Introduction
Osteoarthritis (OA) is a heterogeneous and
multifactorial degenerative joint disease characterized by joint
pain, cartilage degradation and osteophyte formation. Approximately
86% of the societal costs of arthritis are estimated to be
attributable to OA (1) however,
there are currently no therapeutic agents available that halt the
progression of articular cartilage degeneration. Non-surgical
treatments for OA, including non-steroid anti-inflammatory drugs,
oral steroid or knee steroid injection provide temporary
symptomatic relief only (2).
Therefore, the development of a structural OA disease-modifying
drug that provides symptomatic relief and arrests the progression
of cartilage degeneration is required. The lack of progress in the
development of structural OA disease-modifying drugs is primarily
due to limited understanding of the pathogenesis of OA and
insufficient knowledge regarding the molecular targets for
therapeutic intervention.
The biochemical events associated with the
initiation or progression of OA are poorly understood.
Extracellular matrix-degrading enzymes and inflammatory cytokines,
including matrix metollproteinase-13 (MMP-13), ADAM
metallopeptidase with thrombospondin type 1 motif 5, interleukin-1
and cyclooxygenase-2 have been implicated in OA (3–5). The
activation and abnormal phenotypic change of articular chondrocytes
have also been implicated. Chondrocytes in OA articular cartilage
exhibit an aberrant phenotype (6–8),
although the exact mechanisms of chondrocyte activation and
differentiation in OA articular cartilage remain unclear.
Age-associated changes in extracellular matrix molecules or
expression of inflammatory cytokines and growth factors may serve a
role in the activation and differentiation of articular
chondrocytes (7–9). Alternatively, articular chondrocytes
may obtain properties with similarities to those of terminal
differentiating chondrocytes in the growth plate. Terminal
differentiating OA articular chondrocytes are associated with
calcification, apoptosis, neovascularization and ossification
(7–11) and may participate in articular
cartilage degeneration by secreting large amounts of MMP-13
(12–14).
The number of tidemarks, the lines bordering the
calcified cartilage and non-calcified cartilage, increases with age
in human articular cartilage (14).
The movement of tidemarks toward the articular surface is expected
to result in an increase in the thickness of calcified zone
cartilage, however, the calcified zone cartilage instead becomes
thinner during aging (14). This
phenomenon indicates replacement of the calcified cartilage by
subchondral bone in the aging process. OA is also associated with
articular cartilage thinning (15,16).
This association suggests that the replacement of calcified
cartilage by subchondral bone may serve a role in articular
cartilage thinning or cartilage degeneration in OA (17).
Phosphocitrate (PC) is a powerful calcification
inhibitor (18), which prevents soft
tissue calcification (19). PC also
inhibits calcium crystal-induced cell membrane damages,
mitogenesis, expression of extracellular matrix-degrading enzymes
and crystal-induced cell death (20–23).
Based on these results, a hypothesis that PC is a potentially
disease-modifying drug for calcium crystal-associated primary OA
was previously postulated (24). A
subsequent study demonstrated that PC inhibited meniscal
calcification in the Hartley guinea pigs and the reduction in
meniscal calcification was accompanied with decreased articular
cartilage degeneration (25).
It was previously demonstrated that, in cell
cultures, PC inhibited the expression of numerous genes implicated
in OA in the absence of calcium crystals (26–28).
These previous results indicate that PC is a disease modifying drug
for calcium crystal-associated primary OA and injury-induced
secondary posttraumatic OA therapy. However, the molecular
mechanisms underlying the OA disease-modifying activity of PC
remain unclear. The present study aimed to investigate the
hypothesis that PC exerts its OA disease-modifying effect, in part,
by inhibiting a molecular program similar to the endochondral
pathway of ossification.
Materials and methods
Reagents
Safranin O-fast green and hematoxylin and eosin
(H&E) were obtained from Polysciences, Inc. (Warrington, PA,
USA). Antibodies specific to MMP-13 (cat. no. LS-B3168) and type X
collagen (COL-X; cat. no. ab49945) were obtained from LifeSpan
BioSciences, Inc. (Seattle, WA, USA) and Abcam (Cambridge, MA,
USA), respectively. Antibodies specific to runt-related
transcription factor 2 (cat. no. bs-1134R) and sex determining
region Y-box 9 (cat. no. TA335171) were obtained from Bioss
Antibodies, Inc. (Woburn, MA, USA) and OriGene Technologies, Inc.
(Rockville, MD, USA), respectively. Mouse immunoglobulin G was
obtained from BioGenex (Fremont, CA, USA; cat. no. HK119-7M).
Secondary reagent (ImmPRESS reagent kit) was obtained from Vector
Laboratories, Inc. (Burlingame, CA, USA; cat. no. MP-7401-50 ml).
PC and PC-β ethyl ester (PC-E) were synthesized according to
previously detailed procedures (29). All other reagents, including normal
horse serum, bovine serum albumin and 3,3′-diaminobenzidine, were
obtained from Sigma-Aldrich (Merck KgaA, Darmstadt, Germany).
Animals
Hartley guinea pigs develop OA spontaneously 3
months following birth and reach skeletal maturity at the age of ~6
months (30–32). The progressive degenerative changes
in the articular cartilage closely resemble the degenerative
changes in human OA articular cartilage; therefore, Hartley guinea
pigs have been widely used to study the pathogenesis of OA and test
disease modifying drugs (33–37).
Treatment of Hartley guinea pigs with PC and PC-E has been
previously described (38). Briefly,
young male Hartley guinea pigs (mean body weight, 280 g; Charles
River Laboratories, Inc., Wilmington, MA, USA) were housed
individually in solid bottom cages and maintained in a
temperature-controlled room (18–20°C) that was regulated on a 12-h
light-dark cycle. Relative humidity of the room was maintained
between 45–55%. Guinea Pig Chow (cat. no. 5025; Ralston Purina Co.,
Chicago, IL, USA) and water were available ad libitum.
Following acclimation, 15 guinea pigs (age, 5 weeks) were randomly
divided into three groups. The first group (n=5) received
intraperitoneal injection of PC (40 mg/kg) twice per week for a
period of 2 months, the second group (n=5) received intraperitoneal
injection of PC-E (40 mg/kg) twice per week for a period of 2
months, and the last group (n=5) received intraperitoneal injection
of physiological saline twice per week for 2 months. Partial medial
meniscectomy was then performed on the right knee of all guinea
pigs. At 1 week following surgery, administration of treatment
(twice per week at the same dose) was resumed in each group and
continued for a further 5 months. Guinea pigs were then euthanized.
