Introduction
Liver ischemia reperfusion (IR) injury is a process
initiated by hypoxia, which causes cellular damage, followed by the
return of blood flow and oxygen delivery, which exacerbates
cellular damage (1,2). Overproduction of reactive oxygen
species (ROS) and superoxide free radicals in IR causes the
destruction of the hepatic cellular membrane and increases their
permeability, leading to hepatocyte death and hepatic injury
(3). Liver IR occurs in a number of
clinical settings, including liver transplantation, partial hepatic
resection, shock and hepatic failure (4–6) At
present, there is no effective prevention strategy for liver IR
injury and there is there for an urgent need for the development of
new treatments.
Baicalin is a flavonoid glycoside extracted from the
root of the traditional Chinese medicinal herb Scutellaria
baicalensis (7). It has been
documented that baicalin possesses a number of pharmacological
actions, including anti-inflammatory, anti-bacterial,
anti-fibrotic, anti-oxidant and anticancer effects (7–11).
Previous studies have reported that baicalin serves a protective
effect in IR injury in various organs, including the kidneys, brain
and heart (12–14). It has been reported that baicalin may
be able to induce autophagy, as evidenced by elevated expression of
microtubule-associated proteins 1A/1B light chain 3B LC3 and
beclin-1 in human bladder T24 cells (15) and human hepatocellular carcinoma
SMMC-7721 cells following baicalin treatment (16).
Autophagy is an important intracellular process for
degrading macromolecules, as well as recycling cytosolic proteins
and organelles, to maintain cellular homeostasis (17). When cellular components are engulfed
into double or multiple-membrane cytoplasmic vesicles,
autophagosomes are formed, which subsequently fuse with lysosomes,
forming autolysosomes and degrading the captured proteins or
organelles with lysosomal enzymes (15,18).
Autophagy is believed to exert a beneficial effect in IR injury;
autophagy ameliorates liver damage when hepatocytes suffer from
anoxia or nutrient starvation during IR insults (19). It has also been reported that
rapamycin, a mechanistic target of rapamycin (mTOR) inhibitor and
potent autophagy inducer, attenuates tubular injury and renal
dysfunction by promoting autophagy in acute kidney injury (20), demonstrating the protective role of
autophagy in IR injury.
The aim of the present study was to investigate the
effect of baicalin on liver IR injury and its relationship with
autophagy. The results of the present study indicate that baicalin
may be an effective clinical treatment for liver IR injury.
Materials and methods
Cell culture and establishment of
hypoxia/reoxygenation model
Normal human liver LO2 cells were purchased from the
Chinese Academy of Science (Shanghai, China) and maintained in
Dulbecco's modified Eagle's medium/F-12 nutrient mixture (DMEM-F12;
Gibco; Thermo Fisher Scientific, Inc., Waltham, MA, USA)
supplemented with 10% fetal bovine serum (FBS; Gibco; Thermo Fisher
Scientific, Inc.) and 1% penicillin-streptomycin in 21%
O2 at 37°C. Hypoxia/reoxygenation (H/R) was performed as
previously described (21). Briefly,
cells were seeded in 6-well plates at a density of 105
cells/well. At 80% confluence, the 6-well plates were incubated
under hypoxic conditions (94% N2, 5% CO2 and
1% O2 at 37°C) for 24 h in a hypoxia chamber. Cells were
then removed from the hypoxia chamber and incubated for 6 h in an
atmosphere containing 5% CO2 at 37°C. A total of 6
groups (n=3 each) were established: Control group, H/R group, 50
µmol/l baicalin group, 100 µmol/l baicalin group, 200 µmol/l
baicalin group and 3-methyladenine (3-MA; 5 mmol/l) group. In the
control group, LO2 cells were cultured under normal culture
conditions only. In the baicalin and 3-MA groups, baicalin (cat.
no. 572667) or 3-MA (cat. no. M9281) (both Sigma-Aldrich; Merck
KGaA, Darmstadt, Germany) was added 1 h prior to H/R.
Cell viability analysis
Cells were seeded in a 96-well plate at a density of
2,000 cells/well. Following 24 h hypoxia and 6 h reoxygenation,
cell survival was assessed using a Cell Counting kit-8 (CCK-8) kit
(Beyotime Institute of Biotechnology, Haimen, China). Briefly, 10
µl of CCK-8 reagent was added to each well and incubated for 2 h at
37°C. Cell density was determined by measuring the absorbance at
450 nm using a microplate reader (MDC Vacuum Products, LLC,
Hayward, CA, USA).
Lactate dehydrogenase (LDH) activity
assay
The supernatant from the 96-well plates were
collected by centrifugation at 500 × g for 5 min at 25°C. LDH
activity in the supernatant (20 µl) was measured using a LDH assay
kit (Nanjing Jiancheng Bioengineering Institute, Nanjing, China) in
accordance with the manufacturer's protocol.
