Introduction
Colon cancer is a clinically common malignant tumor
in digestive tract, and it has a very high incidence all over the
world (1,2). It is reported that the incidence of
colon cancer is the third highest among all malignant tumors
(3). At present, surgery combined
with radiochemotherapy and immunotherapy are the main ways of
clinical treatments for colon cancer (4–6). In patients with
early-stage colon cancer, the degree of malignancy is not high, and
lymph node metastasis does not occur (7). These patients usually have good
postoperative recovery, and a 5-year survival rate of over 60%
(8). By contrast, patients with
advanced colon cancer usually have poor clinical treatment effects
and a 5-year survival rate of 20%, because of widespread metastasis
and drug resistance (9). The
invasion and metastasis of cancer is the main cause of death in
colon cancer patients, but its molecular mechanism is still
unclear. Forkhead box (Fox) family genes are abundant in fungi and
animal cells, and their main function is to regulate gene
expression by binding with DNA promoter region as a transcription
factor (10). Fox family is first
discovered to regulate embryonic development, and more and more
studies show that Fox family proteins play a key regulatory role in
immune system, cell cycle, energy metabolism, and cell aging
(11,12). FOXA2 gene, a member of Fox family, is
localized at human chromosome 20p11, with a length of 45 kb
(13,14). It contains 3 exons and 2 introns, and
its protein product has 457 amino acids (13,14). The
amino acid sequence of its binding site at DNA is 5′-RYMAYAY-3′
(13,14). It is discovered that FOXA2 can bind
to the promoter region of genes such as CREB and HNF6, thereby
activating their transcription (15,16).
Under normal conditions, FOXA2 is expressed differently in liver,
lungs, pancreas and other tissues, and its activity is regulated by
phosphorylation and acetylation (17). It is also reported that the
expression of FOXA2 is abnormal in many tumor tissues, and abnormal
FOXA2 is involved in the processes of proliferation, invasion and
metastasis, and epithelial mesenchymal transition (EMT) of various
tumors, such as liver cancer, prostate cancer, stomach cancer, and
bladder cancer (18,19). At present, the expression and
function of FOXA2 gene in colon cancer is still unclear.
EMT refers to a transformation process in which
epithelial cells, under the influence by certain factors, lose the
polarity of epithelial cells and acquire the characteristics of
mesenchymal cells (20). This
process is accompanied by changes in cell morphology, function, and
gene expression profiles (21). EMT
is a key step for tumor cells to break away from their original
sites and gain the ability of invasion and metastasis, and
alleviation of EMT suppresses invasion and metastasis of tumor
cells (22). Therefore, it is
necessary to study the molecular mechanism of tumor cell EMT.
Studies show that a variety of transcription factors play important
roles in tumor EMT, including ZEB1, E47 and Twist (23,24).
However, whether FOXA2 is involved in the regulation of EMT and
thereby affecting tumor metastasis has not been reported before. In
the present study, we investigate the expression and mechanism of
action of FOXA2 in colon cancer at tissue and cellular levels.
Materials and methods
Patients
A total of 66 patients with colon cancer who
received treatments at our hospital between January 2014 and
December 2016 were included in the present study. Among the
patients, 49 were males, and 17 were females, with an age range of
35–64 years and an average age of 48.5 years. The inclusion
criterion was that the patients were initially diagnosed of colon
cancer. The exclusion criteria were: i) the patient had other
tumors; ii) the patient received radiotherapy or chemotherapy; iii)
the patient had a long history of drug intake; iv) the patient had
chronic diseases. None of the patients received radiochemotherapy
before surgeries. The clinical and pathological data were
collected, including tumor size, clinical stage, histological
differentiation, lymph node metastasis and distant metastasis.
Colon cancer tissues were collected from all patients. In addition,
tumor-adjacent tissues were collected as control. All procedures
were approved by the Ethics Committee of Qingdao University.
Written informed consents were obtained from all patients or their
families.
Cells
Normal colonic epithelial NCM460 cells, and colon
cancer HCT116 and HT29 cells were cultured in DMEM medium
supplemented with 10% fetal bovine serum, 100 IU/ml penicillin and
100 IU/ml streptomycin under 37°C, 5% CO2, and 70%
humidity. The cells were passaged every three days, and log-phase
cells were collected for experiments.
One day before transfection, log-phase HCT116 and
HT29 cells (2×105) were seeded onto 24-well plates, and
cultured in serum-free DMEM medium until reaching 70% confluency.
In the first vial, 1.5 µl FOXA2 siRNA [20 pmol/µl; FOXA2
interference sequence (siR-FOXA2) group] or 0.5 µg FOXA2 plasmids
(Hanbio Biotechnology Co., Ltd., Shanghai, China) was mixed with 50
µl Opti Memi medium (Thermo Fisher Scientific, Inc., Waltham, MA,
USA). In the second vial, 1 µl Lipofectamine 2000 (Thermo Fisher
Scientific, Inc.) was mixed with 50 µl Opti-MEMI medium. After
standing still for 5 min, the two vials were combined for another
waiting at room temperature for 20 min. Then, the mixtures were
added onto cells in respective groups. Six hours later, the medium
was replaced with DMEM medium containing 10% fetal bovine serum.
After cultivation at 37°C and 5% CO2 for 48 h, the cells
were collected for further assays.