Hind limbs were collected, fixed in 10% formalin for 24 h at room
temperature and transferred to 70% ethanol prior to further use.
The present study was performed according to the guidelines set
forth by the Institutional Animal Care & Use Committee of
Carolinas Medical Center (Charlotte, NC, USA), which also approved
the animal protocol.
Histological examinations
The tibia plateaus of partial meniscectomied right
knees were decalcified in a formic acid/sodium citrate solution
(Thermo Fisher Scientific, Inc., Waltham, MA, USA) and cut in the
coronal plane to produce two equal parts. The posterior part was
embedded in paraffin and sectioned with a Leica RM2025 microtome
(Leica Microsystems GmbH, Wetzlar, Germany) to obtain 4-µm-thick
sections. A total of 3 non-consecutive sets of sections obtained at
400-β m intervals from each tibia plateau (9 total sections
covering the central most degenerated area of the medial tibial
plateau) were stained with Safranin-O for 5 min and countered
stained with fast green for 5 min at room temperature. A total of 2
sections from each tibial plateau were also stained with H&E
for 10 min at room temperature. Safranin-O-fast green-stained
sections were graded according to criteria described previously
(39) with modifications, as
detailed in Table I.
 | Table I.Semi-quantitative histological
grading scheme for medial tibia plateau of the guinea pig. |
Table I.
Semi-quantitative histological
grading scheme for medial tibia plateau of the guinea pig.
| Parameter | Grade | Description |
|---|
| Cartilage
structure | 0 | Normal, smooth,
uninterrupted surface |
|
| 1 | Mild surface
irregularities, no clefts |
|
| 2 | Irregular surface,
1–3 superficial clefts |
|
| 3 | >3 clefts and/or
loss of cartilage to superficial zone |
|
| 4 | 1–3 clefts
extending into the middle zone |
|
| 5 | >3 clefts and/or
loss of cartilage extending into the middle zone |
|
| 6 | 1–3 clefts
extending into the deep zone |
|
| 7 | >3 clefts
extending into the deep zone and/or loss (5–20%) of cartilage to
deep zone |
|
| 8 | Fissures or loss of
cartilage extended to the zone of calcified cartilage |
| Safranin-O
staining | 0 | Uniform staining
throughout articular cartilage |
|
| 1 | Loss of staining in
superficial zone only and for less than half the length of the
plateau |
|
| 2 | Loss of staining in
superficial zone for half the length or greater of the plateau |
|
| 3 | Loss of staining in
superficial and middle zones for less than half the length of the
plateau |
|
| 4 | Loss of staining in
superficial and middle zones for half the length or greater |
|
| 5 | Loss of staining in
all three zones for less than half the length of the condyle or
plateau |
|
| 6 | Loss of staining in
all three zones for half the length or greater of the plateau |
| Cellularity | 0 | Uniform dense
chondrocytes in the superficial zone |
|
| 1 | Cell density is
reduced in the superficial zone |
|
| 2 | Cell density is
reduced in both superficial zone and middle zone |
|
| 3 | Cell density is
also reduced in deep zone |
| Tidemark | 0 | Faint tidemark |
|
| 1 | Moderate visible
tidemark |
|
| 2 | Strong visible
tidemark |
| Subchondral bone
advance | 0 | Distance between
subchondral bone front and tidemark is large (away more than 4
cells) |
|
| 1 | Distance between
subchondral bone front and tidemark is moderate (roughly 3–4
cells) |
|
| 2 | Distance between
subchondral bone front and tidemark is small (2 cells) |
|
| 3 | Distance between
subchondral bone front and tidemark is extremely small (1
cell) |
|
| 4 | Subchondral bone
front reaches to tidemark. |
Growth plate thickness
measurement
Growth plate thickness was determined using ImageJ
photo analysis software (version 1.8.0_45; National Institutes of
Health, Bethesda, MD, USA). Briefly, images of Safranin-O-fast
green-stained sections (3 sections from each tibial plateau) were
captured at ×4 magnification with a digital camera (Sony DXC-S500;
Sony Corporation, Tokyo, Japan) equipped with a light microscope
(Nikon Optiphot-2; Nikon Corporation, Tokyo, Japan). The growth
plate and growth plate cartilage bars or islands (red colored)
embedded within the subchondral bone were selected using the
following image thresholding and LAB color space parameters (L:
0/255, A: 145/255, B: 0/255). Following selection, the areas of
growth plate (particle size setting, ≥0.1 mm2) and
growth plate cartilage bars (particle size setting, ≥0.0001
mm2 and <0.1 mm2) were measured. Growth
plate thickness was calculated by dividing the measured area of
growth plate with the length of the growth plate or by dividing the
measured area of growth plate + the measured area of growth plate
bars by the length of the growth plate. Normalized growth plate
thickness was used in all analyses, with the growth plate thickness
in the untreated guinea pigs set as 1.
Immunostaining
A total of 2 sections from each tibial plateau were
deparaffinized with xylene and rehydrated with graded ethanol.
Endogenous peroxidase activity was blocked via incubation of these
sections with deionized water containing 3%
H2O2 for 15 min at room temperature.
Non-specific binding was blocked via incubation with 100 l of 10%
normal horse serum diluted in base solution (4% bovine serum
albumin and 5% non-fat dry milk in phosphate buffered saline) for
20 min at room temperature. Sections were subsequently incubated
with primary antibodies (1:100) for 1 h at room temperature,
followed with secondary reagent (1:200) for 30 min at room
temperature. A negative control was performed using mouse
immunoglubulin G. Slides were rinsed with PBS three times,
visualized with 3,3′-diaminobenzidine for 5 min at room temperature
and counterstained with light green for 5 min at room temperature.
Slides were viewed under light microscopy and graded on a scale of
0–4 as described previously (38,40).
Briefly, 0, very weak staining; 1, weak staining; 2, moderate
staining; 3, strong staining; and 4, very strong staining.
Statistical analysis
All data, including histological scores of articular
cartilages, growth plate thickness, articular cartilage thickness,
total tissue volume, immunostaining scores of COL-X and MMP13 and
the number of marrow spaces, were presented as the mean ± standard
deviation. The differences between groups were analyzed using
one-way analysis of variance followed by a Tukey or Dunnett's test.
Statistical analysis was performed using the statistical analysis
tool in Sigma Plot software (version 12; Systat Software, Inc., San
Jose, CA, USA). P<0.05 was considered to indicate a
statistically significant difference.
