Introduction
Uterine fibroids is the most common type of tumor to
occur in the female reproductive tract, and develops from the
clonal proliferation of single smooth muscle cells of the
myometrium, initially caused by cellular genetic changes (1,2).
Alterations in complex signaling pathways involving factors such as
steroids, growth factors, transforming growth factor-β
(TGF-β)/Smad, wingless-type (Wnt)/β-catenin, retinoic acid (RA)
serve an important role in the development of uterine fibroids
(3,4). Furthermore, these signaling factors may
regulate multiple pathways through common factors, such as
mitogen-activated protein kinase (MAPK) and protein kinase B (also
known as Akt) (4). The major risk
factors that promote tumor development include genetic, hormonal,
immunological and environmental factors (1–4). At
present, besides surgical approaches such as hysterectomy,
myomectomy, uterine artery embolization (UAE) and magnetic
resonance imaging-guided focused ultrasound surgery (MRgFUS)
(5), the pharmacological strategies
for uterine fibroids focus on relieving symptoms and slowing or
arresting tumor development. Three main types of therapeutic are
used to reduce symptoms and inhibit the proliferation of fibroids
(6,7), namely non-steroidal anti-inflammatory
drugs (NSAIDs) (8), gonadotropin
releasing hormone agonists (GnRHa) (9) and synthetic steroids with
antiprogesterone activity, including mifepristone and asoprisnil
(10). As a therapeutic strategy,
surgery is associated with operative mortality and morbidity
(11), and medicinal therapy is
similarly limited because of its side effects. GnRHa may relieve
bleeding and bulk-related symptoms, but may also cause significant
menopausal side effects (12,13).
Furthermore, progesterone antagonists and other hormonal therapies
that alter estrogen and progesterone production or function may
affect fertility (14).
As an important component of complementary and
alternative medicine, Chinese herbal formulas, including Guizhi
Fuling Wan (GFW), are utilized for the treatment of uterine
fibroids (15). A previous
bibliometrics study of modern literature that analyzed the names of
diseases treated with Guizhi Fuling pills demonstrated that the
formula was most frequently used to treat abdominal masses (zheng
jia) in traditional Chinese medicine (TCM) and uterine fibroids in
western medicine (15).
GFW was first described in Essential Prescriptions
from the Golden Cabinet (Jingui Yaolue) (16). The traditional effects of the GFW
formula are considered to be invigoration of the blood, prevention
of blood stasis and reduction of masses. GFW is composed of
Cinnamomi Ramulus (Gui Zhi), Poria Cocos (Schw.)
Wolf. (Fuling), Cortex Moutan (Mu Dan Pi),
Radix Paeoniae Rubra (Chi Shao) and Persicae
Semen (Tao Ren) (16,17). A
previous systematic review of 38 randomized controlled trials
involving 3,816 participants demonstrated that compared with
mifepristone alone, GFW or GFW plus mifepristone reduced the volume
of fibroids and improved dysmenorrhea to a greater extent, and were
considered to be safer (18). This
suggests that GFW may be a potential alternative medicine for the
treatment of uterine fibroids; however, its pharmacological
mechanism is not well understood.
Chinese formulas are multi-target, multi-component
recipes that achieve their specific therapeutic efficacy through
active components that regulate molecular networks within the body
(19). Therefore, novel methods and
tactics are required to systematically investigate and explain the
mechanism of Chinese formulas. Network pharmacology, combined with
pharmacology and pharmacodynamics, is a novel research field that
is involved in the application of omics and systems biology-based
technologies (20). As Chinese
formulas are considered to have multiple targets, pathways,
components and specificities, as necessary factors for the
treatment of multiple complex illnesses, network pharmacology
methods are suitable for investigating prior knowledge regarding
the combination rules of TCM herbal formulae (21). Based on a previous study by Zheng
et al (22), the present
study selected a network pharmacology approach to uncover the
pharmacological mechanism of GFW in the treatment of uterine
fibroids.
Materials and methods
Data preparation
Composite compounds of the GFW herbs
To collect data on the compounds of GFW, the TCM
Database@Taiwan (http://tcm.cmu.edu.tw/zh-tw/), as a comprehensive TCM
database previously used to identify TCMs for osteoarthritis
(23), and the Traditional Chinese
Medicine Systems Pharmacology Database (ibts.hkbu.edu.hk/LSP/tcmsp.php), as a systems
pharmacology platform for Chinese herbal medicines (24), were used, along with related
literature (25). A total of 565
herbal compounds were identified; 230 in Cinnamomi Ramulus,
55 in Poria Cocos (Schw.) Wolf., 78 in Cortex Moutan,
135 in Radix Paeoniae Rubra and 67 in Persicae Semen.
Combined with related literature (25), 28 active compounds were identified:
Cinnamic acid, cinnamic aldehyde, 3-(2-methoxyphenyl)-2-propenal,
polyporenic acid C, pachymic acid, dehydrotrametenolic acid,
trametenolic acid, gallic acid, oxypaeoniflorin, (+)-catechin,
apiopaeonoside, paeonilide, paeoniflorin, suffruticoside B,
suffruticoside D, galloylpaeoniflorin, tetragalloylglucopyranose,
pentagalloylglucopyranose, benzoic acid, mudanpioside H,
hexagalloylglucopyranose, benzoyloxypaeoniflorin, mudanpioside C,
benzoylalbiflorin, paeonol, benzoylpaeoniflorin, albiflorin and
amygdalin.
Compound target of each GFW herb
All the active compounds were input into SciFinder
(http://scifinder.cas.org), a database of chemical
and bibliographic information provided by the Chemical Abstracts
Service (Columbus, OH, USA), and the molecular structure of each
compound was obtained. The structures were drawn in ChemBioDraw
14.0 (PerkinElmer, Inc., Waltham, MA, USA) and saved as ‘mol2’ file
format. These files were imported into PharmMapper (lilab.ecust.edu.cn/pharmmapper), a web
server that uses a pharmacophore mapping approach for potential
drug target identification (26).
Using this web server, the targets of all compounds were
identified, excluding that of cinnamic aldehyde as the target data
for cinnamic aldehyde was not obtained via the PharmMapper
prediction. Due to non-standard naming of the compound targets, the
UniProt Knowledgebase (http://www.uniprot.org/) was used. The protein names
were input with the species limited to ‘homo sapiens’ to obtain
official symbols. Following these procedures, the compound targets
with official symbols were obtained from the UniProt
Knowledgebase.
Uterine fibroids targets
Genes associated with uterine fibroids were
identified with two resources; i) GeneCards (http://www.genecards.org), as a database containing
information on genes and their products and biomedical
applications, provided by the Weizmann Institute of Science
(Rehovot, Israel) and ii) the Online Mendelian Inheritance in Man
database (http://www.omim.org), which catalogues
all known diseases with a genetic component and, when possible,
links them to relevant genes in the human genome, providing
references for further research and tools for genomic analysis of
catalogued genes (27). These
databases were searched using the keywords ‘uterine fibroids’,
‘uterine fibromas’ and ‘leiomyoma uterine’, which identified a
total of 114 genes.
Protein-protein interaction (PPI) data
Data on PPIs was obtained from STRING (http://string-db.org/; version 10), with the species
limited to ‘homo sapiens’ and a confidence score >0.4, and InAct
(www.ebi.ac.uk/intact/; version 4.2.4).
STRING is a database of known and predicted protein-protein
interactions (28) and InAct
provides an open source database and analytical tool for molecular
interaction data (29).
Network construction
Network construction method
The following networks were constructed: A
compound-compound target network of GFW; a herb-compound
target-uterine fibroids target network of GFW; and a compound
target-uterine fibroids target-other human proteins PPI network.
The network analysis software Cytoscape (www.cytoscape.org; version 3.2.1) was used to
construct networks. Cytoscape is a software that may be used to
visualize biological pathways and intermolecular interaction
networks, among others. Furthermore, it provides a basic set of
features for data integration, analysis and visualization for
complicated network analysis (30).
Network topological feature set definition
The nodes in each network were evaluated based on
three indices: Degree, node betweenness and node closeness. Degree
indicates the number of edges between a node and other nodes in a
network (31). Node betweenness
evaluates the participation of a node in the shortest parts of a
network and reflects the capability of nodes to manage the rate of
information flow in the network (32). Node closeness represents the inverse
of the sum of the distance from node i (any given node) to other
nodes (33). The importance of a
node in a network is indicated by the values of these indices, with
higher values indicating greater importance (22).
Enrichment analysis
Gene ontology (GO) and pathway enrichment analyses
were also performed on the target data, using the Database for
Annotation, Visualization and Integrated Discovery (DAVID;
david.ncifcrf.gov/; version 6.8) (34). P-values were derived from the DAVID
database and are modified Fisher exact P-values. Smaller P-values
indicated greater enrichment (34).