Immunohistochemical data analysis
After logging on The Human Protein Atlas database
(http://www.proteinatlas.org/), the name
of ‘FOXA2’ was searched to query the expression of FOXA2 in colon
cancer and normal colon tissues. In human tissues, FOXA2 scores
were given by the website. Search on the website showed that FOXA2
protein expression is up-regulated. Therefore, we tested protein
and mRNA expression at cellular and tissue levels as shown below.
In addition, positive rate and distribution were analyzed.
Reverse transcription-quantitative polymerase chain
reaction (RT-qPCR). Colon cancer tissues and control tissues (100
mg) were ground into powder in liquid nitrogen and mixed with 1 ml
Trizol (Thermo Fisher Scientific, Inc.) for lysis. Then, total RNA
was extracted using phenol chloroform method. The purity of RNA was
determined by A260/A280 using ultraviolet spectrophotometry
(Nanodrop ND2000; Thermo Fisher Scientific, Inc.). Then, cDNA was
obtained by reverse transcription from 1 µg RNA and stored at
−20°C. Reverse transcription of was carried out using miScript II
RT kit (Qiagen, Hilden, Germany).
For RT-qPCR, miScript SYBR® Green PCR Kit
(Qiagen) was used. The RT-qPCR reaction system was composed of 10
µl RT-qPCR-Mix, 0.5 µl upstream primer (5′-CCCCTGAGTTGGCGGTGGT-3′),
0.5 µl downstream primer (5′-TTGCTCACGGAAGAGTAG-3′), 2 µl cDNA and
7 µl ddH2O. The reaction protocol was: Initial
denaturation at 95°C for 10 min; 40 cycles of denaturation at 95°C
for 1 min and annealing at 60°C for 30 sec.
Western blotting
Cells (1×106) in each group were
trypsinized and collected. Then, precooled radio
immunoprecipitation assay (RIPA) lysis buffer (1,000 µl; 50 mM
Tris-base, 1 mM EDTA, 150 mM NaCl, 0.1% sodium dodecyl sulfate, 1%
Triton X-100, 1% sodium deoxycholate; Beyotime Institute of
Biotechnology, Shanghai, China) was added to the samples. After
lysis for 30 min on ice, the mixture was centrifuged at 12,000 × g
ta 4°C for 10 min. The supernatant was used to determine protein
concentration by bicinchoninic acid (BCA) protein concentration
determination kit (RTP7102; Real-Times Biotechnology Co., Ltd.,
Beijing, China). Protein samples (6 µg) were then mixed with 5X
sodium dodecyl sulfate loading buffer before denaturation in
boiling water bath for 10 min. Afterwards, the samples (10 µl) were
subjected to 10% sodium dodecyl sulfate-polyacrylamide gel
electrophoresis (80 V). The resolved proteins were transferred to
polyvinylidene difluoride membranes on ice (100 V, 1 h) and blocked
with 50 g/l skimmed milk at room temperature for 1 h. Then, the
membranes were incubated with rabbit anti-human FOXA2 polyclonal
primary antibody (1:1,000; Abcam, Cambridge, UK), rabbit anti-human
E-Cadherin polyclonal primary antibody (1:1,000; Abcam), rabbit
anti-human Vimentin polyclonal primary antibody (1:1,000; Abcam),
and rabbit anti-human GAPDH primary antibody (1:4,000; Abcam) at
4°C overnight. After extensive washing with phosphate-buffered
saline with Tween-20 for 5 times of 5 min, the membranes were
incubated with goat anti-rabbit (1:2,000 for FOXA2, E-Cadherin and
Vimentin) or goat anti-mouse (1:4,000 for GAPDH)-labelled
horseradish peroxidase-conjugated secondary antibodies (Abcam) for
1 h at room temperature before washing with phosphate-buffered
saline with Tween-20 for 5 times of 5 min. Then, the membrane was
developed with enhanced chemiluminescence detection kit
(Sigma-Aldrich; Merck KGaA, Darmstadt, Germany) for imaging. Image
lab v3.0 software (Bio-Rad Laboratories, Inc., Hercules, CA, USA)
was used to acquire and analyze imaging signals. The relative
contents of target proteins were expressed against GAPDH.
Cell Counting Kit-8 (CCK-8) assay
Cells were seeded at a density of 2,000/well in
96-well plates. At 0, 24, 48 and 72 h, 20 µl CCK-8 (5 g/l) solution
(Beyotime Institute of Biotechnology) was added onto the cells. On
the last day, 150 µl CCK-8 reaction solution was added and the
cells were incubated at 37°C for 2 h. The absorbance of each well
was measured at 490 nm for plotting cell proliferation curves. Each
group was tested in 3 replicate wells and the values were
averaged.
Transwell assay
Matrigel was thawed at 4°C overnight and diluted
with serum-free DMEM medium (dilution 1:3). The mixture (50 µl) was
evenly smeared into the upper chamber (HyClone; GE Healthcare Life
Sciences, Logan, UT, USA) and incubated at 37°C for 1 h. After
solidification, 1×105 cells from each group were seeded
into the upper chamber containing 200 µl serum-free DMEM medium. In
addition, 500 µl DMEM medium supplemented with 10% fetal bovine
serum was added into the lower chamber. After incubation at 37°C
and 5% CO2 for 24 h, the chamber was removed and the
cells in the upper chamber were wiped off. After being fixed with
4% formaldehyde for 10 min, the membrane was stained using Giemsa
method for microscopic observation of 5 random fields
(magnification, ×200). The number of transwell cells was calculated
for the evaluation of cell invasion and migration ability. All
procedures were carried out on ice with pipetting tips being
precooled at 4°C.