Results
PC and PC-E reduce the histological
score of articular cartilage
Representative Safranin-O-fast green-stained
sections are presented in Fig. 1A.
Sections were previously examined and it was demonstrated that PC
and PC-E inhibited articular cartilage degeneration in the guinea
pig model of posttraumatic OA (38).
Further examination of these sections in the present study
indicated further differences between the articular cartilage in
the untreated and PC-treated guinea pigs. The tidemark in untreated
guinea pigs was more visible and closer to the articular surface
compared with the tidemark in PC-treated guinea pigs. In addition,
severe proteoglycan loss occurred in the superficial and middle
zones, as well as in the calcified zone in the untreated guinea
pigs, but not in the PC-treated guinea pigs. There were also fewer
articular cartilage bars or islands embedded within the subchondral
bone in the untreated guinea pigs compared with PC-treated guinea
pigs. These differences indicated that the advance of calcification
front and active replacement of the calcified cartilage by
subchondral bone occurred in the Hartley guinea pigs and that PC
inhibited this process. Therefore, subchondral bone advance toward
articular cartilage surface or the tidemark may serve a role in
articular cartilage thinning or destruction. Similar differences
were also observed between the articular cartilage in untreated and
PC-E-treated guinea pigs (data not shown).
Representative H&E stained sections are
presented in Fig. 1B. The
subchondral bone was stained dark pink and articular cartilage was
stained pink or light pink. Similar to Safranin-O-fast green
staining, H&E staining demonstrated that the tidemark in the
untreated guinea pigs was more visible and closer to articular
cartilage surface compared with the tidemark in PC-treated guinea
pigs. The superficial, middle, deep and calcified zones of
articular cartilage in PC-treated guinea pigs were stained light
pink. By contrast, only the middle and deep zones in the untreated
guinea pigs were stained light pink, whereas the superficial zone
and the entire calcified zone were stained pink. These differences
indicated that the extracellular matrices within the superficial
and calcified zones in the untreated guinea pigs differed from
those in PC-treated guinea pigs, suggesting that articular
cartilage degeneration or matrix loss occurred in the superficial
and calcified zones in the untreated guinea pigs. Chondrocyte
clusters were observed in the articular cartilage of untreated
guinea pigs, but rarely in the articular cartilage of PC-treated
guinea pigs. In addition, there were fewer chondrocytes within the
articular cartilage in the untreated guinea pigs compared with
PC-treated guinea pigs. These results are consistent with previous
studies that have demonstrated that chondrocytes were activated and
chondrocyte apoptosis was prominent in OA articular cartilage
(6,41–43).
All safranin-O-fast green-stained sections were
graded. PC treatment significantly reduced the histological score
of the medial tibia plateau cartilage, resulting in a 46% reduction
in the histological score, graded according to the criteria
described previously (39) and a 50%
reduction in the histological score, graded according to the
criteria described in Table I,
compared with the untreated control (Fig. 2A). PC-E treatment also reduced the
histological scores, graded according to the criteria described
previously (39) and the criteria
described in Table I, significantly
compared with the untreated control, however, the reductions were
less marked compared with PC in the inhibition of articular
cartilage degeneration (Fig. 2A).
Select sub-category histological scores in the untreated, PC- and
PC-E-treated guinea pigs are presented in Fig. 2B. PC treatment resulted in a 37, 82
and 65% significant reduction in cellularity, tidemark and
subchondral bone advance sub-scores, respectively. By contrast,
PC-E treatment only resulted in a significant reduction in tidemark
and subchondral sub-scores compared with the untreated control,
whereas the difference in the sub-histological score for
cellularity between the control and PC-E treated groups was not
significant, which indicated reduced disease-modifying activity
compared with PC treatment.
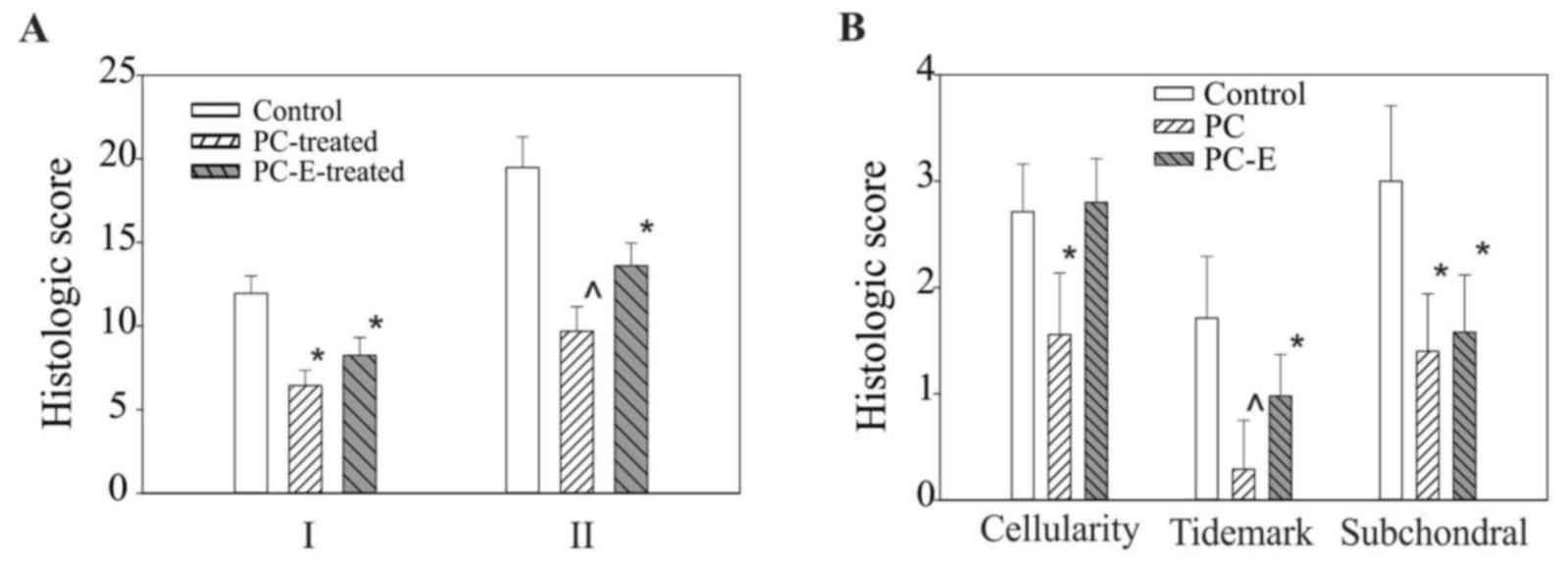 | Figure 2.Histological scores of medial tibial
plateaus cartilage. (A) I, Histologic scores, graded according to
the criteria described previously (39), in untreated, PC-, and PC-E-treated
guinea pigs were 11.95±1.05, 6.43±0.93 and 8.24±1.07, respectively.