Annexin V and phosphatidylinositol
(PI) binding staining
Cells were washed twice with PBS and resuspended in
500 µl staining buffer (Beyotime Institute of Biotechnology) at a
concentration of 105 cells/ml with 5 µl Annexin
V-fluorescein isothiocyanate (FITC; Beyotime Institute of
Biotechnology) and 5 µl PI. Cells were gently mixed and incubated
for 15 min at 37°C in the dark. A total of 400 µl cell suspension
was transferred into flow tubes and analyzed using a BD FACS Aria
II flow cytometer (BD Biosciences, Franklin Lakes, NJ, USA). The
flow cytometry data were analyzed using FlowJo 6.0 software (FlowJo
LLC, Ashland, OR, USA).
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
Total RNA was extract from LO2 cells using TRIzol
(Invitrogen; Thermo Fisher Scientific, Inc.) according to the
manufacturer's protocol. A total of 3–5 µg of RNA was transcribed
into cDNA using a RevertAid First Strand cDNA Synthesis kit (Thermo
Fisher Scientific, Inc.). Reverse transcription was performed at
42°C for 60 min, followed by 70°C for 5 min. Primers used for
RT-qPCR are listed in Table I. qPCR
was conducted using the SYBR Green premix kit from Takara Bio, Inc.
(Otsu, Japan). Thermocycling conditions were as follows: Initial
denaturation at 95°C for 30 sec followed by 50 cycles of 95°C for 5
sec, 60°C for 30 sec and 72°C for 60 sec. Expression levels were
normalized to β-actin in the same samples using the
2−ΔΔCq method (7).
 | Table I.Gene specific primers used for
reverse transcription-quantitative polymerase chain reaction. |
Table I.
Gene specific primers used for
reverse transcription-quantitative polymerase chain reaction.
| Gene | Primer
sequence |
|---|
| Binding of
immunoglobulin protein | Sense:
AAAGAAGACGGGCAAAGATGT |
|
| Antisense:
TGCTTGATGCTGAGAAGACAG |
|
CCAAT/enhancer-binding protein homologous
protein | Sense:
ACCACTCTTGACCCTGCTTCT |
|
| Antisense:
CTCTGGGAGGTGCTTGTGAC |
| β-actin | Sense:
GTTGTCGACGACGAGCG |
|
| Antisense:
GCACAGAGCCTCGCCTT |
Western blotting
LO2 cells were washed with PBS three times and
ice-cold radioimmunoprecipitation assay buffer (Vazyme, Piscataway,
NJ, USA) was added for cell lysis. Lysates were centrifuged at
12,000 × g at 4°C for 25 min. The supernatant was collected and
total proteins were quantified by bicinchoninic acid assay. Samples
(20 µg) were separated by 10% SDS-PAGE, transferred onto a
polyvinylidene fluoride membranes and blocked with 5% skimmed milk
for 1 h at room temperature. Membranes were subsequently incubated
with primary antibodies against the following: B-cell lymphoma 2
(Bcl-2; cat. no. 2872), Bcl-2-associated X protein (Bax; cat. no.
5023), caspase 3 (cat. no. 9662), cleaved caspase 3 (cat. no.
9661), binding of immunoglobulin protein (BIP; cat. no. 3177),
CCAAT/enhancer-binding protein homologous protein (CHOP; cat. no.
2895), beclin-1 (cat. no. 3495), LC3 (cat. no. 4108), activating
transcription factor 4 (ATF4; cat. no. 11815), X-box binding
protein 1 (XBP-1; cat. no. 83418) and GAPDH (cat. no. 5174; all
1:1,000; all Cell Signaling Technology, Inc., Danvers, MA, USA) at
4°C overnight. Membranes were washed with 1X TBS with 1% Tween-20
and incubated with horseradish-conjugated goat anti-rabbit
secondary antibodies (cat. no. ab6721; 1:10,000; Abcam) for 1 h at
room temperature. Immunoreactive bands were visualized using a
chemiluminescence solution (Thermo Fisher Scientific, Inc.) and
normalized to GAPDH using Image J software version 1.8.0 (National
Institutes of Health, Bethesda, MA, USA).
Statistical analysis
Results are presented as the mean ± standard
deviation. Statistical analysis was performed using one-way
analysis of variance with Bonferroni's correction on SPSS 20
software (IBM Corp., Armonk, NY, USA). P<0.05 was considered to
indicate a statistically significant difference.
