Introduction
Spinal cord injury (SCI) is the leading cause of
paralysis, resulting in a loss of function below the injury level
(1). Unlike the peripheral nervous
system, damaged neurons in central nervous system are rarely
regenerated (2). Neural stem cells
(NSCs), which are capable of generating multiple cell lineages,
including neurons and glial cells, have the potential to repair and
replace damaged neurons (3,4). Up until now, stem cell-based repair
strategies for SCI remain a highly investigated area of research.
Various genetically modified stem cells have been transplanted into
SCI models, although the results are not always successful
(5,6).
SCI is a complex injury that includes a primary
injury (mechanical damage to the spinal cord) and a secondary
injury (characterized by prolonged inflammatory cell infiltration,
glial scar formation and tissue damage) (7). The altered microenvironment is no
longer suitable for cell survival, and ultimately excessive glial
scars hinder the regeneration of damaged axons (8,9). A
number of previous studies have confirmed that the transplantation
of NSCs and embryonic stem cells (ESCs) promotes neurological
recovery by decreasing the expression level of inflammatory
cytokines (10,11).
Cadherins, including N-cadherin and E-cadherin, are
a type of cell adhesion molecule that are vital for cell
proliferation and migration (12). A
previous study has demonstrated that N-cadherin promotes
recruitment and migration of NSCs (13). However, the expression of N-cadherin
and E-cadherin in NSCs are negatively correlated (14). Thus far, few studies have
investigated the association between E-cadherin and NSCs.
In the present study, the expression of E-cadherin
was observed in subcultured NSCs and differentiated NSCs.
Furthermore, an E-cadherin overexpression lentivirus was
constructed and transfected into NSCs in order to investigate the
effect of E-cadherin on NSCs viability, migration, differentiation
and neurosphere formation. An in vivo study was used to
assess the survival rate of grafted NSCs-E-cadherin. Furthermore,
by analyzing tissue repair, Basso Mouse Scale (BMS) scores and
inflammatory cytokine expression levels the protective role of
E-cadherin in SCI was assessed. Additionally, NSCs or
NSCs-E-cadherin were co-cultured with mouse bone marrow-derived
macrophages (BMDMs) in order to examine whether E-cadherin
influenced the activation of macrophages.
Materials and methods
Cell culture and differentiation
Primary spinal cord derived NSCs were obtained from
Animal Experimental Center of Tongji University (Shanghai, China)
and cultured in Dulbecco's modified Eagle medium: Nutrient Mixture
F-12 (DMEM/F12) supplemented with 1% B27, 20 ng/ml basic fibroblast
growth factor (bFGF) and 20 ng/ml epidermal growth factor (EGF; all
Thermo Fisher Scientific, Inc., Waltham, MA USA) at 37°C in an
atmosphere containing 5% CO2. Following 3–5 days of
culture, the cells grew to form neurospheres and reached the
highest density on the 7th day. Subsequently, a single cell
suspension was prepared through the mechanical separation of the
neurospheres, and the cells were subcultured every 3–5 days.
Following the removal of bFGF and EGF, the cells were induced to
differentiate using 1% fetal bovine serum (FBS; Gibco; Thermo
Fisher Scientific, Inc.).
Construction of E-cadherin
overexpression lentivirus
DNA was extracted from mouse NSCs
The primers were designed and synthesized with the
following sequences: E-cadherin, forward 5′-GGGTCTTGCTATGTTGCC-3′
and reverse 5′-GTTCCGCTCTGTCTTTGG-3′, to amplify the E-cadherin
sequence (hereafter known as fragment) using the PCR amplification
kit (Omega Bio-tek, Inc., Norcross, GA, USA). The PCR thermocycling
conditions were as follows: 94°C for 10 min, followed by 30 cycles
at 94°C for 30 sec, 58°C for 30 sec and 72°C for 90 sec, and a
final extension of 94°C for 15 sec, 60°C for 1 min, 94°C for 15 sec
and 60°C for 15 sec. The fragment was connected with the plasmid,
PHY-027 (EF1A-MCS-CMV-zsGreen1-IRES-Puro; Shanghai Ruisai
Biotechnology Co., Ltd, Shanghai, China), using EcoR I and
Xba I (Takara Biotechnology Co., Ltd., Dalian, China).
Positive clones were identified using polymerase chain reaction and
subsequently sequenced.
Two groups were established: The E-cadherin
overexpression and control groups. The DNA plasmid mixture for the
experimental group consisted of 1 µg E-cadherin plasmid (2 µg/µl),
0.75 µg psPAX2 and 0.25 µg pMD2.G (both Invitrogen; Thermo Fisher
Scientific, Inc., Waltham, MA, USA) in OPTI-MEM (Promega
Corporation, Madison, WI, USA). The DNA plasmid mixture for the
control group consisted of 1 µg empty plasmid (1.8 µg/µl), 0.75 µg
psPAX2 and 0.25 µg pMD2.G in OPTI-MEM in a total volume of 20 µl. A
total of 6 µl FuGENE®6 (Promega Corporation) was added
to OPTI-MEM, then this mixture was added to the DNA plasmid and
incubated at room temperature for 20 min. The mixture was then
transferred into competent 293T cells (Central Laboratory of
Shanghai Tenth People's Hospital, Shanghai, China) for packaging
and incubated at 37°C for 48 h. Cell medium was then collected, and
purified through ultrafiltration and ultracentrifugation. For each
well, 150 µl lentivirus (2×108 TU/ml) with 5 µl diluted
polybrene in DMEM/F12 (5 µg/ml; Sigma-Aldrich; Merck KGaA,
Darmstadt, Germany) was transfected into NSCs at room temperature
and then the cells were incubated at 37°C for 48 h for subsequent
experiments.
Viability and migration assays
Cell viability was evaluated using an MTT assay
(Sigma-Aldrich; Merck KGaA). Cells were digested with trypsin, and
10% FBS was added to prevent serum interference. The cell
concentration was adjusted to 4×104 cells/well. A total
of 20 µl MTT stock solution was added per well and incubated in
37°C for 4 h. The samples were centrifuged and dimethyl sulfoxide
was then added. Following 10 min of incubation, the absorbance
values were recorded at 490 nm and the growth curve was drawn.
For migration, a Transwell assay was performed with
a Transwell chamber (ECM550, Chemicon International, Inc.,
Billerica, MA, USA) was used. In the upper chamber,
1×105 NSCs in 100 µl DMEM/F12 and 0.05% bovine serum
albumin (R&D Systems, Inc., Minneapolis, MN, USA) were added.
In the lower chamber, 300 µl DMEM/F12 with 10% FBS medium was
added. Following 24 h incubation, the cells were stained with 1%
crystal violet for 20 min at room temperature and counted under a
light microscope (magnification, ×100).
