Introduction
Pseudomonas aeruginosa (P. aeruginosa)
is a ubiquitous Gram negative bacterium capable of surviving in
several environmental niches such as mammals (including humans),
insects, nematodes, soil, water and plants (1,2). In
humans, P. aeruginosa rarely causes community acquired
pneumonia (3,4), but has an increased incidence of
causing hospital infections like hospital-acquired pneumonia,
sepsis, urinary tract infections, and is prevalent among wound and
burn patients (1,5). The bacterium causes severe pulmonary
damage and is a leading cause of mortality in cystic fibrosis
patients (6–8). This opportunistic pathogen poses a huge
challenge in clinical settings as compared to other pathogens
because of its highly intrinsic resistance to most antibiotics
including β-lactams, fluoroquinolones and aminoglycosides (9–11). Each
year, the bacterium is estimated to cause 51,000
healthcare-associated infections out of which 6,000 cases are
multidrug resistant with roughly 400 deaths occurring per year in
the United States (9). In addition,
P. aeruginosa has the ability to switch from free-living
(planktonic) to biofilm phenotypic mode of living. This ability
contributes to antibiotic resistance and is governed by quorum
sensing systems that regulate virulence factor production as well
(1,12–14).
Unlike other pathogens, P. aeruginosa has a unique virulence
potential, its natural resistance to several antibiotics and other
resistance mechanisms like antibiotic modification and
energy-dependant drug efflux make infections very problematic to
control (15,16). Therefore, there is need to search for
more compounds with antimicrobial potential to curb P.
aeruginosa-related infections.
In the recent past, fatty acids (FA) have received
attention because of their antimicrobial and anti-inflammatory
properties (17,18). They possess beneficial effects
against cancerous cells, body fats, and in conditions such as
depression, heart problems, neurodegenerative diseases, joint and
bone conditions (19,20). One of the widely discussed fatty acid
family based on nutrition and human health benefits is the omega-3
family in which linolenic acid (C18:3n-3, LNA) is a parent fatty
acid compound (21). Evidence has
shown that LNA and its derivatives, eicosapentaenoic acid (EPA) and
docosahexaenoic acid (DHA) possess antimicrobial properties against
many microorganisms (22–26). Thus far, bacterial resistance towards
free fatty acids is yet to be reported due to their broader
spectrum activities (27). However,
the last option of antimicrobial agents for multidrug resistant
P. aeruginosa related infections include aminoglycosides and
polymyxins, which besides losing their efficacy are also
accompanied with side effects such as neurotoxicity, nephrotoxicity
and ototoxicity (28–30). Nonetheless, these toxic side effects
can be managed with reduced concentration and duration of
tobramycin therapy. Besides, some studies have shown that
phytochemical compounds can produce synergistic and additive
effects with aminoglycosides in various bacterial infections
thereby improving their (aminoglycosides) efficacious potential
(31).
Despite several reports on the antibacterial
activity of LNA and its omega-3 derivatives (EPA and DHA), the
anti-biofilm mechanism and their interaction with aminoglycosides
on P. aeruginosa pathogens have not been elucidated.
Therefore, the present study aimed at assessing the anti-biofilm
activity and mechanism of linolenic acid alone and in combination
with tobramycin on P. aeruginosa, with the view to
evaluating the potency of the compound alone or in combination for
treating infections associated with this pathogen.
Materials and methods
Materials
Linolenic acid (Sigma-Aldrich), Azocasein
(Sigma-Aldrich), AlarmaBlue cell viability assay kit (KeyGen
BioTECH, China), Tobramycin (Solarbio, China), Crystal violet
(Solarbio, China), LIVE/DEAD BacLight Bacterial viability kit
(Molecular Probes, USA), Biozol Total RNA extraction reagent
(BioFlux, China), TransScript Green Two-Step qRT-PCR SuperMix kit
(Transgen Biotech, China), Trans2 K DNA ladder (Transgen Biotech,
China), 2X Taq PCR MasterMix (TianGen, China), Ethidium bromide
(Sigma, China), Cetrimide agar base (Xiya, China), Dimethyl
sulfoxide (AMRESCO, Ohio-USA), and any other reagents were of
analytical grade.
Bacterial strains
Pseudomonas aeruginosa ATCC 27853 strain, and
Pseudomonas aeruginosa clinical strains were collected from
the Laboratory of Pathogen Biology, Experimental Teaching Center
for Basic Medical Sciences, Dalian Medical University while
environmental strains were isolated from beach soil, water pond
soil and garden soil in different locations on Dalian Medical
University campus.
Primer design
The quantitative real time polymerase chain reaction
(qRT-PCR) primers used in this study were designed with an online
Primer-Blast tool (https://www.ncbi.nlm.nih.gov/tools/primer-blast/;
accessed March 11, 2016) whereas identification primers for P.
aeruginosa were those published by Jaffe et al (32) (Table
I) and all were synthesized by Invitrogen Biotechnology Co.
Ltd, China.
 | Table I.Primer list for PCR and RT-qPCR. |
Table I.
Primer list for PCR and RT-qPCR.
| Target gene | Primer name | Sequence
(5′-3′) | Amplicon (bp) | (Refs.) |
|---|
| oprL (PA0973) | oprL-Forward | ATGGAA
ATGCTGAAATTCGGC | 504 | (32) |
|
| oprL-Reverse |
CTTCTTCAGCTCGACGCGACG |
|
|
| 16SrRNA |
16SrRNA-Forward |
GAGGAAGGTGGGGATGACGT | 233 | (32) |
|
|
16SrRNA-Reverse |
AGGCCCGGGAACGTATTCAC |
|
|
| lasI (PA1432) | LasI-Forward |
TGAAGCCCAGGTTTTCGGTT | 131 | This study |
|
| LasI-Reverse |
AACGGCTGAGTTCCCAGATG |
|
|
| lasR (PA1430) | LasR-Forward |
TCGAACATCCGGTCAGCAAA | 128 | This study |
|
| LasR-Reverse |
GTTCACATTGGCTTCCGAGC |
|
|
| rhlI (PA3476) | RhlI-Forward |
CATCCGCAAACCCGCTACAT | 124 | This study |
|
| RhlI-Reverse |
GGGTTTCGCTGCACAGGTA |
|
|
| rhlR (PA3477) | RhlR-Forward |
GAAATGGTGGTCTGGAGCGA | 132 | This study |
|
| RhlR-Reverse |
GGAAAGCACGCTGAGCAAAT |
|
|
| pilJ (PA0411) | pilJ-Forward |
GACAAGCAGTACATCGGCCA | 137 | This study |
|
| pilJ-Reverse |
CGTTTCTCGAAGTCGTTGCG |
|
|
| algU (PA0762) | algU-Forward |
CCATCAACACCGCGAAGAAC | 148 | This study |
|
| algU-Reverse |
ATCTCATCCCGCAACATCGC |
|
|
| pqsH (PA2587) | pqsH-Forward |
AGACGCTGATCCTGTTCCAG | 131 | This study |
|
| pqsH-Reverse |
GCGAACGAGGGTATTCCTCA |
|
|
| mvfR (PA1003) | mvfR-Forward |
TCGTTCTGCGATACGGTGAG | 136 | This study |
|
| mvfR-Reverse |
CGTCGATGGTGATGGCGATA |
|
|
| oprF (PA1777) | oprF-Forward |
CAGTACCCGTCCACTTCCAC | 146 | This study |
|
| oprF-Reverse |
TTCACGCGACCACCTTCTAC |
|
|
| oprI (PA2853) | oprI-Forward |
AGAAACCGAAGCTCGTCTGA | 137 | This study |
|
| oprI-Reverse |
CGTTAGCCTCGTCAGCAGT |
|
|
| phzR/qscR
(PA1898) | phzR-Forward |
GCTGACCGCGCCTAAATATC | 134 | This study |
|
| phzR-Reverse |
TCCAGATCAGCGGGGTGTAT |
|
|
| lasB (PA3724) | lasB-Forward |
TGTCCAAACTCCCCAGCAAG | 149 | This study |
|
| lasB-Reverse |
GAATTGCTCGTAGCGGGTGA |
|
|
Culture and identification of bacteria
strains
Clinical isolates of P. aeruginosa and the
ATCC strain were inoculated on cetrimide agar and incubated at 37°C
for 24 h. For environmental strains, 1 g of soil was dissolved in 1
ml PBS, pH 7.4 with vigorous shaking and centrifuged at 5,000 g for
one minute then 0.1 ml was inoculated on cetrimide agar and
incubated at 37°C for 24 h. A bacterial suspension was prepared by
dissolving a single colony from an overnight culture plate into 50
µl PBS pH 7.4 from which, a) 30 µl was added to a 15 ml-tube
containing 3 ml fresh LB media for overnight culture at 37°C with
180 rpm shaking, then stored in 8% (v/v) glycerol at −80°C for use
in subsequent assays; b) 1 µl was used for identification with
duplex colony polymerase chain reaction (PCR).