Results
Baicalin pretreatment attenuates
H/R-induced damage to LO2 cells
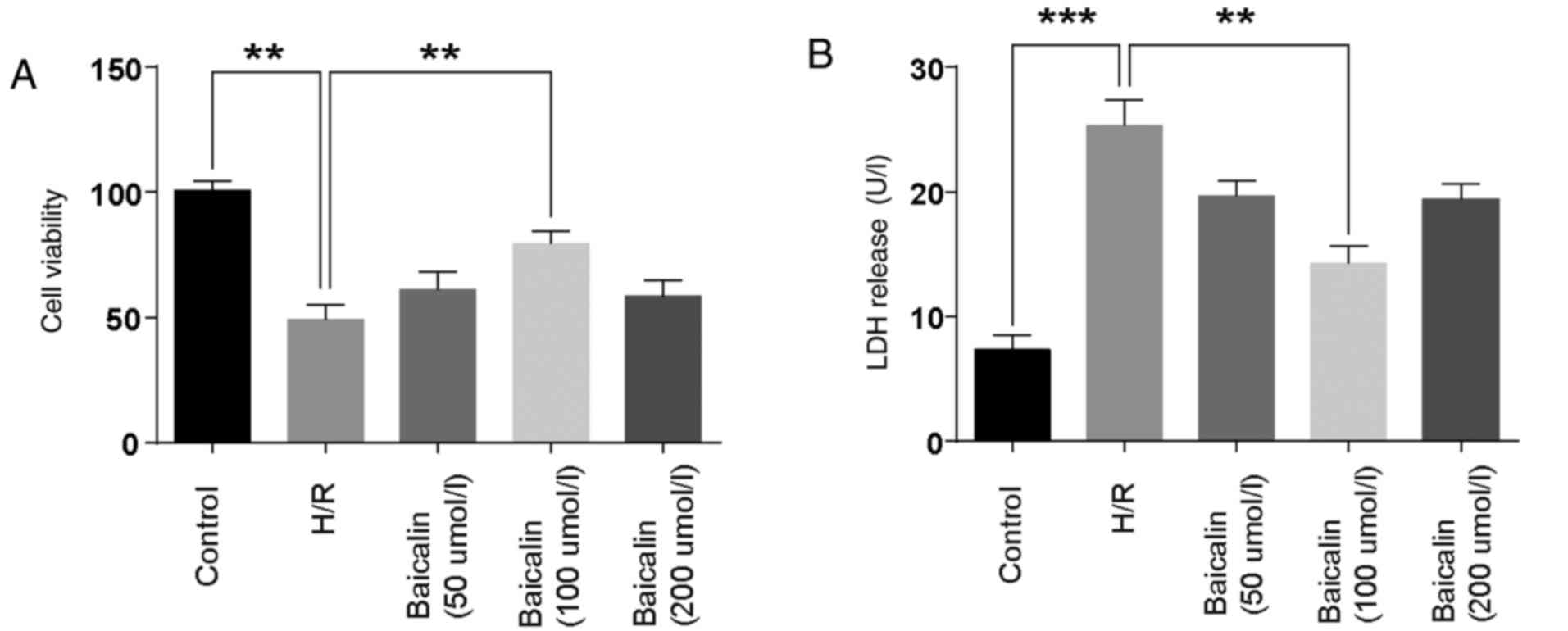
Cell viability and LDH levels were detected to
investigate the effect of baicalin on LO2 cell damage. The results
demonstrated that, compared with the control group, cell viability
was decreased in the H/R group. Treatment with 100 µmol/l baicalin
significantly ameliorated the effects of H/R (Fig. 1A). LDH, a marker of cell injury, was
significantly elevated in the H/R group, while pretreatment with
100 µmol/l baicalin significantly reversed this effect (Fig. 1B). These results suggest that
baicalin pretreatment attenuated H/R-induced damage in LO2
cells.
Baicalin pretreatment inhibits
apoptosis following H/R
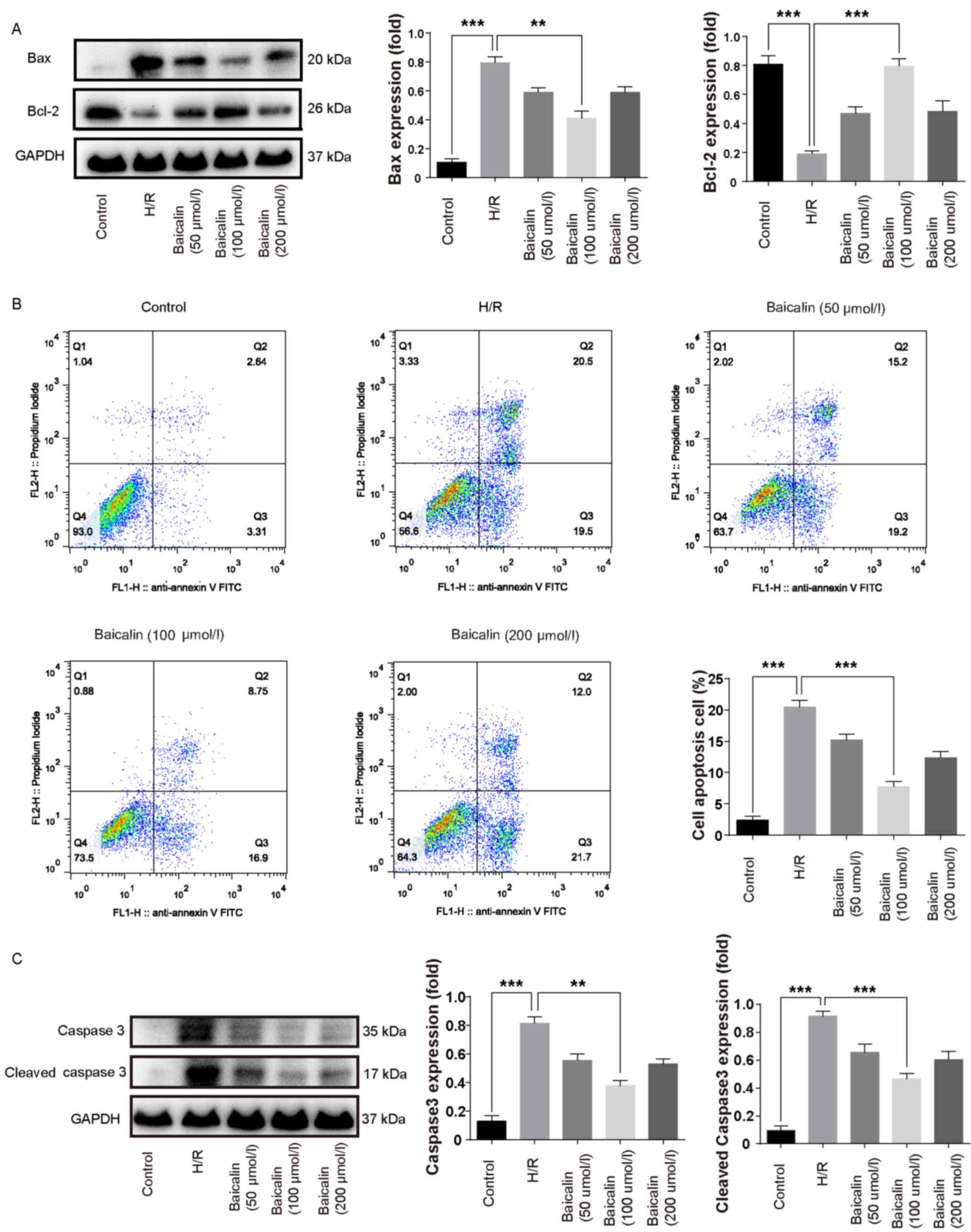
Bcl-2 is an anti-apoptotic protein, whereas Bax is a
pro-apoptotic protein (12). Bcl-2
protein expression was lower in the H/R group compared with the
control, whereas Bax expression was increased (Fig. 2A). Pretreatment with baicalin
significantly increased the expression of Bcl-2 and inhibited the
expression of Bax compared with the H/R group (Fig. 2A). The results of flow cytometry
revealed that the percentage of apoptotic cells in the H/R group
was increased compared with the control group; however,
pretreatment with 100 µmol/l baicalin significantly decreased
apoptosis compared with the H/R group (Fig. 2B). These results suggest that
baicalin pretreatment inhibits apoptosis following H/R.
Baicalin pretreatment decreases
caspase 3 expression following H/R
Caspase 3 is a downstream effector in the caspase
cascade that directly mediates apoptosis upon activation by
multiple upstream signals (22).
Therefore, caspase 3 is regarded as an important pro-apoptotic
indicator. Caspase 3 and cleaved caspase 3 were increased in the
H/R group compared with the control group (Fig. 2C). In contrast, pretreatment with 100
µmol/l significantly decreased the expression of caspase 3 and
cleaved caspase 3 compared with the H/R group (Fig. 2C).
Baicalin pretreatment inhibits