Results
Compound-compound target network
analysis
All active compounds, their targets and the
interactions between them (excluding cinnamic aldehyde) are
presented in Fig. 1. This network
includes 389 nodes (362 compound targets and 27 compounds) and
3,500 edges. Nodes closer to the center exhibit more interactions
with compounds than peripheral nodes, which indicates that numerous
compound targets may be regulated by multiple compounds rather than
a single compound. Coagulation factor II (also known as
prothrombin), matrix metalloproteinase 3, carbonic anhydrase 2,
aldo-keto reductase family 1 member B and cyclin dependent kinase 2
(in Fig. 1, F2, MMP3, CA2, AKR1B1
and CDK2, respectively) may be controlled by all of the active
compounds.
Herb-compound target-uterine fibroids
target network analysis
To understand the relationship between herbs of the
GFW formula, compound targets and uterine fibroids targets, a
herb-compound target-uterine fibroids target network was
constructed. It was composed of 459 nodes (5 herbs, 362 compound
targets, 78 uterine fibroids targets and 14 compound-uterine
fibroids targets) and 3,736 edges (Fig.
2).
 | Figure 2.Herb-compound target-uterine fibroids
target network of Guizhi Fuling Wan. Green diamonds, pink circles,
blue hexagons and green hexagon represent the herbs, compound
targets, uterine fibroids targets and compound-uterine fibroids
targets, respectively; light lines indicate associations between
herbs and other nodes; and dark lines indicate associations between
fibroids targets, compound-uterine fibroids targets and compound
targets. C.R., Cinnamomi Ramulus; C.M., Cortex
Moutan; P.S., Persicae Semen; P.C.W., Poria Cocos
(Schw.) Wolf.; R.P.R., Radix Paeoniae Rubra. |
According to GO enrichment analysis, these targets
were significantly associated with the steroid hormone-mediated
signaling pathway (GO ID: 0043401; fold enrichment=16.1;
P<0.001), response to estrogen (GO ID: 0043627; fold
enrichment=8.4; P<0.001), response to estradiol (GO ID: 0032355;
fold enrichment=7.5; P<0.001), response to progesterone (GO ID:
0032570; fold enrichment=6.4; P<0.001), vascular endothelial
growth factor (VEGF) receptor signaling pathway (GO ID: 0048010;
fold enrichment=5.3; P<0.001), epidermal growth factor (EGF)
receptor signaling pathway (GO ID: 0007173; fold enrichment=4.3;
P<0.001), fibroblast growth factor (FGF) receptor signaling
pathway (GO ID: 0008543; fold enrichment=4.4; P<0.001),
insulin-like growth factor (IGF) receptor signaling pathway (GO ID:
0048009; fold enrichment=10.4; P=0.0058), negative regulation of
cell proliferation (GO ID: 0008285; fold enrichment=3.1;
P<0.001), negative regulation of smooth muscle cell
proliferation (GO ID: 0048662; fold enrichment=8.9; P<0.001), RA
receptor signaling pathway (GO ID: 0048384; fold enrichment=15.1;
P<0.001), response to vitamin A (GO ID: 0033189; fold
enrichment=13.5; P<0.001), response to RA (GO ID: 0032526; fold
enrichment=6.2; P<0.001), TGF-β receptor signaling pathway (GO
ID: 0007179; fold enrichment=4.1; P<0.001), negative regulation
of the TGF-β receptor signaling pathway (GO ID: 0030512; fold
enrichment=4.4; P<0.001), positive regulation of apoptotic
processes (GO ID: 0043065; fold enrichment=2.8; P<0.001) and
apoptotic processes (GO ID: 0006915; fold enrichment=2.0;
P<0.001; Table I and Fig. 3).
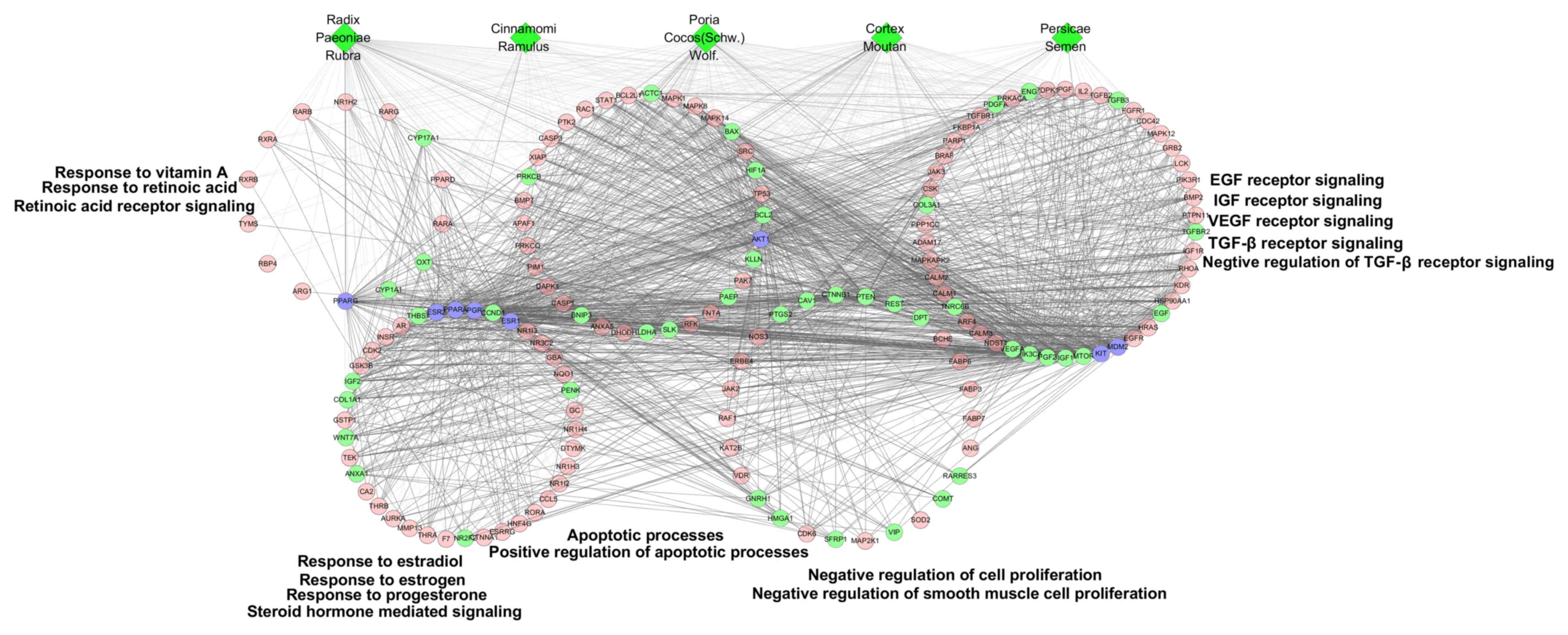 | Figure 3.GO enrichment analysis of compound
targets, uterine fibroids targets and compound target/uterine
fibroids targets. According to the associated biological processes,
compound targets of Guizhi Fuling Wan and uterine fibroids targets
were related to various molecular mechanisms of uterine fibroids.
Green diamonds, pink circles, green circles and blue circles
represent the herbs, compound targets, uterine fibroids targets and
compound-uterine fibroids targets, respectively; light lines
indicate associations between herbs and other nodes; and dark lines
indicate associations between compound-uterine fibroids targets and
compound targets. EGF, epidermal growth factor; FGF, fibroblast
growth factor; IGF, insulin-like growth factor; VEGF, vascular
endothelial growth factor; TGF-β, transforming growth factor-β. |
 | Table I.GO enrichment analysis of compound
targets, uterine fibroids targets and compound targets/uterine
fibroids targets. |
Table I.
