Introduction
During the repair of vascular injury, various
cytokines, such as platelet-derived growth factor (PDGF), that are
able to stimulate the proliferation and migration of vascular
smooth muscle cells (VSMCs) are released (1). Unbalanced proliferation and migration
of VSMCs has been demonstrated to serve key roles in neointimal
formation and thus are important for arteriosclerosis and
restenosis following coronary intervention or vein grafting
(2). Therefore, exploring the
underlying molecular mechanism may be beneficial for the
development of effective strategies for inhibiting this process,
and thus reducing the incidence of cardiovascular diseases
(3,4).
microRNAs (miRs), a type of non-coding RNAs that are
18–25 nucleotides in length, are key regulators for gene expression
through binding to the 3′-untranslated region (UTR) of target
mRNAs, causing mRNA degradation or translation inhibition (5,6). Through
inhibiting the protein expression of their target genes, miRs serve
important roles in various cellular biological processes, including
cell survival, differentiation, proliferation, apoptosis,
migration, angiogenesis and tumorigenesis (7–9).
Recently, miRs have been found to participate in the development
and progression of atherosclerosis and restenosis, and alterations
of miR expression profiles have been identified in these vascular
diseases (10,11). For instance, in arterial lesions
following balloon injury, many miRs have been found to be
significantly upregulated or downregulated (11). Furthermore, some miRs have been
reported to have promoting or suppressive roles in abnormal
proliferation and migration of VSMCs (2). For instance, Liu et al (12) reported that miR-221 and miR-222 were
necessary for the proliferation of VSMCs and neointimal
hyperplasia, and inhibition of miR-221 and miR-222 expression in
rat carotid arteries reduced VSMC proliferation and suppressed
neointimal formation following angioplasty. Sun et al
(13) demonstrated that miR-146a
serves a promoting role in VSMC proliferation in vitro and
vascular neointimal hyperplasia in vivo, at least in part,
by directly targeting Kruppel-like factor 4. Recently, miR-612 has
been demonstrated to have suppressive effects on the stemness of
liver cancer, as well as tumor growth and metastasis in colorectal
cancer (14,15). However, the molecular mechanism of
miR-612 on vascular cell dynamics remains to be elucidated.
The aim of the present study was to investigate the
exact role of miR-612 in platelet-derived growth factor
(PDGF)-BB-induced proliferation and migration of VSMCs, as well as
the underlying mechanism.
Materials and methods
Animals
The present study was approved by the Animal Care
and Use Committee of the Affiliated Hospital of Binzhou Medical
College (Binzhou, China). Animal experiments were consistent with
the National Institutes of Health Guide for the Care and Use of
Laboratory Animals. A total of 5 male Sprague Dawley rats (200–250
g, 8 weeks old) were purchased from the Shandong Laboratory Animal
Center (Jinan, China) and housed in light-controlled (12 h
light/dark cycle) and temperature-controlled (22±2°C) room with
free access to food and water.
Cell culture and treatment
The present study was approved by the Ethics
Committee of the Affiliated Hospital of Binzhou Medical College
(Binzhou, China). VSMCs were isolated from the thoracic aortas of
rats, and cultured in Dulbecco's modified Eagle's medium (DMEM;
Thermo Fisher Scientific, Inc., Waltham, MA, USA) supplemented with
10% fetal bovine serum (FBS; Thermo Fisher Scientific, Inc.), 10
mmol/l 4-(2-hydroxyethyl)-1-piperazineethanesulphonic acid
(Sigma-Aldrich; Merck KGaA, Darmstadt, Germany), 100 U/ml
penicillin (Sigma-Aldrich; Merck KGaA) and 100 mg/ml streptomycin
(Sigma-Aldrich; Merck KGaA) at 37°C in a humidified atmosphere
containing 5% CO2 and 95% air. VSMCs of fifth passage
were used. VSMCs without any treatment were used as the control
group. VSMCs in the PDGF-BB group were cultured to 70% confluence,
and subsequently incubated at 37°C with PDGF-BB (20 ng/ml; Thermo
Fisher Scientific, Inc.) for 6 h.
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
TRIzol reagent (Thermo Fisher Scientific, Inc.) was
used to extract total RNA from VSMCs, which was to produce cDNA
using a PrimeScript RT reagent kit (Takara Biotechnology Co., Ltd.,
Dalian, China), according to the manufacturer's protocol. qPCR was
conducted using a mirVana miRNA Detection kit (Thermo Fisher
Scientific, Inc.) with an ABI 7500 thermocycler (Thermo Fisher
Scientific, Inc.). The sequences for miR-612 (cat. no. HmiR0173)
and the reference gene U6 (cat. no. HmiRQP9001) were not supplied
by the manufacturer (Fulengen, Guangzhou, China). The reaction
conditions for PCR were as follows: Pre-degeneration at 95°C for 3
min and 40 cycles of 95°C for 30 sec, 60°C for 30 sec. Independent
experiments were repeated three times. The relative expressions of
mRNA were analyzed by use of the 2−∆∆Cq method (16).
Cell transfection
Lipofectamine® 2000 (Thermo Fisher
Scientific, Inc.) was used to perform cell transfection, in
accordance with the manufacturer's protocol. Briefly, VSMCs were
cultured to 70% confluence, and resuspended in serum-free DMEM.
Scramble miR mimic (miR-NC) or miR-612 mimic (100 nM, Yearthbio,
Changsha, China) were diluted in OPTI-MEM (Thermo Fisher
Scientific, Inc.), respectively, which was then added with diluted
Lipofectamine® 2000. The sequences for scramble miR
mimic (cat. no. AM1011) and miR-612 (cat. no. AM1423) mimic were
not supplied by the manufacturer (Fulengen). Following incubation
for 20 min at room temperature, the mixture was added into the cell
suspension. Following incubation at 37°C and 5% CO2 for
6 h, the transfection mixture was replaced with DMEM with 10% FBS.