endoplasmic reticulum (ER) stress following H/R
ER stress was assessed by measuring the expression
of BIP and CHOP. Compared with the control group, levels of BIP and
CHOP mRNA were higher in the H/R group, whereas pretreatment with
100 µmol baicalin significantly ameliorated the effects of H/R
(Fig. 3A). Furthermore, the
expression of BIP and CHOP at the protein level was increased in
the H/R group compared with the control and significantly decreased
in the 100 µmol/l baicalin group compared with the H/R group
(Fig. 3B). These results suggest
that baicalin pretreatment inhibits ER stress following H/R.
Baicalin pretreatment induced
autophagy in vitro
To investigate whether baicalin pretreatment is able
to induce autophagy, the expression of LC3 and beclin-1 was
assessed. The results of western blotting revealed that the
expression of LC3-II and beclin-1 were increased in the H/R group
compared with the control (Fig. 4A).
Furthermore, 100 µmol/l baicalin pretreatment significantly
increases the expression of LC3-II and beclin 1 following H/R
(Fig. 4A).
Baicalin pretreatment inhibits ER
stress-induced autophagy in vitro
It has been reported that ER stress induces
autophagy, whereas ATF4 and XBP1 are regarded as important inducers
of autophagy following ER stress (23,24).
Western blotting revealed that ATF4 and XBP1 was upregulated in the
H/R group compared with the control (Fig. 4B). In contrast, baicalin pretreatment
significantly reduced the expression of ATF4 and XBP1 compared with
the H/R group (Fig. 4B).
Autophagy inhibition abrogates the
protective effect of baicalin in vitro
To further confirm the role of autophagy in H/R
injury, the autophagy inhibitor 3-MA was used. The results revealed
that LC3-II and beclin-1 levels were increased in the H/R group
compared with the control and further increased with baicalin
pretreatment (Fig. 5A). However,
co-administration with 3-MA significantly decreased the expression
of LC3-II and beclin-1 compared with baicalin treatment alone
(Fig. 5A).