Duplex colony PCR
The PCR conditions were performed according to Jaffe
et al (32) with
modifications. Briefly, a 10 µl reaction mixture was prepared to
consist of 1 µl bacteria suspension, 0.5 µM forward and reverse
primers apiece, 5 µl of 2X Taq PCR MasterMix, 5% Dimethyl sulfoxide
(DMSO) and 1.5 µl distilled water to bring the total volume to 10
µl. DMSO was included to improve primer binding specificity during
amplification since Pseudomonas species have a high GC
content. The thermal cycling conditions were set as follows:
Initial denaturation at 94°C for 3 min, a 30 cycle involving
denaturation at 94°C for 30 sec, annealing at 58°C for 30 sec and
extension at 72°C for 30 sec, and a final extension at 72°C for 45
sec. After thermocycling, 10 µl was then loaded into a 1% (w/v)
agarose gel and the electrophoresis was run for 40 min at 70 volts
in 1X Tris-acetate-EDTA (TAE) Buffer mixed with 0.2 mM ethidium
bromide, then the images were taken by ChemiDoc XRS+
machine (Bio-ad).
Minimum inhibition concentration
determination
The minimum inhibition concentration (MIC) of
linolenic acid (LNA) and tobramycin (TOB) were performed following
CLSI recommendation using the macrodilution (Tube) broth method and
broth microdilution method, respectively (33). The stock solution of LNA (100 mg/ml)
was prepared in DMSO and a surfactant Tween 20 (2% final
concentration) was added for the uniform distribution of LNA and
two-fold serial dilutions were made. Firstly, an overnight P.
aeruginosa culture was diluted to an optical density (OD) of
0.1 at 600 nm (Multiskan Ascent Microplate Reader, Thermo
Scientific) with fresh LB broth (~108 CFU/ml) and 10 µl
of bacteria suspension (to achieve a final bacterial concentration
of ~106 CFU/ml) was added to the test tubes containing 1
ml media with serial dilutions of LNA, incubated at 37°C for 24 h
with gentle shaking at 110 rpm. Similarly, a two-fold dilutions of
TOB from 20 mg/ml to 0.0391 mg/ml were prepared in a 96-well plate
and incubated at 37°C for 24 h. The highest dilution of LNA or TOB
showing visible inhibition of bacterial growth after 24 h of
incubation was taken as MIC of the drug. Three independent assays
were performed.
Growth curve analysis
To understand the activity of LNA alone or in
combination with TOB on the P. aeruginosa, sub-MIC
concentrations of both LNA and TOB were selected for this study and
were assessed through the growth curve analysis as described by
Kalia et al (34). Briefly,
overnight bacteria culture was inoculated into 10 ml of LB broth
supplemented with five different sub-MIC concentrations of LNA
(i.e. 3/4, 5/8, 1/2, 3/8 and 1/4 of MIC) and TOB (1/4 of
MIC). The flasks were incubated at 37°C and OD600 was
monitored at time intervals 0, 3, 6, 9, 12, 18 and 24 h.
Biofilm control assay
As described elsewhere (34,35),
P. aeruginosa biofilm inhibition and biofilm metabolic
activity quantification were assessed in the presence/absence of
LNA, TOB or in combination (LNA+TOB). Briefly, one microliter of
overnight bacteria suspension adjusted to OD600 of 0.1
(~108 CFU/ml) was added to a well in a microtiter plate
containing 100 µl of fresh LB media mixed with appropriate doses of
drugs, and this plate was incubated at 37°C for 24 h from which
subsequent assays were performed as follows:
Biofilm mass quantification
After incubation, the unattached cells were gently
aspirated and discarded, and the wells were washed with 0.85%
sodium chloride (normal saline) twice and stained with 200 µl of
0.1% crystal violet for 15 min. The stain was discarded, and the
wells were washed with distilled water, air dried and the dye was
re-solubilized with 160 µl of 33% (v/v) glacial acetic acid
(36). The OD was measured at 570 nm
using Multiskan Ascent Microplate Reader (Thermo Scientific).
Biofilm inhibition was given by:
%BI={(ODC–ODT)/ODC}x100.
where %BI is the percentage of biofilm inhibition,
ODC is the 570 nm absorbance value of untreated sample
and ODT is the 570 nm absorbance value of treated sample
(35).
Biofilm metabolic activity
quantification
In order to assess the bacterial activity of biofilm
cells, the alamarblue (7-hydroxy-3H-phenoxazin-3-one-10-oxide) cell
viability assay was applied as a commended, reliable and
reproducible assay for assessing biofilm susceptibility (35,37).
After incubation as mentioned above, the supernatant was removed
and the wells were gently washed twice with LB media. Then, 200 µl
fresh LB media containing 10% (v/v) alarmablue staining reagent was
added and incubated at 37°C for 5 h, as recommended by the
manufacturer (KeyGen Biotech, China). The absorbance was then
measured at 570 nm and Biofilm inactivation was given by:
%BI={(AC–AT)/AC}x100.
where %BI is the percentage of biofilm inactivation,
AC is the 570 nm absorbance value of the untreated
sample and AT is the 570 nm absorbance value of treated
sample (35).
Microscopic analysis
To visualize the effect of biofilm formation in the
presence/absence of sub-MIC doses of LNA, TOB or LNA+TOB,
microscopic analysis of P. aeruginosa clinical strain C2
biofilms was performed as described previously (11,38,39) with
modifications. Briefly, 10 µl of an overnight culture as described
above, was added to 1 ml LB media supplemented with appropriate
sub-MIC doses in a 24-well plate. Positive control wells contained
media supplemented with the greatest concentration of DMSO used
(0.75%) while the negative control wells contained media only (for
sterility check), and the plate was then incubated at 37°C for 24
h. Afterwards, the wells were gently washed with sterile normal
saline and 200 µl of staining solution in LB media containing 2.09
µM of Syto 9 and 12.5 µM of Propidium iodide (PI) using LIVE/DEAD
BacLight Bacterial Viability Kit L7012 was added to each well,
incubated at room temperature for 15 min in the dark and images
were captured with an inverted fluorescence microscope (Olympus
IX71).
Synergy analysis
The interaction of LNA with TOB was analyzed by a
checkerboard method in an 8×8 arrangement using a 96-well plate as
previously described (24). Briefly,
biofilms were allowed to build up for 24 h and then wells were
gently washed with sterile LB medium and later supplemented with LB
medium consisting of different combinations of LNA and TOB. The
combinations were done in such a way that a fixed dose of one agent
and increasing doses of the second agent was put in each column (or
row). Then incubated at 37°C for 24 h, washed gently and the
biofilm metabolic activity was assessed with alamarblue staining
reagent for assessing biofilm susceptibility (35,37). For
all the wells with combined drug concentrations, the sum of the
fractional inhibitory concentrations (FIC) index was calculated
according to the equation below:
FICindex=FICA+FICB=A/MICA+B/MICB.
where A and B are the MICs of LNA (A) and TOB (B) in
combination, MICA and MICB are the MICs of
LNA and TOB alone, FICA and FICB are the FICs
of LNA and TOB, respectively. The FIC index results were
interpreted as synergistic effect (FIC index ≤0.5), additive or
indifferent effect (FIC index >0.5 and ≤1) and antagonistic
effect (FIC index >1) (40).
Inhibition of virulence factors
mediated by QS
Swarming assay
The swarming ability of P. aeruginosa C2
strain was investigated in small 35×10 mm plates containing
swarming motility media 0.5% (w/v) Bacto agar, 0.5% (w/v) peptone,
0.2% (w/v) yeast extract and 1.0% (w/v) glucose (41). A 2.5 µl aliquot from an overnight
culture in the presence/absence of drug was spotted at the center
of the agar surface and the plates were incubated at 37°C for 24 h,
and later 2 h at room temperature (42) and the diameters were measured. Three
independent assays were performed.
Pyocyanin production
As described by Das et al (11) and Kalia et al (34), the pyocyanin production was assayed
by collecting supernatants from overnight cultures of P.
aeruginosa grown in the presence/absence of sub-MIC doses of
LNA, TOB and LNA+TOB at 37°C for 24 h. A 5 ml culture supernatant
was extracted with 3 ml of chloroform and then with 1 ml of 0.2 N
hydrochloric acid to produce an orange-yellow to a pink colored
solution which was measured at 520 nm.
LasA staphylolytic assay
In the presence/absence of LNA, TOB and LNA+TOB
drugs, the ability of P. aeruginosa culture supernatants to
lyse boiled Staphylococcus aureus cells were determined by
LasA protease activity (11). An
overnight culture of S. aureus cells (OD595 of
1.0) was centrifuged at 7,000 rpm for 3 min and the pellet was
suspended in 0.02 M Tris-HCl (pH 8.5) then boiled for 10 min and
diluted with the same buffer to an OD595nm of 0.8.
Diluted S. aureus suspension was added to each cell-free
culture supernatant of P. aeruginosa in the ratio 9:1 and
the absorbance was measured at 595 nm after 0, 15, 30, 45 and 60
min.