GO enrichment analysis of compound
targets, uterine fibroids targets and compound targets/uterine
fibroids targets.
| GO ID | Pathway | Gene count | % | P-value | Fold
enrichment | Benjamini |
|---|
| 0043401 | Steroid hormone
mediated signaling | 26 | 0.040 | 4.80E-24 | 16.1 | 5.60E-21 |
| 0032355 | Response to
estradiol | 24 | 0.037 | 9.40E-14 | 7.5 | 2.50E-11 |
| 0043627 | Response to
estrogen | 15 | 0.023 | 1.70E-9 | 8.4 | 1.40E-7 |
| 0032570 | Response to
progesterone | 7 | 0.011 | 6.90E-4 | 6.4 | 1.30E-2 |
| 0048010 | Vascular
endothelial growth factor receptor signaling | 41 | 0.063 | 5.80E-18 | 5.3 | 3.40E-15 |
| 0007173 | Epidermal growth
factor receptor signaling | 38 | 0.058 | 1.40E-13 | 4.3 | 3.40E-11 |
| 0008543 | Fibroblast growth
factor receptor signaling | 35 | 0.054 | 8.70E-13 | 4.4 | 1.90E-10 |
| 0048009 | Insulin-like growth
factor receptor signaling | 4 | 0.006 | 5.80E-3 | 10.4 | 8.10E-2 |
| 0008285 | Negative regulation
of cell proliferation | 34 | 0.052 | 1.40E-8 | 3.1 | 9.30E-7 |
| 0048662 | Negative regulation
of smooth muscle cell proliferation | 8 | 0.012 | 2.50E-5 | 8.9 | 7.50E-4 |
| 0048384 | Retinoic acid
receptor signaling | 7 | 0.011 | 3.80E-6 | 15.1 | 1.40E-4 |
| 0033189 | Response to vitamin
A | 7 | 0.011 | 7.90E-6 | 13.5 | 2.70E-4 |
| 0032526 | Response to
retinoic acid | 8 | 0.012 | 2.60E-4 | 6.2 | 5.70E-3 |
| 0007179 | Transforming growth
factor-β receptor signaling | 15 | 0.023 | 1.90E-5 | 4.1 | 6.10E-4 |
| 0030512 | Negative regulation
of transforming growth factor-β receptor signaling | 9 | 0.014 | 9.70E-4 | 4.4 | 1.80E-2 |
| 0043065 | Positive regulation
of apoptotic processes | 23 | 0.035 | 2.50E-5 | 2.8 | 7.60E-4 |
| 0006915 | Apoptotic
processes | 33 | 0.051 | 2.30E-4 | 2.0 | 5.00E-3 |
Through pathway enrichment, it was observed that
compound targets and uterine fibroids targets were primarily
related to the phosphoinositide 3-kinase (PI3K)-Akt signaling
pathway (fold enrichment=3.0; P<0.001), Ras signaling pathway
(fold enrichment=3.6; P<0.001), MAPK signaling pathway (fold
enrichment=2.3; P<0.001), estrogen signaling pathway (fold
enrichment=4.7; P<0.001), VEGF signaling pathway (fold
enrichment=6.7; P<0.001), mechanistic target of rapamycin (mTOR)
signaling pathway (fold enrichment=5.1; P<0.001), Wnt signaling
pathway (fold enrichment=1.9; P=0.032), apoptosis (fold
enrichment=3.6; P<0.001), EGF signaling pathway (fold
enrichment=3.8; P<0.001), IGF-1 signaling pathway (fold
enrichment=4.5; P<0.001), role of ErbB2 receptor tyrosine kinase
2 in signal transduction and oncology (fold enrichment=4.1;
P<0.001), platelet-derived growth factor (PDGF) signaling
pathway (fold enrichment=3.3; P=0.0016), TGF-β signaling pathway
(fold enrichment=2.2; P=0.033), nuclear factor-κB (NF-κB) signaling
pathway (fold enrichment=2.1; P=0.043), extracellular
signal-regulated kinase (Erk)1/Erk2 MAPK signaling pathway (fold
enrichment=2.5; P=0.033; Table II
and Fig. 4)
 | Figure 4.Pathway enrichment analysis of
compound targets, uterine fibroids targets and compound
targets/uterine fibroids targets. According to pathway enrichment
analysis, compound targets of GFW and uterine fibroids targets were
related to various pathways. Green diamonds and pink, orange and
blue circles represent the herbs, compound targets, uterine
fibroids targets and compound-uterine fibroids targets,
respectively; Red squares indicate the pathway; red lines indicate
the associations between pathways and targets; green lines indicate
the associations between herbs and targets; orange lines indicate
the associations between uterine fibroids targets and other nodes;
and blue lines indicate the associations between compound-uterine
fibroids targets and other nodes. PI3K, phosphoinositide 3-kinase;
Akt, protein kinase B; Erk, extracellular signal-regulated kinase;
NF-κB, nuclear factor-κB; TGF-β, transforming growth factor-β; IGF,
insulin-like growth factor; EGF, epidermal growth factor; Wnt,
wingless-type; VEGF, vascular endothelial growth factor; mTOR,
mechanistic target of rapamycin; MAPK, mitogen-activated protein
kinase; PDGF, platelet-derived growth factor; C.R., Cinnamomi
Ramulus; C.M., Cortex Moutan; P.S., Persicae
Semen; P.C.W., Poria Cocos(Schw.) Wolf.; R.P.R.,
Radix Paeoniae Rubra. |
| Table II.Pathway enrichment analysis of
compound targets, uterine fibroids targets and compound
targets/uterine fibroids targets. |
Table II.
Pathway enrichment analysis of
compound targets, uterine fibroids targets and compound
targets/uterine fibroids targets.
| Pathway | Gene count | % | P-value | Fold
enrichment | Benjamini |
|---|
| Phosphoinositide
3-kinase/Akt signaling | 56 | 0.086 | 9.10E-14 | 3.0 | 2.70E-12 |
| Ras signaling | 44 | 0.068 | 1.20E-13 | 3.6 | 3.30E-12 |
| MAPK signaling | 32 | 0.049 | 1.30E-5 | 2.3 | 6.80E-5 |
| Estrogen
signaling | 25 | 0.038 | 2.30E-10 | 4.7 | 2.80E-9 |
| Vascular
endothelial growth factor signaling | 22 | 0.034 | 2.10E-12 | 6.7 | 4.60E-11 |
| Mechanistic target
of rapamycin signaling | 16 | 0.025 | 2.40E-7 | 5.1 | 1.60E-6 |
| Wingless-type
signaling | 14 | 0.022 | 3.20E-2 | 1.9 | 7.10E-2 |
| Apoptosis | 12 | 0.018 | 3.90E-4 | 3.6 | 1.50E-3 |
| Epidermal growth
factor signaling | 11 | 0.017 | 2.40E-4 | 3.8 | 4.50E-3 |
| Insulin-like growth
factor-1 signaling | 10 | 0.015 | 1.30E-4 | 4.5 | 3.20E-3 |
| Role of ErbB2
receptor tyrosine kinase 2 in signal transduction and oncology | 10 | 0.015 | 3.10E-4 | 4.1 | 5.30E-3 |
| Platelet-derived
growth factor signaling | 10 | 0.015 | 1.60E-3 | 3.3 | 1.70E-2 |
| Transforming growth
factor-β signaling | 10 | 0.015 | 3.30E-2 | 2.2 | 7.20E-2 |
| Nuclear factor-κB
signaling | 10 | 0.015 | 4.30E-2 | 2.1 | 9.00E-2 |
| Extracellular
signal-regulated kinase 1/2-MAPK signaling | 8 | 0.012 | 3.30E-2 | 2.5 | 1.50E-1 |
Compound target-uterine fibroids
target-other human proteins PPI network analysis
This network contained 2,112 nodes and 67,861 edges
(Fig. 5). In this network, nodes
with indices higher than the average values (gegree ≥0.0006257,
node betweenness ≥0.4363, closeness ≥64.26) were regarded as main
nodes. A total of 337 main nodes were selected for GO and pathway
enrichment analyses.
Based on GO enrichment analysis, a direct
interaction network between the main nodes was established. As
depicted in Fig. 6, the main nodes
were divided into four functional modules. Module 1 is associated
with cell proliferation including negative regulation of cell
proliferation (GO ID: 0008285; fold enrichment=4.6; P<0.001) and
negative regulation of smooth muscle cell proliferation (GO ID:
0048662; fold enrichment=8.9; P<0.001). Module 2 is associated
with the response of cells to steroid hormones including steroid
hormone-mediated signaling pathway (GO ID: 0043401; fold
enrichment=10.0; P<0.001), response to estrogen (GO ID: 0043627;
fold enrichment=9.8; P<0.001) and response to estradiol (GO ID:
0032355; fold enrichment=9.2; P<0.001). Module 3 is associated
with apoptosis including positive regulation of apoptotic processes
(GO ID: 0043065; fold enrichment=5.6; P<0.001) and the apoptotic
signaling pathway (GO ID: 0097190; fold enrichment=4.1;
P<0.001). Module 4 is associated with the response of cells to
growth factors including VEGF receptor signaling pathway (GO ID:
0048010; fold enrichment=9.3; P<0.001), EGF receptor signaling
pathway (GO ID: 0007173; fold enrichment=9.7; P<0.001), FGF
receptor signaling pathway (GO ID: 0008543; fold enrichment=10.1;
P<0.001), IGF receptor signaling pathway (GO ID: 0048009; fold
enrichment=14.0; P=0.0026), TGF-β receptor signaling pathway (GO
ID: 0007179; fold enrichment=10.9; P<0.001), negative regulation
of the TGF-β receptor signaling pathway (GO ID: 0030512; fold
enrichment=7.2; P<0.001). These data are presented in Table III.