Western blotting
The protein samples were extracted using 1X SDS
loading buffer. Protein concentration was determined using
bicinchoninic acid assay quantification. A total of 20 µg protein
was loaded in each well, separated with SDS-PAGE (10% gels) and
blotted on polyvinylidene fluoride membranes (100 v for 120 min).
The membranes were blocked with Tris-buffered saline with Tween-20
(TBST) solution containing 5% fat-free milk for 1 h at room
temperature. Primary antibodies for E-cadherin (1:1,000; cat. no.
MAB7481; R&D Systems, Inc.) and β-actin (cat. no. ab8229;
Abcam, Cambridge, UK) were incubated overnight at 4°C. The
membranes were washed using TBST 3 times and incubated with
horseradish peroxidase-conjugated whole immunoglobulin G
Affinity-Purified Antibodies (Goat Anti-Rabbit; 1:5,000; cat. no.
111-005-045; Jackson Immuno Research Laboratories, Inc., West
Grove, PA, USA) at room temperature for 1 h. Signals were detected
using electrochemiluminescence reagent (Sigma-Aldrich; Merck KGaA).
The bands were analyzed using Image Lab software (version 3.0;
Bio-Rad Laboratories, Inc., Hercules, CA, USA).
Reverse transcription-quantitative PCR
(RT-qPCR)
Total RNA was extracted from NSCs using TRIzol
Reagent (Invitrogen; Thermo Fisher Scientific, Inc.). RNA
quantification was performed using a NanoDrop™ 2000c
Spectrophotometer (Thermo Fisher Scientific, Inc.). cDNA was
synthesized using the Reverse Transcription kit (Applied
Biosystems; Thermo Fisher Scientific, Inc.). qPCR was performed
using SYBR-Green (Thermo Fisher Scientific, Inc.) and CFX384 Touch
Real-Time (Bio-Rad Laboratories, Inc.). The PCR cycling conditions
were as follows: 94°C for 10 min, 30 cycles at 94°C for 30 sec,
58°C for 30 sec and 72°C for 1.5 min, and finally 72°C for 5 min.
The primer sequences used were as follows: GAPDH, forward
5′-CCTGGAGAAACCTGCCAAGTA-3′ and reverse
5′-TCATACCAGGAAATGAGCTTGAC-3′; interleukin (IL)-1β, forward
5′-TGCCACCTTTTGACAGTGATG-3′ and reverse 5′-TGTGCTGCTGCGAGATTTG-3′;
IL-6, forward 5′-CAACCACGGCCTTCCCTACT-3′ and reverse
5′-CACAACTCTTTTCTCATTTCCACGA-3′; IL8, forward
5′-GCATCTCGGTGTAGAGCAAGG-3′ and reverse
5′-CAGTGTCCCTATAGCCCAAGTTA-3′; monocyte chemoattractant protein 1
(MCP-1), forward 5′-CCTGGATCGGAACCAAATG-3′ and reverse
5′-TGGTTGTGGAAAAGGTAGTGG-3′; inducible nitric oxide synthase
(iNOS), forward 5′-TGTGCGAAGTGTCAGTGGCT-3′ and reverse
5′-CCCTTTGTGCTGGGAGTCAT-3′; tumor necrosis factor α (TNFα), forward
5′-GAGTCCGGGCAGGTCTACTT-3′ and reverse 5′-GTCACTGTCCCAGCATCTTGT-3′;
arginase-1 (Arg1), forward 5′-CTTTCCACGACGGTGACACG-3′ and reverse
5′-ACGCTAGGATCTGATCGCGG-3′. GAPDH was used as an internal reference
gene for normalization using the 2−ΔΔCq method (15).
SCI model
A total of 60 female, adult wild-type C57BL/6 mice
(weighing 20 g, aged 8–10 weeks old) were obtained from Animal
Experimental Center of Tongji University (Shanghai, China) and used
for injury model construction. The groups were as follows (15
mice/group): Sham, PBS treatment, NSCs treatment and E-cadherin
overexpression NSCs (NSCs-E-cad) treatment. The mice were
intraperitoneally anesthetized and a T9 laminectomy was performed
to expose the spinal cord. Subsequently, the injury model was
constructed by clamping the spinal cord at T9 for 30 sec and the
wound was stitched and disinfected. The mice were kept warm during
the procedure. Following recovery from anesthesia, the mice
displayed a lower limb motor deficit (motor function loss) and were
housed in a temperature-controlled room at 27°C. Food and water
were provided ad libitum and the mice received manual
bladder evacuation once a day following surgery.
The sham group did not undergo the SCI procedure.
The NSCs and NSCs-E-cad group underwent the SCI procedure and
subsequently had cell transplantations. The PBS group underwent the
SCI procedure and were subsequently administered 2 µl PBS 7 days
after the procedure; PBS was injected at two sites (1-mm rostral
and caudal to the injury epicenter).
Cell transplantation
Cell transplants were performed 7 days post injury
(dpi). The mice were anesthetized and the laminectomy site was
re-exposed. The NSCs and NSCs-E-cad were resuspended using PBS and
the final concentration was adjusted to 1×105 cells/µl
(kept on ice prior to injection). For each mouse, 2-µl cell
suspensions were injected at two sites (1-mm rostral and caudal to
the injury epicenter) using a 5-µl microinjector (Hamilton syringe;
Hamilton Bonaduz AG, Bonaduz, Switzerland). The mice were injected
at a rate of 0.5 µl/min.
Immunohistochemistry
A total of 2 weeks after cell transplantation,
animals were anesthetized, and perfused with PBS and 4%
paraformaldehyde. The spinal cords were removed and post-fixed
overnight at 4°C. The fixed tissues were cryoprotected in 30%
sucrose (Sigma-Aldrich; Merck KGaA) in PBS at 4°C, and then
embedded in optimal cutting temperature compound (Tissue Tek;
Sakura Finetek USA, Inc., Torrance, CA, USA). Cryostat sections
that were 10 µm thick were cut and affixed to glass slides and
stored at −20°C. Following this, the sections were processed for
histochemistry, PBS with Tween 20 was used for permeabilization and
10% FBS was used for blocking for 1 h at room temperature. Sections
were then incubated with the primary antibodies for Nestin (cat.
no. ab22035), neuron-specific class III β-tubulin (cat. no.
ab18207) and glial fibrillary acidic protein (cat. no. ab7260; all
1:500; Abcam) at 4°C overnight. Following this, goat anti-rabbit
secondary antibodies (cat. no. ab150117; Abcam) were incubated for
1 h at room temperature. Subsequent to washing with PBS, nuclei
were stained using 4′,6-diamidino-2-phenylindole (Abcam) at room
temperature for 30 min. The processed slides were assessed under a
fluorescent microscope (magnification, ×200 or ×400).