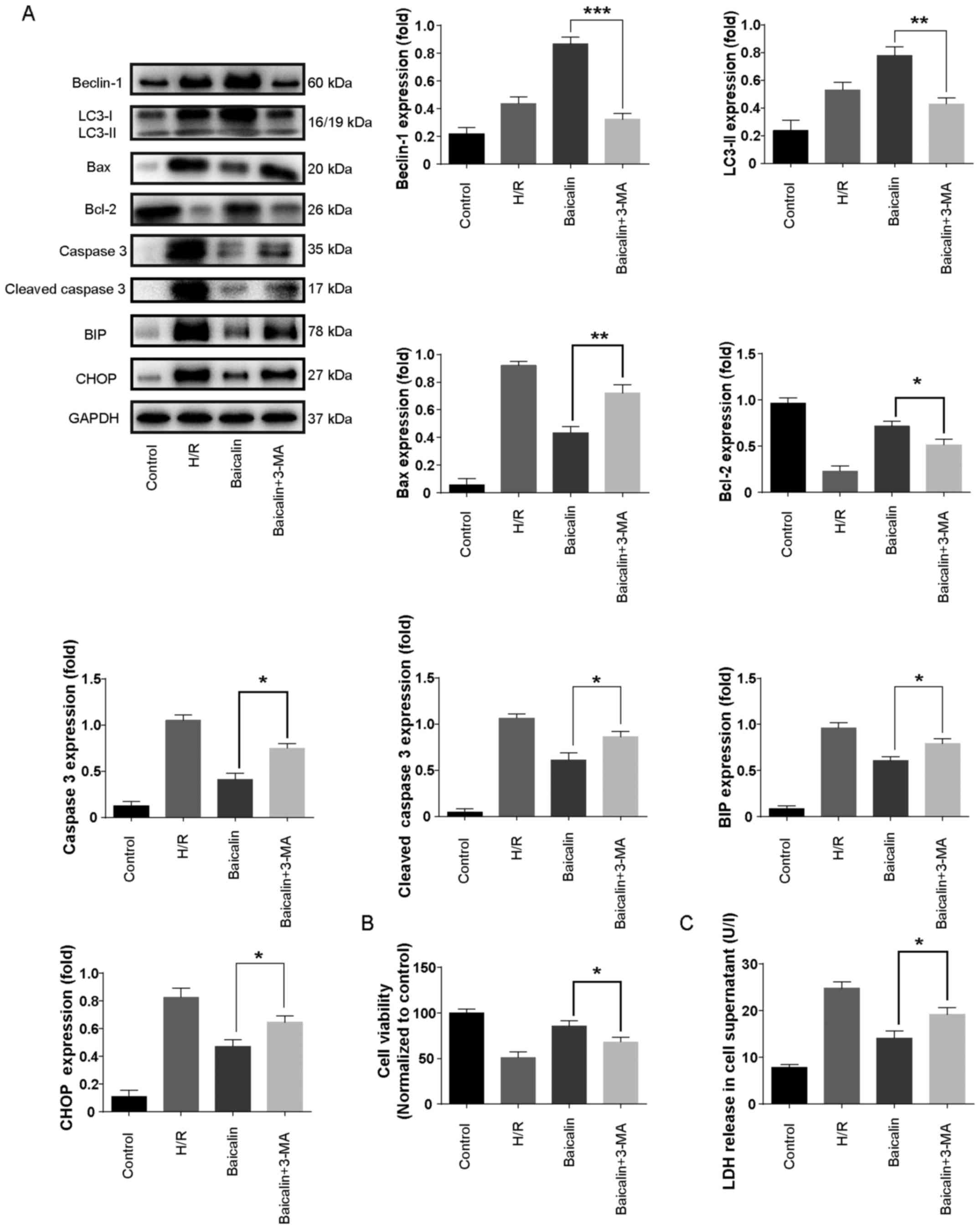 | Figure 5.Autophagy inhibition abrogated the
protective effect of baicalin in vitro. (A) Western blotting
was used to measure the expression of beclin, LC3, Bcl-2, Bax,
caspase 3 and cleaved caspase 3 in LO2 cells. (B) LO2 cell
viability was measured using a Cell Counting Kit-8 assay. (C)
Effects of 3-MA on LDH release in LO2 cells subjected to
hypoxia/reoxygenation. **P<0.05, **P<0.01 and ***P<0.001.