At 48 h following transfection, the expression levels of miR-612
were examined using RT-qPCR following the procedure as above.
Following transfection, VSMCs were treated with PDGF-BB for 6
h.
MTT assay
VSMCs (5×103) in each group were seeded
in a 96-well plate, and 100 µl fresh serum-free DMEM with 0.5 g/l
MTT was added to each well. Following incubation at 37°C for 0, 24,
48 and 72 h, the medium containing MTT was removed, and 100 µl
dimethyl sulfoxide was added. Following incubation at 37°C for 10
min, the absorbance at 570 nm of each sample was measured using a
plate reader (Infinite M200; Tecan Group, Ltd., Männedorf,
Switzerland).
Transwell assay
Transwell assay was conducted to examine cell
migration using a 24-well Transwell chamber (Chemicon; EMD
Millipore, Billerica, MA USA). VSMCs cell suspension (containing
5×105 cells) was added to the upper chamber, and DMEM
containing 10% FBS was added into the lower chamber. Following
incubation in a 37°C humidified atmosphere at 5% CO2 for
24 h, cells on the interior of the inserts were removed using a
cotton-tipped swab. Migratory cells on the lower surface of the
membrane were stained at room temperature for 10 min with gentian
violet (Sigma-Aldrich; Merck KGaA), rinsed with water at room
temperature for 10 min, dried in air at room temperature for 20
min, and counted under a light microscope (magnification, ×200;
Nikon Corporation, Tokyo, USA).
Cell cycle analysis
VSMCs (5×106) in each group were fixed in
70% ethanol overnight at −20°C, and centrifuged at 4°C at 1,000 × g
for 5 min, washed in PBS, and then centrifuged at 4°C at 1,000 × g
for 5 min. Cells were resuspended in 300 µl propidium iodide
staining buffer (Beyotime Institute of Biotechnology, Haimen,
China) and incubated for 30 min at room temperature. DNA content
analyses were performed using a flow cytometer (C6; BD Biosciences,
Franklin Lakes, NJ, USA).
Western blot analysis
Cold RIPA lysis buffer (Beyotime Institute of
Biotechnology) was used to solubilize VSMCs. The concentration of
protein was determined with a BCA Protein Assay kit (Thermo Fisher
Scientific, Inc.). Protein (50 µg) was separated by 10% SDS-PAGE,
and then transferred to a polyvinylidene fluoride membrane (Thermo
Fisher Scientific, Inc.). The membrane was blocked at room
temperature in 5% non-fat dried milk in TBST (Thermo Fisher
Scientific, Inc.) for 2 h. Following three 5 min washes with TBST,
the membrane was incubated with rabbit anti-total AKT2 (1:100;
ab175354), rabbit anti-phosphorylated AKT2 (1:100; ab38513) or
rabbit anti-GAPDH antibody (1:100; ab9485; all Abcam, Cambridge,
UK) at 4°C overnight. Following three 5 min washes with TBST, the
membrane was incubated with horseradish peroxidase conjugated goat
anti-rabbit secondary antibody (1:5,000; ab7090; Abcam) for 40 min
at room temperature. Following three 5 min washes with TBST, the
immune complexes on PVDF membrane were then detected using an
enhanced chemiluminescent western blotting kit (Thermo Fisher
Scientific, Inc.), according to the manufacturer's protocol. The
relative protein expression was analyzed using ImageJ software 1.4
(National Institutes of Health, Bethesda, MD, USA), and represented
as the density ratio vs. GAPDH.
Bioinformatics analysis and
dual-luciferase reporter assay
TargetScan Human 3.1 (www.targetscan.org) was used to predict the potential
target of miR-216. The mutant type (MT) of AKT2 3′UTR was
constructed using an Easy Mutagenesis System kit (Promega
Corporation, Madison, WI, USA), in accordance with the
manufacturer's protocol. The wild-type (WT) or MT of AKT2 3′UTR was
then inserted into the pMIR-REPORT miR Expression Reporter vector
(Promega Corporation). VSMCs in the control group were
co-transfected with WT-AKT2-3′UTR or MT-AKT2-3′UTR plasmid, and
miR-NC or miR-216 mimic, using Lipofectamine® 2000,
respectively. At 48 h following transfection, the Dual-Luciferase
Reporter Assay System (Promega Corporation) was used to determine
the luciferase activity, and the Renilla luciferase activity
was normalized to the firefly luciferase activity.
Statistical analysis
Data are presented as the mean ± standard deviation.
Statistical analysis was performed using SPSS 20 (IBM Corp.,
Armonk, NY, USA). The differences between two groups were analyzed
using Student's t-test. P<0.05 was considered to indicate a
significant difference.