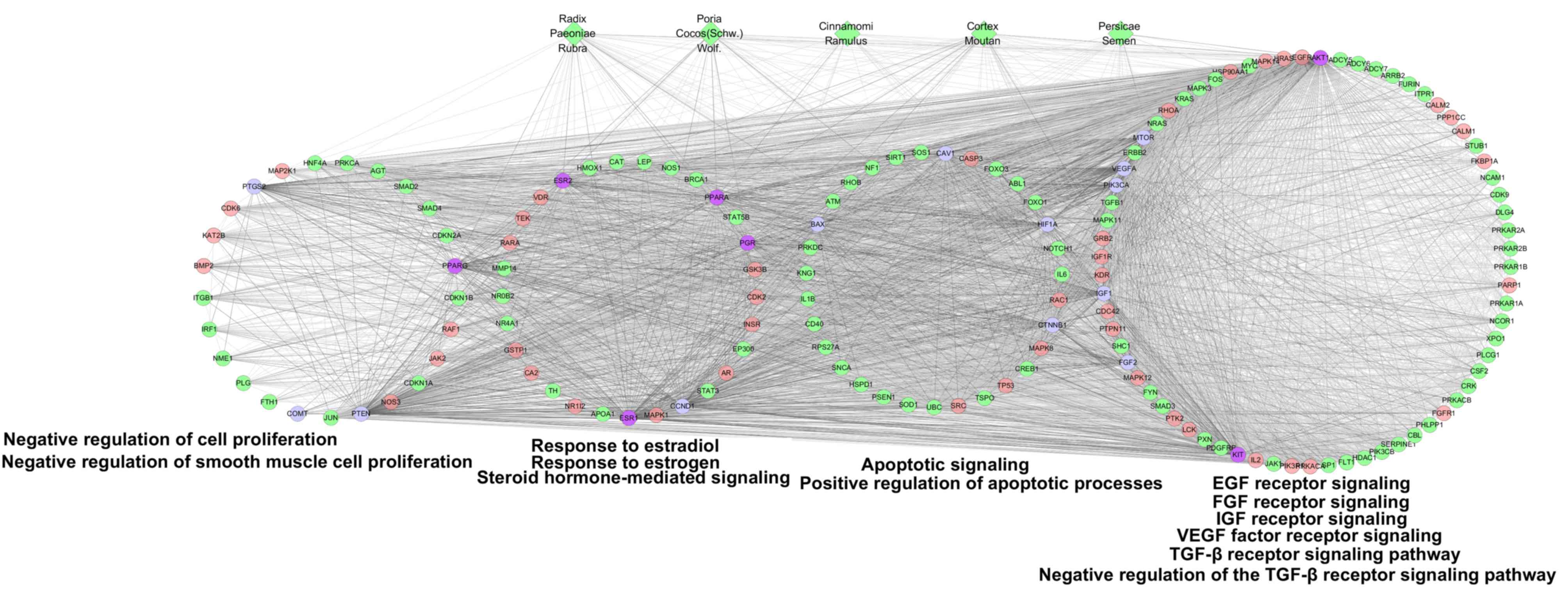 | Figure 6.GO enrichment analysis of compound
targets, uterine fibroids targets, compound targets/uterine
fibroids targets and other human proteins. According to the
associated biological processes, the nodes were categorized into
four modules. Green diamonds and pink, green, blue and purple
circles indicate the herbs, compound targets, other human proteins,
uterine fibroids targets and compound-uterine fibroids targets,
respectively; light lines represent associations between herbs and
other nodes; and dark lines represent associations between compound
targets, other human proteins, uterine fibroids targets and
compound-uterine fibroids targets. EGF, epidermal growth factor;
FGF, fibroblast growth factor; IGF, insulin-like growth factor;
VEGF, vascular endothelial growth factor. |
| Table III.GO enrichment analysis of compound
targets, uterine fibroids targets, compound targets/uterine
fibroids targets and other human proteins. |
Table III.
GO enrichment analysis of compound
targets, uterine fibroids targets, compound targets/uterine
fibroids targets and other human proteins.
| GO ID | Pathway | Gene count | % | P-value | Fold
enrichment | Benjamini |
|---|
| 0007173 | Epidermal growth
factor receptor signaling | 64 | 0.124 | 5.80E-44 | 9.7 | 5.60E-41 |
| 0008543 | Fibroblast growth
factor receptor signaling | 60 | 0.117 | 4.60E-42 | 10.1 | 3.60E-39 |
| 0048010 | Vascular
endothelial growth factor receptor signaling | 53 | 0.103 | 4.90E-35 | 9.3 | 2.40E-32 |
| 0048009 | Insulin-like growth
factor receptor signaling | 4 | 0.008 | 2.60E-3 | 14.0 | 3.00E-2 |
| 0007179 | Transforming growth
factor-β receptor signaling | 30 | 0.058 | 8.60E-22 | 10.9 | 1.70E-19 |
| 0030512 | Negative regulation
of transforming growth factor-β receptor signaling | 11 | 0.021 | 2.70E-6 | 7.2 | 6.10E-5 |
| 0032355 | Response to
estradiol | 22 | 0.043 | 2.40E-14 | 9.2 | 2.10E-12 |
| 0043627 | Response to
estrogen | 13 | 0.025 | 6.10E-9 | 9.8 | 2.20E-7 |
| 0043401 | Steroid
hormone-mediated signaling | 12 | 0.023 | 2.40E-8 | 10.0 | 7.60E-7 |
| 0008285 | Negative regulation
of cell proliferation | 37 | 0.072 | 5.20E-14 | 4.6 | 3.90E-12 |
| 0048662 | Negative regulation
of smooth muscle cell proliferation | 6 | 0.012 | 5.00E-4 | 8.9 | 6.90E-3 |
| 0097190 | Apoptotic
signaling | 10 | 0.019 | 7.60E-4 | 4.1 | 1.00E-2 |
| 0043065 | Positive regulation
of apoptotic processes | 34 | 0.066 | 2.10E-15 | 5.6 | 2.20E-13 |
Pathway enrichment analysis of the major nodes
indicated that the nodes were primarily related to the PI3K-Akt
signaling pathway (fold enrichment=4.6; P<0.001), Ras signaling
pathway (fold enrichment=5.1; P<0.001), MAPK signaling pathway
(fold enrichment=3.8; P<0.001), estrogen signaling pathway (fold
enrichment=8.5; P<0.001), ErbB signaling pathway (fold
enrichment=9.0; P<0.001), VEGF signaling pathway (fold
enrichment=10.7; P<0.001), mTOR signaling pathway (fold
enrichment=2.8; P=0.0056), Wnt signaling pathway (fold
enrichment=3.6; P<0.001), apoptosis (fold enrichment=4.9;
P<0.001), EGF signaling pathway (fold enrichment=5.4;
P<0.001), IGF-1 signaling pathway (fold enrichment=5.9;
P<0.001), TGF-β signaling pathway (fold enrichment=3.9;
P<0.001) and NF-κB signaling pathway (fold enrichment=4.0;
P<0.001; Table IV and Fig. 7).
 | Figure 7.Pathway enrichment analysis of
compound targets, uterine fibroids targets, compound
targets/uterine fibroids targets and other human proteins. Pathway
enrichment indicated that the major nodes were primarily linked to
the indicated pathways. Green diamonds and light pink, dark pink,
orange and blue circles indicate the herbs, compound targets, other
human proteins, uterine fibroids targets and compound-uterine
fibroids targets, respectively; red squares indicate the pathways;
red lines indicate the associations between pathways and targets;
green lines indicate the associations between herbs and targets;
orange lines indicate the associations between uterine fibroids
targets and other nodes; and blue lines indicate the associations
between compound-uterine fibroids targets and other nodes. PI3K,
phosphoinositide 3-kinase; Akt, protein kinase B; mTOR, mechanistic
target of rapamycin; TGF-β, transforming growth factor-β; NF-κB,
nuclear factor-κB; IGF, insulin-like growth factor; EGF, epidermal
growth factor; Wnt, wingless-type; VEGF, vascular endothelial
growth factor; MAPK, mitogen-activated protein kinase. |
| Table IV.Pathway enrichment analysis of
compound targets, uterine fibroids targets, compound
targets/uterine fibroids targets and other human proteins. |
Table IV.
