Introduction
As a vital and regional growth site, mandibular
condylar cartilage is important to the function of the
temporomandibular joints and the morphogenesis of maxillofacial
areas (1). Therefore, an abundance
of studies have focused on the biological and pathological
mechanisms of mandibular condylar cartilage formation. Unlike other
articular cartilage within limb joints, mandibular condylar
cartilage is considered a secondary cartilage and its growth has
been categorized into two phases, maturation and mineralization
(2). Due to the cellular and
phenotypic responses of chondrocytes, in histology mandibular
condylar cartilage is consecutively categorized from the outside
in; the layers are as follows: Surface articular, resting
chondrocyte, proliferative chondrocyte, (non-mineralized)
hypertrophic chondrocyte and mineralized hypertrophic chondrocyte
(3). The morphogenesis of the
mandibular condyle is determined by chondrogenesis within each of
the layers.
Various growth factors and signaling pathways are
involved in the formation of mandibular condylar cartilage,
including Notch signaling members, insulin-like growth factors and
transforming growth factor β (TGF-β) superfamily members (4–6). Among
these factors and members, the TGF-β/bone morphogenic protein (BMP)
signaling pathway is noteworthy. Data from previous studies has
revealed that the TGF-β/BMP signaling pathway, which is
specifically transmitted through mothers against decapentaplegic
homolog (Smad) proteins, is involved in the proliferation and
differentiation of chondroblasts, and the ossification of the
extracellular matrix, in order to regulate the chondrogenesis of
mandibular condylar cartilage (7–9). Smads
are intracellular proteins that transduce the extracellular signals
from TGF-β ligands to the nucleus, where they activate downstream
gene transcription (1–4). Previous studies have demonstrated that
signal transduction via the Smad signaling pathway is crucial in
regulating TGF-β-mediated gene transcription and exerts versatile
regulatory functions on chondrogenesis (9,10). TGF-β
family members bind to their corresponding receptors and trigger
receptor phosphorylation, activating specific receptor-regulated
Smad (R-Smad) proteins to initiate intracellular signaling. R-Smads
combine with the common mediator Smad4 to form a complex; following
phosphorylation, the complex translocates into nuclei and interacts
with transcription factors to trigger the expression of specific
chondroblast genes (11). Smad2 and
3 are inhibitory mediators of TGF-β signaling during the maturation
of chondrocytes (12). Additionally,
Smad3 inhibits the terminal differentiation of chondrocytes, which
is essential for maintaining the morphology of the formed articular
cartilage (13). In Smad7
conditional knockout models, chondrocyte differentiation was
inhibited and malformed cartilage was observed as Smad7 disrupted
the formation of Smad2/3 and 4 complexes (14).
Based on the aforementioned evidence, the functions
of Smad proteins are key to understanding the effects of the
TGF-β/Smad signaling pathway on the development of articular
cartilage. However, to the best of our knowledge, there have been
few studies on the mechanism of Smad signaling in the development
of mandibular condylar cartilage. The present study investigated
the spatial and temporal protein expression of Smad2, 3, 4 and 7,
and phosphorylated (p-)Smad2/3, during the development of
mandibular condylar cartilage. The results of the present study may
provide novel insights into the development of mandibular condylar
cartilage and form a basis for further studies.
Materials and methods
Animals and tissues preparation
A total of 5 male and 20 female C57BL/6J mice (5
weeks old; weight, 25–30 g) were obtained from the Experimental
Animal Laboratory of Sichuan University (Chengdu, China) and
provided with access to food and water ad libitum under
climate-controlled conditions (25°C; 55% humidity; 12 h light/dark
cycle). At night, 3 females and 1 male were transferred into a cage
and left to mate. At 7 am the next day, the vaginal plug was
examined to confirm gestation, which was designated as embryonic
day (E) 0.5. Pregnant mice were anaesthetized with ether and
sacrificed by cervical dislocation at three selected time points
(E14.5, E15.5 and E16.5). Postnatal newborn mice were sacrificed in
the same way at postnatal day (P) 0 (following birth), 4 and 7. The
mandibular condyles of the fetuses and newborn mice were dissected.