Results
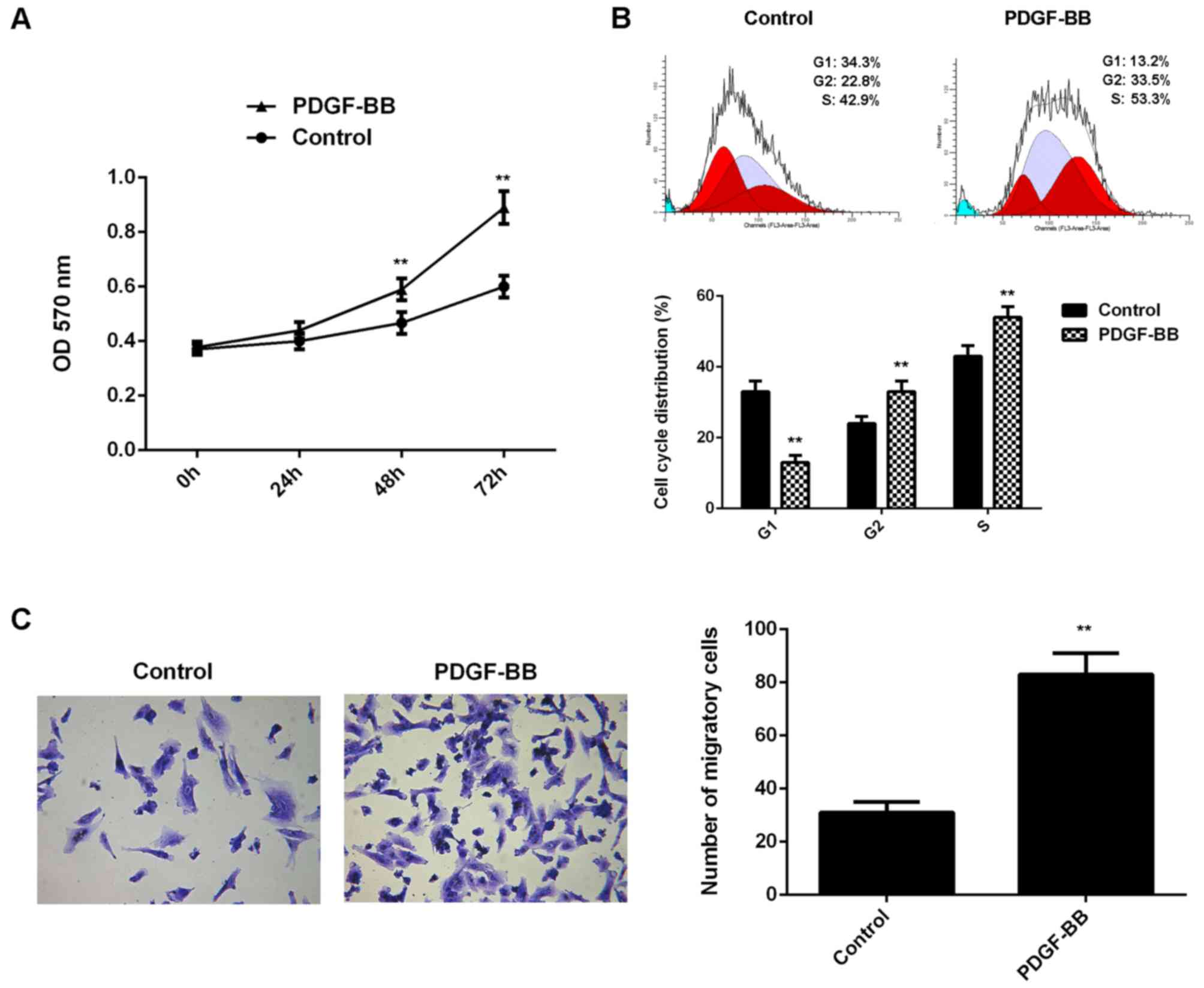
Treatment with PDGF-BB promoted the
proliferation and migration of VSMCs
In the present study, VSMCs in PDGF-BB group were
treated with PDGF-BB for 6 h. VSCMs without any treatment were used
as the control group. Following treatment, the proliferation of
VSMCs was evaluated. As shown in Fig.
1A, the proliferation of VSMCs was significantly increased in
the PDGF-BB group compared with the control group at 48 and 72 h.
Flow cytometry revealed that the percentage of VSMCs at G1 stage
was significantly lower in the PDGF-BB group compared with the
control group, which suggested that treatment with PDGF-BB is able
to promote cell cycle progression (Fig.
1B). Cell migration in each group was subsequently evaluated,
and it was indicated that the migration of VSMCs was significantly
upregulated in the PDGF-BB group when compared with the control
group (Fig. 1C). As such, these
findings indicated that treatment with PDGF-BB promoted the
proliferation and migration of VSMCs.
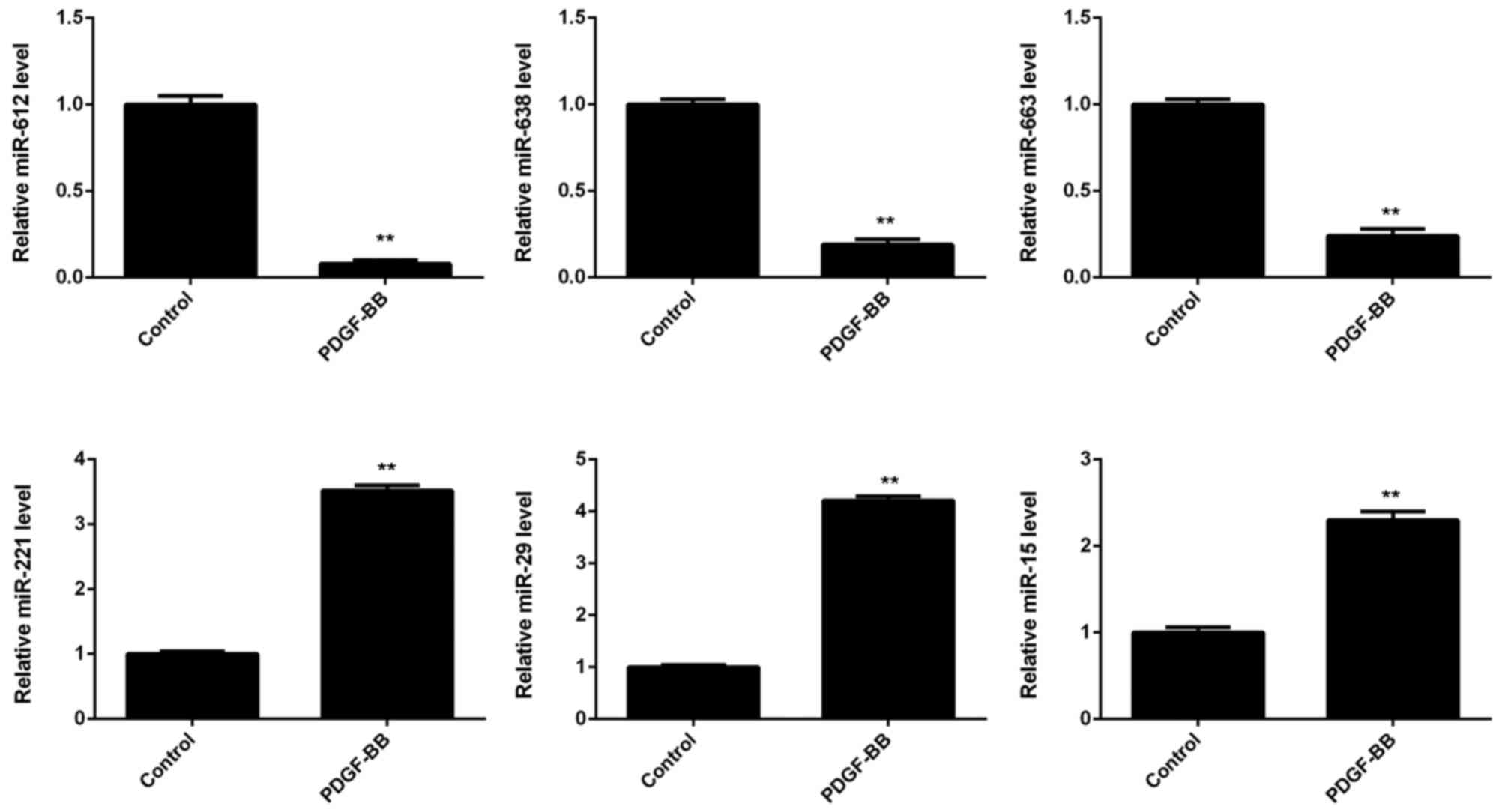
Treatment with PDGF-BB downregulated
miR-612 expression in VSMCs
The expression of several miRs in VSMCs was
subsequently evaluated, with or without PDGF-BB treatment. As shown
in Fig. 2, miR-612, miR-638, and
miR-663 were significantly downregulated in the PDGF-BB group
compared with controls, whereas miR-221, miR-29, and miR-15 were