Azocasein protease assay
The assay was performed to study the effect of
protease production by C2 strain in the presence/absence of LNA,
TOB and LNA+TOB. This was performed following methods published
elsewhere with minor modifications (11,34).
Briefly, cell-free supernatant was collected from centrifuged
overnight cultures in the presence/absence of drugs at 10,000 rpm
for 5 min. 150 µl of supernatant (in absence/presence of the drug)
was added to 1 ml of 0.3% azocasein in 0.05 M Tris-HCl (pH 7.5) and
incubated at 37°C for 15 min. The reaction was then stopped with
0.5 ml 10% trichloroacetic acid and the mixture was later
centrifuged at 10,000 rpm for 5 min. The clear supernatant was
collected and absorbance was measured at 420 nm.
Quantitative real-time polymerase chain reaction
(qRT-PCR)
The real-time PCR reactions for quantification of
Pseudomonas aeruginosa quorum sensing target genes (lasI,
lasR, rhlI, rhlR,), virulence factor-related genes, and the
reference gene (16S rRNA) were performed on ABI StepOne
analyser using TransScript Green Two-Step qRT-PCR SuperMix kit
following the manufacturer's instructions (Transgen Biotech, China)
with the primers listed in Table I.
Firstly, P. aeruginosa was cultured in the absence/presence
of sub-MIC concentrations of LNA, TOB and LNA+TOB for 24 h at 37°C.
Total RNA was then extracted using biozol total RNA extraction
reagent following the manufacturer's instructions (BioFlux, China)
and the concentration was measured with NanoDrop 2000C (Thermo
Scientific). One microgram of the respective extracted RNA was used
for cDNA synthesis using TransScript Green Two-Step qRT-PCR
SuperMix kit as per manufacturer's instructions. Secondly, the
synthesized cDNA was diluted ten-fold (1:10 ratio) and one
microliter of diluted cDNA was mixed with 0.2 µM forward and
reverse primers apiece, 10 µl of 2X TransStart Tip Green qPCR
SuperMix, 1X passive reference dye I and water to a total volume of
20 µl as per instructions. The dissociation stage consisted of 94°C
for 30 sec, followed by 40 cycles involving 94°C for 5 sec, 58°C
for 15 sec, and 72°C for 10 sec. Target gene primers were designed
to give products between 120 and 150 bp, and the 16SrRNA
gene was used as an internal control. Each qRT-PCR run was
performed in triplicate and three independent experiments were
performed. The calculated threshold cycle (Ct) was normalized to
the Ct of 16SrRNA amplified from the corresponding sample.
Finally, the relative quantification of target transcripts were
calculated by the comparative 2−ΔΔCt method (43).
Statistical analysis
To gain statistical significance, each experiment
was performed at least in triplicate. Data values of experimental
results were recorded as mean ± SEM or median with interquartile
range. Significance was determined by using analysis of variance
(one- and two-way), and cited as P<0.05 (*), P<0.01 (**) and
P<0.001 (***). Statistical analyses were performed using
GraphPad Prism 5.0 statistical software.
Results
Antimicrobial and anti-biofilm effects
of linolenic acid and tobramycin on Pseudomonas aeruginosa
isolates
The present study examined five clinical strains,
five environmental strains, and one standard (ATCC 27853) strain.
The clinical and environmental strains were identified by duplex
colony PCR using intragenic primer sets for bacterial 16S
rRNA and the peptidoglycan-associated lipoprotein (oprL:
PA0973) gene sequences for specific detection of P.
aeruginosa (data not shown).
Several reports suggest that FA of the omega-3
family are effective on various pathogenic microorganisms (22,23,25,26,44). We
therefore attempted to assess the antimicrobial activity of LNA on
P. aeruginosa ATCC 27853, clinical and environmental
strains. The median MIC values of LNA and TOB were 1.56 and 0.3125
mg/ml, respectively. Due to this anti-pseudomonas activity, we
hypothesized that LNA can affect biofilm formation when used alone
or in combination with TOB. Therefore, five sub-MIC doses of LNA
(1.17, 0.975, 0.78, 0.585 & 0.39 mg/ml), and 1/4th MIC (0.078
mg/ml) of TOB were selected for this study and their activity on
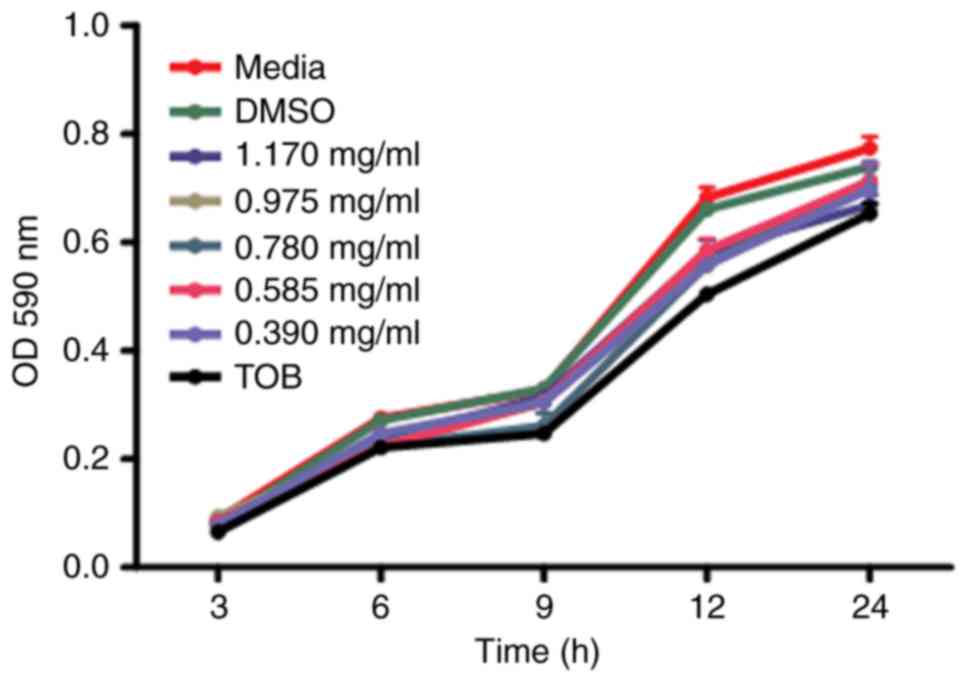
planktonic cells were assessed through the growth curve analysis.
The growth curve results showed nonsignificant results on
planktonic cells (Fig. 1), but we
went on to evaluate the anti-biofilm activities.
Interestingly, the ATCC strain was highly sensitive
to TOB and LNA+TOB treatment in comparison with clinical and
environmental stains (Fig. 2). An
identical dose dependant pattern was exhibited in LNA+TOB and LNA
groups with their respective highest median inhibitory percentage
being 48 and 38% (Fig. 2A). However,
clinical strain C2 was noticed with extreme resistance to TOB
dosage used while the rest of the strains were significantly
affected (P<0.001, Fig. 2B). The
moderate inhibitory effect of LNA alone or in combination (Fig. 2A) provoked further analysis of
biofilm cells. We attempted to assess the metabolic effect with
alarmablue staining reagent to quantify the viability of cells
(37). The assay revealed a
significant inactivation of biofilm cells (P<0.001, Fig. 2C); 60% inactivation with LNA+TOB
involving 1.17 mg/ml dosage, 50% for LNA (1.17 mg/ml) alone and 42%
for TOB alone (Fig. 2C). The slight
difference between the two assays (inhibition and inactivated) was
probably due to their different staining principles. Unlike
alarmablue reagent, crystal violet staining does not differentiate
live cells from dead cells but stains every attached biofilm cell
whether live, about to die or dead ones, thus giving a slight
difference between the two assays. After analyzing the inhibition
and inactivation effects, P. aeruginosa clinical strain C2
was seen with more resistance especially with TOB (Fig. 2B and D). Therefore, we hypothesized
that understanding the anti-biofilm action of LNA alone or in
combination treatment could be better in a strain where TOB is
exerting an insignificant effect. Moreover, the ATCC strain very
sensitive to the selected sub-MIC doses that further dilution was
required to re-establish its MIC doses (data not shown). So,
clinical strain C2 was selected for microscopic evaluation,
virulence factor production and gene expression with qRT-PCR.
