Introduction
Lower back pain (LBP) is one of the most common
complaints in the orthopedic outpatient department, with total
costs incurred by complications associated with LBP amounting to
~$100 billion/year in the US (1,2).
Intervertebral disc (IVD) degeneration (IDD) is an important cause
of LBP (3,4). The mechanism of IDD onset involves a
complex biochemical cascade. An important feature of IDD is the
loss of proteoglycan content in nucleus pulposus cells, leading to
a change in biomechanics (5).
However, in spite of accumulating research data, the relationship
between mechanical load and IDD onset remains unclear.
The IVD serves an important role in the biomechanics
of the spine as it experiences varying degrees of mechanical stress
during daily tasks (6). Previous
studies have found that mechanical load is an important factor in
the pathogenesis of IDD (7,8). Indeed, excessive mechanical pressure
has been demonstrated to promote apoptosis, upregulate the
expression of extracellular matrix (ECM)-degrading enzymes whilst
downregulating those associated with ECM synthesis in nucleus
pulposus cells, ultimately leading to IDD (7,8).
However, unlike excessive mechanical loading, moderate amounts of
mechanical pressure may exert beneficial effects on nucleus
pulposus cells (9). Therefore, the
relationship between mechanical loading and associated
intracellular mechanism in nucleus pulposus cells requires further
study.
NF-κB is an important pathway that regulates gene
expression. A number of studies have previously revealed that
mechanical forces exerted physiological influences on a variety of
different cell types by NF-κB activation, including chondrocytes,
endothelial cells and cardiomyocytes (10–12). In
addition, the NF-κB pathway serves an important role in the
regulation of the inflammatory response and has been reported to be
closely associated with IDD (13).
NF-κB activation leads to the upregulation of inflammatory
cytokines and ECM-degrading enzymes, resulting in ECM degradation
and IDD (13,14). On the basis of these previous
findings, it was hypothesized that NF-κB activation may be
associated with mechanical loading in nucleus pulposus cells.
In the present study, a human nucleus pulposus cell
line model was used to examine the physiological consequences of
differential patterns of mechanical loading. Data from the present
study may clarify the potential mechanism and relationship between
mechanical pressure and IDD.
Materials and methods
Cell culture
Primary human nucleus pulposus (NP) cells (cat. no.
4800; ScienCell Research Laboratories, Inc.) were cultured using
high-glucose DMEM (Gibco; Thermo Fisher Scientific, Inc.)
supplemented with 10% fetal bovine serum (Gibco; Thermo Fisher
Scientific, Inc.), 100 U/ml penicillin (HyClone, Thermo Scientific,
USA) and 100 µg/ml streptomycin (HyClone; GE Healthcare Life
Sciences) up to passage 2–3. NP cells were incubated at 37°C with
5% CO2.
Mechanical loading
Mechanical loading was achieved using the FX-5000TM
Tension or Compression system according to default parameters
(Flexcell International Corporation). Cells were cultured at 37°C
between a piston and stationary platen, positive air pressure was
exhibited to achieve compression loading. NP cells
(2×105/ml) were seeded into six-well BioFlex plates
(Flexcell International Corporation) and subsequently designated as
the unloaded control, load or overload groups. The NP cells in the
load group were treated with two or five cycles of mechanical
loading, with each cycle consisting of 0.8 MPa 0.5 Hz for 4 h
followed by 0.1 MPa 0.5 Hz for 20 h. Cells in the overload group
were subjected to two or five cycles of mechanical pressure, and
each cycle consisted of 0.8 MPa 0.5 Hz for 20 h followed by 0.1 MPa
0.5 Hz for 4 h. Cells were loaded for 2 or 5 days depending on the
number of mechanical pressure cycles. In the control group, cells
were cultured without mechanical loading.