Flow cytometry
At 24 h after transfection, cells (1×106)
in each group were washed with pre-cooled phosphate-buffered saline
twice and subjected to cell cycle detection using Cycletest™ Plus
DNA Reagent kit (BD Biosciences, Franklin Lakes, NJ, USA) following
the manufacturer's manual. The data were analyzed using ModFit
software (Verity Software House, Topsham, ME, USA).
Laser scanning confocal
microscopy
At 24 h after transfection with siR-FOXA2, cells
(1×105) in each group were seeded onto petri-dishes
(diameter, 6 cm) and incubated at 37°C and under 5% CO2
for 24 h. After discarding medium, the cells were washed with
phosphate-buffered saline for three times, and fixed with 4%
formaldehyde for 10 min. After washing with phosphate-buffered
saline for 3 times, the cells were stained with 5 µM rhodamine for
5 min. After additional washing with phosphate-buffered saline for
3 times, the cells were visualized under a laser scanning confocal
microscope (SP8; Leica, Wetzlar, Germany).
In vivo assay in animal model
Sixteen BALB/C nude mice were divided evenly into
two groups, negative control (NC) and siR-FOXA2 group. NC and
siR-FOXA2 sequences were transfected into HCT116 cells using
Lipofectamine 2000 (Thermo Fisher Scientific, Inc.) following the
manufacturer's manual. At 24 h after transfection, the cells
(2×106) in each group were trypsinized and resuspended
with sterile phosphate-buffered saline (0.2 ml). Then, the cell
suspension was inoculated in nude mouse armpit to construct
xenograft tumor model nude mice. Vital signs of nude mice were
observed, and the mice were sacrificed in week 5 to extract tumor
tissues. After fixation with 4% formaldehyde, the tumor tissues
were dehydrated, paraffin-embedded, and sliced (4 nm thick) for
immunohistochemistry test. E-Cadeherin and Vimentin rabbit
anti-human polyclonal antibodies (Beyotime Institute of
Biotechnology) were diluted at a ratio of 1:50 with water, and used
to incubate the slices at 4°C overnight. After washing with
phosphate-buffered saline twice, goat anti-rabbit secondary
antibody was added, followed by incubation at 37°C for 30 min
before color development. All animal experiments were conducted
according to the ethical guidelines of Qingdao University (Qingdao,
China).
Statistical analysis
Statistical analysis was performed using SPSS 17.0
software (IBM Corp., Armonk, NY, USA). The data were expressed as
mean ± standard deviation and intergroup comparison was carried out
using Student's t-test. Comparison of multiple groups was performed
using ANOVA followed by Tukey's or Dunnett's test. P<0.05 was
considered to indicate a statistically significant difference.
Results
Expression of FOXA2 in colon cancer
tissues is elevated, and closely related with the metastasis and
clinical staging of colon cancer
To examine the localization of FOXA2, we used
immunohistochemistry. By comparing with public immunohistochemical
database (http://www.proteinatlas.org/), we discovered that
FOXA2 expression in colon cancer tissues was significantly higher
than that in normal colon tissues (all data from Protein Atlas
database; Fig. 1A).
Immunohistochemical data showed that FOXA2 was localized in the
nucleus of NCM460 cells, and the nucleus, cytoplasm and membrane of
HCT116 and HT29 cells (Fig. 1B). To
measure the expression of FOXA2, RT-qPCR was performed. The data
showed that FOXA2 expression in colon cancer tissues was
significantly higher than that in normal colon tissues (Fig. 1C). In addition, FOXA2 expression in
colon cancer tissues from patients with lymphatic metastasis was
significantly higher than that from patients without lymphatic
metastasis (Fig. 1D). Clinical
staging showed that FOXA2 level in colon cancer tissues at IV stage
was significantly higher than those at I, II and III stages
(Fig. 1E). These results suggest
that the expression of FOXA2 in colon cancer tissues is elevated,
and closely related with the metastasis and clinical staging of
colon cancer.