Functional recovery assessment
Tissue sparing of the injured spinal cord was
quantified using ImageJ 1.50 software (National Institutes of
Health, Bethesda, MD, USA) on the basis of immunohistochemical
stains. Motor function was assessed using BMS scores (9). The BMS score was recorded at the 1st
day post-transplantation. A total of 3 days post-transplantation,
5/15 mice in each group were sacrificed to obtain spinal cord
tissue to investigate the differences in the expression of
inflammatory cytokines among the three groups. A total of 6 weeks
post transplantation, 7/10 in each group were sacrificed to assess
cell survival and tissue sparing. The remaining mice were
sacrificed at 3 months post-transplantation for checking cell
survival in the long run.
Mouse BMDM preparation
BMDMs were harvested from bone marrow, which was
collected from the femoral shafts of 5 adult female C57BL/6 mice
(Animal Experimental Center of Tongji University, Shanghai, China)
aged 8 weeks old and weighing 20 g by flushing the marrow cavity
with DMEM. Animals were housed in a room at a temperature of 24°C
and 40% humidity with 12-h dark and light cycle; they had free
access to standard food and water. Cells were cultured in DMEM
supplemented with 10% FBS, 1% Penicillin-Streptomycin and 15%
conditioned medium (with macrophage colony-stimulating factor;
Gibco; Thermo Fisher Scientific, Inc.).
Interferon-γ induction (INF-γ)
BMDMs and NSCs were co-cultured in 6-well plate
(5×105 cells/well) at 37°C in a 5% CO2
atmosphere for 12 h. The cells were cultured in DMEM supplemented
with 5% newborn calf serum (Rocky Mountain Biologicals, Inc.,
Missoula, MT, USA). Then, treated with INF-γ at a final
concentration of 5 ng/ml for 6 h. INF-γ is usually used to induce
the activation of M1 macrophages (16). The cells were centrifuged at 3,000 ×
g for 20 min at room temperature and the supernatant collected for
subsequent experiments.
Statistical analysis
Images were quantified using ImageJ software with
Java 1.8.0. Data are presented as the mean ± standard deviation
from three independent experiments. For MTT and BMS results,
one-way analysis of variance (ANOVA) followed by the
Student-Newman-Keuls (SNK) post hoc test for repeated measurements
was used for statistical analysis. Non-normal distribution data was
converted to normal distribution data by using logarithmic
transformation performed by SPSS13.0 (SPSS, Inc., Chicago, IL,
USA). A Student's t-test and one-way ANOVA followed by the SNK post
hoc test were used for comparing the difference between two and
more than two groups, respectively. Statistical analysis was
performed with GraphPad Prism 7 software (GraphPad Software, Inc.,
La Jolla, CA, USA). P<0.05 was considered to indicate a
statistically significant difference.
Results
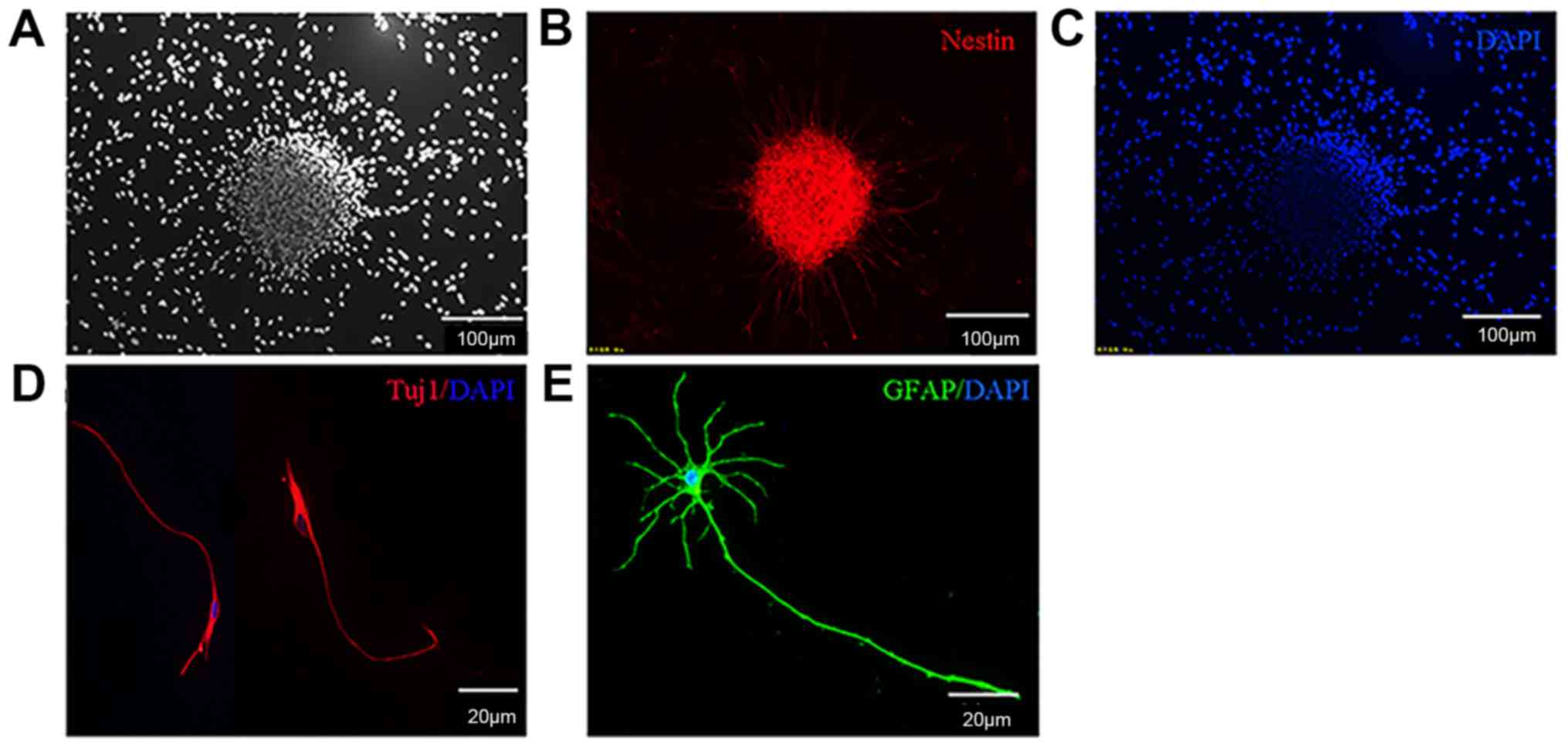
NSCs form neurospheres and
differentiate into neurons and astrocytes
NSCs were obtained from E13.5 of fetal mice spinal
cord tissues and cultured in serum-free growth medium. The
undifferentiated NSCs expressed the specific marker Nestin.
Following 7 days in culture, NSCs were able to proliferate and form
neurospheres (Fig. 1A-C). These
primary neurospheres were mechanically separated into single cell
suspensions and were further passaged every 2–3 days. A total of 3
days following the removal of bFGF and EGF, the cells
differentiated into neurons and astrocytes (Fig. 1D and E).