Cell Counting kit-8 (CCK-8) assay
Following 24 or 48 h of mechanical loading, cell
viability was measured using the CCK-8 assay kit (Dojindo Molecular
Technologies, Inc.) according to manufacturer's protocol. CCK-8
solution diluted in DMEM was added to the six-well BioFlex plates
prior to incubation for 2 h at 37°C. The solution was then
transferred to 96-well plates for optical density (OD) measurement
at 450 nm. DMEM that was not conditioned by NP cells was used as
blank control. Viability was calculated using the following
formula: Viability = [OD(24 or 48
h)-OD(blank)]/[OD(0 h)-OD(blank)].
Cell apoptosis assay
An Annexin V APC-PI Apoptosis kit was used for cell
apoptosis assay (550474, BD Biosciences). NP cells were collected
and suspended in 500 µl binding buffer after mechanical loading. A
total of 5 µl Annexin-V-fluorescein isothiocyanate and 5 µl
propidium iodide were then added into the buffer before incubation
for 15 min in the dark. Flow cytometry analysis was performed to
detect apoptotic cells. Flowjo 10.0 software (FlowJo, LLC) was used
for data analysis.
Western blot analysis
NP cells were either treated with or without the
NF-κB pathway inhibitor BAY11-7082 (20 µM; Selleck Chemicals) 48 h
prior to mechanical loading. After treatment, the cells were washed
twice using ice-cold PBS and lysed with RIPA buffer (50 mM Tris (pH
7.4); 150 mM NaCl; 1% NP-40; 0.5% sodium deoxycholate) supplemented
with different inhibitors (1 mM Na3VO4, 10 mM NaF or 1%
phosphatase/protease inhibitor cocktail). Protein concentration was
quantified using a bicinchoninic acid protein assay kit (Thermo
Fisher Scientific, Inc.). The samples (20 µg per lane) were then
separated using 10% SDS-PAGE and transferred onto PVDF membranes.
The membranes were blocked in 5% fat-free milk diluted in TBS
supplemented with 5% w/v BSA (Gibco; Thermo Fisher Scientific,
Inc.) and 0.1% Tween-20 (TBS-T) for 1 h at room temperature,
followed by incubation with primary antibodies against NF-κB
inhibitor-α (IκBα; cat. no. 4814), phosphorylated-IκBα (p-IκBα;
cat. no. 2859), p65 (cat. no. 8284) or p-p65 (cat. no. 3039;
1:1,000; Cell Signaling Technology, Inc.) at 4°C overnight. The
membranes were then washed with TBS-T three times, before
incubation with appropriate secondary antibodies conjugated with
IRDye® 800CW at room temperature for 1 h (cat. no.
925-32211, 1:20,000, LI-COR corporate). Immunoreactive bands were
visualized using Odyssey infrared imaging system (LI-COR
Biosciences). The β-actin antibody (1:2,000; cat. no. 3700; Cell
Signaling Technology, Inc.) at 4°C overnight was used as loading
control. Positive bands were quantified using Quantity One 1-D
version 4.6.9 (Bio-Rad Laboratories, Inc.) and normalized to
β-actin.
Gene expression assay
Following 48 h mechanical pressure, total RNA was
extracted from cultured cells using TRIzol® reagent
(Thermo Fisher Scientific, Inc.) according to the manufacturer's
protocol. cDNA was then generated from 1 µg purified total RNA
using PrimeScriptTM RT Master Mix (Takara Bio, Inc.) according to
manufacturer's protocol. For quantitative PCR, thermocycling was
performed by denaturation at 95°C for 5 min, followed by 40 cycles
of 95°C for 5 sec and 60°C for 30 sec. For semi-quantitative PCR,
thermocycling was performed using 40 cycles of 95°C for 30 sec,
72°C for 45 sec and 74°C for 60 sec. Gene expression was determined
using semi-quantitative and quantitative PCR (qPCR).