Following fixation in 4% paraformaldehyde at room temperature for
24 h, the specimens were embedded in paraffin. Subsequently,
5-µm-thick serial sections were cut and mounted on
poly-L-lysine-coated glass slides. All procedures performed on the
animals were approved by the Ethics Committee of West China College
of Stomatology, Sichuan University (Chengdu, China).
Immunohistochemistry (IHC)
Primary antibodies directed against Smad2, 3, 4 and
7, and p-Smad2/3, were purchased from Santa Cruz Biotechnology,
Inc. (Dallas, TX, USA). The tissue sections were heated overnight
at 60°C, deparaffinized in xylene, rehydrated in ethanol, and
incubated for 20 min in 3% hydrogen peroxide
(H2O2) in methanol at room temperature to
block endogenous peroxidase activity. Sections were incubated in
the dark for 30 min at 37°C with 3% H2O2 in
trypsin-PBS (0.1 mg/ml; Sigma-Aldrich; Merck KGaA, Darmstadt,
Germany) and subsequently incubated overnight at 4°C with the
following primary antibodies: Mouse anti-Smad2 (cat. no.
sc-101153), 3 (cat. no. sc-101154), 4 (cat. no. sc-7966) and 7
(cat. no. sc-365846), and goat anti-p-Smad2/3 (cat. no. sc-11769)
at a dilution of 1:300. Negative control sections were incubated
with PBS instead of primary antibodies. Subsequently, the sections
were incubated with the appropriate secondary antibodies at 37°C
for 30 min, as follows: Biotin-conjugated secondary rabbit
anti-goat antibodies (cat. no. sc-2774; Santa Cruz Biotechnology,