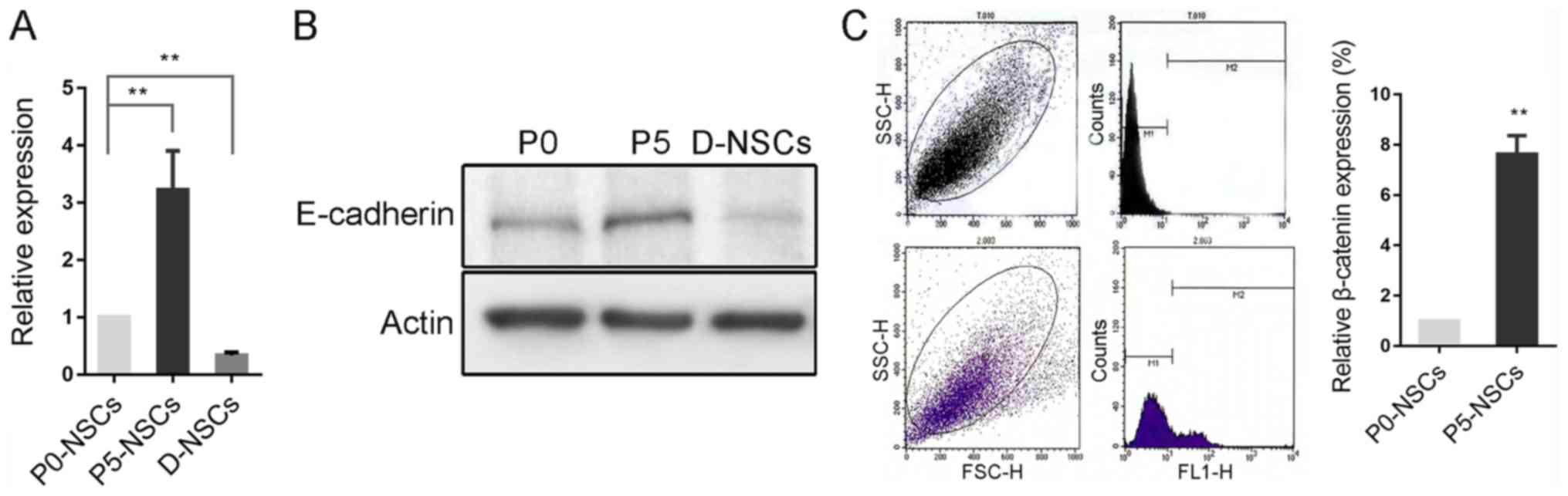
E-cadherin expression increases in
subcultured NSCs but decreases following induced
differentiation
E-cadherin expression was significantly increased in
passage 5 (P5) NSCs compared with P0 cells, and decreased following
differentiation, which was further confirmed by western blotting
(P<0.01; Fig. 2A and B). However,
according to flow cytometry results, β-catenin, a key component of
the E-cadherin/catenin complex (17)
was also significantly increased in this process (P<0.01;
Fig. 2C).
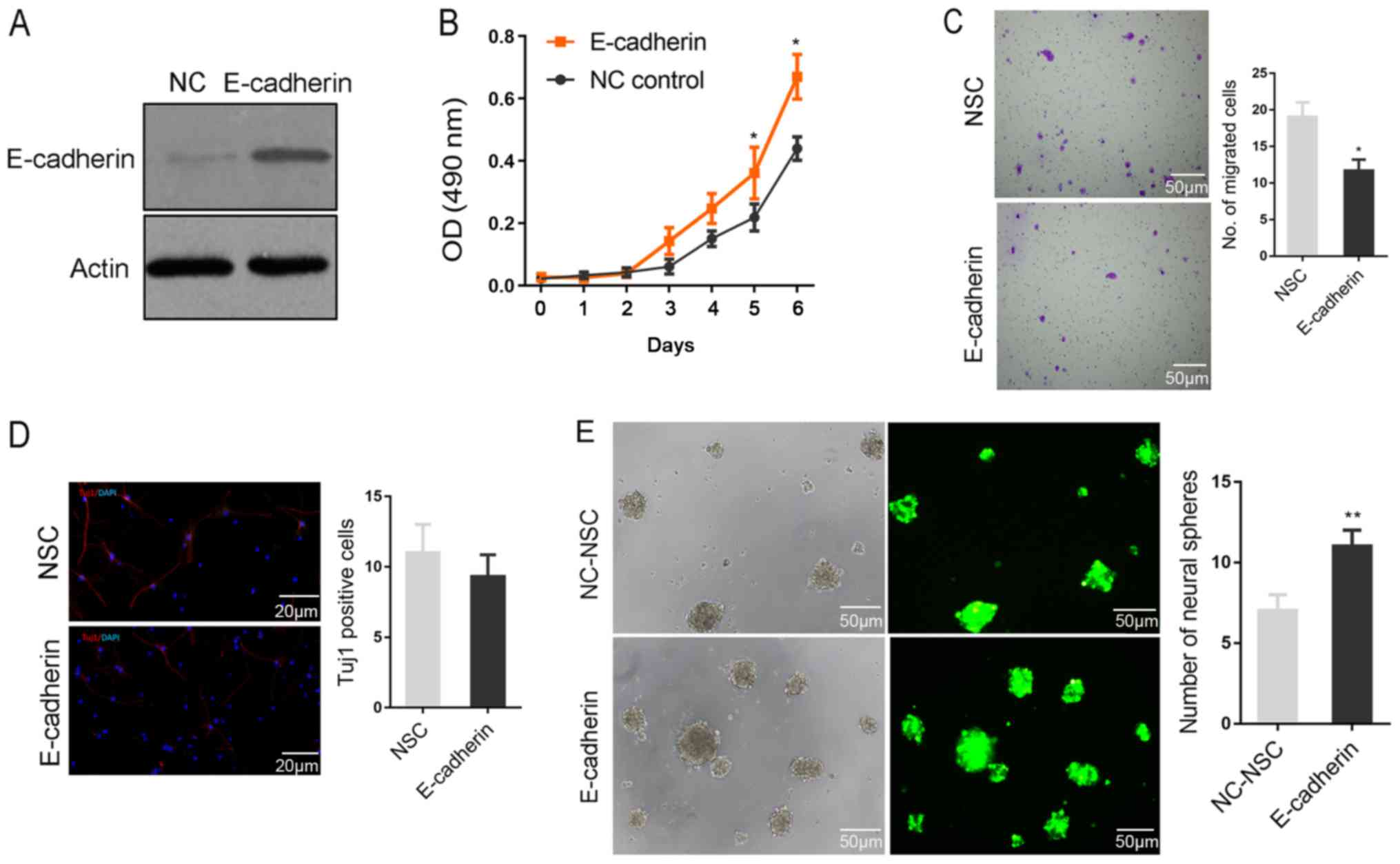
Function of E-cadherin in regulating
the biological behaviors of NSCs
To further investigate the function of E-cadherin in
regulating the biological behaviors of NSCs, the lentivirus vector
PHY-027 carrying E-cadherin and green fluorescent protein was
constructed. As presented in Fig.