Semi-quantitative PCR was performed using GoTaq® Green
Master Mix (Promega Corporation) according to manufacturer's
protocol. PCR products were verified using 1.5% agarose gel
electrophoresis. Positive immunoreactive bands were observed using
a GelDoc XR System (Bio-Rad Laboratories, Inc.) and quantified
using Quantity One 1-D version 4.6.9 software (Bio-Rad
Laboratories, Inc.). qPCR was performed using a SYBR®
Premix Ex TaqTM kit (Takara Bio, Inc.) with the Applied Biosystems
Prism® 7500 Fast Real-Time PCR system (Applied
Biosystems; Thermo Fisher Scientific, Inc.) according to
manufacturer's protocol. The primer sequences for metallopeptidase
(MMP)-3 and −13, A disintegrin and metalloproteinase with
thrombospondin motifs (ADAMTS)-4 and −5, aggrecan, collagen-II,
Bcl-2, Bax and β-actin were designed and selected using BLAST, and
are listed in Table I. Gene
expression was measured using the 2−ΔΔCq method
(15), with β-actin used as internal
control.
 | Table I.Sequences of primers used in
polymerase chain reaction. |
Table I.
Sequences of primers used in
polymerase chain reaction.
| Gene | Primer sequence
(5′-3′) |
|---|
| Bcl-2 | Forward
GAACTGGGGGAGGATTGTGG |
|
| Reverse
CCGTACAGTTCCACAAAGGC |
| Bax | Forward
CCAGAGGCGGGGTTTCAT |
|
| Reverse
GGAAAAAGACCTCTCGGGGG |
| MMP-3 | Forward
TTTGGCCGTCTCTTCCATCC |
|
| Reverse
GCATCGATCTTCTGGACGGT |
| MMP-13 | Forward
ACCATCCTGTGACTCTTGCG |
|
| Reverse
TTCACCCACATCAGGCACTC |
| ADAMTS-4 | Forward
ACCGATTACCAGCCTTTGGG |
|
| Reverse
CCGACTCCGGATCTCCATTG |
| ADAMTS-5 | Forward
CCGAACGAGTTTACGGGGAT |
|
| Reverse
TGTGCGTCGCCTAGAACTAC |
| Aggrecan | Forward
CAGATGGCACCCTCCGATAC |
|
| Reverse
GACACACCTCGGAAGCAGAA |
| Collagen-II | Forward
GGCCAGGATGCCCGAAAATT |
|
| Reverse
ACCCCTCTCTCCCTTGTCAC |
| β-actin | Forward
AACCTTCTTGCAGCTCCTCCG |
|
| Reverse
CCATACCCACCATCACACCCT |
ELISA
NP cells were subjected to mechanical loading for 2
and 5 days respectively. The levels of C-telopeptide of type II
collagen (CTX–II) in the culture medium of NP cells were
subsequently measured using a commercially available ELISA kit
(cat. no. MOCT00; R&D Systems, Inc.) according to
manufacturer's protocol.
Immunocytochemistry
NP cells were seeded onto coverslips and subjected
to mechanical loading for 2 or 5 days. For the overload +
BAY11-2082 group, BAY11-7082 was added to the culture medium at the
beginning of mechanical loading for 2 or 5 days at 37°C.
Immunocytochemistry was then performed on days 2 and 5, after
mechanical loading, respectively. The cells were fixed using 4%
paraformaldehyde for 10 min at room temperature prior to incubation
with 0.2% Triton X-100 for 15 min at room temperature. Following
blocking at room temperature for 2 h with 5% fat-free milk diluted
in TBS and supplemented with 5% w/v BSA, cells were incubated with
anti-Collagen II antibody (cat. no. ab24118; 1:500 dilution;
Abcam,) overnight at 4°C and treated with horseradish
peroxidase-conjugated secondary antibody (1:1,000; cat. no. M00172;
Boster Biological Technology) for 1 h at room temperature, prior to
immersion into DAB at room temperature for 5 min. All
immunocytochemistry images were captured using a Nikon ECLIPSE 80i
microscope (Nikon Corporation) with NIS-Elements D software
(version 4.50; Nikon Corporation). For all images captured,
integral optical density analysis was calculated using Image Pro
Plus software 6.0 (Media Cybernetics, Inc.).
Statistical analysis
All experiments were repeated three times. All data
are presented as the mean ± standard deviation. Statistical
analysis was performed using one-way ANOVA, followed by Duncan's
post-hoc test using SPSS 19.0 (IBM Corp.). P<0.05 was considered
to indicate a statistically significant difference.