Inhibition of FOXA2 reduces the
proliferation of colon cancer cells in vitro
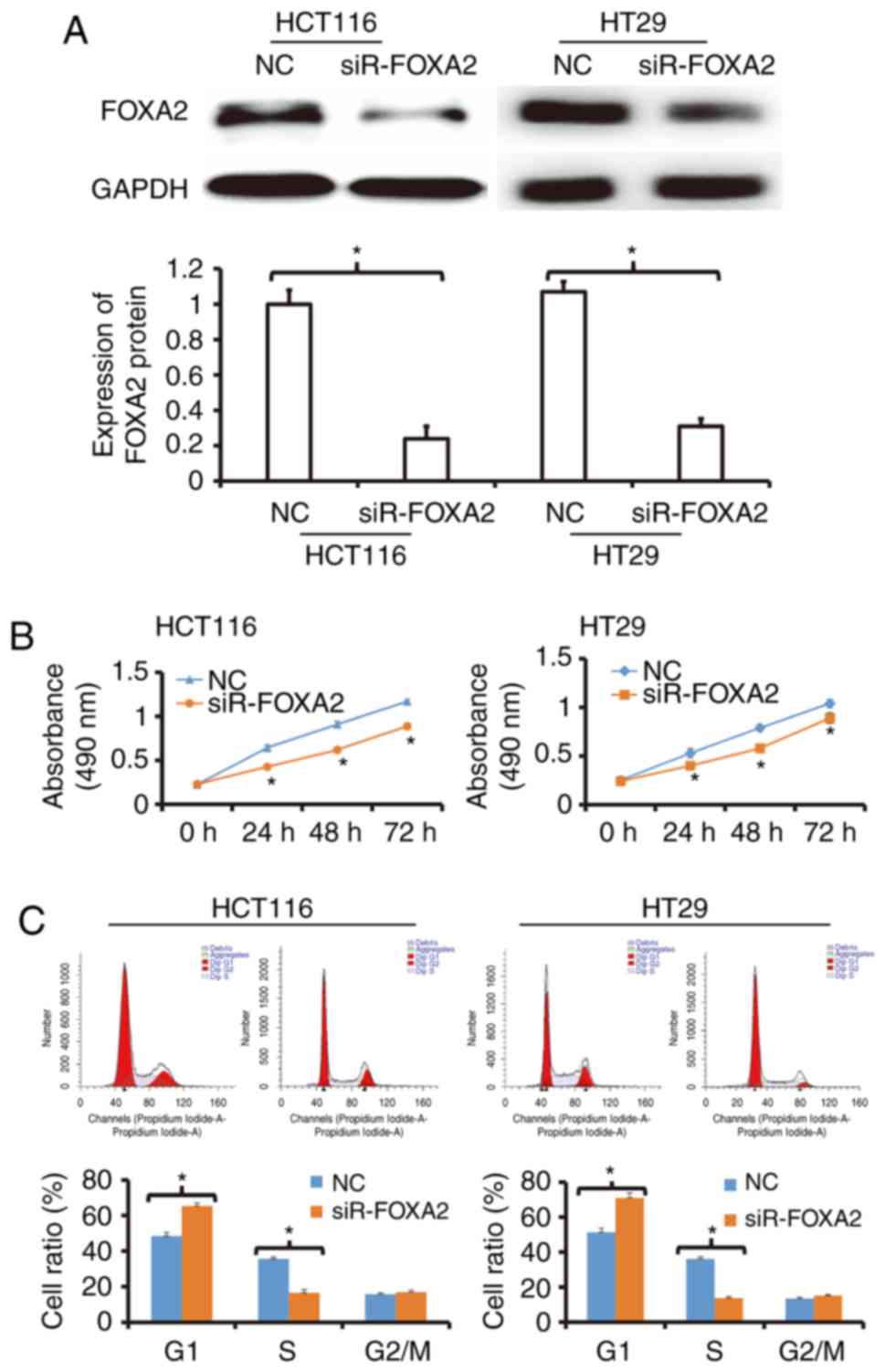
To determine the expression of FOXA2 protein,
western blotting was used. The data showed that transfection with
siR-FOXA2 reduced the protein expression of FOXA2 in HCT116 and
HT29 cells (Fig. 2A). To detect the
proliferation of HCT116 and HT29 cells, CCK-8 assay was carried
out. The data showed that the proliferation of HCT116 and HT29
cells transfected with siR-FOXA2 was significantly reduced compared
with negative control group (P<0.05; Fig. 2B). Flow cytometry showed that G1/S
phase transition in HCT116 and HT29 cells transfected with
siR-FOXA2 was reduced compared with negative control group
(Fig. 2C). These results indicate
that inhibition of FOXA2 reduces the proliferation of colon cancer
cells in vitro, and suppresses the transition from G1 phase
to S phase.