Inc.) or goat anti-mouse antibodies (Biotin-Streptavidin HRP
Detection system; cat. no. sp-9002; OriGene Technologies, Inc.,
Beijing, China). Secondary antibodies were used at a dilution of
1:50. The peroxidase activity was visualized by immersing the
sections in 3,3′-diaminobenzidine at room temperature for 3–10 min.
Lastly, images were captured using an optical microscope
(magnification, ×100, 160 or 400).
Results
Histological observation
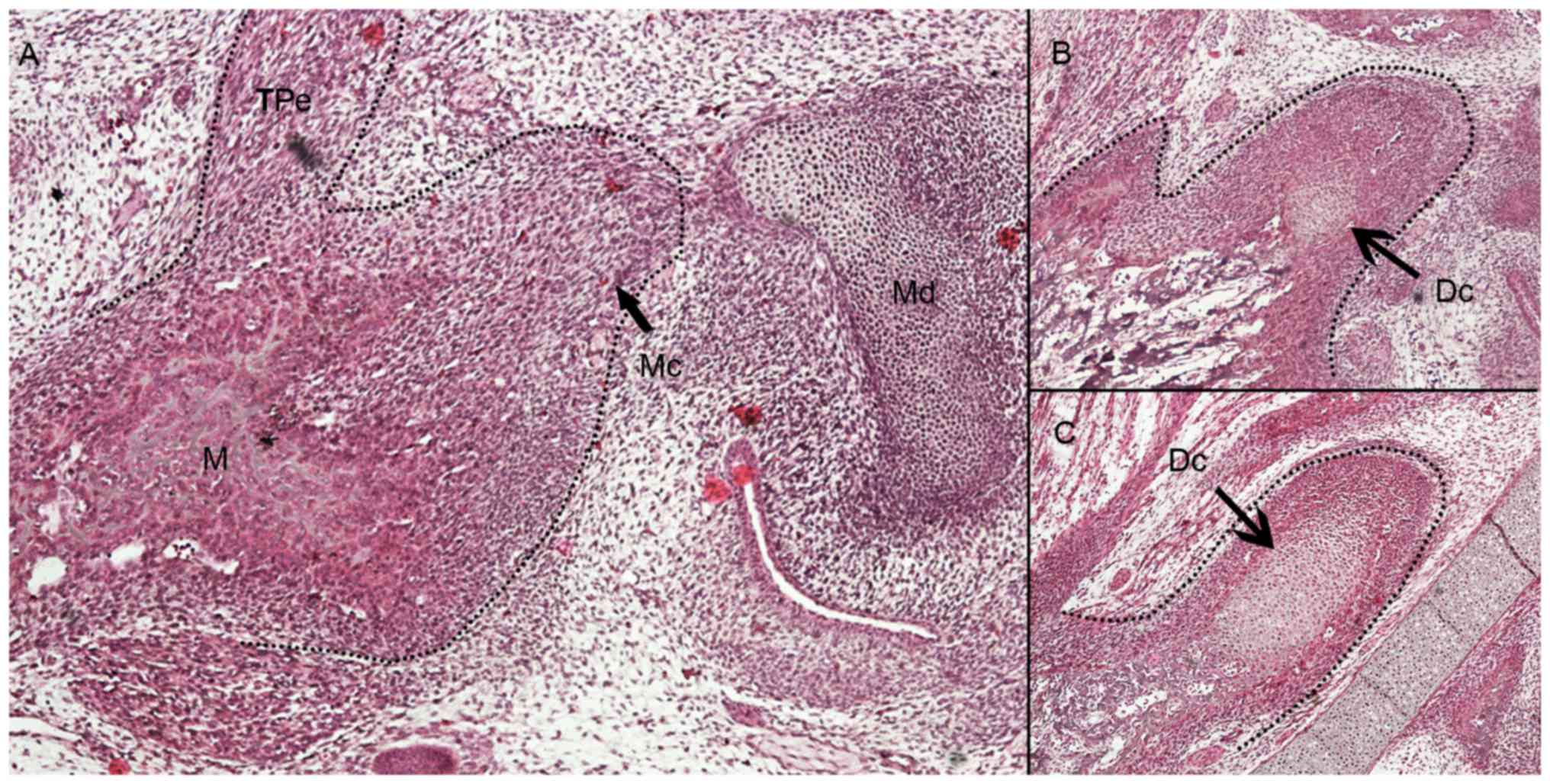
At E14.5, mesenchymal cells began to condense at the
distal upper portion of developing mandible (Fig. 1A). At E15.5, the condensation of the
mesenchymal cells was more recognizable but without defined
cartilaginous layers of condylar architectures (Fig. 1B). In the central region of the
developing condylar cartilage, differentiated chondrocytes were
first observed. At E16.5, the condylar cartilage was evident and
the cartilaginous cells exhibited layer-separated growth, including
undifferentiated outer layer and differentiated inner layer
chondrocytes (Fig. 1C).
At E17.5, a sequential arrangement of chondroblast
layers was observed (Fig. 2A and B).
Osteoblasts were identified at the periphery of the hypertrophic
chondrocytes. After E17.5, endochondral ossification progressed and
the length of the mandibular condyle increased (Fig. 2C and D). The primary structure of
mature condylar cartilage had taken shape by P7 (Fig. 2E).