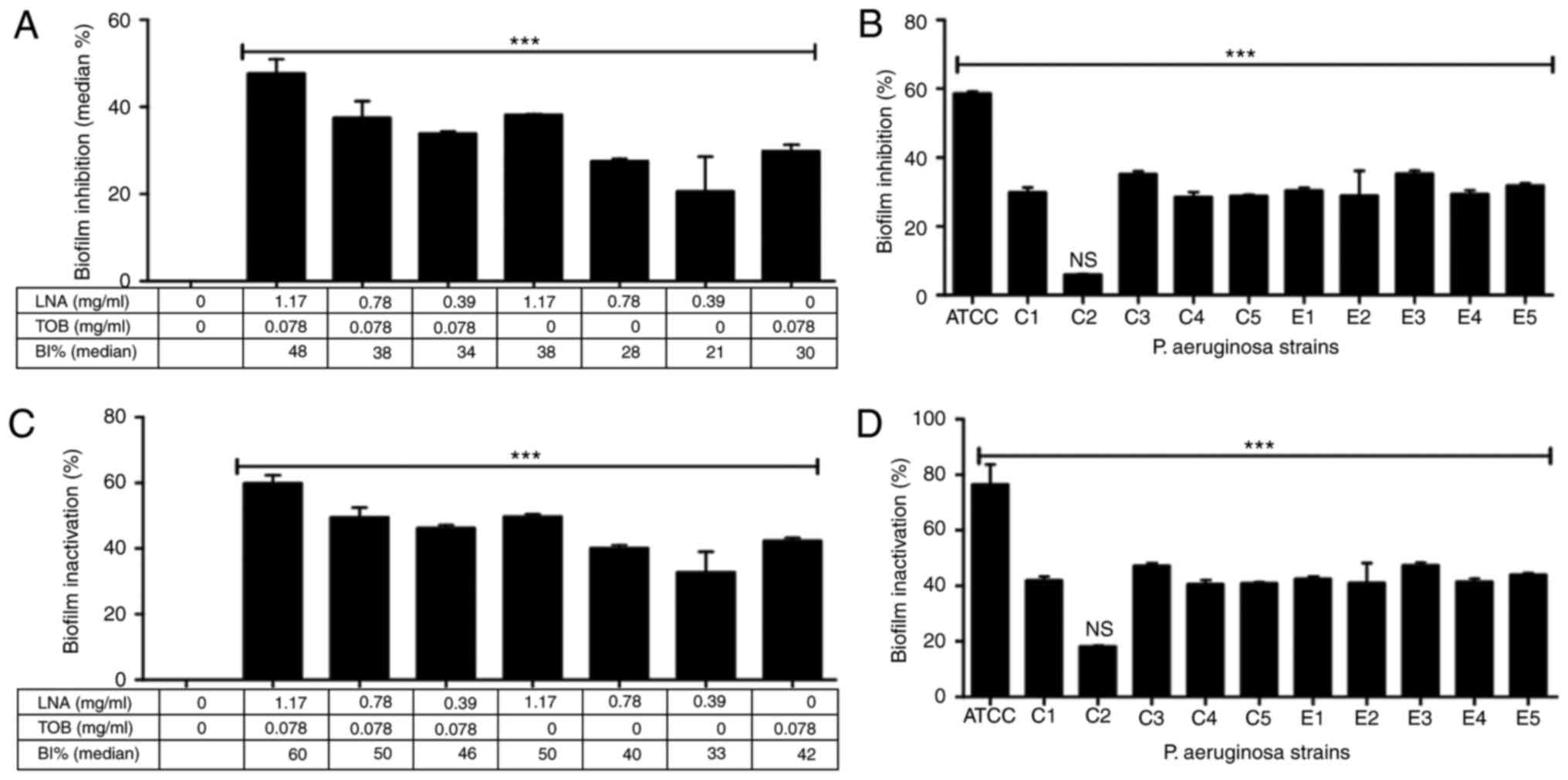 | Figure 2.Biofilm inhibition and inactivation
effects of linolenic acid, tobramycin, and LNA+TOB on P.
aeruginosa strains. (A) Median inhibition percentages of all
strains with respect to sub-MIC doses, (B) the inhibitory effect of
TOB alone, (C) the median inactivation percentages of all strains
with respect to sub-MIC doses, and (D) the inactivation effect of
TOB alone are presented. The data are presented as median with
interquartile ranges. LNA, linolenic acid; TOB, tobramycin; ATCC,
ATCC 27853 strain; C, clinical strains; E, environmental strain;
and ***P<0.001. Three independent assays were performed. |
Microscopic visualization was performed with a
fluorescence microscope (Olympus IX71). Images of the untreated
control showed robust biofilm biomass but slightly reduced in TOB
treated cells. LNA treated cells managed to reduce the biofilm
formation by disintegrating the biomass while combination treatment
exhibited strong inhibitory activity (Fig. 3). This effect reduced significantly
in a dose-dependent manner in combined treatment groups. The
findings here were in agreement with the biofilm inhibition
(Fig. 2A) and inactivation (Fig. 2C) assays which also showed similar
effects with reference to drug concentration used. Moreover, the
interaction effect between LNA and TOB determined by the FIC index
showed additive and synergistic effects on C2 strain (Table II), which could explain the strong
attenuation effects of biofilm in combined treatment with reference
to untreated control cells.
| Table II.Interaction study between LNA and
TOB. |
Table II.
Interaction study between LNA and
TOB.
| P. aeruginosa
strain | Combined doses
(LNA+TOB) (mg/ml) | FIC index | Interpretation |
|---|
| C2 | 1.17+0.078 | 1.00 | Additive |
| C2 | 0.78+0.078 | 0.75 | Additive |
| C2 | 0.39+0.078 | 0.50 | Synergy |
Effect of linolenic acid and
tobramycin on virulence factor production
P. aeruginosa is known to produce virulence
factors that include pyocyanin and protease production that tends
to counteract host defenses and can directly damage host tissues
(45). Since virulence factor
production and biofilm formation are controlled by the QS systems,
we hypothesized that LNA alone or in combination with TOB can
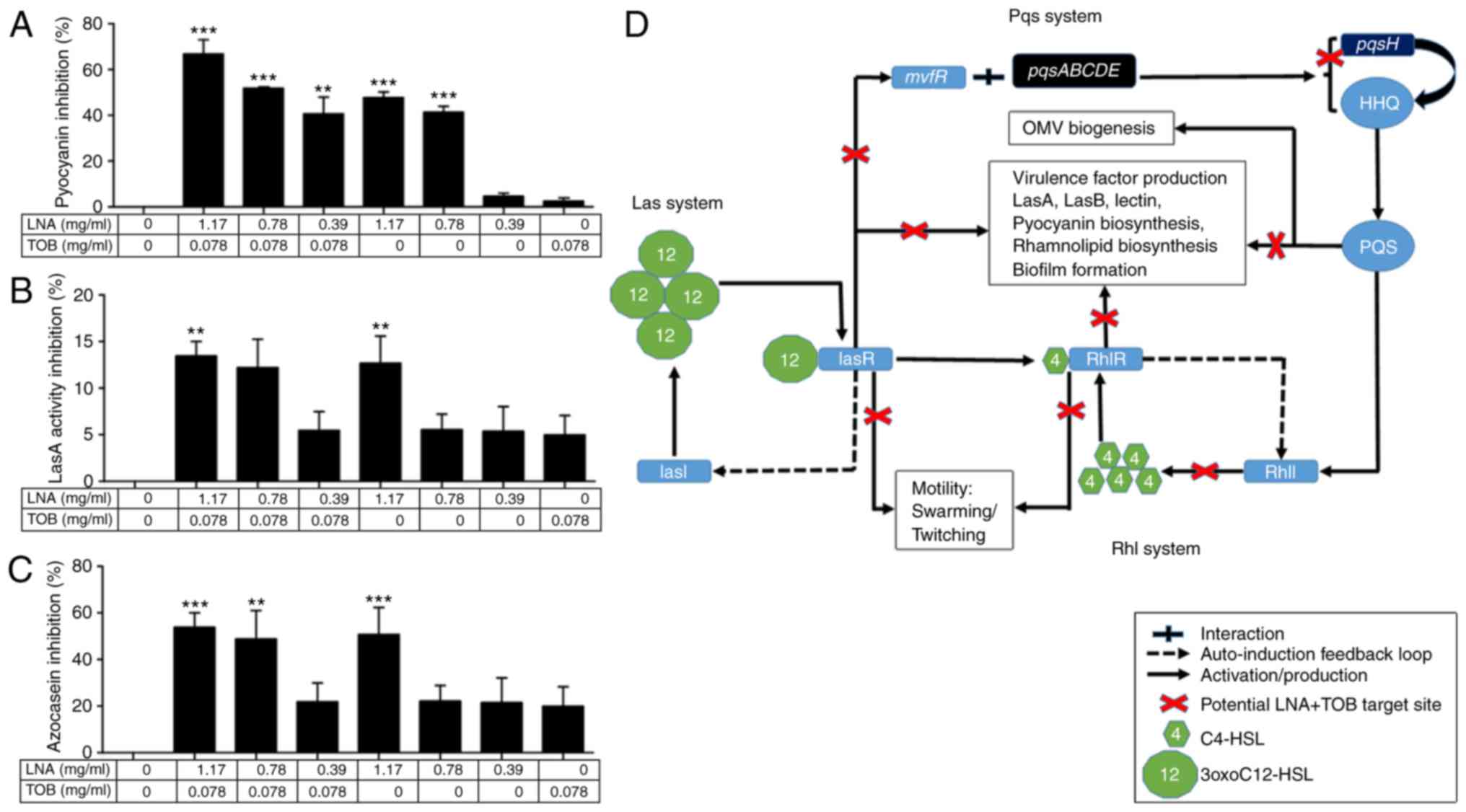
attenuate virulent factor production. So we assessed swarming
motility, pyocyanin production, lasA and azocasein activities on
P. aeruginosa strain C2.
Swarming motility
Swarming motility is the ability of bacteria to move
across a semisolid surface and P. aeruginosa uses a
flagellar motility (42) and
participates in increasing production of virulence factors and
antibiotic resistance (46). We
found that the treated group in comparison with the untreated
control inhibited the swarming movement of P. aeruginosa C2
strain (Fig. 4). Interestingly, all
the sub-MIC doses used in combination and single treatment showed
significant inhibition effect on swarming motility with reference
to the untreated control cells of P. aeruginosa C2 strain
(P<0.001, Fig. 4).