LC3, microtubule-associated proteins 1A/1B light chain 3B; Bcl-2,
B-call lymphoma 2; Bax, Bcl-2-associated X protein; 3-MA,
3-methyladenine; LDH, lactate dehydrogenase. |
To further investigate the effect of autophagy
inhibition in H/R, the expression of caspase-3, cleaved caspase 3,
Bcl-2, Bax, BIP and CHOP, as well as cell viability and LDH levels
were measured. Co-treatment with 3-MA significantly increased the
expression of caspase-3, cleaved caspase 3, Bcl-2, Bax, BIP and
CHOP (Fig. 5A). Furthermore, cell
viability was significantly decreased in the baicalin + 3-MA
treatment group compared with baicalin treatment alone (Fig. 5B). LDH levels were significantly
increased with 3-MA treatment compared with baicalin treatment
alone (Fig. 5C). These results
suggest that autophagy inhibition abrogates the protective effect
of baicalin on apoptosis and ER stress in LO2 cells.
Discussion
As a flavonoid glycoside extracted from the root of
the traditional Chinese medicinal herb Scutellaria
baicalensis, baicalin has been reported to possess a protective
effect against IR injury in multiple organs (12–14).
Zheng et al (13)
demonstrated that baicalin treatment led to significantly reduced
neurological deficit, smaller infarct volume and enhanced
expression of induced myeloid leukemia cell differentiation
protein-1 and BCL-2 in a rat model of cerebral artery
occlusion/reperfusion, suggesting that baicalin's protective effect
against cerebral IR injury is achieved via the inhibition of
ischemia-induced neuronal apoptosis. Kong et al (14) revealed that baicalin improved left
ventricular function, reduced creatine kinase and LDH release in
the coronary effluent, and increased SOD and MDA activity in rats
with IR injury. Lin et al (12) demonstrated that baicalin ameliorates
kidney IR injury by inhibiting the production of proinflammatory
cytokines, including tumor necrosis factor-α and interleukin-1β. In
agreement with previous studies, the results of the present study
demonstrated that baicalin pretreatment is able to attenuate
hepatic injury and inhibit LO2 apoptosis, possibly via the
induction of autophagy.
The upregulation of Bcl-2, downregulation of Bax and
decreased percentage of apoptotic cells in the baicalin group
indicated that apoptosis was decreased following baicalin
pretreatment. Decreased apoptosis is associated with deactivation
of the caspase cascade (22). As a
downstream effector in the caspase cascade, caspase 3 directly
mediates apoptosis when activated by various upstream signals
(22) and as such has been reported
to be a crucial modulator of apoptosis. In the present study,
caspase 3 and cleaved caspase 3 were upregulated in the H/R group,
but baicalin pretreatment was able to significantly decrease their
expression, indicating that baicalin inhibits LO2 cells
apoptosis.
ER stress is a cellular stress condition that
disrupts ER function and results in massive accumulation of
unfolded or misfolded proteins in the ER lumen (25). To maintain ER homeostasis, a series
of adaptive responses called the unfolded protein response (UPR) is
triggered upon ER stress (12). This
process is mediated by 3 major stress sensors, namely
inositol-requiring protein 1 (IRE1), ATF6 and protein kinase
RNA-like ER kinase (22). UPR is
associated with decreased protein load and upregulated ER
chaperones, including BIP (22).
With persisting ER stress, UPR fails and cells progress into the
apoptosis phase, in which CHOP accumulation, IRE1 phosphorylation
and c-JUN N-terminal kinase are activated (22). Herein, BIP and CHOP are regarded as
vital markers of ER stress. In the present study, the expression of
BIP and CHOP were increased in the H/R group and decreased by
baicalin pretreatment, implying that ER stress occurs following H/R
and may be successfully inhibited by baicalin pretreatment.
Autophagy is a lysosome-dependent dynamic
intracellular pathway that involving the delivery and degradation
of unfolded or misfolded proteins (26). It has been documented that ER stress
is a potent trigger of autophagy (27). In the process of UPR, ATF4 and XBP-1
induce autophagy (23,24). In the present study, increased ATF4
and XBP-1 levels were observed in the H/R group. Similarly, beclin
1 and LC3-II were upregulate in the H/R group. Pretreatment with
baicalin inhibited the expression of ATF4 and XBP-1; however,
beclin 1 and LC3-II levels remained high in the baicalin groups,
particularly at the 100 µmol/l dosage, revealing that baicalin
pretreatment inhibited ER-induced autophagy and that baicalin is a
potent inducer of autophagy. This ability is similar to that of a
previously reported immunosuppressive drug, rapamycin (28). Zhu et al (28) revealed that, in a mouse model of,
ischemia reperfusion injury and thapsigargin-treated primary
hepatocytes, ATF4 and XBP1 were markedly upregulated, whereas
rapamycin treatment downregulated their expression; in addition, it
was also revealed that rapamycin is able to enhance autophagy.