Results
Effects of mechanical loading on cell
viability and apoptosis
To study the effects of mechanical loading on NP
cells, CCK-8 assay was used to assess cell viability. Cell
viability in the load group was found to be enhanced at 48 h
compared with the control group, whilst that in the overload group
was significantly reduced at 24 and 48 h (Fig. 1A). Compared with the control group,
although moderate loading did not significantly alter Bax and Bcl-2
expression, overloading significantly increased Bax expression
whilst downregulating Bcl-2 expression (Fig. 1B and C). According to flow cytometry
analysis, the apoptotic rate was significantly increased in the
overload group compared with the load group, but no significant
differences were observed between the load and control groups
(Fig. 1D and E).
ECM synthesis is affected by different
patterns of mechanical loading
To measure ECM synthesis related gene and protein
expression in NP cells, reverse transcription-qPCR (RT-qPCR) and
immunocytochemistry were used to evaluate mRNA and protein levels
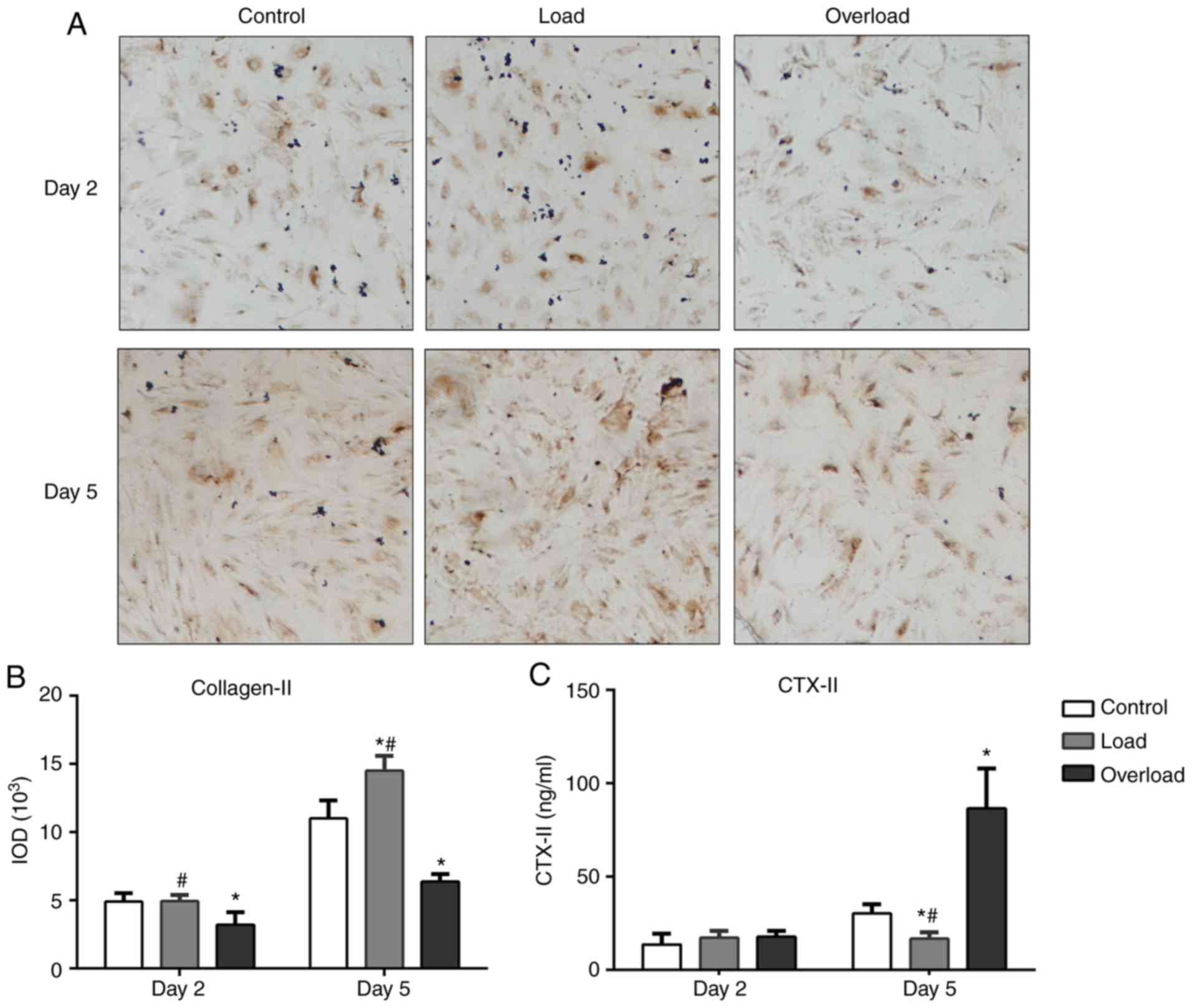
of Collagen II and aggrecan, respectively. Collagen-II synthesis
was significantly increased in the load group compared with the
control group, the production of Collagen-II increased in the load
group compared with the control group (Fig. 2); however, collagen-II mRNA and
protein expression levels were significantly reduced in the
overload group compared with the load group (Figs. 2, 3A and
B). In terms of aggrecan, gene expression was reduced in the
overload group compared with the control group; while moderate
loading increased NP cell aggrecan expression (Fig. 2).