Virulence factor production
P. aeruginosa virulence and pathogenesis
include secretion of virulence factors such as pyocyanin and
pyoverdine. Pyocyanin disrupts the redox system and electron
transport pathways of a host cell whereas pyoverdine is a
siderophore that captures iron usually from iron-binding proteins
such as ferritin, lactoferrin, and transferrin (45,47), and
are important for virulence and biofilm formation (48). On the other hand, LasA protease
possesses a high staphylolytic activity on cleaving the peptide
bonds of pentaglycine bridges within the peptidoglycan of S.
aureus cells and enhances the activity of LasB elastase in
degrading the Gly-Gly peptide bonds in elastin, a component of
connective tissue, blood vessels, and lung tissue (49). The effect of these virulence factors
were examined in the presence/absence of sub-MIC doses of the
drugs. The activity of LNA+TOB was more effective than single dose
treatments (Fig. 5A-C). Therefore,
combining the phenotypic findings and genotypic expression pattern,
we presented the schematic diagram (Fig.
5D) illustrating the suggested targeted sites of LNA+TOB in the
QS system.
The anti-biofilm mechanism of
linolenic acid, tobramycin, and LNA+TOB on P. aeruginosa C2
strain
To analyze the effects of QS genes and virulence
factor-related gene expressions by P. aeruginosa C2 strain
in the presence/absence of sub-MIC doses of the drugs, quantitative
real-time PCR (qRT-PCR) was performed to assess the relative
expression of our target genes (lasI, lasR, rhlI, rhlR, pqsH,
mvfR, oprF, oprI, pilJ, algU, phzR/qscR, and lasB). For
this analysis, 0.39 mg/ml (LNA) and 0.078 mg/ml (TOB) were used in
single and combined treatment on C2 strain because these
concentrations exhibited synergism in the interaction study as
shown by the FIC index (0.5; Table
II).
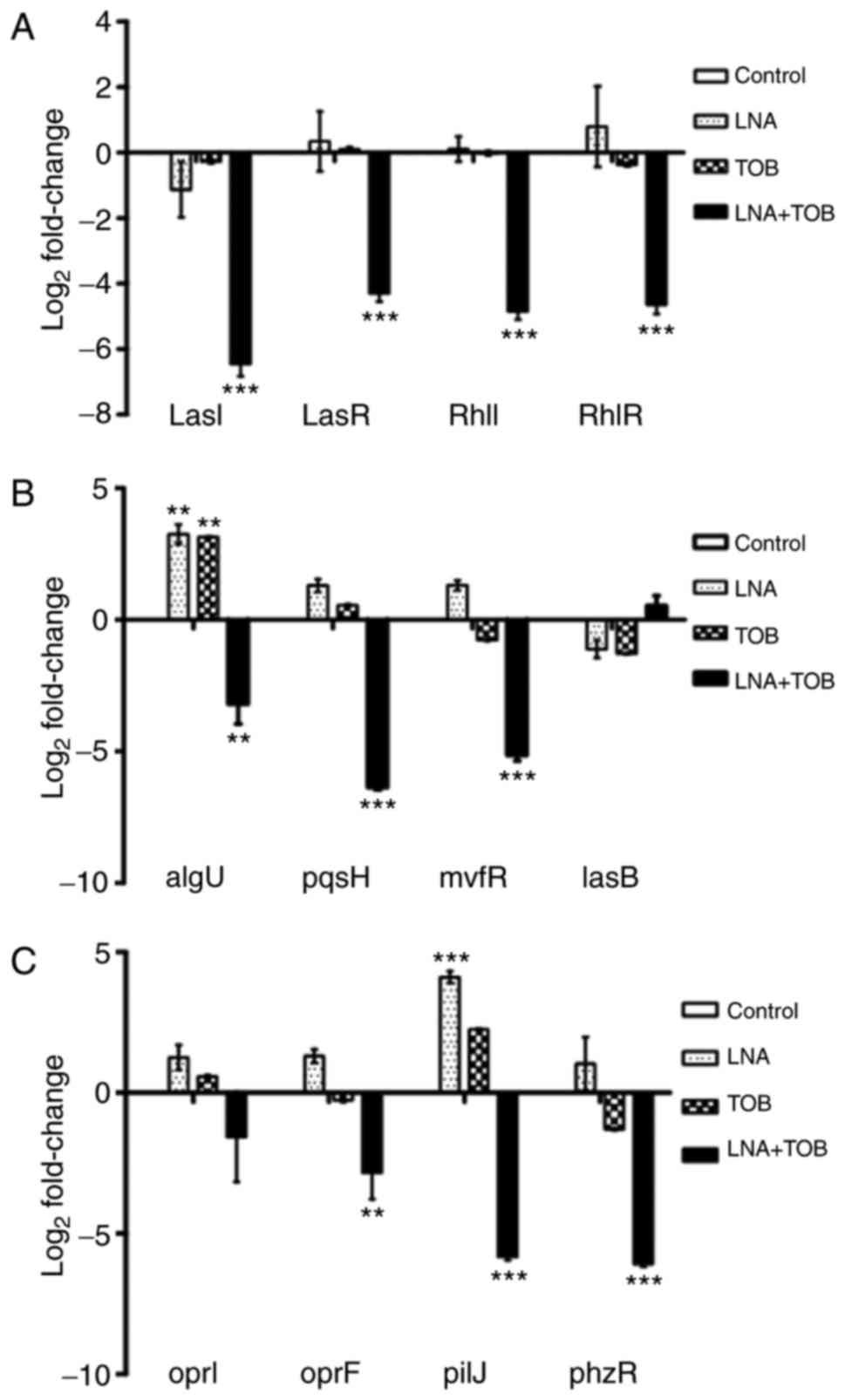
Quantitative RT-PCR results showed nonsignificant
effects of both LNA and TOB on QS genes but their expressions
reduced by −91fold (lasI; PA1432), −20 fold (lasR;
PA1430), −29 fold (rhlI; PA3476) and −25 fold (rhlR;
PA3477) due to LNA+TOB treatment (P<0.001; Fig. 6A). These genes are the major
regulators of Las system (lasI/lasR) and Rhl system (rhlI/rhlR). In
spite of these two systems, the bacterium uses another system, the
Pseudomonas quinolone signal system. For this system, the activated
lasR expresses mvfR that interacts with pqsABCDE loci to
control the production of pqsH and 2-heptyl-4-quinolone
(HHQ), and the latter is converted into
2-heptyl-3-hydroxy-4-quinolone (PQS) by pqsH (50), as depicted in Fig. 5D. The product, PQS regulates various
virulence factor related genes including elastase and pyocyanin
coding genes (50). Also, evidence
suggests that alterations in mvfR expression disrupt PQS
synthesis and pyocyanin production (51,52). In
addition, mvfR mutant was shown to reduce virulence in an
animal model (53). In the present
study, we observed that the expression of pqsH (PA2587;
P<0.001) and mvfR (PA1003; P<0.001) were downregulated
by LNA+TOB whereas the cells treated with LNA or TOB showed no
statistical significance (Fig. 6B).
This suggested that LNA had no influence on the expression of our
selected genes belonging to the three interrelated QS systems.