3A, the lentivirus was able to effectively promote E-cadherin
protein expression following transfection into NSCs. The ectopic
expression of E-cadherin appeared to significantly facilitate NSC
viability, but significantly inhibited cell migration compared with
the NSC group (all P<0.05; Fig. 3B
and C). Subsequent to induced differentiation, no significant
difference in Tuj1-positive cells was identified between the two
groups, indicating that E-cadherin did not significantly affect the
fate determination of NSCs (Fig.
3D). The rate of neurosphere formation is considered a primary
characteristic of stem cells (18).
Notably, the rate of neurosphere formation was significantly higher
in the E-cadherin group compared with the NC-NSC group, indicating
that E-cadherin maintained the stemness of NSCs (P<0.01;
Fig. 3E).
E-cadherin improves the survival of
grafted NSCs in the short term
A T9 laminectomy was performed to expose the spinal
cord (Fig. 4A) The injury model was
constructed by clamping the spinal cord at T9 for 30 sec, and two
clear red lines were observed at either side of the midline
(Fig. 4B). Subsequent to surgery,
the mice displayed a lower limb motor deficit. Disruption of tissue
was revealed in the injured spinal cord (Fig. 4C). A total of 3 months post-surgery,
the injured spinal cord was able to form a large cavity surrounded
by GFAP-positive glial cells (Fig.
4D). The grafted NSCs were able to migrate to the lesion site
and increased cell survival around the epicenter was observed in
the E-cadherin group at 14 dpi compared with the control group
(P<0.05; Fig. 4E).
 | Figure 4.E-cadherin improved the survival of
grafted NSCs in the short term. (A) T9 laminectomy was performed to
exposure spinal cord. (B) Subsequent to clamping the spinal cord at
T9 for 30 sec, two red lines were observed at either sides of the
midline (black arrow). (C) The disruption of the tissue was
observed in the injured spinal cord. (D) A total of 3 months post
surgery, the injured spinal cord was able to form a large cavity
surrounded by GFAP positive glial cells. (E) Transplanted NSCs were
able to migrate to the lesion site, and E-cadherin improved the
survival of NSCs around the epicenter in the short term. *P<0.05
vs. NSC. NSC, neural stem cells; GFAP, glial fibrillary acidic
protein. DAPI, 4′,6-diamidino-2-phenylindole; GFAP, glial
fibrillary acidic protein; GFP, green fluorescent protein; SCI,
spinal cord injury. |
E-cadherin served as a
‘neuroprotector’ following SCI
A total of 6 weeks post-transplantation, mice
tissues were sectioned and stained with GFAP. As presented in
Fig. 5A, compared with the sham
group, PBS treatment resulted in large tissue cavities, and the
caudal and rostral of the lesion site were not fully connected.
Furthermore, the total volume of the spinal cord was substantially
reduced. Conversely, the E-cadherin group demonstrated increased
tissue sparing compared with the PBS and NSCs groups, with the
exception of −2,000 and −3,000 um; at these distances, tissue
sparing in the E-cadherin group was significantly increased
compared with the PBS group (all P<0.05; Fig. 5B). In line with the aforementioned
results, the highest BMS scores were obtained in the E-cadherin
group. BMS scores in the E-cadherin group were significantly
greater compared with the PBS and NSC groups 3–6 weeks after
transplantation, but only significantly greater compared with the
PBS group 2 weeks after transplantation (all P<0.05; Fig. 5C). These results indicate improved
motor functional recovery in the E-cadherin group compared with the
PBS and NSC groups. Therefore, suggesting that grafted NSCs with
highly expressed E-cadherin may promote motor function recovery and
create an improved microenvironment for neural regeneration.
E-cadherin alleviates secondary damage
by reducing the release of inflammatory cytokines
To determine whether E-cadherin was able to promote
motor function recovery via regulating inflammatory responses
following SCI, several associated pro-inflammatory cytokines,
including IL-1β, IL-6, IL-8, MCP-1, iNOS and TNF-α were detected in
the injured spinal cord tissue in each group 3 days
post-transplantation. These cytokines were significantly higher in
the injured spinal cord group compared with the sham group,
suggesting that the inflammatory response was activated following
SCI (all P<0.01; Fig. 6A). The
release of these cytokines was significantly reduced in the
E-cadherin group compared with the NSC group (all P<0.05).
Notably, iNOS and TNF-α are regarded as M1 macrophage markers, and
M1 macrophage infiltration is considered to be the primary cause of
further damage by secreting inflammatory cytokines following
primary mechanical injury (16). On
these grounds, mouse BMDMs were cultured with NSCs or
NSCs-E-cadherin to investigate whether E-cadherin influenced M1
macrophage activation. Upon type II IFN-γ induction, the expression
of IL-1β, iNOS and TNF-α in the PBS group was significantly
increased compared with the control group (all P<0.01; Fig. 6B), indicating that the M1 macrophage
was activated. Compared with the NSC group, E-cadherin exhibited
the most significant inhibitory effect on the release of these
cytokines (all P<0.05). To rule out the possibility that the
pro-cytokines were partly released by NSCs, cyrtokine release was
assessed in the NSC in response to INF-γ treatment. The release of
cytokines was only significantly increased, compared with the NSCs,
in NSC and macrophage co-cultures (Fig.
6C). No significant difference was observed prior to and
post-INF-γ treatment in NSCs alone, indicating that NSCs may
respond to INF-γ, but not release inflammatory cytokines. These
results may partly explain why E-cadherin promoted motor function
recovery.
 | Figure 6.E-cadherin alleviated secondary
damage by reducing the release of inflammatory cytokines. (A) The
release of inflammatory cytokines (IL-1β, IL-6, IL-8, MCP-1, iNOS
and TNF-α) was increased in the PBS group 3 days after
transplantation. (B) INF-γ was able to induce the ectopic
expression of IL-1β, iNOS and TNF-α released by activated
macrophages, and NSCs alone were able to reduce the expression of
these cytokines, while NSCs with E-cadherin exhibited the most
marked inhibitory effect. (C) Cytokines were detected in NSCs alone
in response to INF-γ treatment. No obvious difference was observed
before and after INF-γ treatment. *P<0.05, **P<0.01 as
indicated; ##P<0.01 vs. NSCs. NSCs, neural stem
cells; IL, interleukin; MCP-1, monocyte chemoattractant protein 1;
iNOS, inducible nitric oxide synthase; TNF-α, tumor necrosis factor
α; IFN-γ, type II interferon. |
Discussion
The traditional concept that injured neurons cannot
regenerate has been challenged since NSCs were identified in 1992
(19). The hallmark characteristics
of NSCs include the ability to self-renew and generate multiple
cell lineages, including neurons, astrocytes and oligodendrocytes
(20). Consequently, grafted NSCs
have the potential to replace injured neurons and reconstruct
neural circuits in neurodegenerative/traumatic diseases (21). Basic transplantation research
including clinical applications have been performed for decades,
but despite the few favorable results gained, there are still
numerous unsolved issues, including a lower grafted cell survival
rate, restricted differentiation of glial lineage and the cells
rarely integrating with host cells (22–24).