Pathway enrichment analysis of
compound targets, uterine fibroids targets, compound
targets/uterine fibroids targets and other human proteins.
| Pathway | Gene count | % | P-value | Fold
enrichment | Benjamini |
|---|
| Phosphoinositide
3-kinase/Akt signaling | 73 | 0.142 | 1.30E-29 | 4.6 | 3.80E-28 |
| Ras signaling | 53 | 0.103 | 2.20E-23 | 5.1 | 3.30E-22 |
| Mitogen-activated
protein kinase signaling | 45 | 0.087 | 7.20E-15 | 3.8 | 4.40E-14 |
| Estrogen
signaling | 39 | 0.076 | 4.60E-26 | 8.5 | 9.90E-25 |
| ErbB signaling | 36 | 0.070 | 6.90E-25 | 9.0 | 1.40E-23 |
| Vascular
endothelial growth factor signaling | 30 | 0.058 | 2.60E-23 | 10.7 | 3.70E-22 |
| Wingless-type
signaling | 23 | 0.045 | 2.30E-7 | 3.6 | 7.20E-7 |
| Epidermal growth
factor signaling | 20 | 0.039 | 1.70E-11 | 5.4 | 8.20E-10 |
| Insulin-like growth
factor-1 signaling | 17 | 0.033 | 1.10E-10 | 5.9 | 3.90E-9 |
| Nuclear factor-κB
signaling | 16 | 0.031 | 8.10E-6 | 4.0 | 2.20E-5 |
| Transforming growth
factor-β signaling | 15 | 0.029 | 2.10E-5 | 3.9 | 5.40E-5 |
| Apoptosis | 14 | 0.027 | 3.50E-6 | 4.9 | 9.70E-6 |
| Mechanistic target
of rapamycin signaling | 10 | 0.037 | 5.60E-3 | 2.8 | 2.30E-2 |
Discussion
At present there is a lack of effective treatment
for uterine fibroids. The therapeutic strategies used in western
medicine include surgical treatment and pharmacological strategies,
though both achieve unsatisfactory outcomes (12–14). TCM
recipes exert therapeutic effects on a number of incurable
diseases, including uterine fibroids (15). Due to the multi-component and
multi-target features of TCM, the research approach for TCM should
be different to that of western medicine. However, many studies
still apply the conventional research approach of ‘one drug, one
target, one illness’, which does not account for the multi-target
and multi-component characteristics of TCM recipes (19–22).
Due to the development of bioinformatics, the
network approach has become a novel means of efficiently and
systemically identifying the potential molecular mechanisms of TCM
recipes. In the present study, a number of network-based
computational methods and algorithm-based approaches were used to
predict targets and construct networks, in order to assess the
molecular interactions associated with GFW when used as a uterine
fibroids therapy.
Other studies have demonstrated that there are
numerous pathways associated with the development of uterine
fibroids (35–42). In particular, steroid signaling
(estrogen and progesterone) has been implicated as a key factor in
the progression of uterine fibroids (35–38).
Alterations in other signaling pathways involving growth factors
and their cognate receptors also promote the growth and development
of uterine fibroids (39–42).
Regarding the steroid pathway, aberrant and rapid
MAPK signaling responses to estradiol may effect on leiomyoma
proliferation (43). It has been
observed that compared with the surrounding myometrium, the mRNA
transcription of estrogen receptor (ER)α and ERβ is elevated in
leiomyoma tissue (44–45). Maekawa et al (46) demonstrated that epigenetic regulation
of ERα through DNA methylation may serve a role in leiomyoma. In
addition, it has been reported that progresterone as well as
estrogen may serve a significant role in leiomyoma development
(38). Compared with
estrogen-mediated myometrium proliferation during the menstrual
cycle, secretion of progesterone promotes mitotic activity in
uterine leiomyomas (35,38). Furthermore, leiomyoma xenograft
animal models have indicated the necessity of progesterone for the
growth of uterine leiomyoma (47).
Progesterone-bound progesterone receptor (PR) can not only
accelerate the transcription of specificity protein-1 (SP-1)
(35), as a transcription factor
itself, but may also activate signaling pathways. For instance,
ligand-bound PRs may activate protein kinases involved in growth
factor signaling, such as MAPK and MEK (48).
The role of progesterone in the development of
uterine fibroids is complex. Estradiol may induce an upregulation
in PRs in leiomyoma cells (47), and
interactions between progesterone and growth factor signaling may
also promote the development of leiomyoma. For instance,
progesterone may downregulate the expression of IGF-1 in human
leiomyoma cells (49), upregulate
the expression of proliferating cell nuclear antigen (PCNA) and
EGF, as established regulators of leiomyoma cellular proliferation
(50,51), and activate the Akt pathway to
mediate leiomyoma proliferation (4).
These findings suggest that progesterone signaling is involved in
complex signaling networks associated with leiomyoma.
Regarding growth factors, previous studies suggest
that alterations in certain growth factors and their cognate
receptors or signaling pathways serve a significant role in the
growth and development of uterine fibroids (39–42).
These factors include IGF-1 (52,53),
PDGF (42), VEGF (54), EGF (55), and FGF (56). Activation of receptor tyrosine
kinases (RTKs) is a critical biological process; growth factor
binding to RTKs leads receptor dimerization and
autophosphorylation, thereby activating the downstream pathways
Grb2/Sos/Ras/Raf/MEK/Erk and PI3K/PIP3/Akt to regulate
proliferation, differentiation, survival and metabolism (57,58).
Previous results have indicated that IGF-1 signaling is regulated
by estrogen, and that 17β estradiol treatment leads to increases in
IGF-1 mRNA and Myb, a transcription factor that promotes the
expression of cell cycle progression genes in leiomyoma cells
(59). Furthermore, estradiol has
been demonstrated to promote the upregulation of growth factors and
RTKs in uterine leiomyomas, indicating that growth factors and RTKs
represent intermediate effectors of sex steroids in leiomyomas
(60). It has also been demonstrated
that under the influence of estrogen, the Ras/Raf/MEK/ERK signaling
pathway (43,59) and PI3K/Akt/mTOR signaling pathway
(57,61,62),
activated by RTK-ligand complexes, exert significant effects on the
pathological growth and development of fibroids. Expression of the
Smad signaling pathway mediated by ligands including TGF-β,
activin, mystatin, BMP and others belonging to the TGF-β
superfamily is also associated with leiomyoma, and is now becoming
a potential therapeutic target (63). Furthermore, compared with normal
smooth muscle cells, IGF-2 mRNA is upregulated in uterine leiomyoma
samples, and levels of IGF-1 are associated with Akt activation
(53). In uterine fibroids, both EGF
and PDGF have been found to stimulate protein synthesis in
leiomyoma and myometrial cells (64). Notably, downstream signaling induced
by EGF stimulation is altered in leiomyoma cells when compared with
myometrial cells (65).
VEGF was an essential factor for the growth of
leiomyoma xenografts in vivo (66–68).
Furthermore, its cognate receptors VEGFR-1 and VEGFR-2, and VEGF-A,
were significantly overexpressed in leiomyoma when compared with
adjacent myometrium (66–68). A recent study on Wnt signaling in the
growth and development of uterine fibroids demonstrated that the
Wnt/β-catenin signaling pathway mediated a novel interaction
between leiomyoma stem cells (representing 1% of tumor cells; also
known as a leiomyoma side-population), and mature leiomyoma cells,
which promoted tumor growth (69).
Furthermore, the paracrine effects of estrogen and progesterone may
stimulate leiomyoma cells to proliferate through Wnt/β-catenin
signaling (69). As an additional
factor, RA is an active metabolite of vitamin A (retinol) and
primarily promotes cellular growth and development (70). Previous results suggest that
receptors of RA signaling (RA and retinoid X receptors) are
expressed in leiomyoma cells (70).
The characteristics of fibroids are principally due
to the clonal proliferation of single smooth muscle cells in the
myometrium and alterations in complex signal pathways (3,4). The
associated signaling pathways are mediated by multiple factors,
including steroids, growth factors, TGF-β/Smad, Wnt/β-catenin and
RA (3,4). These signaling molecules in leiomyoma
cells serve a common role of mediating secretion from peripheral
stromal cells to regulate leiomyoma growth (3,4). The
regulation of estrogen and progesterone signaling may be an
important method for treating uterine fibroids. For instance,
continuous administration of gonadotropin-releasing GnRHa induced
menopausal status and lowered estrogen level, which was associated
with tumor shrinkage (71). However,
due to the side effects of long-term GnRHa use, including loss of
bone mineral density (71), it may
only be used for a relatively short period. Progesterone antagonist
and selective progesterone receptor modulator (SPRM) may inhibit
proliferation and induce apoptosis in leiomyoma cells (72,73).
Furthermore, SPRM was able to reduce the expression of IGF-1
(43), VEGF (74), EGF and TGF-β (75), which inhibits estrogen and
progesterone signaling, and the Ras/Raf/MEK/Erk and PI3K/Akt/mTOR
signaling pathways (75).
Asoprisnil, a SPRM, decreased the expression of certain growth
factors and growth factors receptors in leioyoma, including EGFR,
IGF-1Rα and TGFRII (75). However,
its side effects included endometrial hyperplasia and breast pain
and discomfort (76).
The TGF-β/Smad and RA signaling pathways also
represent potential targets for therapeutic development. Previous
data indicate that GnRHa may decrease the expression of TGF-β
receptors, Smad4 and phosphorylated Smad3 (63), and all-trans RA may inhibit the
proliferation of leiomyoma cells (77). At present, the major therapeutic
agents for uterine leiomyomas are steroids, SPRMs and selective
estrogen receptor modulators. However, long-term usage might cause
reproductive system side effects (12–14). The
present experimental data demonstrated that uterine fibroids may
also be alleviated by GFW through its downregulatory effects on
estrogen, progesterone and their cognate receptors and production
of TGF-β/Smad.