However, due to the anti-biofilm effects that were exhibited in
treatment groups (Fig. 2), we
thought to assess the expression levels of oprI (PA2853),
oprF (PA1777) and algU (PA0762) genes. OprF is an
outer membrane protein that plays a role in maintaining the cell
shape and contribute to P. aeruginosa virulence (54,55). The
counterpart outer membrane protein oprI also play a part in
membrane integrity and normal cell shape (56). A study by Fito-Boncompte et al
(55) reported that the absence of
OprF disturbs bacterial cell adhesion to animal cells, release of
ExoT and ExoS toxins through the type III secretion system (T3SS),
and production of the QS-associated virulence factors such as
lectin PA-1 L, elastase, pyocyanin, and exotoxin A. In addition,
there was reduction and retardation in production of the signalling
molecules 3oxoC12HSL and C4HSL respectively in the oprF mutant. In
our study, LNA+TOB reduced the expression of oprF
(P<0.01) but not the outer membrane lipoprotein precursor
(oprI) as compared to the untreated control (Fig. 6C). Moreover, algU is involved
in regulating alginate production and inactivation of this gene
increased the susceptibility of P. aeruginosa PAO1 killing
by chemically generated reactive oxygen species, J774 murine
macrophages and human neutrophils (57). Moreover, algU regulates rsmA
expression, a posttranscriptional regulator involved in virulence
factor production and biofilm formation (58). Interestingly, algU was
downregulated by LNA+TOB (P<0.01) but upregulated in LNA or TOB
treated groups with similar statistical magnitude (Fig. 6B). Furthermore, we selectively
decided to assess the expression levels of pilJ (PA0411),
phzR/qscR (PA1898) and lasB (PA3724) that contribute
to bacterial virulence. We observed the downregulation of
pilJ (PA0411; P<0.001) and phzR/qscR (PA1898;
P<0.001) genes in LNA+TOB treated cells while LNA alone
upregulated the pilJ (P<0.001) expression (Fig. 6C). Surprisingly, no statistically
significant effect was noticed on lasB gene expression in
all treatment groups as compared to the untreated control.
Discussion
Pseudomonas aeruginosa is a medically
important pathogen that causes nosocomial and serious life
threating infections due to its ability to produce biofilm that has
contributed to the emergence of drug resistance towards currently
utilized antibiotics. Considering the antimicrobial and
anti-inflammatory properties of fatty acids (17,18),
assessing their biofilm prevention properties on P.
aeruginosa may supplement in finding alternative agents for
antimicrobial resistance problem. Moreover, evidence suggest that
LNA and its derivatives, EPA and DHA possess antimicrobial
properties against various pathogenic microorganisms (22–26).
Biofilm formation is an active process that is
dependent highly on environmental signals which sensitize a cell to
undergo stages of cycle growth. This process involves production of
a matrix comprising of polysaccharides, proteins, lipids and
extracellular DNA (eDNA) that provide a physiological barrier to
antimicrobial diffusion and concentrate secreted extracellular
enzymes such as β-lactamases, whereas the limitation of oxygen and
nutrients support the anaerobic growth of P. aeruginosa
which decelerates cell division (2,10,59).
During this stage, the bacterial community develops resistance to
various antimicrobial agents, like β-lactam and aminoglycoside
antibiotics that target actively and aerobically growing bacterial
cells, respectively (59). Our study
has shown that LNA, indeed possesses antibacterial and anti-biofilm
effect against P. aeruginosa as evidenced by the MIC and
biofilm inhibition/inactivation assays. We had observed that LNA
alone and in combination with TOB had the ability to disrupt
biofilm formation and decrease the metabolic activities of biofilm
cells by causing cell death in a dose-dependent manner (Figs. 2 and 3). Besides, the stronger effects exhibited
by the combined treatment suggested that LNA did not antagonize the
activity of TOB but synergistically and additively exerted their
combined effects on biofilm as supported by their FIC index results
(Table II).
Furthermore, quorum sensing is a major player in
P. aeruginosa virulence factor production and biofilm
formation, making it an interesting target for an antipseudomonal
compound search. Evidence has shown that inhibiting P.
aeruginosa QS system disrupts biofilm formation and virulence
factor production (34,60). Our study showed that LNA+TOB had a
great impact in downregulating the genes associated with the three
interrelated QS systems (Las, Rhl and Pqs systems) (Fig. 6A and B). Since the bacterium uses QS
systems to control biofilm formation and virulence factor
secretion, attempts were made to analyze some virulence factor
associated genes. P. aeruginosa produces outer membrane
vesicles (OMV) as a tool for genetic information and toxins
dissemination (61), and they
require the signaling molecule PQS for their biogenesis which
accumulates in the LPS-rich outer leaflet of the outer membrane and
causes bleb formation (62).
Previous evidence suggests that outer membrane proteins oprI and
oprF participate in OMV formation but the absence of these proteins
and presence of PQS still increased OMV production by P.
aeruginosa (63). This implies
that PQS play a major role in OMV biogenesis making it an
attractive target for anti-biofilm exploration. We have shown that
LNA+TOB is capable of interfering with OMV biogenesis at gene
transcription level (Fig. 6B and C).
On the other hand, alginate is associated with chronic lung
infection though only secreted by a subsection of P.
aeruginosa species, as most strains can either secrete Psl
(polysaccharide synthesis locus) or Pel (pectate lyase)
polysaccharides (64,65). Our study has shown that LNA+TOB is
effective in disrupting alginate production (Fig. 6B). This suggests that LNA+TOB can be
effective in reducing chronicity resulting from alginate production
in lung infections. Moreover, we have shown that LNA+TOB treatment
can prevent swarming and twitching motility thereby reducing the
spread of infection. Therefore, we can deduce that LNA+TOB is more
effective in preventing biofilm formation and virulence factor
production than single LNA or TOB in targeting the QS system of
P. aeruginosa. This could be due to the additive and
synergistic effects that may occur once the two compounds are
combined.
Although tobramycin is currently used in cystic
fibrosis lung infections, it fails to clear chronic infection
associated with P. aeruginosa completely (66). However, various compounds combined
with tobramycin had shown synergism in attenuating biofilm
formation by this pathogen (67,68). Our
study revealed that LNA can improved the efficacy of TOB and could
be considered in P. aeruginosa related infections. In spite
of the differences in the genotypic and phenotypic assays'
outcomes, it is no doubt that LNA is capable of reducing biofilm
formation and virulence factor production as shown from our
phenotypic analyses such as biofilm, swarming, phenazine, and
proteases for being affected by LNA alone. However, the mechanism
is definitely not associated with QS system but perhaps utilizing a
different route to cause biofilm inhibition. For that, we
hypothesized that LNA could be interacting with the cell membrane,
increasing the membrane fluidity thereby disrupting its
permeability causing the cell to leak out as well as transporting
TOB in biofilm cells (17,69), but substantialized analysis is
needed. On the other hand, diet supplementation of omega-3 PUFA
have been shown to improve the cardiovascular and respiratory
conditions (19,70), and was reported to be safe for human
use (71), suggesting that LNA
supplementation may serve as an immunomodulatory and antibiotic
agent.
In conclusion, the present study demonstrates that
linolenic acid has the capacity to interfere with P.
aeruginosa biofilm formation and virulence factor production
though experimented on the C2 strain. This strain exhibited
stronger resistance to the selected doses than any other strain
used in this study. It is therefore not acceptable to generalize
the findings of one strain with the rest. However, it is imperative
to note that these findings have shown the effectiveness of a
combination therapy (LNA+TOB), which should stimulate critical
analysis for antimicrobial consideration. Moreover, the combination
therapy has shown strong synergism in disrupting the P.
aeruginosa quorum sensing systems and decreasing its virulence
factor production. Therefore, it can be deduced that linolenic acid
used alone can attenuate the formation of biofilm and can improve
the action of tobramycin in targeting the quorum sensing systems.
This may decrease tobramycin concentration when in fact treated in
combination with linolenic acid (in the form of dietary
supplements) thereby reducing the adverse effects of
aminoglycosides. However, this study is only based on in
vitro investigations that may not mimic the clinical settings.
Thus, the findings here may not be conclusive but stimulate
interest in considering such compounds for adjunctive therapy.
Hence more studies are required, especially in vivo studies
with model systems that can simulate the clinical settings to
substantiate our findings.
Acknowledgements
We would like to convey our appreciation to Mr.
Richardson Joseph (Ph.D. Fellow), Mr. Emeka Okoye (Ph.D. Fellow)
and Dr. Brian Ayuka (MD) for reviewing this manuscript. We would
like to also thank the Chinese Scholarship Council for their
financial support.
References
|
1
|
Bai AJ and Rai VR: Quorum-Sensing Systems
in PseudomonasQuorum Sensing vs Quorum Quenching: A Battle
with No End in Sight. Kalia VC: Springer; India: pp. 73–84.
2015
|
|
2
|
Sharma G, Rao S, Bansal A, Dang S, Gupta S
and Gabrani R: Pseudomonas aeruginosa biofilm: Potential
therapeutic targets. Biologicals. 42:1–7. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Takajo D, Iwaya K, Katsurada Y, Miyai K,
Takasu A, Matsubara O, Sakamoto T, Tamai S and Tsuda H:
Community-acquired lobar pneumonia caused by Pseudomonas
aeruginosa infection in Japan: A case report with histological
and immunohistochemical examination. Pathol Int. 64:224–230. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Gharabaghi MA, Abdollahi SM, Safavi E and
Abtahi SH: Community acquired Pseudomonas pneumonia in an
immune competent host. BMJ Case Rep. 2012:pii: bcr0120125673. 2012.