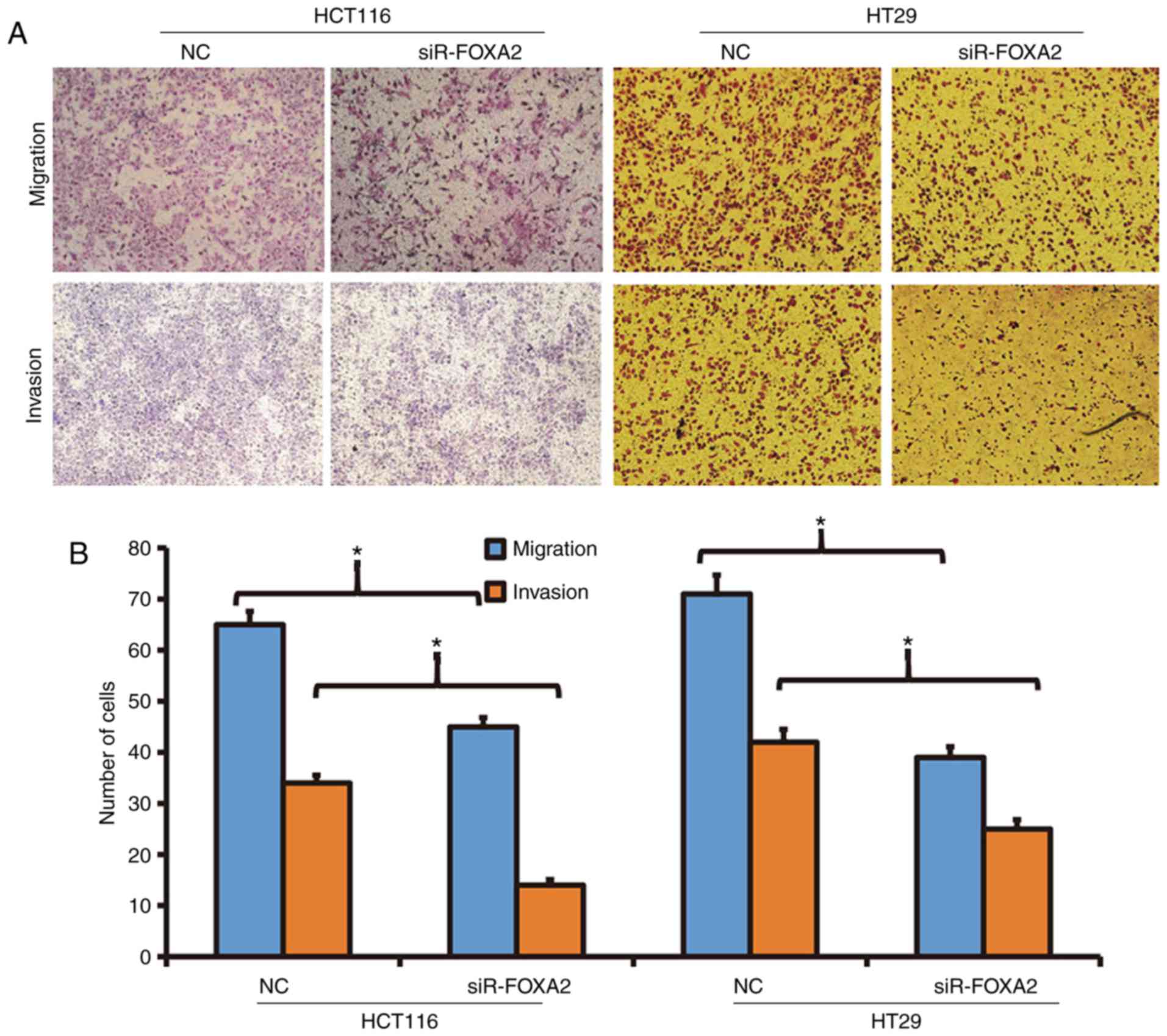
Reduced expression of FOXA2 is able to
decrease the migration and invasion abilities of colon cancer
cells
To determine the migration and invasion abilities of
HCT116 and HT29 cells, Transwell assay was employed. The data
showed that the number of transwell HCT116 cells after transfection
with siR-FOXA2 was significantly reduced than that in negative
control group in both migration and invasion assays (P<0.05;
Fig. 3). Similarly, the number of
transwell HT29 cells after transfection with siR-FOXA2 was
significantly lower than that in negative control group in both
migration and invasion assays (P<0.05; Fig. 3). The results suggest that reduced
expression of FOXA2 is able to decrease the migration and invasion
abilities of colon cancer cells.
FOXA2 may promote EMT, inhibit
apoptosis, and enhance the invasion ability of colon cancer
cells
To detect EMT, apoptosis and cytoskeleton, Western
blotting, flow cytometry and laser scanning confocal microscopy
were carried out, respectively. Western blots showed that the
expression of E-Cadherin in HCT116 and HT29 cells transfected with
siR-FOXA2 was significantly higher than that in NC group, while
expression of Vimentin in HCT116 and HT29 cells transfected with
siR-FOXA2 was lower than that in NC group (Fig. 4A). In the meantime, flow cytometry
showed that the apoptosis of HCT116 and HT29 cells transfected with
siR-FOXA2 was enhanced than those of NC groups (Fig. 4B). Laser scanning confocal microscopy
showed that protrusion of the cell membrane in HCT116 and HT29
cells transfected with siR-FOXA2 was reduced than that in NC group
(Fig. 4C). These results indicate
that FOXA2 may promote EMT, inhibit apoptosis, and enhance the
invasion ability of colon cancer cells.
Decreased expression of FOXA2 inhibits
tumorigenesis of colon cancer cells in nude mice
To test the effect of FOXA2 on the formation of
colon tumor, nude mice were used. Tumor diameter in siR-FOXA2 group
seemed smaller than that in NC group (Fig. 5A). Similarly, weights of colon tumors
from mice inoculated with HCT116 or HT29 cells transfected with
siR-FOXA2 were significantly lower than that in NC group,
respectively (P<0.05; Fig. 5B).
Immunohistochemistry showed that E-Cadherin expression in siR-FOXA2
group seemed higher than that in NC group, while Vimentin
expression in in siR-FOXA2 group seemed lower than that in NC group
(Fig. 5C). The results suggest that
decreased expression of FOXA2 inhibits tumorigenesis of colon
cancer cells in nude mice.
Discussion
Tumor recurrence and metastasis are the key factors
that restrict the postoperative survival of colon cancer patients
(25). Studies show that the
recurrence and metastasis of colon cancer are closely related to
gene expression, methylation, mutation, tumor stem cells, drug
resistance and immunosuppression (7,26).