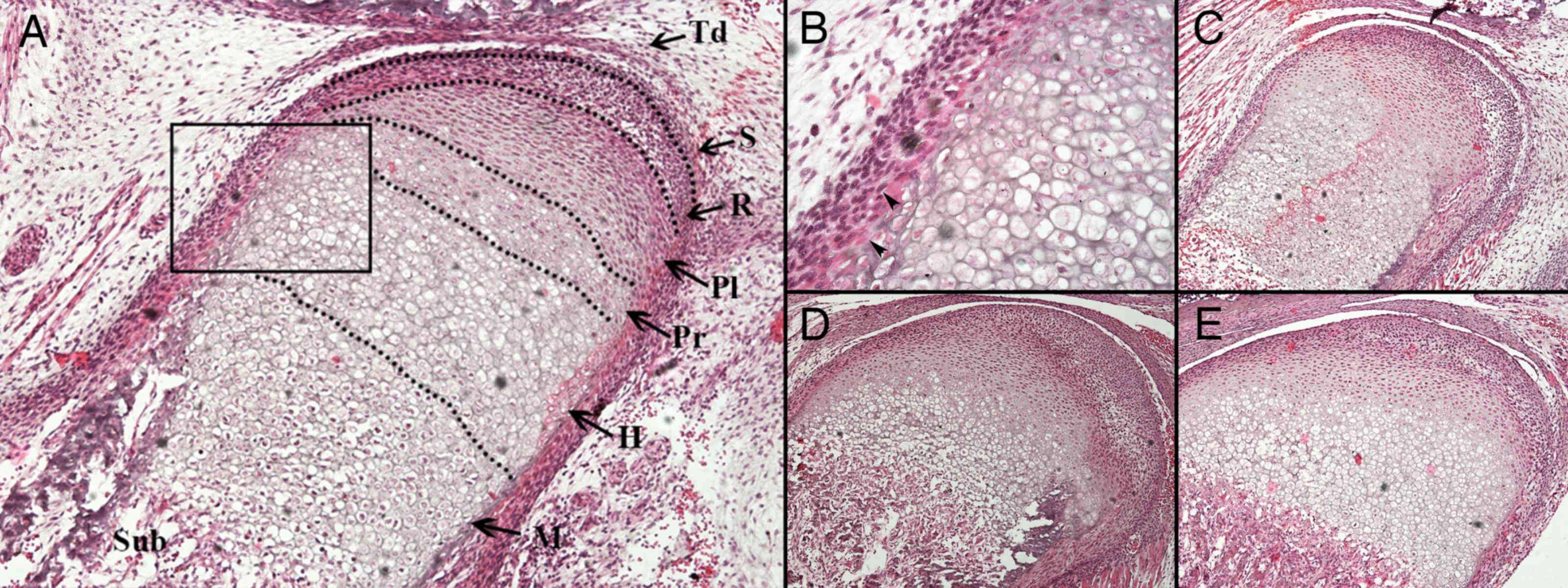 | Figure 2.Histological observation of the
developing mandibular condylar cartilage between E17.5 and P7. (A)
At E17.5, four layers of chondrocytes were arranged regularly and
possessed good continuity. The dotted line marks the edge of the
developing mandibular ramus and condyle. The boxed part is Fig. 2B. Magnification, ×100. (B) A
magnification of a section from Fig.
2A. The arrows indicate perichondral osteoblasts at the
perichondrium/periosteum of the developing condyle. Magnification,
×400. (C) At P0 each layer of the mandibular condylar cartilage was
observed. At (D) P4 and (E) P7, the length of the hypertrophic
chondrocyte layers in the vertical axis decreased, while the
subchondral bone increased in volume. Magnification, ×100. Td,
temporomandibular joint disk; S, surface articular layer; R,
resting chondrocyte layer; Pl, proliferative chondrocyte layer; Pr,
pre-hypertrophic chondrocyte layer; H, hypertrophic chondrocyte