II, Histologic scores, graded according to the criteria listed in
Table I, in untreated, PC- and
PC-E-treated guinea pigs were 19.47±1.83, 9.68±1.47 and 13.60±1.37,
respectively. (B) Sub-category histological scores for cellularity,
tidemark and subchondral bone advance in untreated, PC-, and
PC-E-treated guinea pigs. *P<0.05 vs. control; ^P<0.05 vs.
control and PC-E. PC, phosphocitrate; PC-E, PC-β ethyl ester. |
PC and PC-E inhibit growth plate
thinning
In addition to cartilage degeneration, PC also
inhibited growth plate thinning. The growth plate in PC-treated
guinea pigs was markedly thicker than the growth plate in untreated
guinea pigs (Fig. 3A). In addition,
there were more growth plate cartilage bars or islands embedded
within the metaphyseal bone in PC-treated guinea pigs compared with
untreated guinea pigs. A similar phenomenon was observed in
PC-E-treated guinea pigs (data not shown). The differences were
quantified by measuring growth plate thickness using ImageJ
software. The growth plates in PC- and PC-E-treated guinea pigs
were 54 and 36% thicker compared with the growth plate in untreated
guinea pigs, respectively (Fig. 3B).
If the area of growth plate cartilage bars was also included in the
calculation the growth plate in the PC- and PC-E-treated guinea
pigs was 97 and 41% thicker compared with the growth plate in
untreated guinea pigs, respectively.
PC and PC-E appeared to inhibit the conversion of
growth plate cartilage into bone, therefore, the effects of
treatment on metaphyseal bone was investigated by measuring the
areas of bone matrices under the growth plate using ImageJ
software. The differences between the areas of bone matrices, an
indirect indicator of bone volume, in the untreated and PC- or
PC-E-treated guinea pigs were not significant (Fig. 3C). However, the total areas of
matrices (areas of bone matrices + areas of growth plate cartilage
bars or islands) in the PC-or PC-E-treated guinea pigs was
significantly greater than that in the untreated guinea pigs
(Fig. 3C).
Analyses using ImageJ software were also performed
to determine the thickness of articular cartilage. The articular
cartilage in the central most degenerated area of the medial tibial
plateau cartilage in the PC- and PC-E-treated guinea pigs was 40
and 29% thicker compared with that in untreated guinea pigs,
respectively and the differences were significant (Fig. 3D). If the area of articular cartilage
bars or islands was also included in the calculation the articular
cartilage in the central most degenerated area of the medial tibial
plateau cartilage in the PC- and PC-E-treated guinea pigs was 53
and 38% thicker compared with the articular cartilage in the
untreated guinea pigs, respectively.
PC and PC-E inhibit vascular invasion
into growth plate
Representative H&E-stained sections of growth
plates are presented in Fig. 4. The
growth plate in the untreated guinea pigs was markedly thinner and
had fewer growth plate cartilage bars compared with that in the PC
or PC-E treated guinea pigs (Fig.
4A). In addition, there was an increased number of marrow
spaces or vascular channels at the junction between the growth
plate and metaphysis in the untreated guinea pigs compared with PC-
or PC-E-treated guinea pigs. The number of marrow spaces is an
indirect indicator of vascular invasion into the growth plate. The
PC or PC-E treatment resulted in reduced vascular invasion into the
growth plate compared with the untreated group (Fig. 4B). There were 39 and 18% fewer marrow
spaces or vascular channels at the junction in the PC- and PC-E
treated guinea pigs compared with the untreated guinea pigs,
respectively and these differences were significant.
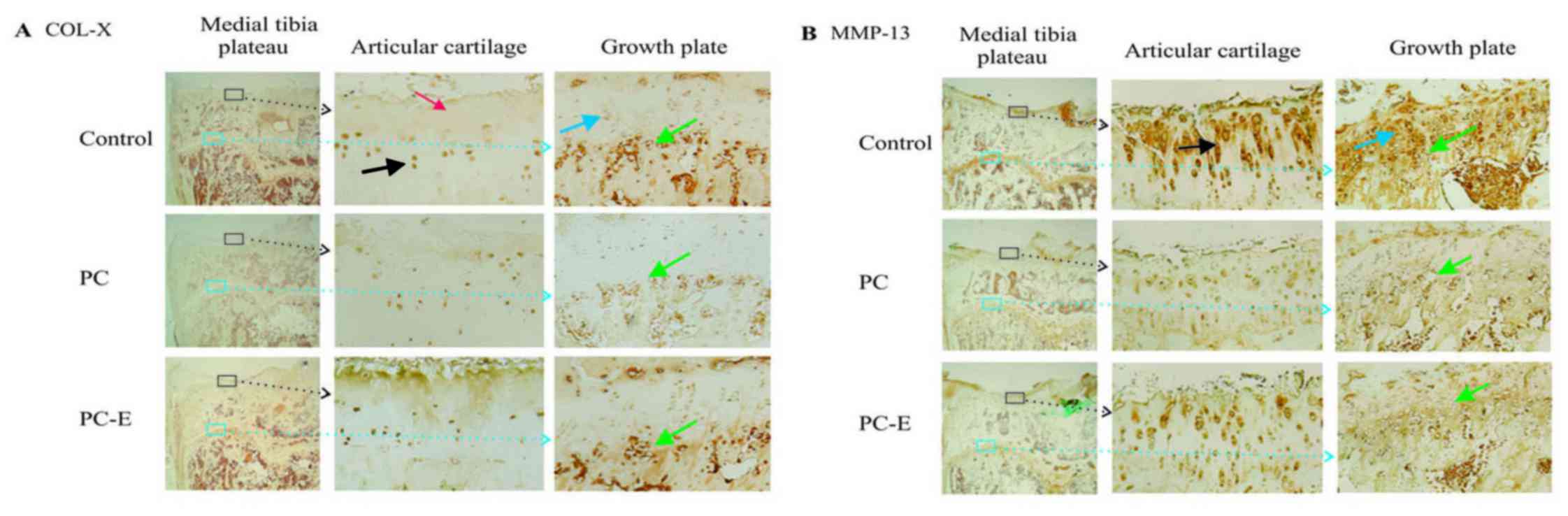
PC and PC-E reduce the levels of COL-X
and MMP-13 protein
Representative immunostaining images of the
expression of COL-X are presented in Fig. 5A. High levels of COL-X were detected
in the nucleus and cytoplasm of articular chondrocytes in the
untreated guinea pigs. These positively stained chondrocytes were
primarily located in the deep and calcified zones. Moderate or low
levels of COL-X were detected in the extracellular matrices within
the superficial and middle zones in the untreated guinea pigs.