Transcription factors play key regulatory roles in the process of
gene expression regulation, and they are important drug targets
(27). In the present study, we
discover that the expression of FOXA2 is up-regulated in colon
cancer tissues, and correlated with lymphatic metastasis and
clinical staging of colon cancer. In vitro and in
vivo experiments show that FOXA2 can promote the proliferation,
migration and invasion of colon cancer cells, and EMT may be the
key factor for FOXA2 to promote metastasis of colon cancer. These
results suggest that FOXA2 plays an oncogene role by promoting EMT
and colon cancer recurrence and metastasis.
Transcription factors can bind to the binding sites
of the promoter region of genes, and initiate gene transcription,
having important regulatory functions (28,29). In
cancer, transcription factors are often able to initiate
transcription of multiple downstream oncogenes or tumor-suppressor
genes, thereby inducing a cascade of responses that regulate cell
proliferation, invasion, metastasis, differentiation, drug
resistance, and apoptosis (30,31).
Studies show that multiple transcription factors are closely
related to the recurrence and metastasis of colon cancer. For
example, Ma et al discover that KLF4 inhibits the
proliferation of colon cancer by activating NDRG2 expression
(32). Stein et al discover
that MACC1 indirectly activates the HGF/c-Met signaling pathway by
activating transcription of c-Met gene, and promotes the recurrence
and metastasis of colon cancer (33). In the present study, we find that
transcription factor FOXA2 is up-regulated in colon cancer tissues.
Moreover, our immunohistochemical data show that FOXA2 expression
is distributed in the nucleus, cytoplasm and membrane in colon
cancer, which is abnormal. Our RT-qPCR data demonstrate that
expression of FOXA2 is up-regulated in colon cancer tissues, and
positively correlated with lymphatic metastasis and clinical
staging. These results indicate that FOXA2 is associated with the
development and progression of colon cancer and may play the role
of an oncogene.
Some studies show that the function of FOXA2 in
tumors is two-sided. For example, Li et al report that
miR-187 can target the expression of FOXA2 and promote the
proliferation and metastasis of gastric cancer, suggesting that
FOXA2 may be a tumor-suppressor gene in gastric cancer (34). Down-regulation of FOXA2 may promote
EMT in pancreatic cancer, suggesting that FOXA2 may inhibit tumor
EMT (35). Tu et al discover
that miR-1291 inhibits proliferation and metastasis of pancreatic
cancer by targeting FOXA2 (36). Our
results in the present study show that FOXA2 expression is elevated
in HCT116 and HT29 cells. CCK-8 assay shows that interference of
FOXA2 expression reduces the proliferation of HCT116 and HT29
cells. In addition, HCT116 and HT29 cells in FOXA2 interference
group have shown G1/S arrest, suggesting that down-regulation of
FOXA2 inhibits the proliferation of colon cancer cells. Transwell
assay shows that the number of cells that cross the membrane in
FOXA2 interference group is lower than that in control group,
suggesting that FOXA2 is able to promote the migration and invasion
of colon cancer cells. Furthermore, our Western blotting data show
that expression of E-Cadherin and Vimentin in FOXA2 interference
group is up-regulated and down-regulated, respectively than that in
control group, suggesting that EMT of colon cancer is inhibited by
down-regulation of FOXA2. Of note, the apoptosis of cells in FOXA2
interference group is enhanced than control group, and laser
scanning confocal microscopy shows that the cytoskeleton in the
cell membrane of the FOXA2 interference group is more abundant,
suggesting that these cells have stronger mobility. Tumor formation
in nude mice shows that the growth rate of tumor in FOXA2
interference group is significantly lower than that in NC group. In
addition, E-Cadherin expression is up-regulated and Vimentin
expression is down-regulated in tumor tissues of FOXA2 interference
group, suggesting that EMT is inhibited. These results indicate
that FOXA2 promotes the proliferation, migration and EMT of colon
cancer cells.
In conclusion, the present study demonstrates that
FOXA2 is an oncogene in colon cancer, and the up-regulation of its
expression promotes the proliferation, migration and invasion of
colon cancer. As a transcription factor, up-regulation of FOXA2
expression has clinical values in the diagnosis and prognosis
evaluation of colon cancer. Because FOXA2 initiates the expression
of a large number of downstream genes, it can also be a potential
diagnostic and therapeutic target for colon cancer, being
consistent with a previous report (37). In future studies, we will investigate
the molecular mechanisms of FOXA2 in colon cancer, such as
downstream transcriptional genes of FOXA2 and relevant signaling
pathways.
Acknowledgements
The authors wish to thank their department and
research team for their help and dedication.
Funding
The present work was supported by the National
Natural Science Foundation of China (grant no. 81172031).
Availability of data and materials
The analyzed data sets generated during the study
are available from the corresponding author on reasonable
request.
Authors' contributions
BW and YL designed the study; BW, GL, LD and JZ were
involved in designing and performing experiments; BW, GL and YL
analyzed the data. The final version of the manuscript has been
read and approved by all authors, and each author believes that the
manuscript represents honest work.
Ethics approval and consent to
participate
All procedures performed in the current study were
approved by the Ethics Committee of Qingdao University. Written
informed consent was obtained from all patients or their
families.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Chen Y, Fang L, Li G, Zhang J, Li C, Ma M,
Guan C, Bai F, Lyu J and Meng QH: Synergistic inhibition of colon
cancer growth by the combination of methylglyoxal and silencing of
glyoxalase I mediated by the STAT1 pathway. Oncotarget.