layer (non-mineralized hypertrophic chondrocyte zone); M,
mineralized hypertrophic chondrocyte layer; Sub, subchondral bone;
E, embryonic day; P, postnatal day. |
IHC results
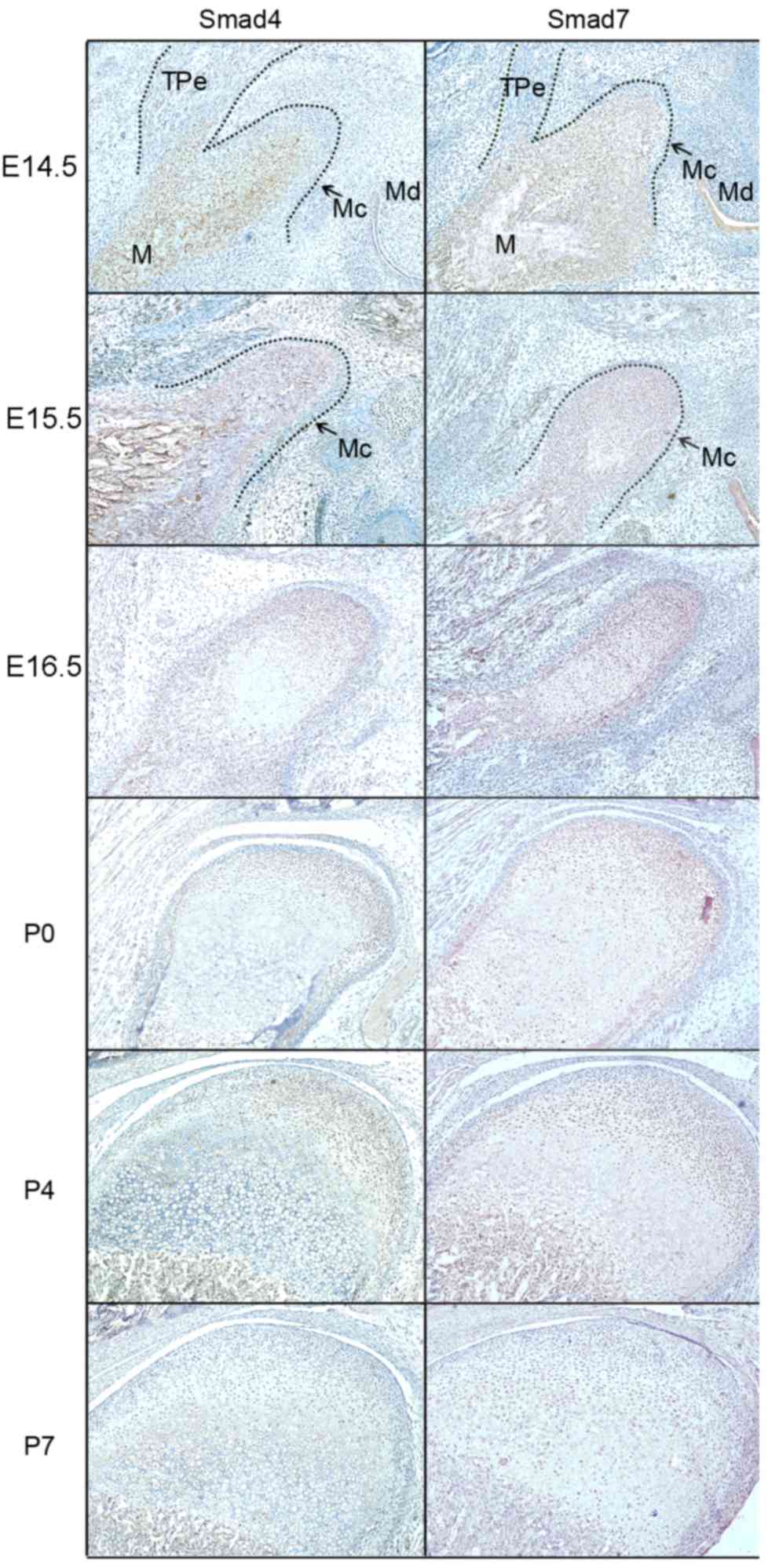
The IHC results (Figs.
3 and 4) demonstrated that
positive products of Smad2, 3, 4 and 7 were observed in the
cytoplasm of coagulating mesenchymal cells at the initial stage of
condylar development at E14.5 and E15.5. Notably, the expression
patterns of these products were not exactly accordant since the
expression of Smad4 was localized to the central section of the
mesenchymal condensation. At E16.5, the mesenchymal cells located
in the center of the developing condylar cartilage differentiated
into chondrocytes and the preliminary morphology of the mandibular
condylar cartilage was formed. During these processes, Smad2, 3, 4
and 7 were identified in differentiating chondrocytes. However,
Smad3 was mainly located in the differentiated chondrocytes, while
Smad4 was identified in the undifferentiated chondrocytes.