COL-X was also detected in the nucleus and cytoplasm of
chondrocytes in the growth plate cartilage. The positively stained
growth plate chondrocytes were primarily located in the maturation
and hypertrophic zones. COL-X was also detected in the cells
located at the junction between the growth plate and metaphyseal
bone in the untreated guinea pigs. PC treatment reduced the levels
of COL-X in the articular chondrocytes located in the deep and
calcified zones, however, it had little effect on the levels of
COL-X in the extracellular matrices within the superficial and
middle zones. PC treatment reduced the levels of COL-X in the
growth plate chondrocytes located in the maturation zone,
hypertrophic zone and at the junction between growth plate and
metaphyseal bone, as well as the levels of COL-X in the
extracellular matrices within the middle zone compared with
untreated samples. PC-E displayed minor inhibitory effects on the
levels of COL-X.
Representative immunostaining images of the
expression of MMP-13 protein are presented in Fig. 5B. MMP-13 protein was detected in the
nucleus and cytoplasm of articular chondrocytes and pericellular
and extracellular areas in the untreated guinea pigs. The highest
levels of MMP-13 protein were detected in the superficial, middle
and deep zones. By contrast, the highest levels of MMP-13 protein
in the growth plate were detected in the maturation and
hypertrophic zones. Consistent with its OA disease-modifying
activities, PC treatment reduced the levels of MMP-13 protein in
the articular cartilage and growth plate compared with the
untreated group. In addition, consistent with its reduced
inhibitory effects on cartilage degeneration, PC-E exhibited
reduced inhibitory effects on the levels of MMP-13 protein compared
with PC. The levels of runt-related transcription factor 2 and sex
determining region Y-box 9 were also investigated, however, the
differences in the levels of these proteins between the untreated
and PC-treated guinea pigs were not significant (data not
shown).
Immunostaining scores of COL-X and MMP13 proteins
are presented in Fig. 6. The
immunostaining scores of COL-X in the articular chondrocytes and
cells located at the junction between the growth plate and
metaphyseal bone in the PC-treated guinea pigs were decreased by 32
and 37% compared with the scores in untreated guinea pigs,
respectively and the differences were significant. The
immunostaining scores of COL-X in PC-E-treated guinea pigs were
also decreased compared with the score of COL-X in untreated guinea
pigs, however, the differences were not significant. Immunostaining
scores of MMP-13 protein in the articular chondrocytes and cells
located within the hypertrophic zone of growth plate in the
PC-treated guinea pigs were decreased by 49 and 37% compared with
the score in the untreated guinea pigs, respectively and the
differences were significant (Fig.
6B). The immunostaining scores of MMP-13 protein in
PC-E-treated guinea pigs were also decreased compared with that in
untreated guinea pigs, however, only the difference between the
immunostaining score of MMP-13 protein in the articular chondrocyte
was significant.
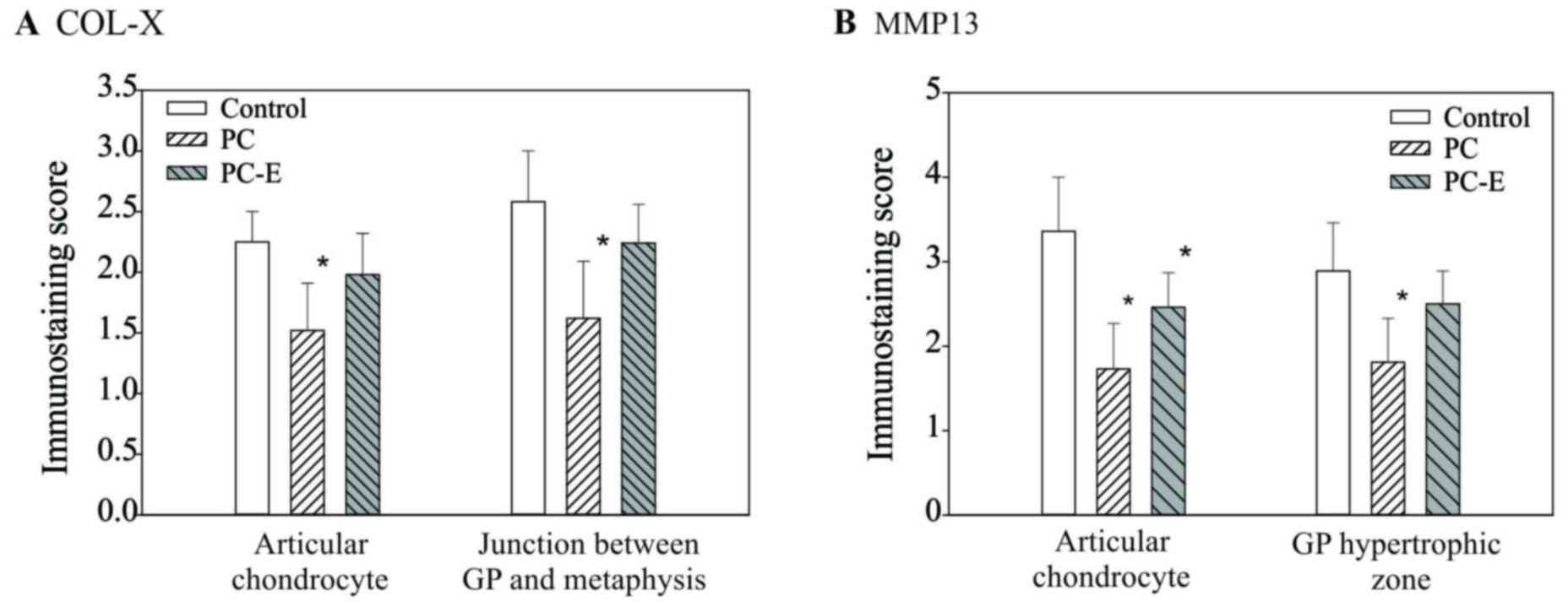 | Figure 6.Immunostaining scores of the levels
of COL-X and MMP13 proteins. (A) Scores for COL-X in articular
chondrocytes in the control, PC- and PC-E-treated guinea pigs were
2.25±0.26, 1.52±0.39 and 1.98±0.34, respectively. Scores for COL-X
protein in the cells located at the junction between the growth
plate and metaphyseal bone in the control, PC- and PC-E-treated
guinea pigs were 2.58±0.42, 1.62±0.49 and 2.24±0.32, respectively.