View Article : Google Scholar
|
|
5
|
Stryjewski M and Sexton D: Pseudomonas
aeruginosa infections in specific types of patients and
clinical settingsSevere infections caused by Pseudomonas
aeruginosa. Hauser AR and Rello J: Kluwer Academic Publishers;
Boston: pp. 1–15. 2003, View Article : Google Scholar
|
|
6
|
Caron E, Desseyn JL, Sergent L, Bartke N,
Husson MO, Duhamel A and Gottrand F: Impact of fish oils on the
outcomes of a mouse model of acute Pseudomonas aeruginosa
pulmonary infection. Br J Nutr. 113:191–199. 2015. View Article : Google Scholar
|
|
7
|
Kollef MH, Chastre J, Fagon JY, François
B, Niederman MS, Rello J, Torres A, Vincent JL, Wunderink RG, Go KW
and Rehm C: Global prospective epidemiologic and surveillance study
of ventilator-associated pneumonia due to Pseudomonas
aeruginosa. Crit Care Med. 42:2178–2187. 2014. View Article : Google Scholar
|
|
8
|
Bodey GP, Bolivar R, Fainstein V and
Jadeja L: Infections caused by Pseudomonas aeruginosa. Rev
Infect Dis. 5:279–313. 1983. View Article : Google Scholar
|
|
9
|
CDC: Antibiotic Resistance Threats in the
United States, 2013. Journal. 69–70. 2013.
|
|
10
|
Breidenstein EB, de la Fuente-Núñez C and
Hancock RE: Pseudomonas aeruginosa: All roads lead to
resistance. Trends Microbiol. 19:419–426. 2011. View Article : Google Scholar
|
|
11
|
Das MC, Sandhu P, Gupta P, Rudrapaul P, De
UC, Tribedi P, Akhter Y and Bhattacharjee S: Attenuation of
Pseudomonas aeruginosa biofilm formation by Vitexin: A
combinatorial study with azithromycin and gentamicin. Sci Rep.
6:233472016. View Article : Google Scholar
|
|
12
|
Antunes LC, Ferreira RB, Buckner MM and
Finlay BB: Quorum sensing in bacterial virulence. Microbiology.
156:2271–2282. 2010. View Article : Google Scholar
|
|
13
|
Juhas M, Eberl L and Tümmler B: Quorum
sensing: The power of cooperation in the world of
Pseudomonas. Environ Microbiol. 7:459–471. 2005. View Article : Google Scholar
|
|
14
|
Winzer K, Falconer C, Garber NC, Diggle
SP, Camara M and Williams P: The Pseudomonas aeruginosa
lectins PA-IL and PA-IIL are controlled by quorum sensing and by
RpoS. J Bacteriol. 182:6401–6411. 2000. View Article : Google Scholar
|
|
15
|
Lyczak JB, Cannon CL and Pier GB:
Establishment of Pseudomonas aeruginosa infection: Lessons
from a versatile opportunist. Microbes Infect. 2:1051–1060. 2000.
View Article : Google Scholar
|
|
16
|
Hancock RE and Speert DP: Antibiotic
resistance in Pseudomonas aeruginosa. Mechanisms and impact
on treatment. Drug Resist Updat. 3:247–255. 2000. View Article : Google Scholar
|
|
17
|
Desbois AP and Smith VJ: Antibacterial
free fatty acids: Activities, mechanisms of action and
biotechnological potential. Appl Microbiol Biotechnol.
85:1629–1642. 2010. View Article : Google Scholar
|
|
18
|
Desbois AP: Potential applications of
antimicrobial fatty acids in medicine, agriculture and other
industries. Recent Pat Antiinfect Drug Discov. 7:111–122. 2012.
View Article : Google Scholar
|
|
19
|
Olveira G, Olveira C, Acosta E, Espíldora
F, Garrido-Sánchez L, García-Escobar E, Rojo-Martínez G, Gonzalo M
and Soriguer F: Fatty acid supplements improve respiratory,
inflammatory and nutritional parameters in adults with cystic
fibrosis. Arch Bronconeumol. 46:70–77. 2010.(In English, Spanish).
View Article : Google Scholar
|
|
20
|
Nieuwenhove CPV, Terán V and González
SN: Conjugated Linoleic and Linolenic Acid Production by Bacteria:
Development of Functional FoodsProbiotics. Rigobelo EC: INTECH;
2012
|
|
21
|
WHO and FAO joint consultation: fats and
oils in human nutrition. Nutr Rev. 53:202–205. 1995.
|
|
22
|
Sun M, Zhou Z, Dong J, Zhang J, Xia Y and
Shu R: Antibacterial and antibiofilm activities of docosahexaenoic
acid (DHA) and eicosapentaenoic acid (EPA) against periodontopathic
bacteria. Microb Pathog. 99:196–203. 2016. View Article : Google Scholar
|
|
23
|
Correia M, Michel V, Matos AA, Carvalho P,
Oliveira MJ, Ferreira RM, Dillies MA, Huerre M, Seruca R,
Figueiredo C, et al: Docosahexaenoic acid inhibits Helicobacter
pylori growth in vitro and mice gastric mucosa colonization.
PLoS One. 7:e350722012. View Article : Google Scholar
|
|
24
|
Huang CB, George B and Ebersole JL:
Antimicrobial activity of n-6, n-7 and n-9 fatty acids and their
esters for oral microorganisms. Arch Oral Biol. 55:555–560. 2010.
View Article : Google Scholar
|
|
25
|
Desbois AP and Lawlor KC: Antibacterial
activity of long-chain polyunsaturated fatty acids against
Propionibacterium acnes Staphylococcus aureus. Mar Drugs.
11:4544–4557. 2013. View Article : Google Scholar
|
|
26
|
Mil-Homens D, Bernardes N and Fialho AM:
The antibacterial properties of docosahexaenoic omega-3 fatty acid
against the cystic fibrosis multiresistant pathogen Burkholderia
cenocepacia. FEMS Microbiol Lett. 328:61–69. 2012. View Article : Google Scholar
|
|
27
|
Desbois AP, Mearns-Spragg A and Smith VJ:
A fatty acid from the diatom Phaeodactylum tricornutum is
antibacterial against diverse bacteria including multi-resistant
Staphylococcus aureus (MRSA). Mar Biotechnol (NY). 11:45–52.
2009. View Article : Google Scholar
|
|
28
|
Kaushik KS, Stolhandske J, Shindell O,
Smyth HD and Gordon VD: Tobramycin and bicarbonate synergise to
kill planktonic Pseudomonas aeruginosa, but antagonise to
promote biofilm survival. NPJ Biofilms Microbiomes. 2:160062016.
View Article : Google Scholar
|
|
29
|
Zobell JT, Young DC, Waters CD, Ampofo K,
Stockmann C, Sherwin CM and Spigarelli MG: Optimization of
anti-pseudomonal antibiotics for cystic fibrosis pulmonary
exacerbations: VI. Executive summary. Pediatr Pulmonol. 48:525–537.
2013. View Article : Google Scholar
|
|
30
|
Hirsch EB and Tam VH: Impact of
multidrug-resistant Pseudomonas aeruginosa infection on
patient outcomes. Expert Rev Pharmacoecon Outcomes Res. 10:441–451.
2010. View Article : Google Scholar
|
|
31
|
Morais-Braga MF, Souza TM, Santos KK,
Guedes GM, Andrade JC, Tintino SR, Sobral-Souza CE, Costa JG,
Saraiva AA and Coutinho HD: Phenolic compounds and interaction
between aminoglycosides and natural products of Lygodium
venustum SW against multiresistant bacteria. Chemotherapy.
58:337–340. 2012. View Article : Google Scholar
|
|
32
|
Jaffe RI, Lane JD and Bates CW: Real-time
identification of Pseudomonas aeruginosa direct from
clinical samples using a rapid extraction method and polymerase
chain reaction (PCR). J Clin Lab Anal. 15:131–137. 2001. View Article : Google Scholar
|
|
33
|
CLSI: Methods for Dilution Antimicrobial
Susceptibility Tests for Bacteria That Grow Aerobically; Approved
Standard. 9th edition. CLSI document M07-A9. 29:17–19. 2012.
|
|
34
|
Kalia M, Yadav VK, Singh PK, Sharma D,
Pandey H, Narvi SS and Agarwal V: Effect of cinnamon oil on quorum
sensing-controlled virulence factors and biofilm formation in
Pseudomonas aeruginosa. PLoS One. 10:e01354952015.
View Article : Google Scholar
|
|
35
|
Borges A, Simões LC, Saavedra MJ and
Simões M: The action of selected isothiocyanates on bacterial
biofilm prevention and control. Int Biodeterior Biodegrad.