Expression of ECM-degrading enzymes
and collagen-II degradation are stimulated by excessive mechanical
loading
The effects of mechanical loading on the expression
of the main ECM-degrading enzymes in NP cells and collagen-II
degradation were examined using RT-qPCR analysis and ELISA. The
expression of ECM-degrading enzymes MMP-3, MMP-13, ADAMTS-5 and
ADAMTS-4 in the overload group was significantly increased compared
with the control group (Fig. 2);
whilst no significant differences were observed between load and
control groups. According to the ELISA data, CTX–II levels in the
culture medium collected from the overload group were significantly
increased at day 5 compared with that from the control group,
suggesting accelerated collagen-II degradation in the overload
group (Fig. 3C). Additionally,
CTX–II level was reduced in load group at day 5 when compared with
the control group (Fig. 3C).
Excessive mechanical loading promotes
ECM degradation by activating NF-κB in NP cells
To evaluate the intracellular mechanism of excessive
mechanical loading in NP cells, the activation of the NF-κB pathway
was measured using western blot analysis following BAY11-7082
treatment. In the overload group, the degrees of p65 and IκBα
phosphorylation were significantly elevated on day 5 compared with
the load group and the control group. Therefore, excessive
mechanical loading significantly increased NF-κB pathway activation
in NP cells, which was reversed by BAY11-7082 treatment (Fig. 4A and B). Collagen-II degradation in
the overload group was partially reversed in the presence of
BAY11-7082 on day 5 (Fig. 4C and D),
suggesting a role for the NF-κB signaling pathway in collagen-II
degradation induced by mechanical overload.
Discussion
The L4/5 and L5/S1 lumbar segments are the most
susceptible to degeneration due to the heavy mechanical pressure
endured (16). However, the specific
value of loading of spine is difficult to quantify. Wilke et
al (17) directly measured the
pressure in the L4/5 disc of a healthy volunteer, which was found
to be 0.1 MPa during sleep and 0.3–2.3 MPa during activities
performed in daily life (sitting, 0.3–0.83 MPa; standing, 0.5–1.1
MPa; lifting weight, 1.1–2.3 MPa). Based on the findings of this
study, the current study used 0.1 and 0.8 MPa pressure to mimic
rest and working activities, respectively.
Sleep deprivation is considered to be a possible
risk factor for the progressive deterioration of LBP. Appropriate
amounts of sleep were previously reported to mitigate chronic pain
(18). Therefore, in the present
study, a 4-h resting period was set for the overload group to mimic
patients with sleep problems and study the relationship between
rest time and IDD. The results suggested that poor sleep may be an
underlying factor for IDD, whilst ample rest exhibited beneficial
effects for IVD.
Static or dynamic compression techniques have been
frequently applied to research disc degeneration (19,20). It
was previously reported that dynamic compression was more
physiologically relevant compared with static compression in
vivo (17). In the current
study, dynamic compression was used to gain a better result.