In uterine fibroids, the upregulation of five key
factors, namely steroids, growth factors, TGF-β/Smad, Wnt/-catenin
and RA, is closely related with tumor growth and development
(4). Leiomyoma and peripheral
stromal cells cooperate to promote leiomyoma proliferation and
increase the synthesis of peripheral matrix proteins (proteoglycans
and fibronectins) by paracrine signaling, which forms a complex
network involving alterations in cell shape and cytoskeleton
(78,79). Thus, regulation of paracrine
signaling molecules, as a therapeutic strategy for uterine
fibroids, may inhibit the synthesis of matrix proteins and
proteoglycans, inhibit the proliferation of leiomyoma cells and
promote leiomyoma cell apoptosis (4). Proliferation-related signaling pathways
mediated by estrogen and progesterone serve complex and important
roles in the pathology of uterine fibroids (39). Thus, use of estrogen and progesterone
antagonists, including GnRHa and synthetic steroids, with
progesterone antagonists such as mifepristone and asoprisnil, may
effectively inhibit the development of leiomyoma, and warrants
further study. Future research should also focus on the potential
regulatory effects of GFW on estrogen and progesterone
receptors.
Cell apoptosis, particularly of leiomyoma,
peripheral stromal and fibroids vascular endothelial cells and
leiomyoma stem cells, has been closely associated with fibroids
development (4). Leiomyoma stem
cells serve roles in the organizational structure and generation of
leiomyoma cells, and peripheral matrix synthesis, which contributes
to the development of fibroids (4).
Thus, inducing the apoptosis of various types of leiomyoma cells is
a terminal method for treating uterine fibroids. Previous studies
have demonstrated that total paeony glucosides, including
paeoniflorin, oxypaeoniflorin, benzoyloxypaeoniflorin,
benzoylalbiflorin and albiflorin, may induce tumor cell apoptosis
by increasing intracellular Ca2+ concentration,
inhibiting the mRNA transcription of B-cell lymphoma 2 (Bcl-2),
Bcl-extra large and upregulating Bcl-2-associated X protein
expression (79,80). Furthermore, human leiomyoma cell
proliferation may be inhibited by paeonol, pachymic acid,
albiflorin and paeoniflorin (80,81). It
has also been reported that GFW may promote tumor cell apoptosis
and inhibit human leiomyoma cell proliferation (81,82).
Therefore, GFW may prevent the growth of uterine fibroids by
promoting tumor cell apoptosis.
The uterine fibroids are characterized by excessive
deposition of extracellular matrix and proliferation of fibroids,
which is mediated by a variety of signaling pathways (4). For instance, receptor-bound growth
factors, estradiol and progesterone can activate Ras/Raf/MEK/Erk,
and thus inhibition of Ras/Raf/MEK/Erk may be an effective
therapeutic strategy, indicating the importance of double- and
multi-target treatments. At present, therapeutics that regulate
multiple targets and pathways are regarded as an alternative method
in the management of cancers such as prostate cancer (83). GFW may exert therapeutic effects
against uterine fibroids through multi-pathway and -target
activity.
According to the current predictions based on
network pharmacology, a number of novel signaling pathways and
biological processes underlying the effects of GFW on uterine
fibroids were identified. The results also provided a rationale for
the combination of herbs within GFW. This network pharmacology
method may aid the systematical study of herbal formulae and make
TCM drug discovery more predictable. Evaluating the efficacy of TCM
recipes and identifying the corresponding pharmacological mechanism
on a systematic level may be a useful method for future
studies.
Acknowledgements
The present study was supported by the National
Natural Science Foundation of China (grant no. 81303123)
Glossary
Abbreviations
Abbreviations:
|
GFW
|
Guizhi Fuling Wan
|
|
GO
|
gene ontology
|
|
TGF-β
|
transforming growth factor-β
|
|
Wnt
|
wingless-type
|
|
MAPK
|
mitogen-activated protein kinase
|
|
AKT
|
protein kinase B
|
|
UAE
|
uterine artery embolization
|
|
MRgFUS
|
magnetic resonance imaging-guided
focused ultrasound surgery
|
|
NSAIDs
|
non-steroidal anti-inflammatory
drugs
|
|
GnRHa
|
gonadotropin releasing hormone
agonists
|
|
TCM
|
traditional Chinese medicine
|
|
PPI
|
protein-protein interaction
|
|
IGF
|
insulin-like growth factor
|
|
PCNA
|
proliferating cell nuclear antigen
|
|
EGF
|
epidermal growth factor
|
|
PDGF
|
platelet-derived growth factor
|
|
VEGF
|
vascular endothelial growth factor
|
|
FGF
|
fibroblast growth factor
|
|
RTKs
|
receptor tyrosine kinase
|
|
RA
|
retinoic acid
|
|
SPRM
|
selective progesterone receptor
modulator
|
|
PI3K
|
phosphoinositide 3-kinase
|
|
mTOR
|
mechanistic target of rapamycin
|
|
NF-κB
|
nuclear factor-κB
|
|
Erk
|
extracellular signal-regulated
kinase
|
|
MEK
|
mitogen-activated protein kinase
kinase
|
|
ER
|
estrogen receptor
|
|
PR
|
progesterone receptor
|
|
Bcl-2
|
B-cell lymphoma 2
|
References
|
1
|
Townsend DE, Sparkes RS, Baluda MC and
McClelland G: Unicellular histogenesis of uterine leiomyomas as
determined by electrophoresis by glucose-6-phosphate dehydrogenase.
Am J Obstet Gynecol. 107:1168–1173. 1970. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Pandis N, Heim S, Bardi G, Flodérus UM,
Willén H, Mandahl N and Mitelman F: Chromosome analysis of 96
uterine leiomyomas. Cancer Genet Cytogenet. 55:11–18. 1991.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Rein MS: Advances in uterine leiomyoma
research: The progesterone hypothesis. Environ Health Perspect. 108
Suppl 5:S791–S793. 2000. View Article : Google Scholar
|
|
4
|
Borahay MA, Al-Hendy A, Kilic GS and
Boehning D: Signaling pathways in leiomyoma: Understanding
pathobiology and implications for therapy. Mol Med. 21:242–256.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Levy BS: Modern management of uterine
fibroids. Acta Obstet Gynecol Scand. 87:812–823. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Maruo T, Ohara N, Wang J and Matsuo H: Sex
steroidal regulation of uterine leiomyoma growth and apoptosis. Hum
Reprod Update. 10:207–220. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Grigorieva V, Chen-Mok M, Tarasova M and
Mikhailov A: Use of a levonorgestrel-releasing intrauterine system
to treat bleeding related to uterine leiomyomas. Fertil Steril.
79:1194–1198. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Falcone T and Bedaiwy MA: Minimally
invasive management of uterine fibroids. Curr Opin Obstet Gynecol.
14:401–407. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Lethaby A, Vollenhoven B and Sowter M:
Pre-operative GnRH analogue therapy before hysterectomy or
myomectomy for uterine fibroids. Cochrane Database Syst Rev:
CD000547. 2001. View Article : Google Scholar
|
|
10
|
Tristan M, Orozco LJ, Steed A,
Ramírez-Morera A and Stone P: Mifepristone for uterine fibroids.
Cochrane Database Syst Rev. doi:
10.1002/14651858.CD007687.pub2.
|
|
11
|
Lumsden MA: Modern management of fibroids.
Obstet Gynaecol Reprod Med. 20:82–86. 2010. View Article : Google Scholar
|
|
12
|
Walker CL and Stewart EA: Uterine
fibroids: The elephant in the room. Science. 308:1589–1592. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Lethaby A, Vollenhoven B and Sowter M:
Efficacy of pre-operative gonadotrophin hormone releasing analogues
for women with uterine fibroids undergoing hysterectomy or
myomectomy: A systematic review. BJOG. 109:1097–1108. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Sankaran S and Manyonda IT: Medical
management of fibroids. Best Pract Res Clin Obstet Gynaecol.
22:655–676. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Yan J, Li CP and Liu SS: Study of disease
names treated by Guizhi Fuling pills based on data mining
technology. Chin J Mod Drug Appl. 6:85–86. 2012.(In Chinese).
|
|
16
|
Fan YS: Jin Gui Yao Lue. China Press of
Traditional Chinese Medicine; Beijing: 2007
|
|
17
|
National Commission of Chinese
Pharmacopoeia, . Pharmacopoeia of the People's Republic of China.