8:54838–54857. 2017.PubMed/NCBI
|
|
2
|
Chino XMS, Martinez CJ, Garzón VRV,
González IÁ, Treviño SV, Bujaidar EM, Ortiz GD and Hoyos RB: Cooked
chickpea consumption inhibits colon carcinogenesis in mice induced
with azoxymethane and dextran sulfate sodium. J Am Coll Nutr.
36:391–398. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Cusimano A, Balasus D, Azzolina A, Augello
G, Emma MR, Di Sano C, Gramignoli R, Strom SC, McCubrey JA,
Montalto G and Cervello M: Oleocanthal exerts antitumor effects on
human liver and colon cancer cells through ROS generation. Int J
Oncol. 51:533–544. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Cho N, Ransom TT, Sigmund J, Tran T,
Cichewicz RH, Goetz M and Beutler JA: Growth inhibition of colon
cancer and melanoma cells by versiol derivatives from a
paraconiothyrium species. J Nat Prod. 80:2037–2044. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Myint ZW and Goel G: Role of modern
immunotherapy in gastrointestinal malignancies: A review of current
clinical progress. J Hematol Oncol. 10:862017. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Goel G and Sun W: Advances in the
management of gastrointestinal cancers-an upcoming role of immune
checkpoint blockade. J Hematol Oncol. 8:862015. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Hou PC, Li YH, Lin SC, Lin SC, Lee JC, Lin
BW, Liou JP, Chang JY, Kuo CC, Liu YM, et al: Hypoxia-induced
downregulation of DUSP-2 phosphatase drives colon cancer stemness.
Cancer Res. 77:4305–4316. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Pahwa M, Harris MA, MacLeod J, Tjepkema M,
Peters PA and Demers PA: Sedentary work and the risks of colon and
rectal cancer by anatomical sub-site in the Canadian census health
and environment cohort (CanCHEC). Cancer Epidemiol. 49:144–151.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Múnera JO, Sundaram N, Rankin SA, Hill D,
Watson C, Mahe M, Vallance JE, Shroyer NF, Sinagoga KL,
Zarzoso-Lacoste A, et al: Differentiation of human pluripotent stem
cells into colonic organoids via transient activation of BMP
signaling. Cell Stem Cell. 21:51–64.e6. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Ancey PB, Ecsedi S, Lambert MP, Talukdar
FR, Cros MP, Glaise D, Narvaez DM, Chauvet V, Herceg Z, Corlu A and
Hernandez-Vargas H: TET-catalyzed 5-hydroxymethylation precedes
HNF4A promoter choice during differentiation of bipotent liver
progenitors. Stem Cell Reports. 9:264–278. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Von Stetina SE, Liang J, Marnellos G and
Mango SE: Temporal regulation of epithelium formation mediated by
FoxA, MKLP1, MgcRacGAP, and PAR-6. Mol Biol Cell. 28:2042–2065.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
McFadden VC, Shalaby RE, Iram S, Oropeza
CE, Landolfi JA, Lyubimov AV, Maienschein-Cline M, Green SJ,
Kaestner KH and McLachlan A: Hepatic deficiency of the pioneer
transcription factor FoxA restricts hepatitis B virus biosynthesis
by the developmental regulation of viral DNA methylation. PLoS
Pathog. 13:e10062392017. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Li J, Machado Dantas AC, Guo M, Sagendorf
JM, Zhou Z, Jiang L, Chen X, Wu D, Qu L, Chen Z, et al: Structure
of the forkhead domain of FOXA2 bound to a complete DNA consensus
site. Biochemistry. 56:3745–3753. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Bastidas-Ponce A, Roscioni SS, Burtscher
I, Bader E, Sterr M, Bakhti M and Lickert H: Foxa2 and Pdx1
cooperatively regulate postnatal maturation of pancreatic β-cells.
Mol Metab. 6:524–534. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Rausa FM III, Hughes DE and Costa RH:
Stability of the hepatocyte nuclear factor 6 transcription factor
requires acetylation by the CREB-binding protein coactivator. J
Biol Chem. 279:43070–43076. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Zhang L, Rubins NE, Ahima RS, Greenbaum LE
and Kaestner KH: Foxa2 integrates the transcriptional response of
the hepatocyte to fasting. Cell Metab. 2:141–148. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Lehner F, Kulik U, Klempnauer J and Borlak
J: Inhibition of the liver enriched protein FOXA2 recovers HNF6
activity in human colon carcinoma and liver hepatoma cells. PLoS
One. 5:e133442010. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Berg DT, Gerlitz B, Sharma GR, Richardson
MA, Stephens EJ, Grubbs RL, Holmes KC, Fynboe K, Montani D, Cramer
MS, et al: FoxA2 involvement in suppression of protein C, an
outcome predictor in experimental sepsis. Clin Vaccine Immunol.
13:426–432. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Kanaki M, Tiniakou I, Thymiakou E and
Kardassis D: Physical and functional interactions between nuclear
receptor LXRα and the forkhead box transcription factor FOXA2
regulate the response of the human lipoprotein lipase gene to
oxysterols in hepatic cells. Biochim Biophys Acta. 1860:848–860.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Li D, He C, Wang J, Wang Y, Bu J, Kong X
and Sun D: MicroRNA-138 inhibits cell growth, invasion and EMT of
non-small cell lung cancer via SOX4/p53 feedback loop. Oncol Res.