significantly upregulated. Furthermore, miR-612 demonstrated the
greatest downregulation in VSMCs treated with PDGF-BB, when
compared with the control group (Fig.
2).
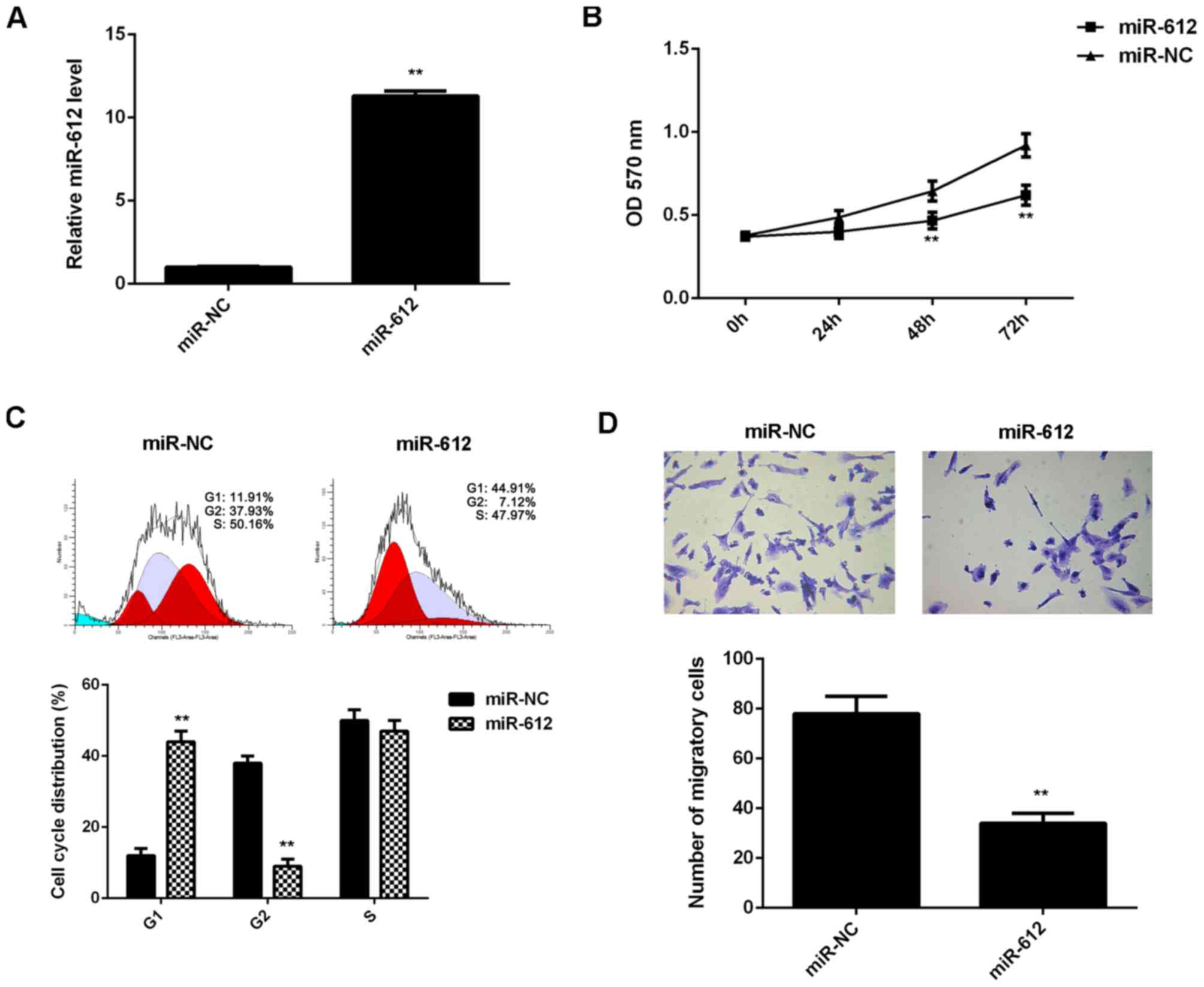
Overexpression of miR-612 attenuated
the proliferation and migration of VSMCs induced by PDGF-BB
treatment
The regulatory effects of miR-612 on the
proliferation and migration of VSMCs induced by PDGF-BB treatment
were then evaluated. VSMCs were transfected with miR-612 mimic or
miR-NC mimic and after transfection the miR-612 levels were
significantly increased in the miR-612 group compared with the
miR-NC group (Fig. 3A). VSMCs in
each group were then treated with PDGF-BB for 6 h. MTT assay data
indicated that the proliferation of VSMCs was significantly reduced
in miR-612 group compared with the miR-NC group at 48 and 72 h
(Fig. 3B). Flow cytometry data
indicated that the cell percentage in the G1 stage was
significantly higher in the miR-612 group compared with the miR-NC
group, suggesting that overexpression of miR-612 led to a
significant cell cycle arrest at G1 stage, which partially
contributes to decreased VSMC proliferation (Fig. 3C). Further investigation revealed
that the migration of VSMCs was also significantly reduced in the
miR-612 group compared with the miR-NC group (Fig. 3D). Therefore, overexpression of
miR-612 attenuated the proliferation and migration of VSMCs induced
by PDGF-BB treatment.