From P0, the chondrocytes underwent proliferation
and differentiation to elongate the condylar cartilage, which
corresponded with chondrogenesis in the mandibular condyles. Smad2
and 4 expression was associated with condylar cartilage growth;
their expression was identified in proliferative and mineralized
hypertrophic chondrocytes at the postnatal stage, but was reduced
in the hypertrophic zone. Smad3 and 7 were expressed in all
proliferative and hypertrophic chondrocytes of the developing
condylar cartilage, including pre-hypertrophic and mineralized
hypertrophic chondrocytes. Although Smad3 and 7 were expressed in
the cytoplasm of proliferative chondrocytes, Smad3 was observed in
the nuclei while Smad7 was observed in the cytoplasm of mineralized
hypertrophic chondrocytes. At P7, the expression of Smad3 in
hypertrophic and mineralized hypertrophic chondrocytes was
reduced.
As a direct downstream activator of the TGF-β/Smad
signaling pathway, p-Smad2/3 was expressed in different regions of
the developing condylar cartilage, which was similar to the
observed expression pattern of Smad3. During the chondrogenesis
stage of condylar cartilage development (prior to E16.5), p-Smad2/3
was primarily expressed in the cytoplasm of coagulating mesenchymal
cells in the condylar blastema. Notably, with the differentiation
and maturation of chondrocytes, the expression of p-Smad2/3
relocated from the cytoplasm to the nucleus of proliferative
chondrocytes and mineralized hypertrophic chondrocytes at P0 and
P4, and reduced in hypertrophic and mineralized hypertrophic
chondrocytes at P7. Notably, p-Smad2/3 was expressed in the
cytoplasm of proliferative chondrocytes at P7.
Discussion
Previous studies have reported that the TGF-β
signaling pathway serves an important role in cartilaginous
remodeling, osteoarthritis, osteochondromas and hyperplasia
(15–17). The expression of Smads has been
identified to be associated with the function of TGF-β signaling
(18,19). Typically, Smad2, 3 and 4 are
considered to be the intracellular mediator Smads that transduce
TGF-β signaling to the nuclei. During the differentiation of
chondrocytes, Smad2 and 3 form a complex with Smad4, which enters
the nuclei and modulates gene expression (5). As inhibitors of chondrocyte
differentiation, Smad2 and 3 are activated by TGF-β in
chondrocytes, and are involved in the reduction of chondrocyte
terminal differentiation, which mediates the suppressive effect of
TGF-β on the maturation of chondrocytes (15). Previous studies have reported that
Smad2 and 3 are co-expressed during the development of different
organs. During the development of Meckel's cartilage, Smad2 and 3
were identified to be expressed in differentiated chondrocytes
(18). The present study
demonstrated that the expression of Smad3 markedly overlapped with
Smad2 expression during chondrogenesis and the maturation of
proliferative chondrocytes. Similarly, in the epiphyseal growth
plate of limb joints, Smad2 and 3 are markedly expressed in
proliferating chondrocytes and maturing chondrocytes (12). Such coexpression implies a functional
synergy of Smad2 and 3. When p-Smad2/3 signaling is absent,
chondrocytes exit their quiescent state and undergo anomalous
terminal differentiation (18).
During endochondral ossification, Smad2 and 3 are
distinct in their expression patterns. Smad3 has been identified to
be predominantly expressed in the perichondrium of developing
cartilage undergoing endochondral ossification (20). The present study demonstrated that
along with the maturation and mineralization of chondrocytes, Smad3
expression was diminished in hypertrophic and mineralized
hypertrophic chondrocytes during the postnatal stages, despite the
expression of Smad3 being persistent in the cytoplasm of
proliferative chondrocytes. This expression pattern indicates that
the decreasing expression of Smad3 was potentially self-regulatory
to allow for endochondral ossification during the development of
condylar cartilage (21).