To demonstrate the role of autophagy in the
baicalin-mediated protective effect on H/R, the autophagy inhibitor
3-MA was administered. 3-MA is a commonly used autophagy inhibitor
and is a specific inhibitor of autophagic/lysosomal protein
degradation and the formation of autophagosomes (29). The phosphoinositide 3 kinase
(PI3K)/protein kinase B (AKT)/mammalian target of rapamycin (mTOR)
pathway has been implicated in autophagy (29). AKT is downstream of class I and class
III PI3K (30). Class I PI3K is a
major upstream modulator of mTOR that causes the suppression of
autophagy, while the class III PI3K pathway has been recognized to
activate autophagy through sequestration of cytoplasmic material
(30). It has been reported that
3-MA is able to inhibit the class III PI3K pathway, thus inhibiting
autophagy (31). The results of the
present study revealed that pretreatment with 3-MA markedly
inhibited autophagy, as evidenced by low expression of beclin-1 and
LC3-II. Furthermore, high expression of BIP and CHOP were detected
following the inhibition of autophagy, suggesting that autophagy
inhibition aggravates ER stress. This is consistent with a previous
study, in which autophagy was reported to eliminate accumulated
misfolded proteins and degrade damaged organelles to ameliorate ER
stress (32). Autophagy inhibition
also increased LDH levels, increased the expression of caspase-3,
cleaved caspase 3 and Bax and decreased cell viability and Bcl-2
expression, which indicates a protective effect of baicalin in H/R.
In conclusion, the results of the present study demonstrate that
baicalin may have a protective effect in H/R, possibly mediated by
the induction of autophagy. Although the action and mechanism of
baicalin on liver hypoxia/reoxygenation injury were investigated,
there are certain limitations to the present study. The effect of
baicalin on liver hypoxia/reoxygenation injury was only performed
in vitro and requires further study in vivo. In
addition the PI3K/AKT/mTOR signaling pathway was not investigated
in the present study and represents an important pathway in
autophagy. This should also be further investigated in any
subsequent animal studies.
Acknowledgements
Not applicable.
Funding
The present study was supported by the grants from
the National Natural Science Foundation of China (grant nos.
81400675 and 81603406).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
ZM and FL conceived the study and participated in
the design of the study. JZ and JQ performed the experiments. FL
analyzed the data and drafted the manuscript. GW revised the
manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Carchman EH, Rao J, Loughran PA, Rosengart
MR and Zuckerbraun BS: Heme oxygenase-1-mediated autophagy protects
against hepatocyte cell death and hepatic injury from
infection/sepsis in mice. Hepatology. 53:2053–2062. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Wang Y, Shen J, Xiong X, Xu Y, Zhang H,
Huang C, Tian Y, Jiao C, Wang X and Li X: Remote ischemic
preconditioning protects against liver ischemia-reperfusion injury
via heme oxygenase-1-induced autophagy. PLoS One. 9:e988342014.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Zhang W, Wang M, Xie HY, Zhou L, Meng XQ,
Shi J and Zheng S: Role of reactive oxygen species in mediating
hepatic ischemia-reperfusion injury and its therapeutic
applications in liver transplantation. Transplant Proc.
39:1332–1337. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Serracino-Inglott F, Habib NA and Mathie
RT: Hepatic ischemia-reperfusion injury. Am J Surg. 181:160–166.
2001. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Klune JR and Tsung A: Molecular biology of
liver ischemia/reperfusion injury: Established mechanisms and
recent advancements. Surg Clin North Am. 90:665–677. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Kato Y, Shimazu M, Wakabayashi G, Tanabe
M, Morikawa Y, Hoshino K, Harada H, Kadomura T, Obara H, Urakami H,
et al: Significance of portal venous flow in graft regeneration
after living related liver transplantation. Transplant Proc.
33:1484–1485. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Zheng L, Zhang C, Li L, Hu C, Hu M,
Sidikejiang N, Wang X, Lin M and Rong R: Baicalin ameliorates renal
fibrosis via inhibition of transforming growth factor β1 production
and downstream signal transduction. Mol Med Rep. 15:1702–1712.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Zhu J, Wang J, Sheng Y, Zou Y, Bo L, Wang
F, Lou J, Fan X, Bao R, Wu Y, et al: Baicalin improves survival in
a murine model of polymicrobial sepsis via suppressing inflammatory
response and lymphocyte apoptosis. PLoS One. 7:e355232012.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Waisundara VY, Siu SY, Hsu A, Huang D and
Tan BK: Baicalin upregulates the genetic expression of antioxidant
enzymes in Type-2 diabetic Goto-Kakizaki rats. Life Sci.
88:1016–1025. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Shieh DE, Cheng HY, Yen MH, Chiang LC and
Lin CC: Baicalin-induced apoptosis is mediated by Bcl-2-dependent,
but not p53-dependent, pathway in human leukemia cell lines. Am J
Chin Med. 34:245–261. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Motoo Y and Sawabu N: Antitumor effects of
saikosaponins, baicalin and baicalein on human hepatoma cell lines.