86:25–33. 2014. View Article : Google Scholar
|
|
36
|
Stepanovic S, Vukovic D, Dakic I, Savic B
and Svabic-Vlahovic M: A modified microtiter-plate test for
quantification of staphylococcal biofilm formation. J Microbiol
Methods. 40:175–179. 2000. View Article : Google Scholar
|
|
37
|
Pettit RK, Weber CA and Pettit GR:
Application of a high throughput Alamar blue biofilm susceptibility
assay to Staphylococcus aureus biofilms. Ann Clin Microbiol
Antimicrob. 8:282009. View Article : Google Scholar
|
|
38
|
Musken M, Di Fiore S, Römling U and
Häussler S: A 96-well-plate-based optical method for the
quantitative and qualitative evaluation of Pseudomonas
aeruginosa biofilm formation and its application to
susceptibility testing. Nat Protoc. 5:1460–1469. 2010. View Article : Google Scholar
|
|
39
|
Johnson MB and Criss AK: Fluorescence
microscopy methods for determining the viability of bacteria in
association with mammalian cells. J Vis Exp. Sep 5–2013.doi:
10.3791/50729. View
Article : Google Scholar
|
|
40
|
Konaté K, Mavoungou JF, Lepengué AN,
Aworet-Samseny RR, Hilou A, Souza A, Dicko MH and M'batchi B:
Antibacterial activity against β-lactamase producing Methicillin
and Ampicillin-resistants Staphylococcus aureus. Fractional
inhibitory concentration index (FICI) determination. Ann Clin
Microbiol Antimicrob. 11:182012. View Article : Google Scholar
|
|
41
|
Krishnan T, Yin WF and Chan KG: Inhibition
of quorum sensing-controlled virulence factor production in
Pseudomonas aeruginosa PAO1 by Ayurveda spice clove
(Syzygium aromaticum) bud extract. Sensors (Basel). 12:4016–4030.
2012. View Article : Google Scholar
|
|
42
|
Ha DG, Kuchma SL and O'Toole GA:
Plate-based assay for swarming motility in Pseudomonas
aeruginosa. Methods Mol Biol. 1149:67–72. 2014. View Article : Google Scholar
|
|
43
|
Schmittgen TD and Livak KJ: Analyzing
real-time PCR data by the comparative C(T) method. Nat Protoc.
3:1101–1108. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Cheng CL, Huang SJ, Wu CL, Gong HY, Ken
CF, Hu SY and Wu JL: Transgenic expression of omega-3 PUFA
synthesis genes improves zebrafish survival during Vibrio
vulnificus infection. J Biomed Sci. 22:1032015. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Gellatly SL and Hancock RE: Pseudomonas
aeruginosaNew insights into pathogenesis and host defenses.
Pathog Dis. 67:159–173. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Overhage J, Bains M, Brazas MD and Hancock
RE: Swarming of Pseudomonas aeruginosa is a complex
adaptation leading to increased production of virulence factors and
antibiotic resistance. J Bacteriol. 190:2671–2679. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Smith KD: Iron metabolism at the host
pathogen interface: Lipocalin 2 and the pathogen-associated iroA
gene cluster. Int J Biochem Cell Biol. 39:1776–1780. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Lamont IL, Konings AF and Reid DW: Iron
acquisition by Pseudomonas aeruginosa in the lungs of
patients with cystic fibrosis. Biometals. 22:53–60. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Andrejko M, Zdybicka-Barabas A, Janczarek
M and Cytryńska M: Three Pseudomonas aeruginosa strains with
different protease profiles. Acta Biochim Pol. 60:83–90.
2013.PubMed/NCBI
|
|
50
|
Diggle SP, Cornelis P, Williams P and
Cámara M: 4-quinolone signalling in Pseudomonas aeruginosa:
Old molecules, new perspectives. Int J Med Microbiol. 296:83–91.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Gallagher LA, McKnight SL, Kuznetsova MS,
Pesci EC and Manoil C: Functions required for extracellular
quinolone signaling by Pseudomonas aeruginosa. J Bacteriol.
184:6472–6480. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Déziel E, Lépine F, Milot S, He J,
Mindrinos MN, Tompkins RG and Rahme LG: Analysis of Pseudomonas
aeruginosa 4-hydroxy-2-alkylquinolines (HAQs) reveals a role
for 4-hydroxy-2-heptylquinoline in cell-to-cell communication. Proc
Natl Acad Sci USA. 101:1339–1344. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Déziel E, Gopalan S, Tampakaki AP, Lépine
F, Padfield KE, Saucier M, Xiao G and Rahme LG: The contribution of
MvfR to Pseudomonas aeruginosa pathogenesis and quorum
sensing circuitry regulation: Multiple quorum sensing-regulated
genes are modulated without affecting lasRI, rhlRI or the
production of N-acyl-L-homoserine lactones. Mol Microbiol.
55:998–1014. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Rawling EG, Brinkman FS and Hancock RE:
Roles of the carboxy-terminal half of Pseudomonas aeruginosa
major outer membrane protein OprF in cell shape, growth in
low-osmolarity medium, and peptidoglycan association. J Bacteriol.
180:3556–3562. 1998.PubMed/NCBI
|
|
55
|
Fito-Boncompte L, Chapalain A,
Bouffartigues E, Chaker H, Lesouhaitier O, Gicquel G, Bazire A,
Madi A, Connil N, Véron W, et al: Full virulence of Pseudomonas
aeruginosa requires OprF. Infect Immun. 79:1176–1186. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Lin YM, Wu SJ, Chang TW, Wang CF, Suen CS,
Hwang MJ, Chang MD, Chen YT and Liao YD: Outer membrane protein I
of Pseudomonas aeruginosa is a target of cationic
antimicrobial peptide/protein. J Biol Chem. 285:8985–8994. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Yu H, Boucher JC, Hibler NS and Deretic V:
Virulence properties of Pseudomonas aeruginosa lacking the
extreme-stress sigma factor AlgU (sigmaE). Infect Immun.
64:2774–2781. 1996.PubMed/NCBI
|
|
58
|
Stacey SD and Pritchett CL: Pseudomonas
aeruginosa AlgU contributes to posttranscriptional activity by
increasing rsma expression in a mucA22 strain. J Bacteriol.
198:1812–1826. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Prince AS: Biofilms, antimicrobial
resistance, and airway infection. N Engl J Med. 347:1110–1111.
2002. View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Kim HS, Lee SH, Byun Y and Park HD:
6-Gingerol reduces Pseudomonas aeruginosa biofilm formation
and virulence via quorum sensing inhibition. Sci Rep. 5:86562015.
View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Gujrati VB and Jon S: Bioengineered
bacterial outer membrane vesicles: What is their potential in
cancer therapy? Nanomedicine (Lond). 9:933–935. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Schertzer JW and Whiteley M: A
bilayer-couple model of bacterial outer membrane vesicle
biogenesis. MBio. 3:pii: e00297. –11. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Wessel AK, Liew J, Kwon T, Marcotte EM and
Whiteley M: Role of Pseudomonas aeruginosa
peptidoglycan-associated outer membrane proteins in vesicle
formation. J Bacteriol. 195:213–219. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Ramsey DM and Wozniak DJ: Understanding
the control of Pseudomonas aeruginosa alginate synthesis and
the prospects for management of chronic infections in cystic
fibrosis. Mol Microbiol. 56:309–322. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Franklin MJ, Nivens DE, Weadge JT and
Howell PL: Biosynthesis of the Pseudomonas aeruginosa
Extracellular Polysaccharides, Alginate, Pel, and Psl. Front
Microbiol. 2:1672011. View Article : Google Scholar : PubMed/NCBI
|
|
66
|
Price KE, Orazi G, Ruoff KL, Hebert WP,
O'Toole GA and Mastoridis P: Mannitol does not enhance tobramycin
killing of Pseudomonas aeruginosa in a cystic fibrosis model
system of biofilm formation. PLoS One. 10:e01411922015. View Article : Google Scholar : PubMed/NCBI
|
|
67
|
Furiga A, Lajoie B, El Hage S, Baziard G
and Roques C: Impairment of Pseudomonas aeruginosa biofilm
resistance to antibiotics by combining the drugs with a new
quorum-sensing inhibitor. Antimicrob Agents Chemother.
60:1676–1686. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
68
|
Anderson GG, Kenney TF, Macleod DL, Henig
NR and O'Toole GA: Eradication of Pseudomonas aeruginosa
biofilms on cultured airway cells by a fosfomycin/tobramycin
antibiotic combination. Pathog Dis. 67:39–45. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
69
|
Lawrence GD: The Fats of Life: Essential
Fatty Acids in Health and Disease. Rutgers University Press;
2010
|
|
70
|
Pontes-Arruda A, Martins LF, de Lima SM,
Isola AM, Toledo D, Rezende E, Maia M and Magnan GB: Investigating
Nutritional Therapy with EPA, GLA and Antioxidants Role in Sepsis
Treatment (INTERSEPT) Study Group: Enteral nutrition with
eicosapentaenoic acid, γ-linolenic acid and antioxidants in the
early treatment of sepsis: Results from a multicenter, prospective,
randomized, double-blinded, controlled study: The INTERSEPT study.
Crit Care. 15:R1442011. View Article : Google Scholar : PubMed/NCBI
|
|
71
|
Chen W, Jiang H, Zhou ZY, Tao YX, Cai B,
Liu J, Yang H, Lu CD and Zeng J: Is omega-3 fatty acids enriched
nutrition support safe for critical ill patients? A systematic
review and meta-analysis. Nutrients. 6:2148–2164. 2014. View Article : Google Scholar : PubMed/NCBI
|