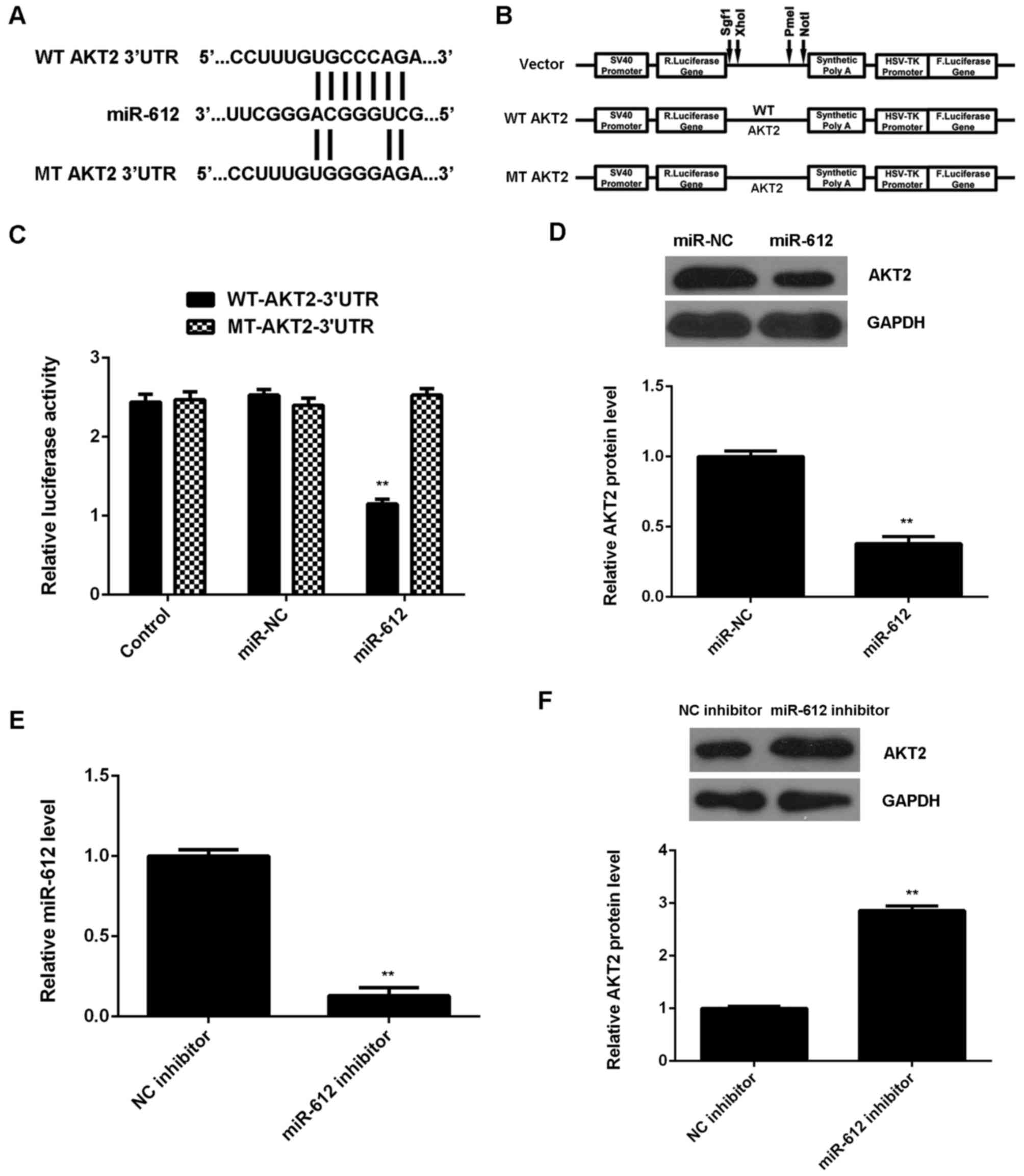
AKT2 is a direct target gene of
miR-612 in VSMCs
As miRs function through regulation of their target
genes, the potential target gene of miR-612 in VSMCs was
investigated. Data from the TargetScan Human 3.1 database indicated
that AKT2 was a putative target gene of miR-612. To confirm this
prediction, WT- and MT-AKT2-3′UTR luciferase reporter plasmids were
constructed (Fig. 4A and B), and a
luciferase reporter gene assay was performed using VSMCs. As shown
in Fig. 4C, the luciferase activity
was significantly downregulated in VSMCs transfected with
WT-AKT2-3′UTR luciferase reporter plasmid and miR-612 mimic, when
compared with the control group; however, this was ameliorated by
transfection with MT-AKT2-3′UTR luciferase reporter plasmid. These
findings indicate that miR-612 directly binds to the 3′UTR of AKT2
mRNA in VSMCs.