Beijing China Med Scie Technol Press; 1. pp. 9842010, (In
Chinese).
|
|
18
|
Chen NN, Han M, Yang H, Yang GY, Wang YY,
Wu XK and Liu JP: Chinese herbal medicine Guizhi Fuling Formula for
treatment of uterine fibroids: A systematic review of randomised
clinical trials. BMC Complement Altern Med. 14:22014. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Liu AL and Du GH: Network pharmacology:
New guidelines for drug discovery. Yao Xue Xue Bao. 45:1472–1477.
2010.(In Chinese). PubMed/NCBI
|
|
20
|
Liang X, Li H and Li S: A novel network
pharmacology approach to analyse traditional herbal formulae: The
Liu-Wei-Di-Huang pill as a case study. Mol Biosyst. 10:1014–1022.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Li S, Zhang B, Jiang D, Wei Y and Zhang N:
Herb network construction and co-module analysis for uncovering the
combination rule of traditional Chinese herbal formulae. BMC
Bioinformatics. 11 Suppl 11:S62010. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Zheng CS, Xu XJ, Ye HZ, Wu GW, Li XH, Xu
HF and Liu XX: Network pharmacology-based prediction of the
multi-target capabilities of the compounds in Taohong Siwu
decoction, and their application in osteoarthritis. Exp Ther Med.
6:125–132. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Chen FP, Chang CM, Hwang SJ, Chen YC and
Chen FJ: Chinese herbal prescriptions for osteoarthritis in Taiwan:
Analysis of National Health Insurance dataset. BMC Complement
Altern Med. 14:912014. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Ru J, Li P, Wang J, Zhou W, Li B, Huang C,
Li P, Guo Z, Tao W, Yang Y, et al: TCMSP: A database of systems
pharmacology for drug discovery from herbal medicines. J
Cheminform. 6:132014. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Chen L, Wang D, Wu J, Yu B and Zhu D:
Identification of multiple constituents in the traditional Chinese
medicine formula GuiZhiFuLing-Wan by HPLC-DAD-MS/MS. J Pharm Biomed
Anal. 49:267–275. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Liu X, Ouyang S, Yu B, Liu Y, Huang K,
Gong J, Zheng S, Li Z, Li H and Jiang H: PharmMapper server: A web
server for potential drug target identification using pharmacophore
mapping approach. Nucleic Acids Res. 38:(Web Server Issue).
W609–W614. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Hamosh A, Scott AF, Amberger JS, Bocchini
CA and McKusick VA: Online Mendelian Inheritance in Man (OMIM), a
knowledgebase of human genes and genetic disorders. Nucleic Acids
Res. 33(Database Issue): D514–D517. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Szklarczyk D, Franceschini A, Wyder S,
Forslund K, Heller D, Huerta-Cepas J, Simonovic M, Roth A, Santos
A, Tsafou KP, et al: STRING v10: Protein-protein interaction
networks, integrated over the tree of life. Nucleic Acids Res.
43(Database Issue): D447–D452. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Orchard S, Ammari M, Aranda B, Breuza L,
Briganti L, Broackes-Carter F, Campbell NH, Chavali G, Chen C,
del-Toro N, et al: The MIntAct project-IntAct as a common curation
platform for 11 molecular interaction databases. Nucleic Acids Res.
42(Database Issue): D358–D363. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Franz M, Lopes CT, Huck G, Dong Y, Sumer O
and Bader GD: Cytoscape.js: A graph theory library for
visualisation and analysis. Bioinformatics. 32:309–311.
2016.PubMed/NCBI
|
|
31
|
Missiuro PV, Liu K, Zou L, Ross BC, Zhao
G, Liu JS and Ge H: Information flow analysis of interactome
networks. PLoS Comput Biol. 5:e10003502009. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Raman K, Damaraju N and Joshi GK: The
organisational structure of protein networks: Revisiting the
centrality-lethality hypothesis. Syst Synth Biol. 8:73–81. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Zhang Y, Bai M, Zhang B, Liu C, Guo Q, Sun
Y, Wang D, Wang C, Jiang Y, Lin N and Li S: Uncovering
pharmacological mechanisms of Wu-tou decoction acting on rheumatoid
arthritis through systems approaches: Drug-target prediction,
network analysis and experimental validation. Sci Rep. 5:94632015.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Huang da W, Sherman BT and Lempicki RA:
Systematic and integrative analysis of large gene lists using DAVID
bioinformatics resources. Nat Protoc. 4:44–57. 2009.PubMed/NCBI
|
|
35
|
Farber M, Conrad S, Heinrichs WL and
Herrmann WL: Estradiol binding by fibroid tumors and normal
myometrium. Obstet Gynecol. 40:479–486. 1972.PubMed/NCBI
|
|
36
|
Puukka MJ, Kontula KK, Kauppila AJ, Janne
OA and Vihko RK: Estrogen receptor in human myoma tissue. Mol Cell
Endocrinol. 6:35–44. 1976. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Kim JJ, Sefton EC and Bulun SE:
Progesterone receptor action in leiomyoma and endometrial cancer.
Prog Mol Biol Transl Sci. 87:53–85. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Kim JJ, Kurita T and Bulun SE:
Progesterone action in endometrial cancer, endometriosis, uterine
fibroids, and breast cancer. Endocr Rev. 34:130–162. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Islam MS, Protic O, Stortoni P, Grechi G,
Lamanna P, Petraglia F, Castellucci M and Ciarmela P: Complex
networks of multiple factors in the pathogenesis of uterine
leiomyoma. Fertil Steril. 100:178–193. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Ciarmela P, Islam MS, Reis FM, Gray PC,
Bloise E, Petraglia F, Vale W and Castellucci M: Growth factors and
myometrium: Biological effects in uterine fibroid and possible
clinical implications. Hum Reprod Update. 17:772–790. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Marsh EE and Bulun SE: Steroid hormones
and leiomyomas. Obstet Gynecol Clin North Am. 33:59–67. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Sozen I and Arici A: Interactions of
cytokines, growth factors, and the extracellular matrix in the
cellular biology of uterine leiomyomata. Fertil Steril. 78:1–12.
2002. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Nierth-Simpson EN, Martin MM, Chiang TC,
Melnik LI, Rhodes LV, Muir SE, Burow ME and McLachlan JA: Human
uterine smooth muscle and leiomyoma cells differ in their rapid
17beta-estradiol signaling: Implications for proliferation.
Endocrinology. 150:2436–2445. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Benassayag C, Leroy MJ, Rigourd V, Robert
B, Honoré JC, Mignot TM, Vacher-Lavenu MC, Chapron C and Ferré F:
Estrogen receptors (ERalpha/ERbeta) in normal and pathological
growth of the human myometrium: Pregnancy and leiomyoma. Am J
Physiol. 276:E1112–E1118. 1999.PubMed/NCBI
|
|
45
|
Kovács KA, Oszter A, Göcze PM, Környei JL
and Szabó I: Comparative analysis of cyclin D1 and oestrogen
receptor (alpha and beta) levels in human leiomyoma and adjacent
myometrium. Mol Hum Reprod. 7:1085–1091. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Maekawa R, Sato S, Yamagata Y, Asada H,
Tamura I, Lee L, Okada M, Tamura H, Takaki E, Nakai A and Sugino N:
Genome-wide DNA methylation analysis reveals a potential mechanism
for the pathogenesis and development of uterine leiomyomas. PLoS
One. 8:e666322013. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Ishikawa H, Ishi K, Serna VA, Kakazu R,
Bulun SE and Kurita T: Progesterone is essential for maintenance
and growth of uterine leiomyoma. Endocrinology. 151:2433–2442.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Lange CA: Integration of progesterone
receptor action with rapid signaling events in breast cancer
models. J Steroid Biochem Mol Biol. 108:203–212. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Boonyaratanakornkit V, Scott MP, Ribon V,
Sherman L, Anderson SM, Maller JL, Miller WT and Edwards DP:
Progesterone receptor contains a proline-rich motif that directly
interacts with SH3 domains and activates c-Src family tyrosine
kinases. Mol Cell. 8:269–280. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Maruo T, Matsuo H, Samoto T, Shimomura Y,
Kurachi O, Gao Z, Wang Y, Spitz IM and Johansson E: Effects of
progesterone on uterine leiomyoma growth and apoptosis. Steroids.
65:585–592. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Shimomura Y, Matsuo H, Samoto T and Maruo
T: Up-regulation by progesterone of proliferating cell nuclear
antigen and epidermal growth factor expression in human uterine
leiomyoma. J Clin Endocrinol Metab. 83:2192–2198. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Burroughs KD, Howe SR, Okubo Y,
Fuchs-Young R, LeRoith D and Walker CL: Dysregulation of IGF-I
signaling in uterine leiomyoma. J Endocrinol. 172:83–93. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Peng L, Wen Y, Han Y, Wei A, Shi G,
Mizuguchi M, Lee P, Hernando E, Mittal K and Wei JJ: Expression of
insulin-like growth factors (IGFs) and IGF signaling: Molecular
complexity in uterine leiomyomas. Fertil Steril. 91:2664–2675.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Chang CC, Hsieh YY, Lin WH and Lin CS:
Leiomyoma and vascular endothelial growth factor gene
polymorphisms: A systematic review. Taiwan J Obstet Gynecol.