Nevertheless, certain studies have revealed that
Smad3 is dispensable in the early stages of cartilage formation
(13), and that a synergistic and
negative regulation exists between Smad2 and 3 signaling in the
regulation of chondrocyte differentiation (22). The present study demonstrated that
p-Smad2/3 was expressed in the nuclei of postnatal proliferative
and mineralized hypertrophic chondrocytes, but not hypertrophic
chondrocytes.
Smad proteins continuously move between the
cytoplasm and nuclei, thus Smads reach a steady state (23). It is well known that Smad4 combines
with Smad2/3 to form a complex, Smad4 is then activated when it
undergoes phosphorylation. The present study demonstrated that as a
central mediator for the TGF-β/BMP signaling pathway, Smad4, was
expressed in proliferative and hypertrophic chondrocytes during the
development of condylar cartilage, which was in accordance with the
Smad4 expression pattern identified in the epiphyseal growth plate
in a previous study (24).
In the present study, Smad7 was markedly expressed
in coagulating mesenchymal cells during chondrogenesis, and in
proliferative and hypertrophic chondrocytes during maturation of
condylar cartilage. As a negative regulator of the TGF-β/Smad
signaling pathway, Smad7 controls the function of R-Smads by
targeting the TGF-β receptor for degradation (25). A previous study demonstrated that
Smad7 antagonized TGF-β signaling in the nucleus by interfering
with functional Smad-DNA complex formation, and repressing Smad3/4,
Smad2/4 and Smad1/4 complex formation (26). Additionally, an abnormal feedback
regulation of Smad7 may cause the pathological growth of
chondrocytes (14).
In conclusion, the spatial and temporal expression
of Smad2, 3, 4 and 7, and p-Smad2/3, in the development of
mandibular condylar cartilage was explored in the present study.
The results demonstrated that these proteins were involved in the
development of mandibular condylar cartilage. In addition, these
Smad proteins were identified to be localized to different regions
and thus may exert distinct functions on chondrogenesis and
morphogenesis. The results of the present study may stimulate
further understanding of the biological function of Smad signaling
in the development of mandibular condylar cartilage.
References
|
1
|
Ranly DM: Craniofacial growth. Dent Clin
North Am. 44:457–470. 2000.PubMed/NCBI
|
|
2
|
Inoue H, Nebgen D and Veis A: Changes in
phenotypic gene expression in rat mandibular condylar cartilage
cells during long-term culture. J Bone Miner Res. 10:1691–1697.
1995. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Shen G and Darendeliler MA: The adaptive
remodeling of condylar cartilage-a transition from chondrogenesis
to osteogenesis. J Dent Res. 84:691–699. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Shimizu T, Nakano K, Tsujigiwa H,
Nagatsuka H, Watanabe T, Okafuji N, Kurihara S, Hasegawa H, Nagai N
and Kawakami T: Notch signaling in mandibular condylar cartilage
development. Eur J Med Res. 12:515–519. 2007.PubMed/NCBI
|
|
5
|
Van der Kraan PM, Davidson EN Blaney and
van den Berg WB: A role for age-related changes in TGFbeta
signaling in aberrant chondrocyte differentiation and
osteoarthritis. Arthritis Res Ther. 12:2012010. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Patil AS, Sable RB and Kothari RM: Role of
insulin-like growth factors (IGFs), their receptors and genetic
regulation in the chondrogenesis and growth of the mandibular
condylar cartilage. J Cell Physiol. 227:1796–1804. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Chai Y, Ito Y and Han J: TGF-beta
signaling and its functional significance in regulating the fate of
cranial neural crest cells. Crit Rev Oral Biol Med. 14:78–88. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Li TF, O'Keefe RJ and Chen D: TGF-beta
signaling in chondrocytes. Front Biosci. 10:681–688. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Chen G, Deng C and Li YP: TGF-β and BMP
signaling in osteoblast differentiation and bone formation. Int J
Biol Sci. 8:272–288. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Katagiri T and Takahashi N: Regulatory
mechanisms of osteoblast and osteoclast differentiation. Oral Dis.