The effects of miR-612 on the protein expression of
AKT2 in VSMCs were subsequently evaluated. As shown in Fig. 4D, overexpression of miR-612
significantly reduced the protein levels of AKT2 in VSMCs. To
further confirm these findings, VSMCs were transfected with miR-612
inhibitor, and qPCR data indicated that the miR-612 levels were
significantly decreased in the miR-612 inhibitor group, when
compared with those in the NC inhibitor group (Fig. 4E). Western blot data then showed that
knockdown of miR-612 significantly increased the protein expression
of AKT2 in VSMCs (Fig. 4F).
Together, these data suggest that miR-612 negatively regulates the
protein expression of its target AKT2 in VSMCs.
Overexpression of miR-612 inhibited
the PDGF-BB-induced upregulation of AKT2 in VSMCs
Finally, the molecular mechanism of miR-612 in
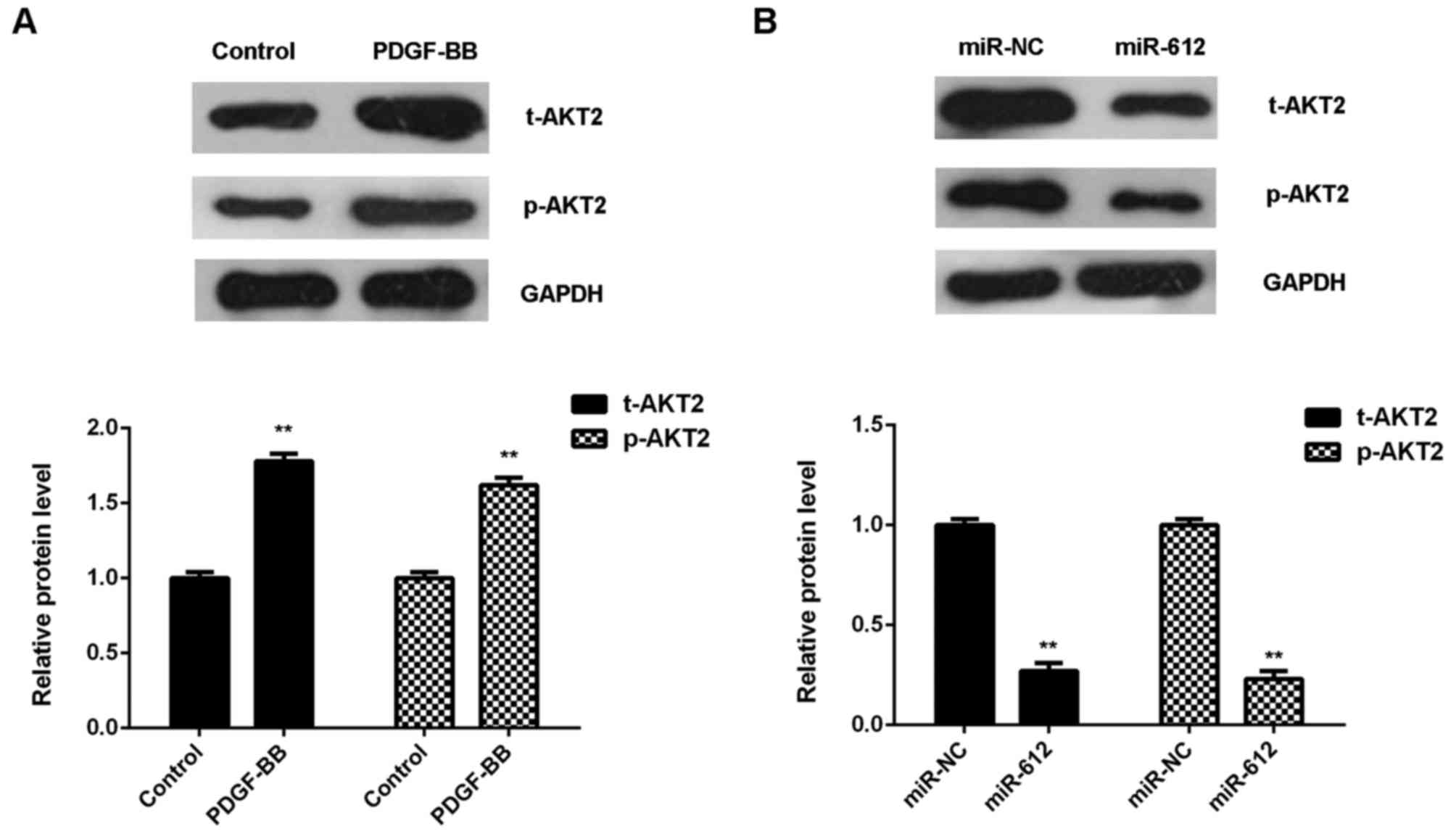
PDGF-BB-treated VSMCs was evaluated. As shown in Fig. 5A, treatment with PDGF-BB
significantly upregulated the protein expression of total AKT2 and
phosphorylated AKT2 in VSMCs, when compared with the control group.
However, overexpression of miR-612 significantly downregulated the
protein expression of t-AKT2 and p-AKT2 in VSMCs treated with
PDGF-BB, when compared with the miR-NC group (Fig. 5B). These findings suggest that
miR-612 has inhibitory effects on the PDGF-BB-induced upregulation
of AKT signaling in VSMCs.