Cancer Lett. 86:91–95. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Lin M, Li L, Li L, Pokhrel G, Qi G, Rong R
and Zhu T: The protective effect of baicalin against renal
ischemia-reperfusion injury through inhibition of inflammation and
apoptosis. BMC Complement Altern Med. 14:192014. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Zheng WX, Cao XL, Wang F, Wang J, Ying TZ,
Xiao W, Zhang Y, Xing H, Dong W, Xu SQ, et al: Baicalin inhibiting
cerebral ischemia/hypoxia-induced neuronal apoptosis via
MRTF-A-mediated transactivity. Eur J Pharmacol. 767:201–210. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Kong F, Luan Y, Zhang ZH, Cheng GH, Qi TG
and Sun C: Baicalin protects the myocardium from
reperfusion-induced damage in isolated rat hearts via the
antioxidant and paracrine effect. Exp Ther Med. 7:254–259. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Lin C, Tsai SC, Tseng MT, Peng SF, Kuo SC,
Lin MW, Hsu YM, Lee MR, Amagaya S, Huang WW, et al: AKT
serine/threonine protein kinase modulates baicalin-triggered
autophagy in human bladder cancer T24 cells. Int J Oncol.
42:993–1000. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Zhang X, Tang X, Liu H, Li L, Hou Q and
Gao J: Autophagy induced by baicalin involves downregulation of
CD147 in SMMC-7721 cells in vitro. Oncol Rep. 27:1128–1134. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Shintani T and Klionsky DJ: Autophagy in
health and disease: A double-edged sword. Science. 306:990–995.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Geronimo-Olvera C, Montiel T,
Rincon-Heredia R, Castro-Obregón S and Massieu L: Autophagy fails
to prevent glucose deprivation/glucose reintroduction-induced
neuronal death due to calpain-mediated lysosomal dysfunction in
cortical neurons. Cell Death Dis. 8:e29112017. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Rautou PE, Mansouri A, Lebrec D, Durand F,
Valla D and Moreau R: Autophagy in liver diseases. J Hepatol.
53:1123–1134. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Kaushal GP: Autophagy protects proximal
tubular cells from injury and apoptosis. Kidney Int. 82:1250–1253.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Yao W, Li H, Han X, Chen C, Zhang Y, Tai
WL, Xia Z and Hei Z: MG53 anchored by dysferlin to cell membrane
reduces hepatocyte apoptosis which induced by ischaemia/reperfusion
injury in vivo and in vitro. J Cell Mol Med. 21:2503–2513. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Lin M, Li L, Zhang Y, Zheng L, Xu M, Rong
R and Zhu T: Baicalin ameliorates H2O2 induced cytotoxicity in HK-2
cells through the inhibition of ER stress and the activation of
Nrf2 signaling. Int J Mol Sci. 15:12507–12522. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Xue LX, Liu HY, Cui Y, Dong Y, Wang JQ, Ji
QY, He JT, Yao M, Wang YY, Shao YK, et al: Neuroprotective effects
of Activin A on endoplasmic reticulum stress-mediated apoptotic and
autophagic PC12 cell death. Neural Regen Res. 12:779–786. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Kishino A, Hayashi K, Hidai C, Masuda T,
Nomura Y and Oshima T: XBP1-FoxO1 interaction regulates ER
stress-induced autophagy in auditory cells. Sci Rep. 7:44422017.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Hoyer-Hansen M and Jäättelä M: Connecting
endoplasmic reticulum stress to autophagy by unfolded protein
response and calcium. Cell Death Differ. 14:1576–1582. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
B'Chir W, Maurin AC, Carraro V, Averous J,
Jousse C, Muranishi Y, Parry L, Stepien G, Fafournoux P and Bruhat
A: The eIF2α/ATF4 pathway is essential for stress-induced autophagy
gene expression. Nucleic Acids Res. 41:7683–7699. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Margariti A, Li H, Chen T, Martin D,
Vizcay-Barrena G, Alam S, Karamariti E, Xiao Q, Zampetaki A, Zhang
Z, et al: XBP1 mRNA splicing triggers an autophagic response in
endothelial cells through BECLIN-1 transcriptional activation. J
Biol Chem. 288:859–872. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Zhu J, Hua X, Li D, Zhang J and Xia Q:
Rapamycin attenuates mouse liver ischemia and reperfusion injury by
inhibiting endoplasmic reticulum stress. Transplant Proc.
47:1646–1652. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Hermann R, Vélez DE, Rusiecki TM,
Fernández Pazos Mde L, Cordero Mestre VE, Prendes Marina MG,
Rossini Perazzo JC, Savino EA and Varela A: Effects of
3-methyladenine on isolated left atria subjected to simulated
ischaemia-reperfusion. Clin Exp Pharmacol Physiol. 42:41–51. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Kim SH, Son KM, Kim KY, Yu SN, Park SG,
Kim YW, Nam HW, Suh JT, Ji JH and Ahn SC: Deoxypodophyllotoxin
induces cytoprotective autophagy against apoptosis via inhibition
of PI3K/AKT/mTOR pathway in osteosarcoma U2OS cells. Pharmacol Rep.
69:878–884. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Chen W, Sun Y, Liu K and Sun X: Autophagy:
A double-edged sword for neuronal survival after cerebral ischemia.
Neural Regen Res. 9:1210–1216. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Cai Y, Arikkath J, Yang L, Guo ML,
Periyasamy P and Buch S: Interplay of endoplasmic reticulum stress
and autophagy in neurodegenerative disorders. Autophagy.
12:225–244. 2016. View Article : Google Scholar : PubMed/NCBI
|