Jun 13–2017.(Epub ahead of print). View Article : Google Scholar
|
|
21
|
Zhou Q, Dai J, Chen T, Dada LA, Zhang X,
Zhang W, DeCamp MM, Winn RA, Sznajder JI and Zhou G: Downregulation
of PKCζ/Pard3/Pard6b is responsible for lung adenocarcinoma cell
EMT and invasion. Cell Signal. 38:49–59. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Wahl GM and Spike BT: Cell state
plasticity, stem cells, EMT, and the generation of intra-tumoral
heterogeneity. NPJ Breast Cancer. 3:142017. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Cao Z, Zheng X, Cao L and Liang N:
MicroRNA-539 inhibits the epithelial-mesenchymal transition of
esophageal cancer cells by twist-related protein 1-mediated
modulation of melanoma associated antigen A4 (MAGEA4). Oncol Res.
Jun 12–2017.(Epub Ahead of Print). View Article : Google Scholar
|
|
24
|
Lazarova D and Bordonaro M: ZEB1 mediates
drug resistance and EMT in p300-deficient CRC. J Cancer.
8:1453–1459. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Aquino RGF, Vasques PHD, Cavalcante DIM,
Oliveira ALS, Oliveira BMK and Pinheiro LGP: Invasive ductal
carcinoma: Relationship between pathological characteristics and
the presence of axillary metastasis in 220 cases. Rev Col Bras Cir.
44:163–170. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Laudato S, Patil N, Abba ML, Leupold JH,
Benner A, Gaiser T, Marx A and Allgayer H: P53-induced miR-30e-5p
inhibits colorectal cancer invasion and metastasis by targeting
ITGA6 and ITGB1. Int J Cancer. 141:1879–1890. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Schayek H, Laitman Y, Katz LH, Pras E,
Ries-Levavi L, Barak F and Friedman E: Colorectal and endometrial
cancer risk and age at diagnosis in BLMAsh mutation carriers. Isr
Med Assoc J. 19:365–367. 2017.PubMed/NCBI
|
|
28
|
Liu Z, Zhao Y, Fang J, Cui R, Xiao Y and
Xu Q: SHP2 negatively regulates HLA-ABC and PD-L1 expression via
STAT1 phosphorylation in prostate cancer cells. Oncotarget.
8:53518–53530. 2017.PubMed/NCBI
|
|
29
|
Nelson AC, Cutty SJ, Gasiunas SN, Deplae
I, Stemple DL and Wardle FC: In vivo regulation of the zebrafish
endoderm progenitor niche by T-Box transcription factors. Cell Rep.
19:2782–2795. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Shen Y, Nar R, Fan AX, Aryan M, Hossain
MA, Gurumurthy A, Wassel PC, Tang M, Lu J, Strouboulis J and
Bungert J: Functional interrelationship between TFII-I and E2F
transcription factors at specific cell cycle gene loci. J Cell
Biochem. 119:712–722. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Seo HS, Ku JM, Choi HS, Woo JK, Lee BH,
Kim DS, Song HJ, Jang BH, Shin YC and Ko SG: Apigenin overcomes
drug resistance by blocking the signal transducer and activator of
transcription 3 signaling in breast cancer cells. Oncol Rep.
38:715–724. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Ma Y, Wu L, Liu X, Xu Y, Shi W, Liang Y,
Yao L, Zheng J and Zhang J: KLF4 inhibits colorectal cancer cell
proliferation dependent on NDRG2 signaling. Oncol Rep. 38:975–984.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Stein U, Walther W, Arlt F, Schwabe H,
Smith J, Fichtner I, Birchmeier W and Schlag PM: MACC1, a newly
identified key regulator of HGF-MET signaling, predicts colon
cancer metastasis. Nat Med. 15:59–67. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Li C, Lu S and Shi Y: MicroRNA-187
promotes growth and metastasis of gastric cancer by inhibiting
FOXA2. Oncol Rep. 37:1747–1755. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Kondratyeva LG, Sveshnikova AA, Grankina
EV, Chernov IP, Kopantseva MR, Kopantzev EP and Sverdlov ED:
Downregulation of expression of mater genes SOX9, FOXA2, and GATA4
in pancreatic cancer cells stimulated with TGFβ1
epithelial-mesenchymal transition. Dokl Biochem Biophys.
469:257–259. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Tu MJ, Pan YZ, Qiu JX, Kim EJ and Yu AM:
MicroRNA-1291 targets the FOXA2-AGR2 pathway to suppress pancreatic
cancer cell proliferation and tumorigenesis. Oncotarget.
7:45547–45561. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Jayachandran A, Dhungel B and Steel JC:
Epithelial-to-mesenchymal plasticity of cancer stem cells:
Therapeutic targets in hepatocellular carcinoma. J Hematol Oncol.
9:742016. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Uhlén M, Fagerberg L, Hallström BM,
Lindskog C, Oksvold P, Mardinoglu A, Sivertsson Å, Kampf C,
Sjöstedt E, Asplund A, et al: Proteomics. Tissue-based map of the
human proteome. Science. 347:12604192015. View Article : Google Scholar : PubMed/NCBI
|