49:247–253. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Rossi MJ, Chegini N and Masterson BJ:
Presence of epidermal growth factor, platelet-derived growth
factor, and their receptors in human myometrial tissue and smooth
muscle cells: Their action in smooth muscle cells in vitro.
Endocrinology. 130:1716–1727. 1992. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Helmke BM, Markowski DN, Müller MH, Sommer
A, Müller J, Möller C and Bullerdiek J: HMGA proteins regulate the
expression of FGF2 in uterine fibroids. Mol Hum Reprod. 17:135–142.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Kolch W: Meaningful relationships: The
regulation of the Ras/Raf/MEK/ERK pathway by protein interactions.
Biochem J. 351:289–305. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Crabtree JS, Jelinsky SA, Harris HA, Choe
SE, Cotreau MM, Kimberland ML, Wilson E, Saraf KA, Liu W,
McCampbell AS, et al: Comparison of human and rat uterine
leiomyomata: Identification of a dysregulated mammalian target of
rapamycin pathway. Cancer Res. 69:6171–6178. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Swartz CD, Afshari CA, Yu L, Hall KE and
Dixon D: Estrogen-induced changes in IGF-I, Myb family and MAP
kinase pathway genes in human uterine leiomyoma and normal uterine
smooth muscle cell lines. Mol Hum Reprod. 11:441–450. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Yu L, Saile K, Swartz CD, He H, Zheng X,
Kissling GE, Di X, Lucas S, Robboy SJ and Dixon D: Differential
expression of receptor tyrosine kinases (RTKs) and IGF-I pathway
activation in human uterine leiomyomas. Mol Med. 14:264–275. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Karra L, Shushan A, Ben-Meir A, Rojansky
N, Klein BY, Shveiky D, Levitzki R and Ben-Bassat H: Changes
related to phosphatidylinositol 3-kinase/Akt signaling in
leiomyomas: Possible involvement of glycogen synthase kinase 3alpha
and cyclin D2 in the pathophysiology. Fertil Steril. 93:2646–2651.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Jeong YJ, Noh EM, Lee YR, Yu HN, Jang KY,
Lee SJ, Kim J and Kim JS: 17β-estradiol induces up-regulation of
PTEN and PPARγ in leiomyoma cells, but not in normal cells. Int J
Oncol. 36:921–927. 2010.PubMed/NCBI
|
|
63
|
Chegini N, Luo X, Ding L and Ripley D: The
expression of Smads and transforming growth factor beta receptors
in leiomyoma and myometrium and the effect of gonadotropin
releasing hormone analogue therapy. Mol Cell Endocrinol. 209:9–16.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Fayed YM, Tsibris JC, Langenberg PW and
Robertson AL Jr: Human uterine leiomyoma cells: Binding and growth
responses to epidermal growth factor, platelet-derived growth
factor, and insulin. Lab Invest. 60:30–37. 1989.PubMed/NCBI
|
|
65
|
Ren Y, Yin H, Tian R, Cui L, Zhu Y, Lin W,
Tang XD, Gui Y and Zheng XL: Different effects of epidermal growth
factor on smooth muscle cells derived from human myometrium and
from leiomyoma. Fertil Steril. 96:1015–1020. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
66
|
Brown LF, Detmar M, Tognazzi K, Abu-Jawdeh
G and Iruela-Arispe ML: Uterine smooth muscle cells express
functional receptors (flt-1 and KDR) for vascular permeability
factor/vascular endothelial growth factor. Lab Invest. 76:245–255.
1997.PubMed/NCBI
|
|
67
|
Sanci M, Dikis C, Inan S, Turkoz E, Dicle
N and Ispahi C: Immunolocalization of VEGF VEGF receptors, EGF-R
and Ki-67 in leiomyoma, cellular leiomyoma and leiomyosarcoma. Acta
Histochem. 113:317–325. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
68
|
Gentry CC, Okolo SO, Fong LF, Crow JC,
Maclean AB and Perrett CW: Quantification of vascular endothelial
growth factor-A in leiomyomas and adjacent myometrium. Clin Sci
(Lond). 101:691–695. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
69
|
Ono M, Yin P, Navarro A, Moravek MB, JS V
Coon, Druschitz SA, Serna VA, Qiang W, Brooks DC, Malpani SS, et
al: Paracrine activation of WNT/β-catenin pathway in uterine
leiomyoma stem cells promotes tumor growth. Proc Natl Acad Sci USA.
110:pp. 17053–17058. 2013; View Article : Google Scholar : PubMed/NCBI
|
|
70
|
Catherino WH and Malik M: Uterine
leiomyomas express a molecular pattern that lowers retinoic acid
exposure. Fertil Steril. 87:1388–1398. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
71
|
Olive DL, Lindheim SR and Pritts EA:
Non-surgical management of leiomyoma: Impact on fertility. Curr
Opin Obstet Gynecol. 16:239–243. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
72
|
Fiscella K, Eisinger SH, Meldrum S, Feng
C, Fisher SG and Guzick DS: Effect of mifepristone for symptomatic
leiomyomata on quality of life and uterine size: A randomized
controlled trial. Obstet Gynecol. 108:1381–1387. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
73
|
Donnez J, Tatarchuk TF, Bouchard P,
Puscasiu L, Zakharenko NF, Ivanova T, Ugocsai G, Mara M, Jilla MP,
Bestel E, et al: Ulipristal acetate versus placebo for fibroid
treatment before surgery. N Engl J Med. 366:409–420. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
74
|
Xu Q, Ohara N, Chen W, Liu J, Sasaki H,
Morikawa A, Sitruk-Ware R, Johansson ED and Maruo T: Progesterone
receptor modulator CDB-2914 down-regulates vascular endothelial
growth factor, adrenomedullin and their receptors and modulates
progesterone receptor content in cultured human uterine leiomyoma
cells. Hum Reprod. 21:2408–2416. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
75
|
Ohara N, Morikawa A, Chen W, Wang J,
DeManno DA, Chwalisz K and Maruo T: Comparative effects of SPRM
asoprisnil (J867) on proliferation, apoptosis, and the expression
of growth factors in cultured uterine leiomyoma cells and normal
myometrial cells. Reprod Sci. 14 8 Suppl:S20–S27. 2007. View Article : Google Scholar
|
|
76
|
Eisinger SH, Bonfiglio T, Fiscella K,
Meldrum S and Guzick DS: Twelve-month safety and efficacy of
low-dose mifepristone for uterine myomas. J Minim Invasive Gynecol.
12:227–233. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
77
|
Boettger-Tong H, Shipley G, Hsu CJ and
Stancel GM: Cultured human uterine smooth muscle cells are retinoid
responsive. Proc Soc Exp Biol Med. 215:pp. 59–65. 1997; View Article : Google Scholar : PubMed/NCBI
|
|
78
|
Norian JM, Owen CM, Taboas J, Korecki C,
Tuan R, Malik M, Catherino WH and Segars JH: Characterization of
tissue biomechanics and mechanical signaling in uterine leiomyoma.
Matrix Biol. 31:57–65. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
79
|
Rogers R, Norian J, Malik M, Christman G,
Abu-Asab M, Chen F, Korecki C, Iatridis J, Catherino WH, Tuan RS,
et al: Mechanical homeostasis is altered in uterine leiomyoma. Am J
Obstet Gynecol. 198:474.e1–e11. 2008. View Article : Google Scholar
|
|
80
|
Xu HY, Chen ZW and Wu YM: Antitumor
activity of total paeony glycoside against human chronic myelocytic
leukemia K562 cell lines in vitro and in vivo. Med Oncol.
29:1137–1147. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
81
|
Tao XQ, Li N, Cao L, Zhang CF, Wang TJ,
Ding G, Wang ZZ and Xiao W: Effect of main components from Guizhi
Fuling capsule on human leiomyoma cell proliferation and
contraction of isolated mouse uterine. Chin J Exp Tradit Med Form.
2:91–96. 2016.
|
|
82
|
Lu CC, Shen CH, Chang CB, Hsieh HY, Wu JD,
Tseng LH, Hwang DW, Chen SY, Wu SF, Chan MW and Hsu CD: Guizhi
Fuling Wan as a novel agent for intravesical treatment for bladder
cancer in mouse model. Mol Med. Jan 13–2016.(Epub ahead of print).
View Article : Google Scholar
|
|
83
|
McCarty MF: Targeting multiple signaling
pathways as a strategy for managing prostate cancer: Multifocal
signal modulation therapy. Integr Cancer Ther. 3:349–380. 2004.
View Article : Google Scholar : PubMed/NCBI
|
