In addition to neuronal differentiation advantages,
the anti-inflammatory effects of NSCs have been demonstrated in
vitro and in vivo following SCI. NSCs have been
demonstrated to influence the phenotypes of macrophages and secrete
immunosuppressive factors (25–28). The
pathophysiology of acute SCI comprises the primary injury
(mechanical damage of the spinal cord) and the secondary injury (a
cascade process initiated hours to days after the primary injury)
that is characterized by a prolonged inflammatory response
resulting in inflammatory cell infiltration, glial scar formation
and further tissue damage (29–32).
The adhesion protein E-cadherin has been identified
as an epithelial-to-mesenchymal transition marker and is vital for
tumor progression and metastasis (33,34). As
a transmembrane protein, its functions involve the regulation of
cell adhesion and mediation of extracellular matrix signaling
(35). E-cadherin has been
recognized as one of the key factors to maintain the morphological
structure and functional integrity of epithelial cells (36–38).
Several studies have confirmed the function of E-cadherin in stem
cells. For example, Eastham et al (39) identified E-cadherin as a marker for
undifferentiated human ESCs (hESCs), which were used to demarcate
differentiated and undifferentiated hESCs. D'Amour et al
(40) demonstrated that E-cadherin
is co-expressed with typical undifferentiated markers of hESCs
(including stage-specific embryonic antigen-4) and its expression
is decreased immediately following the induction of
differentiation. Furthermore, E-cadherin directly regulates hESC
survival and self-renewal. Upregulated E-cadherin substantially
enhances the cloning efficiency and pluripotency of hESCs (41).
However, the function of E-cadherin in regulating
the biological behavior of spinal cord-derived NSCs remains
unknown. In the present study, the increased expression of
E-cadherin in subcultured NSCs (P5) compared with P0 cells was
observed. β-catenin, localized at the core of the
E-cadherin/catenin complex, was also increased. Notably, a previous
study revealed that β-catenin was involved in Wnt signaling, which
has been proven to control mouse embryonic stem cell stemness
(42). MTT and Transwell assays were
performed, and the results revealed that the ectopic expression of
E-cadherin was able to maintain the stemness of NSCs by enhancing
cell viability and abrogating cell motility. Similar results have
been obtained in other cell types (43–45). Jin
et al (46) demonstrated that
germline stem cells (GSCs) expressing more E-cadherin become more
competitive compared with the neighboring GSCs occupying the niche,
which is the fundamental requirement for cell self-renewal.
Furthermore, E-cadherin-mediated cell adhesion has been confirmed
to serve the principal role to preserve the stemness state of mouse
ESCs (47). Karpowicz et al
(48) proposed that E-cadherin
regulates the self-renewal of embryonic forebrain ventricles
derived NSCs.
Since E-cadherin had a positive effect on NSC
viability in vitro, NSCs-E-cadherin were transplanted into
the SCI mice to further investigated whether it promoted long-term
cell survival in vivo. The grafted cells were revealed to be
capable of surviving. Furthermore, they were localized around
damaged axons. Unexpectedly, E-cadherin demonstrated no significant
effect on cell survival in the long term, and the altered niche was
favorable to glial differentiation. However, NSCs with highly
expressed E-cadherin substantially maximized spinal cord integrity
and ultimately improved motor function recovery. Those combined
results indicated that E-cadherin served a neuroprotective role in
SCI.
The expression levels of inflammatory cytokines,
including IL-1β, IL-6, IL-8, MCP-1, iNOS and TNF-α, were recorded.
By comparison, E-cadherin substantially reduced the expression of
these cytokines and demonstrated the highest inhibition compared
with the PBS and NSCs groups. This implies that E-cadherin relieves
the inflammatory response following SCI. Interestingly, as iNOS and
TNF-α are regarded as M1 macrophage markers and M1 macrophage
infiltration is considered to be the main cause of secondary injury
cascade (14), the present study
co-cultured BMDMs with NSCs or NSCs-E-cadherin to investigate
whether E-cadherin influenced M1 macrophage activation. As
expected, compared with the PBS and NSC groups, E-cadherin
exhibited the most significant inhibitory effect on IL-1β, iNOS and
TNF-α expression induced by IFN-γ. These results partly explained
why E-cadherin promoted motor functional recovery following SCI. At
present, there are few studies that have investigated the
association between E-cadherin and inflammation. Shie et al
(49) reported lower E-cadherin
expression in patients with interstitial cystitis, which is
associated with higher apoptotic cell numbers, higher visual analog
scale pain scores and chronic inflammation. The adherens junction
complex (E-cadherin/β-catenin), together with tight junctions and
desmosomes, controls the epithelial cell-to-cell adherence and
forms a selective barrier. Disruption of this barrier is associated
with inflammatory bowel disease (50). It has been suggested that E-cadherin
mediated cell-to-cell connection, which is vital for the
maintenance of epithelial integrity, the disruption of which is
sufficient to initiate the development of inflammation (51–53).
Tsai et al (54) revealed
that proinflammatory cytokines and activated immune cells
contribute to the disarrangement of the E-cadherin/catenin complex
and cause the shedding of E-cadherin from the cell surface, and
substantially elevated E-cadherin in plasma was identified in
patients with pelvic inflammatory disease. In addition, activated
neutrophils have been confirmed to induce the decomposition of the
E-cadherin/β-catenin complex, resulting in the enhanced migration
of immune cells (55). Therefore,
providing an improved cell microenvironment by relieving the
inflammatory response subsequent to SCI, along with cell
transplantation or endogenous NSCs activation, may be a promising
strategy for treating SCI. In conclusion, E-cadherin may maintain
NSCs in an undifferentiated state by promoting cell viability and
neurosphere formation. Grafted NSCs with highly expressed
E-cadherin facilitated motor functional recovery following SCI by
reducing the release of inflammatory cytokines and provide a
favorable microenvironment for neural regeneration.
Acknowledgements
Not applicable.
Funding
No funding was received.
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
DC and SL planned the study. DC wrote the manuscript
and performed the primary experiments. SH and JL were responsible
for the construction of SCI models, provided general coordination
of the study and revised the manuscript. All authors read and
approved the final manuscript.