(B) Scores for MMP-13 protein in the articular chondrocytes in the
control, PC- and PC-E-treated guinea pigs were 3.36±0.64, 1.73±0.55
and 2.46±0.41, respectively. Scores for MMP-13 protein in the cells
located in hypertrophic zone of the growth plate in the untreated,
PC- and PC-E-treated guinea pigs were 2.89±0.56, 1.81±0.52 and
2.50±0.39, respectively. *P<0.05 vs. control. COL-X, type X
collagen; MMP-13, matrix metollproteinase-13; PC, phosphocitrate;
PC-E, PC-β ethyl ester. |
Discussion
The primary function of the calcified zone is to
anchor articular cartilage to the subchondral bone (44). The current study demonstrated that
severe proteoglycan loss occurred in the superficial and middle
zones, as well as in the calcified zone. In addition, there were
fewer articular cartilage bars and islands embedded within the
subchondral bone in the untreated guinea pigs compared with the PC-
or PC-E-treated guinea pigs. These results indicated that articular
cartilage degeneration progressed from the articular surface and
calcified zone toward the inner zones in Hartley guinea pigs.
Degenerative insults to articular cartilage came from the articular
side and epiphyseal side. It is conceivable that degenerative
insults from the epiphyseal side may not only result in articular
cartilage thinning but may also weaken the connection between the
calcified cartilage and the underlying subchondral bone. It appears
that the advance of calcification front and subsequent ossification
or subchondral bone advance towards articular cartilage surface
serves a role in articular cartilage thinning or articular
cartilage destruction. This mechanism was investigated further by
determining the effects of PC and PC-E on the growth plate. The
results indicated that PC and PC-E inhibited growth plate thinning.
The differences in growth plate thickness between the treatment
groups and the control group were larger in the PC group compared
with the PC-E group, suggesting that PC has a larger effect in
inhibiting growth plate thinning, which was consistent with the
inhibitory activities of PC or PC-E on articular cartilage
thinning. This consistency suggests that a similar molecular
mechanism may be associated with the inhibition of articular
cartilage thinning and growth plate thinning by PC and its analogue
PC-E.
Hypertrophic differentiation of growth plate
chondrocytes and vascular invasion into the growth plate are key
processes in the endochondral pathway of ossification. The current
study demonstrated that high levels of two specific hypertrophic
markers COL-X and MMP-13 protein, were detected in the hypertrophic
zone and at the junction between the growth plate and metaphyseal
bone. Their levels were decreased in the PC- or PC-E-treated guinea
pigs, which indicated that PC and its analogue may inhibit
hypertrophic differentiation of growth plate chondrocytes. In
addition, there were more marrow spaces or vascular invasions at
the junction between the growth plate and metaphyseal bone in the
untreated guinea pigs compared with PC- or PC-E-treated guinea
pigs. Consistent with their inhibitory activities on vascular
invasion, PC had a greater effect on the inhibition of growth plate
thinning compared with PC-E. Taken together, these results indicate
that PC and its analogue PC-E inhibit endochondral pathway of
ossification.
COL-X and MMP-13 protein were also detected in
articular cartilage. The highest levels of COL-X were present in
the articular chondrocytes located in the deep and calcified zones.
These results, together with the findings that there were fewer
articular cartilage bars or islands embedded within the subchondral
bone in the untreated guinea pigs compared with the PC- or
PC-E-treated guinea pigs, indicated that a molecular program
similar to the endochondral pathway of ossification occurred in the
articular cartilage. This molecular program is responsible for the
degenerative insults to the articular cartilage from the epiphyseal
side in the control guinea pigs. COL-X was also detected in the
superficial and middle zone matrices, which suggested that this
molecular program is also associated with the degenerative insults
to the articular cartilage from the articular side. Therefore, it
may be deduced that PC and its analogue inhibited articular
cartilage thinning or destruction, in part, by inhibiting the
re-activated molecular program in the OA-prone Hartley guinea
pigs.
The highest levels of MMP-13 protein were detected
in the superficial, middle and deep zones of articular cartilage.
OA synovial fluid contains high levels of MMP-13 protein (45–47) and
may be one of the sources for the superficial zone MMP-13 protein
through diffusion. The strong inhibitory effect of PC on the levels
of MMP-13 protein in the superficial and middle zones may explain
why PC had little effect on the levels of COL-X within the
extracellular matrices in these zones. Decreased MMP-13 activity in
these zones due to PC treatment may result in reduced COL-X
degradation (13) and therefore,
preserved COL-X content in these zones.
The larger volume of growth plate cartilage in the
PC-treated guinea pigs compared with the control guinea pigs
suggested that PC inhibited the conversion of growth plate
cartilage into bone. It was expected that there would be fewer bone
matrices in the PC-treated guinea pigs compared with the untreated
guinea pigs. However, this was not what was observed. A possible
explanation for this contradiction is that PC may have a strong
stimulatory activity on the production of extracellular matrices by
chondrocytes (27,28) and osteoblasts. If the stimulatory
activity of PC is a dominant activity over its inhibitory activity
on the conversion of growth plate cartilage into bone, formation of
a larger cartilaginous template without bone loss in the PC-treated
guinea pigs compared with untreated control guinea pigs would be
expected.
The results presented in the present study provide
further support for the hypothesis that PC or its analogue are
potentially structural disease-modifying drugs for OA therapy.
However, the use of PC as an orally administered OA disease
modifying drug may be limited, as the small molecule PC contains a
P-O-C bond and five negative charges. The P-O-C bond is vulnerable
to degradation by alkaline phosphatases in vivo and the five
negative charges may make PC less permeable to the intestinal
membrane resulting in lower bioavailability in vivo
(48). In order to overcome these
limitations, two strategies are being investigated. The first
strategy is to develop PC analogues that are resistant to
degradation by alkaline phosphatases and/or have fewer negative
changes, which will be tested as orally administered drugs on
Hartley guinea pigs. The second strategy is to develop a hydrogel
that controls the release or extends the release of the small
molecule drug PC. One-time knee injections of the hydrogel for OA
therapy will then be tested in Hartley guinea pigs prior to
consideration for human clinical trials.
