (31). p38 MAPK also regulated TGF-β-mediated
MLC phosphorylation to affect permeability of endothelial cells
(32). During vascular endothelial
growth factor-induced cellular permeability changes, ERK was
identified as an important regulator (33). Although NAMPT promoted inflammation
in human pulmonary microvascular endothelial cells, inhibition of
MAPK blocked NAMPT function, suggesting that NAMPT may regulate
inflammation via MAPK signaling (34). These studies indicated that
NAMPT-mediated lung injury may depend on MAPK signaling.
In the present study, it was observed that A-R
significantly induced the expression of NAMPT and the
phosphorylation of p38 MAPK and ERK in HUVECs. When NAMPT was
depleted by siRNA transfection, the phosphorylation levels
decreased. Clearly, NAMPT controls the A-R-induced activation of
MAPK signaling. Combined with previous research, it can be
concluded that NAMPT promotes A-R-induced inflammation and cellular
permeability through MAPK signaling. The current study indicates
that NAMPT may be a potential drug target for CPB-triggered acute
injury of endothelial cells.
Acknowledgements
The present study was supported by the National
Natural Science Foundation of China (grant no. 81260054).
References
|
1
|
Huffmyer JL and Groves DS: Pulmonary
complications of cardiopulmonary bypass. Best Pract Res Clin
Anaesthesiol. 29:163–175. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Thiessen S, Vanhorebeek I and Van den
Berghe G: Glycemic control and outcome related to cardiopulmonary
bypass. Best Pract Res Clin Anaesthesiol. 29:177–187. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Durandy Y: Minimizing systemic
inflammation during cardiopulmonary bypass in the pediatric
population. Artif Organs. 38:11–18. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Ferrari RS and Andrade CF: Oxidative
stress and lung ischemia-reperfusion injury. Oxid Med Cell Longe.
2015:5909872015. View Article : Google Scholar
|
|
5
|
Chao YK, Wu YC, Yang KJ, Chiang LL, Liu
HP, Lin PJ and Chu Y: Pulmonary perfusion with L-arginine
ameliorates post-cardiopulmonary bypass lung injury in a rabbit
model. J Surg Res. 167:e77–e83. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Samal B, Sun YH, Stearns G, Xie CS, Suggs
S and McNiece I: Cloning and characterization of the cDNA encoding
a novel human pre-B-cell colony-enhancing factor. Mol Cell Biol.
14:1431–1437. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Chang YC, Chang TJ, Lee WJ and Chuang LM:
The relationship of visfatin/pre-B-cell colony-enhancing
factor/nicotinamide phosphoribosyltransferase in adipose tissue
with inflammation, insulin resistance, and plasma lipids.
Metabolism. 59:93–99. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Bajwa EK, Yu CL, Gong MN, Thompson BT and
Christiani DC: Pre-B-cell colony-enhancing factor gene
polymorphisms and risk of acute respiratory distress syndrome. Crit
Care Med. 35:1290–1295. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Yang W, Zeng Y, Li B, Zhou J, Gong Y, Xu J
and Dong X: Pre-B-cell colony enhancing factor (PBEF) increases
endothelial permeability in hypoxia/re-oxygenation model. Int J
Clin Exp Med. 8:8842–8847. 2015.PubMed/NCBI
|
|
10
|
Mordret G: MAP kinase kinase: A node
connecting multiple pathways. Biol Cell. 79:193–207. 1993.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
L'Allemain G: Deciphering the MAP kinase
pathway. Prog Growth Factor Res. 5:291–334. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Munshi A and Ramesh R: Mitogen-activated
protein kinases and their role in radiation response. Genes Cancer.
4:401–408. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Markou T and Chambers DJ: Lung injury
after simulated cardiopulmonary bypass in an isolated perfused rat
lung preparation: Role of mitogen-activated protein kinase/Akt
signaling and the effects of theophylline. J Thorac Cardiovasc
Surg. 148:2335–2344. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Dong X, Liu Y, Du M, Wang Q, Yu CT and Fan
X: P38 mitogen-activated protein kinase inhibition attenuates
pulmonary inflammatory response in a rat cardiopulmonary bypass
model. Eur J Cardiothorac Surg. 30:77–84. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Lum H and Malik AB: Regulation of vascular
endothelial barrier function. Am J Physiol. 267:L223–L241.