Ethics approval and consent to
participate
All applicable international, national and
institutional guidelines for the care and use of animals were
followed. Ethical approval for animal (mice) use was provided by
the Ethics Committee of Shanghai Tenth People's Hospital, Tongji
University School of Medicine (Shanghai, China).
Patient consent to participate
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Gensel JC, Donnelly DJ and Popovich PG:
Spinal cord injury therapies in humans: An overview of current
clinical trials and their potential effects on intrinsic CNS
macrophages. Expert Opin Ther Targets. 15:505–518. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Tavares I: Human neural stem cell
transplantation in spinal cord injury models: How far from clinical
application? Stem Cell Res Ther. 4(61)2013.PubMed/NCBI
|
|
3
|
Lu Y and Wang MY: Neural stem cell grafts
for complete spinal cord injury. Neurosurgery. 71:N13–N15. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Pomeshchik Y, Puttonen KA, Kidin I,
Ruponen M, Lehtonen S, Malm T, Åkesson E, Hovatta O and Koistinaho
J: Transplanted human induced pluripotent stem cell-derived neural
progenitor cells do not promote functional recovery of
pharmacologically immunosuppressed mice with contusion spinal cord
injury. Cell Transplant. 24:1799–1812. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Salewski RP, Mitchell RA, Li L, Shen C,
Milekovskaia M, Nagy A and Fehlings MG: Transplantation of induced
pluripotent stem cell-derived neural stem cells mediate functional
recovery following thoracic spinal cord injury through
remyelination of axons. Stem Cells Transl Med. 4:743–754. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Sugai K, Nishimura S, Kato-Negishi M, Onoe
H, Iwanaga S, Toyama Y, Matsumoto M, Takeuchi S, Okano H and
Nakamura M: Neural stem/progenitor cell-laden microfibers promote
transplant survival in a mouse transected spinal cord injury model.
J Neurosci Res. 93:1826–1838. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Gonzalez R, Glaser J, Liu MT, Lane TE and
Keirstead HS: Reducing inflammation decreases secondary
degeneration and functional deficit after spinal cord injury. Exp
Neurol. 184:456–463. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Conti A, Cardali S, Genovese T, Di Paola R
and La Rosa G: Role of inflammation in the secondary injury
following experimental spinal cord trauma. J Neurosurg Sci.
47:89–94. 2003.PubMed/NCBI
|
|
9
|
Machova Urdzikova L, Karova K, Ruzicka J,
Kloudova A, Shannon C, Dubisova J, Murali R, Kubinova S, Sykova E,
Jhanwar-Uniyal M and Jendelova P: The anti-inflammatory compound
curcumin enhances locomotor and sensory recovery after spinal cord
injury in rats by immunomodulation. Int J Mol Sci. 17(pii):
E492015. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Han D, Wu C, Xiong Q, Zhou L and Tian Y:
Anti-inflammatory mechanism of bone marrow mesenchymal stem cell
transplantation in rat model of spinal cord injury. Cell Biochem
Biophys. 71:1341–1347. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Karavelioglu E, Gönül Y, Kokulu S, Hazman
Ö, Bozkurt F, Koçak A and Eser O: Anti-inflammatory and
antiapoptotic effect of interleukine-18 binding protein on the
spinal cord ischemia-reperfusion injury. Inflammation. 37:917–923.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Gao J, Grill RJ, Dunn TJ, Bedi S,
Labastida JA, Hetz RA, Xue H, Thonhoff JR, DeWitt DS, Prough DS, et
al: Human neural stem cell transplantation-mediated alteration of
microglial/macrophage phenotypes after traumatic brain injury. Cell
Transplant. 25:1863–1877. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Pellegatta S, Tunici P, Poliani PL,
Dolcetta D, Cajola L, Colombelli C, Ciusani E, Di Donato S and
Finocchiaro G: The therapeutic potential of neural stem/progenitor
cells in murine globoid cell leukodystrophy is conditioned by
macrophage/microglia activation. Neurobiol Dis. 21:314–323. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Klingener M, Chavali M, Singh J, McMillan
N, Coomes A, Dempsey PJ, Chen EI and Aguirre A: N-cadherin promotes
recruitment and migration of neural progenitor cells from the SVZ
neural stem cell niche into demyelinated lesions. J Neurosci.
34:9590–9606. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Karin M, Lawrence T and Nizet V: Innate
immunity gone awry: Linking microbial infections to chronic
inflammation and cancer. Cell. 124:823–835. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Nakamura E, Sugihara H, Bamba M and
Hattori T: Dynamic alteration of the E-cadherin/catenin complex
during cell differentiation and invasion of undifferentiated-type
gastric carcinomas. J Pathol. 205:349–358. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Ma ZZ, Fan L, Huang JL and Pan XJ: A novel
method to derive and expand mice neural stem cells efficiently
without neuro-sphere formation. Int J Clin Exp Med. 8:12834–12841.
2015.PubMed/NCBI
|
|
19
|
Reynolds BA, Tetzlaff W and Weiss S: A
multipotent EGF-responsive striatal embryonic progenitor cell
produces neurons and astrocytes. J Neurosci. 12:4565–4574. 1992.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Anderson DJ: Stem cells and pattern
formation in the nervous system: The possible versus the actual.
Neuron. 30:19–35. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Bacigaluppi M, Pluchino S,
Peruzzotti-Jametti L, Kilic E, Kilic U, Salani G, Brambilla E, West
MJ, Comi G, Martino G and Hermann DM: Delayed post-ischaemic
neuroprotection following systemic neural stem cell transplantation
involves multiple mechanisms. Brain. 132:2239–2251. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Robert AA, Zamzami M, Sam AE, Al Jadid M
and Al Mubarak S: The efficacy of antioxidants in functional
recovery of spinal cord injured rats: An experimental study. Neurol
Sci. 33:785–791. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Ravikumar R, Narayanan S, Baskar S,
Senthil Nagarajan R and Abraham S: Autologous stem cell injection
for spinal cord injury-a clinical study from India. J Stem Cells
Regen Med. 3:24–25. 2007.PubMed/NCBI
|
|
24
|
Seebach C, Henrich D, Meier S, Nau C,
Bonig H and Marzi I: Safety and feasibility of cell-based therapy
of autologous bone marrow-derived mononuclear cells in
plate-stabilized proximal humeral fractures in humans. J Transl
Med. 14:3142016. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Kumar AA, Kumar SR, Narayanan R, Arul K
and Baskaran M: Autologous bone marrow derived mononuclear cell
therapy for spinal cord injury: A phase I/II clinical safety and
primary efficacy data. Exp Clin Transplant. 7:241–248.
2009.PubMed/NCBI
|
|
26
|
Einstein O, Karussis D, Grigoriadis N,
Mizrachi-Kol R, Reinhartz E, Abramsky O and Ben-Hur T:
Intraventricular transplantation of neural precursor cell spheres
attenuates acute experimental allergic encephalomyelitis. Mol Cell
Neurosci. 24:1074–1082. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Pluchino S, Zanotti L, Rossi B, Brambilla
E, Ottoboni L, Salani G, Martinello M, Cattalini A, Bergami A,
Furlan R, et al: Neurosphere-derived multipotent precursors promote
neuroprotection by an immunomodulatory mechanism. Nature.