In conclusion, articular chondrocytes in the
OA-prone Hartley guinea pigs exhibit a hypertrophic phenotype and
may recapitulate a developmental molecular program similar to the
endochondral pathway of ossification. This molecular program may
serve a role in articular cartilage thinning and destruction. The
present results indicated that PC or its analogue exerted OA
disease-modifying activity, in part, by inhibiting this reactivated
molecular program in OA knee joints.
Glossary
Abbreviations
Abbreviations:
|
COL-X
|
type X collagen
|
|
H&E
|
hematoxylin and eosin
|
|
MMP13
|
matrix metalloproteinase-13
|
|
OA
|
osteoarthritis
|
|
PC
|
phosphocitrate
|
|
PC-E
|
phosphocitrate-β ethyl ester
|
References
|
1
|
Brown TD, Johnston RC, Saltzman CL, Marsh
JL and Buckwalter JA: Posttraumatic osteoarthritis: A first
estimate of incidence, prevalence, and burden of disease. J Orthop
Trauma. 20:739–744. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Olney RC: Mechanisms of impaired growth:
Effect of steroids on bone and cartilage. Horm Res. 72 Suppl
1:S30–S35. 2009. View Article : Google Scholar
|
|
3
|
Goldring MB, Otero M, Plumb DA, Dragomir
C, Favero M, El Hachem K, Hashimoto K, Roach HI, Olivotto E, Borzì
RM and Marcu KB: Roles of inflammatory and anabolic cytokines in
cartilage metabolism: Signals and multiple effectors converge upon
MMP-13 regulation in osteoarthritis. Eur Cell Mater. 21:202–220.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Martel-Pelletier J, Welsch DJ and
Pelletier JP: Metalloproteases and inhibitors in arthritic
diseases. Best Pract Res Clin Rheumatol. 15:805–829. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Burrage PS and Brinckerhoff CE: Molecular
targets in osteoarthritis: Metalloproteinases and their inhibitors.
Curr Drug Targets. 8:293–303. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Lotz MK, Otsuki S, Grogan SP, Sah R,
Terkeltaub R and D'Lima D: Cartilage cell clusters. Arthritis
Rheum. 62:2206–2218. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
van der Kraan PM and van den Berg WB:
Chondrocyte hypertrophy and osteoarthritis: Role in initiation and
progression of cartilage degeneration? Osteoarthritis Cartilage.
20:223–232. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Tchetina EV, Squires G and Poole AR:
Increased type II collagen degradation and very early focal
cartilage degeneration is associated with upregulation of
chondrocyte differentiation related genes in early human articular
cartilage lesions. J Rheumatol. 32:876–886. 2005.PubMed/NCBI
|
|
9
|
Bertrand J, Cromme C, Umlauf D, Frank S
and Pap T: Molecular mechanisms of cartilage remodelling in
osteoarthritis. Int J Biochem Cell Biol. 42:1594–1601. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Clark JM: The structure of vascular
channels in the subchondral plate. J Anat. 171:105–115.
1990.PubMed/NCBI
|
|
11
|
Kawaguchi H: Endochondral ossification
signals in cartilage degradation during osteoarthritis progression
in experimental mouse models. Mol Cells. 25:1–6. 2008.PubMed/NCBI
|
|
12
|
D'Angelo M, Yan Z, Nooreyazdan M, Pacifici
M, Sarment DS, Billings PC and Leboy PS: MMP-13 is induced during
chondrocyte hypertrophy. J Cell Biochem. 77:678–693. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Inada M, Wang Y, Byrne MH, Rahman MU,
Miyaura C, López-Otín C and Krane SM: Critical roles for
collagenase-3 (Mmp13) in development of growth plate cartilage and
in endochondral ossification. Proc Natl Acad Sci USA. 101:pp.
17192–17197. 2004; View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Wei F, Zhou J, Wei X, Zhang J, Fleming BC,
Terek R, Pei M, Chen Q, Liu T and Wei L: Activation of Indian
hedgehog promotes chondrocyte hypertrophy and upregulation of
MMP-13 in human osteoarthritic cartilage. Osteoarthritis Cartilage.
20:755–763. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Lane LB and Bullough PG: Age-related
changes in the thickness of the calcified zone and the number of
tidemarks in adult human articular cartilage. J Bone Joint Surg Br.
62:372–375. 1980.PubMed/NCBI
|
|
16
|
Oettmeier R, Abendroth K and Oettmeier S:
Analyses of the tidemark on human femoral heads. II. Tidemark
changes in osteoarthrosis-a histological and histomorphometric
study in non-decalcified preparations. Acta Morphol Hung.
37:169–180. 1989.PubMed/NCBI
|
|
17
|
Staines KA, Pollard AS, McGonnell IM,
Farquharson C and Pitsillides AA: Cartilage to bone transitions in
health and disease. J Endocrinol. 219:R1–R12. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Williams G and Sallis JD: Structural
factors influencing the ability of compounds to inhibit
hydroxyapatite formation. Calcif Tissue Int. 34:169–177. 1982.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Shankar R, Crowden S and Sallis JD:
Phosphocitrate and its analogue N-sulpho-2-amino tricarballylate
inhibit aortic calcification. Atherosclerosis. 52:191–198. 1984.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Sallis JD, Shankar R, Rees B and Thomson
R: Protection of crystal-induced polymorphonuclear leukocyte
membranolysis by phosphocitrate. Biochem Med Metab Biol. 41:56–63.
1989. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Nair D, Misra RP, Sallis JD and Cheung HS:
Phosphocitrate inhibits a basic calcium phosphate and calcium
pyrophosphate dihydrate crystal-induced mitogen-activated protein
kinase cascade signal transduction pathway. J Biol Chem.
272:18920–18925. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Cheung HS, Sallis JD and Struve JA:
Specific inhibition of basic calcium phosphate and calcium
pyrophosphate crystal-induction of metalloproteinase synthesis by
phosphocitrate. Biochim Biophys Acta. 1315:105–111. 1996.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Sun Y, Reuben P, Wenger L, Sallis JD,
Demadis KD and Cheung HS: Inhibition of calcium phosphate-DNA
coprecipitates induced cell death by phosphocitrates. Front Biosci.