1994.PubMed/NCBI
|
|
16
|
Mehta D and Malik AB: Signaling mechanisms
regulating endothelial permeability. Physiol Rev. 86:279–367. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Van Lierop JE, Wilson DP, Davis JP,
Tikunova S, Sutherland C, Walsh MP and Johnson JD: Activation of
smooth muscle myosin light chain kinase by calmodulin. Role of
LYS(30) and GLY(40). J Biol Chem. 277:6550–6558. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Sun ZJ, Lei H and Zhang Z: Pre-B cell
colony enhancing factor (PBEF), a cytokine with multiple
physiological functions. Cytokine Growth Factor Rev. 24:433–442.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Dahl TB, Yndestad A, Skjelland M, Øie E,
Dahl A, Michelsen A, Damås JK, Tunheim SH, Ueland T, Smith C, et
al: Increased expression of visfatin in macrophages of human
unstable carotid and coronary atherosclerosis: Possible role in
inflammation and plaque destabilization. Circulation. 115:972–980.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Zakkar M, Guida G, Suleiman M and Angelini
GD: Cardiopul-monary bypass and oxidative stress. Oxid Med Cell
Longe. 2015:1898632015. View Article : Google Scholar
|
|
21
|
Ye SQ, Simon BA, Maloney JP,
Zambelli-Weiner A, Gao L, Grant A, Easley RB, McVerry BJ, Tuder RM,
Standiford T, et al: Pre-B-cell colony-enhancing factor as a
potential novel biomarker in acute lung injury. Am J Respir Crit
Care Med. 171:361–370. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Hong SB, Huang Y, Moreno-Vinasco L,
Sammani S, Moitra J, Barnard JW, Ma SF, Mirzapoiazova T, Evenoski
C, Reeves RR, et al: Essential role of pre-B-cell colony enhancing
factor in ventilator-induced lung injury. Am J Respir Crit Care
Med. 178:605–617. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Liu P, Li H, Cepeda J, Zhang LQ, Cui X,
Garcia JG and Ye SQ: Critical role of PBEF expression in pulmonary
cell inflammation and permeability. Cell Biol Int. 33:19–30. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Liu C, Zhang H, Cheng PY and Zhou FC: The
influence of pre-B-cell colony enhancing factor on adhesive
molecule in pulmonary cells in rats with acute lung injury/acute
respiratory distress syndrome. Zhonghua Wei Zhong Bing Ji Jiu Yi
Xue. 25:159–163. 2013.(In Chinese). PubMed/NCBI
|
|
25
|
Gao W, Mao Q, Feng AW, Sun HM, Sun WK, Lu
X, Su X and Shi Y: Inhibition of pre-B cell colony-enhancing factor
attenuates inflammation and apoptosis induced by pandemic H1N1 2009
in lung endothelium. Respir Physiol Neurobiol. 178:235–241. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Ye SQ, Zhang LQ, Adyshev D, Usatyuk PV,
Garcia AN, Lavoie TL, Verin AD, Natarajan V and Garcia JG:
Pre-B-cell-colony-enhancing factor is critically involved in
thrombin-induced lung endothelial cell barrier dysregulation.
Microvasc Res. 70:142–151. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Liu SW, Qiao SB, Yuan JS and Liu DQ:
Visfatin stimulates production of monocyte chemotactic protein-1
and interleukin-6 in human vein umbilical endothelial cells. Horm
Metab Res. 41:281–286. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Khan TA, Bianchi C, Araujo EG, Ruel M,
Voisine P and Sellke FW: Activation of pulmonary mitogen-activated
protein kinases during cardiopulmonary bypass. J Surg Res.
115:56–62. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Yang D, Xie P, Guo S and Li H: Induction
of MAPK phosphatase-1 by hypothermia inhibits TNF-alpha-induced
endothelial barrier dysfunction and apoptosis. Cardiovasc Res.
85:520–529. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Sun HM, Hong LZ, Shen XK, Lin XQ, Song Y
and Shi Y: Antithrombin-III without concomitant heparin improves
endotoxin-induced acute lung injury rats by inhibiting the
activation of mitogen-activated protein kinase. Chin Med J (Engl).
122:2466–2471. 2009.PubMed/NCBI
|
|
31
|
Liu T, Milia E, Warburton RR, Hill NS,
Gaestel M and Kayyali US: Anthrax lethal toxin disrupts the
endothelial permeability barrier through blocking p38 signaling. J
Cell Physiol. 227:1438–1445. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Goldberg PL, MacNaughton DE, Clements RT,
Minnear FL and Vincent PA: p38 MAPK activation by TGF-beta1
increases MLC phosphorylation and endothelial monolayer
permeability. Am J Physiol Lung Cell Mol Physiol. 282:L146–L154.
2002.PubMed/NCBI
|
|
33
|
Breslin JW, Pappas PJ, Cerveira JJ, Hobson
RW II and Durán WN: VEGF increases endothelial permeability by
separate signaling pathways involving ERK-1/2 and nitric oxide. Am
J Physiol Heart Circ Physiol. 284:H92–H100. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Ming GF, Ma XH, Xu DM, Liu ZY, Ai YH, Liu
HX and Shi ZH: PBEF promotes the apoptosis of pulmonary
microvascular endothelial cells and regulates the expression of
inflammatory factors and AQP1 through the MAPK pathways. Int J Mol
Med. 36:890–896. 2015. View Article : Google Scholar : PubMed/NCBI
|