Discussion
Various miRs have been suggested to serve promoting
or suppressive roles in neointimal formation (17,18);
however, the underlying mechanism remains unclear. In the present
study, it was demonstrated that treatment with PDGF-BB
significantly promoted the proliferation and migration of VSMCs,
and decreased the miR-612 levels in VSMCs. Overexpression of
miR-612 significantly inhibited PDGF-BB-induced migration and
invasion of VSMCs, through inducing a cell cycle arrest at G1
stage. AKT2 was further identified as a direct target gene of
miR-612, and its expression was negatively regulated by miR-612 in
VSMCs. Furthermore, overexpression of miR-612 suppressed the
PDGF-BB-induced upregulation of AKT signaling.
The proliferation and migration of VSMCs have
promoting effects on the development of arteriosclerosis and
restenosis, and PDGF has been demonstrated to stimulate VSMC
proliferation and migration through multiple mechanisms, including
the modulation of miR expression (19–21). For
example, miR-15b expression, which is induced by PDGF signaling, is
required for the proliferation of VSMCs (19). In the present study, treatment with
PDGF-BB induced a significant reduction in the expression levels of
several miRs in VSMCs, accompanied by increased cell proliferation
and migration. It was hypothesized that these downregulated miRs
may have a role in VSMC proliferation and migration. As miR-612
exhibited the greatest downregulation, VSMCs were transfected with
miR-612 mimic to upregulate its expression. Ectopic expression of
miR-612 was found to significantly attenuate the proliferation and
migration of VSMCs induced by PDGF-BB treatment, which confirmed
the hypothesis.
As miRs function through regulating the expression
of their target genes (9), potential
targets of miR-612 were evaluated and AKT2 was identified as a
direct target of miR-612 in VSMCs via luciferase reporter gene
assay. AKT2 encodes AKT serine/threonine kinase 2, which is an
important member of PI3K/AKT signaling and capable of
phosphorylating several downstream proteins such as p70S6 kinase,
mechanistic target of rapamycin and glycogen synthase kinase 3
(22,23). A previous study confirmed that AKT2
has a promoting role in VSMC migration and neointimal formation
(24). Furthermore, AKT2 is
associated with CXCL16-triggered platelet activation and adhesion,
and thus may be associated with vascular inflammation and
thrombo-occlusive diseases (25).
Rotllan et al also reported that hematopoietic AKT2
deficiency attenuated the progression of atherosclerosis (26). In the present study, miR-612 was
demonstrated to negatively regulate the protein expression of AKT2
and to inhibit the PDGF-BB-induced upregulation of AKT2 protein
expression in VSMCs. These findings suggest that miR-612 is able to
inhibit the PDGF-BB-induced VSMC proliferation and invasion through
inhibition of AKT2 signaling.
In addition to miR-612, various other miRs have also
been found to serve a suppressive role in VSMC proliferation and/or
migration. For instance, miR-21 inhibits PDGF-induced proliferation
and migration of human aortic VSMCs through inhibiting the
expression of activator protein-1 (27). miR-638, which is highly expressed in
VSMCs, is able to inhibit PDGF-BB-induced cell proliferation and
migration through directly targeting neuron-derived orphan receptor
1 (28). miR-24 inhibits high
glucose-induced VSMC proliferation and migration by targeting high
mobility group box 1 protein (29).
Furthermore, the targeting association between miR-612 and AKT2 has
also previously been found in colon and liver cancers (15,30). For
instance, Sheng et al (15)
recently reported that miR-612 is able to inhibit colorectal cancer
growth and metastasis by targeting AKT2. Accordingly, the present
study expands the understanding of miR-612 functions.
In conclusion, the present study is, to the best of
our knowledge the first study to demonstrate that miR-612 has
suppressive effects on PDGF-BB-induced proliferation and migration
of VSMCs, at least in part, by directly targeting AKT2. These
findings suggest that miR-612 may be a potential candidate for
inhibiting neointimal formation and thus preventing
arteriosclerosis or restenosis following coronary intervention or
vein grafting.
References
|
1
|
Yi N, Chen SY, Ma A, Chen PS, Yao B, Liang
TM and Liu C: Tunicamycin inhibits PDGF-BB-induced proliferation
and migration of vascular smooth muscle cells through induction of
HO-1. Anat Rec (Hoboken). 295:1462–1472. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Zhang C: MicroRNA-145 in vascular smooth
muscle cell biology: A new therapeutic target for vascular disease.
Cell Cycle. 8:3469–3473. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Obata JE, Nakamura T, Kitta Y, Saito Y,
Sano K, Fujioka D, Kawabata K and Kugiyama K: In-stent restenosis
is inhibited in a bare metal stent implanted distal to a
sirolimus-eluting stent to treat a long de novo coronary lesion
with small distal vessel diameter. Catheter Cardiovasc Interv.
82:E777–E787. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Mazurova VV, Sukhorukov OE and Zakharova
OV: Comparing moderately late results of the application of stents
coated with a medicinal antiproliferative agent for the treatment
of patients with various forms of coronary heart disease: Their
efficacy and safety. Klin Med (Mosk). 90:30–35. 2012.(In Russian).
PubMed/NCBI
|
|
5
|
Moss EG: MicroRNAs: Hidden in the genome.
Curr Biol. 12:R138–R140. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Bartel DP: MicroRNAs: Genomics,
biogenesis, mechanism, and function. Cell. 116:281–297. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Zhou W, Zou B, Liu L, Cui K, Gao J, Yuan S
and Cong N: MicroRNA-98 acts as a tumor suppressor in
hepatocellular carcinoma via targeting SALL4. Oncotarget.