8:147–159. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Guo X and Wang XF: Signaling cross-talk
between TGF-beta/BMP and other pathways. Cell Res. 19:71–88. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Li TF, Darowish M, Zuscik MJ, Chen D,
Schwarz EM, Rosier RN, Drissi H and O'Keefe RJ: Smad3-deficient
chondrocytes have enhanced BMP signaling and accelerated
differentiation. J Bone Miner Res. 21:4–16. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Yang X, Chen L, Xu X, Li C, Huang C and
Deng CX: TGF-beta/Smad3 signals repress chondrocyte hypertrophic
differentiation and are required for maintaining articular
cartilage. J Cell Biol. 153:35–46. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Iwai T, Murai J, Yoshikawa H and Tsumaki
N: Smad7 Inhibits chondrocyte differentiation at multiple steps
during endochondral bone formation and down-regulates p38 MAPK
pathways. J Biol Chem. 283:27154–27164. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Ferguson CM, Schwarz EM, Reynolds PR,
Puzas JE, Rosier RN and O'Keefe RJ: Smad2 and 3 mediate
transforming growth factor-beta1-induced inhibition of chondrocyte
maturation. Endocrinology. 141:4728–4735. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Meng Q, Long X, Deng M, Cai H and Li J:
The expressions of IGF-1, BMP-2 and TGF-β1 in cartilage of condylar
hyperplasia. J Oral Rehabil. 38:34–40. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Cuellar A, Inui A, James MA, Borys D and
Reddi AH: Immunohistochemical localization of bone morphogenetic
proteins (BMPs) and their receptors in solitary and multiple human
osteochondromas. J Histochem Cytochem. 62:488–498. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Ito Y, Bringas P Jr, Mogharei A, Zhao J,
Deng C and Chai Y: Receptor-regulated and inhibitory Smads are
critical in regulating transforming growth factor beta-mediated
Meckel's cartilage development. Dev Dyn. 224:69–478. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Flanders KC, Heger CD, Conway C, Tang B,
Sato M, Dengler SL, Goldsmith PK, Hewitt SM and Wakefield LM:
Brightfield proximity ligation assay reveals both canonical and
mixed transforming growth factor-β/bone morphogenetic protein Smad
signaling complexes in tissue sections. J Histochem Cytochem.
62:846–863. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Verdier MP, Seité S, Guntzer K, Pujol JP
and Boumédiène K: Immunohistochemical analysis of transforming
growth factor beta isoforms and their receptors in human cartilage
from normal and osteoarthritic femoral heads. Rheumatol Int.
25:118–124. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Song B, Estrada KD and Lyons KM: Smad
signaling in skeletal development and regeneration. Cytokine Growth
Factor Rev. 20:379–388. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Alvarez J and Serra R: Unique and
redundant roles of Smad3 in TGF-beta-mediated regulation of long
bone development in organ culture. Dev Dyn. 230:685–699. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Hill CS: Nucleocytoplasmic shuttling of
Smad proteins. Cell Res. 19:36–46. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Sakou T, Onishi T, Yamamoto T, Nagamine T
Sampath and Tk Ten Dijke P: Localization of Smads, the TGF-beta
family intracellular signaling components during endochondral
ossification. J Bone Miner Res. 14:1145–1152. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Shi Y and Massagué J: Mechanisms of
TGF-beta signaling from cell membrane to the nucleus. Cell.
113:685–700. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Zhang S, Fei T, Zhang L, Zhang R, Chen F,
Ning Y, Han Y, Feng XH, Meng A and Chen YG: Smad7 antagonizes
transforming growth factor beta signaling in the nucleus by
interfering with functional SmadDNA complex formation. Mol Cell
Biol. 27:4488–4499. 2007. View Article : Google Scholar : PubMed/NCBI
|