However, the pressure of 2.0 MPa applied in the study was
relatively high and the in vitro loading time of 45 min was
relatively short (19). It has
previously been demonstrated that in rabbit chondrocytes subjected
to stress varying from 0 to 200 kPa, and at 0.1 Hz, yielded the
strongest tissue-engineered cartilage (21). However, the most appropriate pressure
for studying nucleus pulposus cells remains poorly understood. In
the present study, an in vitro loading model was
constructed. It was found that a short loading time followed by a
long period of rest exerted beneficial effects on nucleus pulposus
cells, while a long loading time followed by a short period of rest
led to increased apoptosis and accelerated collagen II
degradation.
Nucleus pulposus cell apoptosis serves an important
role in IDD, as it leads to ECM degradation and the fibrosis of
nucleus pulposus (22,23). Aberrant mechanical stress due to
stretching has been found to induce chondrocyte and nucleus
pulposus cell apoptosis (23,24). The
present study demonstrated that prolonged periods of mechanical
pressure inhibited nucleus pulposus cell viability whilst promoting
apoptosis by disrupting the balance between Bcl-2 and Bax
expression. However, the detailed mechanism underlying this
observation requires further study in the future.
MMPs and ADAMTSs are the major catabolic enzymes
that have been studied extensively in the pathogenesis of IVD
degeneration (25,26). Among these families of enzymes,
MMP-3, MMP-13, ADAMTS-4 and ADAMTS-5 were classified as the major
enzymes related to IDD (27,28). In particular, an in vitro study
previously reported that the expression of ADAMTSs was
significantly elevated in human nucleus pulposus cells after the
application of compressive load (29). This study provided preliminary data
on the underlying mechanism linking compressive load to the
development of IDD. In the present study, it was found that the
expression levels of MMP-3, MMP-13, ADAMTS-4 and ADAMTS-5 were
upregulated after prolonged periods of loading, whereas the
expression of collagen-II and aggrecan were suppressed. These
results suggest that long periods of mechanical pressure may reduce
ECM content by limiting synthesis and promoting degradation.
The NF-κB signaling pathway serves a crucial role in
IDD, as it had been demonstrated to be activated in response to
mechanical stress (30), leading to
the production of ADAMTSs and MMPs (31). A previous study reported that the
expression of ADAMTS-4, ADAMTS-5 and MMP-13 increased following
mechanical stress by activating NF-κB in human chondrocytes, an
effect that could be reversed by the inhibition of NF-κB signaling
(11). The present study yielded
similar observations as excessive mechanical loading induced NF-κB
activation, whilst the inhibition of NF-κB reduced ECM degradation.
However, it was noted that IκBα was also mildly phosphorylated in
the load group. Indeed, the NF-κB pathway has been reported to be
associated with a large number of cellular physiological events
that do not involve ECM degradation and mechanical loading
(30). Therefore, results from the
present study suggest that mechanical loading may influence nucleus
pulposus cell ECM content in a NF-κB-independent manner, and
differential magnitudes of NF-κB activation may produce different
outcomes.
However, the present study had certain limitations.
Firstly, cells were cultured in a monolayer which is inconsistent
with in vivo conditions. Secondly, the mechanism of
mechanical loading on nucleus pulposus cells was not thoroughly
studied. Further studies should focus on using three-dimensional
cell culture models and should including additional studies into
the mechanism behind this.
In conclusion, the results from the present study
demonstrate that excessive mechanical loading inhibits human
nucleus pulposus cell viability and promotes ECM degradation by
NF-κB activation, whilst appropriate mechanical loading exhibited
beneficial effects for nucleus pulposus cells.
Acknowledgements
Not applicable.
Funding
No funding was received.
Availability of data and materials
The analyzed datasets generated during the present
study are available from the corresponding author on reasonable
request.
Authors' contributions
YW conceived and designed the experiments, performed
the experiments, analyzed the data, contributed
reagents/materials/analytical tools, wrote the manuscript, prepared
figures and tables and reviewed drafts of the paper. KZ and CX
performed the experiments, analyzed the data, contributed
reagents/materials/analysis tools, and prepared figures and tables.