436:266–271. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Aharonowiz M, Einstein O, Fainstein N,
Lassmann H, Reubinoff B and Ben-Hur T: Neuroprotective effect of
transplanted human embryonic stem cell-derived neural precursors in
an animal model of multiple sclerosis. PLoS One. 3:e31452008.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Cheng Z, Zhu W, Cao K, Wu F, Li J, Wang G,
Li H, Lu M, Ren Y and He X: Anti-inflammatory mechanism of neural
stem cell transplantation in spinal cord injury. Int J Mol Sci.
17(pii): E13802016. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Mautes AE, Weinzierl MR, Donovan F and
Noble LJ: Vascular events after spinal cord injury: Contribution to
secondary pathogenesis. Phys Ther. 80:673–687. 2000.PubMed/NCBI
|
|
31
|
Fleming JC, Norenberg MD, Ramsay DA,
Dekaban GA, Marcillo AE, Saenz AD, Pasquale-Styles M, Dietrich WD
and Weaver LC: The cellular inflammatory response in human spinal
cords after injury. Brain. 129:3249–3269. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Hausmann ON: Post-traumatic inflammation
following spinal cord injury. Spinal Cord. 41:369–378. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Liu Y, Ye H, Satkunendrarajah K, Yao GS,
Bayon Y and Fehlings MG: A self-assembling peptide reduces glial
scarring, attenuates post-traumatic inflammation and promotes
neurological recovery following spinal cord injury. Acta Biomater.
9:8075–8088. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
van Roy F: Beyond E-cadherin: Roles of
other cadherin superfamily members in cancer. Nat Rev Cancer.
14:121–134. 2014. View
Article : Google Scholar : PubMed/NCBI
|
|
35
|
Onder TT, Gupta PB, Mani SA, Yang J,
Lander ES and Weinberg RA: Loss of E-cadherin promotes metastasis
via multiple downstream transcriptional pathways. Cancer Res.
68:3645–3654. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Vleminckx K, Vakaet L Jr, Mareel M, Fiers
W and van Roy F: Genetic manipulation of E-cadherin expression by
epithelial tumor cells reveals an invasion suppressor role. Cell.
66:107–119. 1991. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Christofori G and Semb H: The role of the
cell-adhesion molecule E-cadherin as a tumour-suppressor gene.
Trends Biochem Sci. 24:73–76. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Frixen UH, Behrens J, Sachs M, Eberle G,
Voss B, Warda A, Löchner D and Birchmeier W: E-cadherin-mediated
cell-cell adhesion prevents invasiveness of human carcinoma cells.
J Cell Biol. 113:173–185. 1991. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Eastham AM, Spencer H, Soncin F, Ritson S,
Merry CL, Stern PL and Ward CM: Epithelial-mesenchymal transition
events during human embryonic stem cell differentiation. Cancer
Res. 67:11254–11262. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
D'Amour KA, Agulnick AD, Eliazer S, Kelly
OG, Kroon E and Baetge EE: Efficient differentiation of human
embryonic stem cells to definitive endoderm. Nat Biotechnol.
23:1534–1541. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Xu Y, Zhu X, Hahm HS, Wei W, Hao E, Hayek
A and Ding S: Revealing a core signaling regulatory mechanism for
pluripotent stem cell survival and self-renewal by small molecules.
Proc Natl Acad Sci USA. 107:8129–8134. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Li L, Wang BH, Wang S, Moalim-Nour L,
Mohib K, Lohnes D and Wang L: Individual cell movement, asymmetric
colony expansion, rho-associated kinase, and E-cadherin impact the
clonogenicity of human embryonic stem cells. Biophys J.
98:2442–2451. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Hao J, Li TG, Qi X, Zhao DF and Zhao GQ:
WNT/beta-catenin pathway up-regulates Stat3 and converges on LIF to
prevent differentiation of mouse embryonic stem cells. Dev Biol.
290:81–91. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Hawkins K, Mohamet L, Ritson S, Merry CL
and Ward CM: E-cadherin and, in its absence, N-cadherin promotes
Nanog expression in mouse embryonic stem cells via STAT3
phosphorylation. Stem Cells. 30:1842–1851. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Maeda K, Takemura M, Umemori M and
Adachi-Yamada T: E-cadherin prolongs the moment for interaction
between intestinal stem cell and its progenitor cell to ensure
Notch signaling in adult Drosophila midgut. Genes Cells.
13:1219–1227. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Jin Z, Kirilly D, Weng C, Kawase E, Song
X, Smith S, Schwartz J and Xie T: Differentiation-defective stem
cells outcompete normal stem cells for niche occupancy in the
Drosophila ovary. Cell Stem Cell. 2:39–49. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
del Valle I, Rudloff S, Carles A, Li Y,
Liszewska E, Vogt R and Kemler R: E-cadherin is required for the
proper activation of the Lifr/Gp130 signaling pathway in mouse
embryonic stem cells. Development. 140:1684–1692. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Karpowicz P, Willaime-Morawek S, Balenci
L, DeVeale B, Inoue T and van der Kooy D: E-Cadherin regulates
neural stem cell self-renewal. J Neurosci. 29:3885–3896. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Shie JH and Kuo HC: Higher levels of cell
apoptosis and abnormal E-cadherin expression in the urothelium are
associated with inflammation in patients with interstitial
cystitis/painful bladder syndrome. BJU Int. 108:E136–E141. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Mehta S, Nijhuis A, Kumagai T, Lindsay J
and Silver A: Defects in the adherens junction complex
(E-cadherin/β-catenin) in inflammatory bowel disease. Cell Tissue
Res. 360:749–760. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Zbar AP, Simopoulos C and Karayiannakis
AJ: Cadherins: An integral role in inflammatory bowel disease and
mucosal restitution. J Gastroenterol. 39:413–421. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Turner JR: Molecular basis of epithelial
barrier regulation: From basic mechanisms to clinical application.
Am J Pathol. 169:1901–1909. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Takeichi M: Cadherin cell adhesion
receptors as a morphogenetic regulator. Science. 251:1451–1455.
1991. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Tsai HT, Lee TH, Yang SF, Lin LY, Tee YT
and Wang PH: Markedly elevated soluble E-cadherin in plasma of
patient with pelvic inflammatory disease. Fertil Steril.
99:490–495. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Reiss K, Ludwig A and Saftig P: Breaking
up the tie: Disintegrin-like metalloproteinases as regulators of
cell migration in inflammation and invasion. Pharmacol Ther.
111:985–1006. 2006. View Article : Google Scholar : PubMed/NCBI
|