7:74059–74073. 2016.PubMed/NCBI
|
|
8
|
Yao J, Deng B, Zheng L, Dou L, Guo Y and
Guo K: miR-27b is upregulated in cervical carcinogenesis and
promotes cell growth and invasion by regulating CDH11 and
epithelial-mesenchymal transition. Oncol Rep. 35:1645–1651. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Ambros V: The functions of animal
microRNAs. Nature. 431:350–355. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Chen LJ, Lim SH, Yeh YT, Lien SC and Chiu
JJ: Roles of microRNAs in atherosclerosis and restenosis. J Biomed
Sci. 19:792012. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Weber C, Schober A and Zernecke A:
MicroRNAs in arterial remodelling, inflammation and
atherosclerosis. Curr Drug Targets. 11:950–956. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Liu X, Cheng Y, Zhang S, Lin Y, Yang J and
Zhang C: A necessary role of miR-221 and miR-222 in vascular smooth
muscle cell proliferation and neointimal hyperplasia. Circ Res.
104:476–487. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Sun SG, Zheng B, Han M, Fang XM, Li HX,
Miao SB, Su M, Han Y, Shi HJ and Wen JK: miR-146a and Krüppel-like
factor 4 form a feedback loop to participate in vascular smooth
muscle cell proliferation. EMBO Rep. 12:56–62. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Tang J, Tao ZH, Wen D, Wan JL, Liu DL,
Zhang S, Cui JF, Sun HC, Wang L, Zhou J, et al: MiR-612 suppresses
the stemness of liver cancer via Wnt/β-catenin signaling. Biochem
Biophys Res Commun. 447:210–215. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Sheng L, He P, Yang X, Zhou M and Feng Q:
miR-612 negatively regulates colorectal cancer growth and
metastasis by targeting AKT2. Cell Death Dis. 6:e18082015.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) Method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Lee J, Lim S, Song BW, Cha MJ, Ham O, Lee
SY, Lee C, Park JH, Bae Y, Seo HH, et al: MicroRNA-29b inhibits
migration and proliferation of vascular smooth muscle cells in
neointimal formation. J Cell Biochem. 116:598–608. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Li TJ, Chen YL, Gua CJ, Xue SJ, Ma SM and
Li XD: MicroRNA 181b promotes vascular smooth muscle cells
proliferation through activation of PI3K and MAPK pathways. Int J
Clin Exp Pathol. 8:10375–10384. 2015.PubMed/NCBI
|
|
19
|
Kim S and Kang H: miR-15b induced by
platelet-derived growth factor signaling is required for vascular
smooth muscle cell proliferation. BMB Rep. 46:550–554. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Gan J, Li P, Wang Z, Chen J, Liang X, Liu
M, Xie W, Yin R and Huang F: Rosuvastatin suppresses
platelet-derived growth factor-BB-induced vascular smooth muscle
cell proliferation and migration via the MAPK signaling pathway.
Exp Ther Med. 6:899–903. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Guan S, Tang Q, Liu W, Zhu R and Li B:
Nobiletin Inhibits PDGF-BB-induced vascular smooth muscle cell
proliferation and migration and attenuates neointimal hyperplasia
in a rat carotid artery injury model. Drug Dev Res. 75:489–496.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Frias A, Lambies G, Viñas-Castells R,
Martínez-Guillamon C, Dave N, García de Herreros A and Díaz VM: A
switch in Akt isoforms is required for notch-induced Snail1
expression and protection from cell death. Mol Cell Biol.
36:923–940. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Shiratsuchi H and Basson MD: Akt2, but not
Akt1 or Akt3 mediates pressure-stimulated serum-opsonized latex
bead phagocytosis through activating mTOR and p70 S6 kinase. J Cell
Biochem. 102:353–367. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Zhang W, Zhang X, González-Cobos JC,
Stolwijk JA, Matrougui K and Trebak M: Leukotriene-C4 synthase, a
critical enzyme in the activation of store-independent Orai1/Orai3
channels, is required for neointimal hyperplasia. J Biol Chem.
290:5015–5027. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Borst O, Münzer P, Gatidis S, Schmidt EM,
Schönberger T, Schmid E, Towhid ST, Stellos K, Seizer P, May AE, et
al: The inflammatory chemokine CXC motif ligand 16 triggers
platelet activation and adhesion via CXC motif receptor 6-dependent
phosphatidylinositide 3-kinase/Akt signaling. Circ Res.
111:1297–1307. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Rotllan N, Chamorro-Jorganes A, Araldi E,
Wanschel AC, Aryal B, Aranda JF, Goedeke L, Salerno AG, Ramírez CM,
Sessa WC, et al: Hematopoietic Akt2 deficiency attenuates the
progression of atherosclerosis. FASEB J. 29:597–610. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Li Y, Yan L, Zhang W, Hu N, Chen W, Wang
H, Kang M and Ou H: MicroRNA-21 inhibits platelet-derived growth
factor-induced human aortic vascular smooth muscle cell
proliferation and migration through targeting activator protein-1.
Am J Transl Res. 6:507–516. 2014.PubMed/NCBI
|
|
28
|
Li P, Liu Y, Yi B, Wang G, You X, Zhao X,
Summer R, Qin Y and Sun J: MicroRNA-638 is highly expressed in
human vascular smooth muscle cells and inhibits PDGF-BB-induced
cell proliferation and migration through targeting orphan nuclear
receptor NOR1. Cardiovasc Res. 99:185–193. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Yang J, Chen L, Ding J, Fan Z, Li S, Wu H,
Zhang J, Yang C, Wang H, Zeng P and Yang J: MicroRNA-24 inhibits
high glucose-induced vascular smooth muscle cell proliferation and
migration by targeting HMGB1. Gene. 586:268–273. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Tao ZH, Wan JL, Zeng LY, Xie L, Sun HC,
Qin LX, Wang L, Zhou J, Ren ZG, Li YX, et al: miR-612 suppresses
the invasive-metastatic cascade in hepatocellular carcinoma. J Exp
Med. 210:789–803. 2013. View Article : Google Scholar : PubMed/NCBI
|