NL and PR performed the experiments. HWP and YW designed the
experiments and reviewed the paper draft.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Hart LG, Deyo RA and Cherkin DC: Physician
office visits for low back pain. Frequency, clinical evaluation,
and treatment patterns from a U.S. national survey. Spine (Phila Pa
1976). 20:11–19. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Katz JN: Lumbar disc disorders and
low-back pain: Socioeconomic factors and consequences. J Bone Joint
Surg Am. 88 (Suppl 2):S21–S24. 2006. View Article : Google Scholar
|
|
3
|
Kuslich SD, Ulstrom CL and Michael CJ: The
tissue origin of low back pain and sciatica: A report of pain
response to tissue stimulation during operations on the lumbar
spine using local anesthesia. Orthop Clin North Am. 22:181–187.
1991.PubMed/NCBI
|
|
4
|
Schwarzer AC, Aprill CN, Derby R, Fortin
J, Kine G and Bogduk N: The relative contributions of the disc and
zygapophyseal joint in chronic low back pain. Spine (Phila Pa
1976). 19:801–806. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Kepler CK, Ponnappan RK, Tannoury CA,
Risbud MV and Anderson DG: The molecular basis of intervertebral
disc degeneration. Spine J. 13:318–330. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Setton LA and Chen J: Mechanobiology of
the intervertebral disc and relevance to disc degeneration. J Bone
Joint Surg Am. 88 (Suppl 2):S52–S57. 2006. View Article : Google Scholar
|
|
7
|
Gawri R, Rosenzweig DH, Krock E, Ouellet
JA, Stone LS, Quinn TM and Haglund L: High mechanical strain of
primary intervertebral disc cells promotes secretion of
inflammatory factors associated with disc degeneration and pain.
Arthritis Res Ther. 16:R212014. View
Article : Google Scholar : PubMed/NCBI
|
|
8
|
Wang C, Gonzales S, Levene H, Gu W and
Huang CY: Energy metabolism of intervertebral disc under mechanical
loading. J Orthop Res. 31:1733–1738. 2013.PubMed/NCBI
|
|
9
|
Chowdhury TT, Arghandawi S, Brand J,
Akanji OO, Bader DL, Salter DM and Lee DA: Dynamic compression
counteracts IL-1beta induced inducible nitric oxide synthase and
cyclo-oxygenase-2 expression in chondrocyte/agarose constructs.
Arthritis Res Ther. 10:R352008. View
Article : Google Scholar : PubMed/NCBI
|
|
10
|
Zhao H, Hiroi T, Hansen BS and Rade JJ:
Cyclic stretch induces cyclooxygenase-2 gene expression in vascular
endothelial cells via activation of nuclear factor kappa-beta.
Biochem Biophys Res Commun. 389:599–601. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Ozawa M, Nishida K, Yoshida A, Saito T,
Harada R, Machida T and Ozaki T: Hyaluronan suppresses mechanical
stress-induced expression of catabolic enzymes by human
chondrocytes via inhibition of IL-1β production and subsequent
NF-κB activation. Inflamm Res. 64:243–252. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Leychenko A, Konorev E, Jijiwa M and
Matter ML: Stretch-induced hypertrophy activates NFkB-mediated VEGF
secretion in adult cardiomyocytes. PLoS One. 6:e290552011.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Li Y, Li K, Mao L, Han X, Zhang K, Zhao C
and Zhao J: Cordycepin inhibits LPS-induced inflammatory and matrix
degradation in the intervertebral disc. PeerJ. 4:e19922016.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Li Y, Li K, Hu Y, Xu B and Zhao J:
Piperine mediates LPS induced inflammatory and catabolic effects in
rat intervertebral disc. Int J Clin Exp Pathol. 8:6203–6213.