10:803–808. 2005. View
Article : Google Scholar : PubMed/NCBI
|
|
24
|
Cheung HS: Phosphocitrate as a potential
therapeutic strategy for crystal deposition disease. Curr Rheumatol
Rep. 3:24–28. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Cheung HS, Sallis JD, Demadis KD and
Wierzbicki A: Phosphocitrate blocks calcification-induced articular
joint degeneration in a guinea pig model. Arthritis Rheum.
54:2452–2461. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Sun Y, Mauerhan DR, Franklin AM, Norton J,
Hanley EN Jr and Gruber HE: Phosphocitrate is potentially a
disease-modifying drug for noncrystal-associated osteoarthritis.
Biomed Res Int. 2013:3262672013. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Sun Y, Roberts A, Mauerhan DR, Sun AR,
Norton HJ and Hanley EN Jr: Biological activities of
phosphocitrate: A potential meniscal protective agent. Biomed Res
Int. 2013:7265812013. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Sun Y, Mauerhan DR, Steuerwald NM, Ingram
J, Kneisl JS and Hanley EN Jr: Expression of
phosphocitrate-targeted genes in osteoarthritis menisci. Biomed Res
Int. 2014:2104692014. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Turhanen PA, Demadis KD, Peräniemi S and
Vepsäläinen JJ: A novel strategy for the preparation of naturally
occuring phosphocitrate and its partially esterified derivatives. J
Org Chem. 72:1468–1471. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Bendele AM, White SL and Hulman JF:
Osteoarthrosis in guinea pigs: Histopathologic and scanning
electron microscopic features. Lab Anim Sci. 39:115–121.
1989.PubMed/NCBI
|
|
31
|
Kuyinu EL, Narayanan G, Nair LS and
Laurencin CT: Animal models of osteoarthritis: Classification,
update and measurement of outcomes. J Orthop Surg Res. 11:192016.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Poole R, Blake S, Buschmann M, Goldring S,
Laverty S, Lockwood S, Matyas J, McDougall J, Pritzker K, Rudolphi
K, et al: Recommendations for the use of preclinical models in the
study and treatment of osteoarthritis. Osteoarthritis Cartilage. 18
Suppl 3:S10–S16. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Jimenez PA, Glasson SS, Trubetskoy OV and
Haimes HB: Spontaneous osteoarthritis in Dunkin Hartley guinea
pigs: Histologic, radiologic and biochemical changes. Lab Anim Sci.
47:598–601. 1997.PubMed/NCBI
|
|
34
|
Huebner JL, Seifer DR and Kraus VB: A
longitudinal analysis of serum cytokines in the Hartley guinea pig
model of osteoarthritis. Osteoarthritis Cartilage. 15:354–356.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Thomsen JS, Straarup TS, Danielsen CC,
Oxlund H and Brüel A: No effect of risedronate on articular
cartilage damage in the Dunkin Hartley guinea pig model of
osteoarthritis. Scand J Rheumatol. 42:408–416. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Thomsen JS, Straarup TS, Danielsen CC,
Oxlund H and Brüel A: Relationship between articular cartilage
damage and subchondral bone properties and meniscal ossification in
the Dunkin Hartley guinea pig model of osteoarthritis. Scand J
Rheumatol. 40:391–399. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Horcajada MN, Sanchez C, Membrez Scalfo F,
Drion P, Comblain F, Taralla S, Donneau AF, Offord EA and Henrotin
Y: Oleuropein or rutin consumption decreases the spontaneous
development of osteoarthritis in the Hartley guinea pig.
Osteoarthritis Cartilage. 23:94–102. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Sun Y, Haines N, Roberts A, Ruffolo M,
Mauerhan DR, Mihalko KL, Ingram J, Cox M and Hanley EN Jr:
Disease-modifying effects of phosphocitrate and
phosphocitrate-beta-ethyl ester on partial meniscectomy-induced
osteoarthritis. BMC Musculoskelet Disord. 16:2702015. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Huebner JL, Hanes MA, Beekman B, TeKoppele
JM and Kraus VB: A comparative analysis of bone and cartilage
metabolism in two strains of guinea-pig with varying degrees of
naturally occurring osteoarthritis. Osteoarthritis Cartilage.
10:758–767. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Sun Y, Mauerhan DR, Kneisl JS, Norton HJ,
Zinchenko N and Ingram JA: Histologic evidence for collagen and
proteoglycan changes in menisci of osteoarthritic patients.
Proceedings of the ACR/ARHP 75th Annual Scientific Meeting.
Chicago, IL. 2011;
|
|
41
|
Kirsch T, Swoboda B and Nah H: Activation
of annexin II and V expression, terminal differentiation,
mineralization and apoptosis in human osteoarthritic cartilage.
Osteoarthritis Cartilage. 8:294–302. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Kim HA, Lee YJ, Seong SC, Choe KW and Song
YW: Apoptotic chondrocyte death in human osteoarthritis. J
Rheumatol. 27:455–462. 2000.PubMed/NCBI
|
|
43
|
Kouri JB, Aguilera JM, Reyes J, Lozoya KA
and González S: Apoptotic chondrocytes from osteoarthrotic human
articular cartilage and abnormal calcification of subchondral bone.
J Rheumatol. 27:1005–1019. 2000.PubMed/NCBI
|
|
44
|
Sophia Fox AJ, Bedi A and Rodeo SA: The
basic science of articular cartilage: Structure, composition and
function. Sports Health. 1:461–468. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Marini S, Fasciglione GF, Monteleone G,
Maiotti M, Tarantino U and Coletta M: A correlation between knee
cartilage degradation observed by arthroscopy and synovial
proteinases activities. Clin Biochem. 36:295–304. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Ozler K, Aktas E, Atay C, Yilmaz B, Arikan
M and Güngör Ş: Serum and knee synovial fluid matrix
metalloproteinase-13 and tumor necrosis factor-alpha levels in
patients with late-stage osteoarthritis. Acta Orthop Traumatol
Turc. 50:356–361. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Molloy ES, Morgan MP, McDonnell B, O'byrne
J and McCarthy GM: BCP crystals increase prostacyclin production
and upregulate the prostacyclin receptor in OA synovial
fibroblasts: Potential effects on mPGES1 and MMP-13. Osteoarthritis
Cartilage. 15:414–420. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Cooper CM and Sallis JD: Studies on the
gastrointestinal absorption of phosphocitrate, a powerful
controller of hydroxyapatite formation. Int J Pharm. 98:165–172.
1993. View Article : Google Scholar
|