2015.PubMed/NCBI
|
|
15
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Lou C, Chen H, Mei L, Yu W, Zhu K, Liu F,
Chen Z, Xiang G, Chen M, Weng Q and He D: Association between
menopause and lumbar disc degeneration: An MRI study of 1,566 women
and 1,382 men. Menopause. 24:1136–1144. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Wilke HJ, Neef P, Caimi M, Hoogland T and
Claes LE: New in vivo measurements of pressures in the
intervertebral disc in daily life. Spine (Phila Pa 1976).
24:755–762. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Nitter AK, Pripp AH and Forseth KØ: Are
sleep problems and non-specific health complaints risk factors for
chronic pain? A prospective population-based study with 17 year
follow-up. Scand J Pain. 3:210–217. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
van der Veen AJ, van Dieën JH, Nadort A,
Stam B and Smit TH: Intervertebral disc recovery after dynamic or
static loading in vitro: Is there a role for the endplate? J
Biomech. 40:2230–2235. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Wang DL, Jiang SD and Dai LY: Biologic
response of the intervertebral disc to static and dynamic
compression in vitro. Spine (Phila Pa 1976). 32:2521–2528. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Anderson DE and Johnstone B: Dynamic
mechanical compression of chondrocytes for tissue engineering: A
critical review. Front Bioeng Biotechnol. 5:762017. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Chen S, Lv X, Hu B, Zhao L, Li S, Li Z,
Qing X, Liu H, Xu J and Shao Z: Critical contribution of RIPK1
mediated mitochondrial dysfunction and oxidative stress to
compression-induced rat nucleus pulposus cells necroptosis and
apoptosis. Apoptosis. 23:299–313. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Yang Q, Zhou Y, Wang J, Fu W and Li X:
Study on the mechanism of excessive apoptosis of nucleus pulposus
cells induced by shRNA-Piezo1 under abnormal mechanical stretch
stress. J Cell Biochem. 120:3989–3997. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Wang W, Qing X, Wang B, Ma K, Wei Y and
Shao Z: Tauroursodeoxycholic acid protects nucleus pulposus cells
from compression-induced apoptosis and necroptosis via inhibiting
endoplasmic reticulum stress. Evid Based Complement Alternat Med.
2018:67194602018.PubMed/NCBI
|
|
25
|
Pockert AJ, Richardson SM, Le Maitre CL,
Lyon M, Deakin JA, Buttle DJ, Freemont AJ and Hoyland JA: Modified
expression of the ADAMTS enzymes and tissue inhibitor of
metalloproteinases 3 during human intervertebral disc degeneration.
Arthritis Rheum. 60:482–491. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Li Y, Li K, Han X, Mao C, Zhang K, Zhao T
and Zhao J: The imbalance between TIMP3 and matrix-degrading
enzymes plays an important role in intervertebral disc
degeneration. Biochem Biophys Res Commun. 469:507–514. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Bachmeier BE, Nerlich A, Mittermaier N,
Weiler C, Lumenta C, Wuertz K and Boos N: Matrix metalloproteinase
expression levels suggest distinct enzyme roles during lumbar disc
herniation and degeneration. Eur Spine J. 18:1573–1586. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Patel KP, Sandy JD, Akeda K, Miyamoto K,
Chujo T, An HS and Masuda K: Aggrecanases and aggrecanase-generated
fragments in the human intervertebral disc at early and advanced
stages of disc degeneration. Spine (Phila Pa 1976). 32:2596–2603.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Huang M, Wang HQ, Zhang Q, Yan XD, Hao M
and Luo ZJ: Alterations of ADAMTSs and TIMP-3 in human nucleus
pulposus cells subjected to compressive load = Implications in the
pathogenesis of human intervertebral disc degeneration. J Orthop
Res. 30:267–273. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Rigoglou S and Papavassiliou AG: The NF-κB
signalling pathway in osteoarthritis. Int J Biochem Cell Biol.
45:2580–2584. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Kobayashi H, Hirata M, Saito T, Itoh S,
Chung UI and Kawaguchi H: Transcriptional induction of ADAMTS5
protein by nuclear factor-κB (NF-κB) family member RelA/p65 in
chondrocytes during osteoarthritis development. J Biol Chem.
288:28620–28629. 2013. View Article : Google Scholar : PubMed/NCBI
|