Introduction
Cervical cancer is one of the most common
malignancies in females worldwide, with 500,000 incident cases and
300,000 mortalities each year; it is commonly caused by infection
with high-risk strains of human papillomavirus (1–3). Despite
improvements in surgical resection combined with radiotherapy and
chemotherapy, the prognosis of patients with advanced cervical
cancer remains poor, primarily due to metastasis and drug tolerance
(2,4,5).
Therefore, it is critically important to explore the molecular
mechanism underlying cervical cancer progression, which may assist
in developing novel and efficient biomarkers and therapeutic
targets.
Long non-coding RNAs (lncRNAs) are a group of
single-strand non-coding RNAs measuring >200 nucleotides in
length, which comprise ~80% of non-coding RNAs (6,7). In
previous years, accumulating evidence has revealed that lncRNAs
serve key roles in a variety of cellular biological processes,
including cell proliferation, apoptosis, differentiation,
viability, autophagy, migration and angiogenesis (8–10). In
addition, a large number of lncRNAs have been demonstrated to serve
promotive or suppressive roles in various types of human cancer,
including cervical cancer, through interactions with the downstream
microRNAs (miRNAs) or proteins, and certain lncRNAs are associated
with cancer recurrence and poor prognosis (11,12). For
example, lncRNA X inactive specific transcript accelerates cervical
cancer progression by upregulating FUS RNA binding protein by
competitively binding with miR-200a (13). The lncRNA Taurine up-regulated 1
promotes cervical cancer progression by regulating the
miR-138-5p-sirtuin 1 axis (14).
Recently, Li et al (15)
suggested that lncRNA NCK1-antisense 1 (AS1) promoted cervical
cancer cell proliferation by inducing cell cycle progression by
regulating the expression of miR-6857 and cyclin dependent kinase
1. However, the underlying molecular mechanism of NCK1-AS1 in
cervical cancer growth and metastasis remains largely unclear.
miRNAs, a group of small non-coding RNAs measuring
22–25 nucleotides in length, are gene expression regulators that
serve key roles in cancer development and progression (15,16).
Among the cancer-associated miRNAs, miR-134 has been demonstrated
to be essential for cancer development and progression, as it
participates in cancer cell proliferation, apoptosis, migration,
invasion and drug resistance (17,18).
However, to the best of our knowledge, the function of miR-134 in
cervical cancer, and the association between miR-134 and NCK1-AS1,
has not been described previously.
The present study aimed to examine the expression of
NCK1-AS1 in cervical cancer tissues and cell lines, and to
investigate the association between NCK1-AS1 expression and
clinicopathological characteristics in cervical cancer. In
addition, in vitro experiments were performed to explore the
regulatory mechanism of NCK1-AS1 in the proliferation and migration
of cervical cancer cells involving miR-134.
Materials and methods
Tissue collection
The present study was approved by the Ethics
Committee of Chengdu Women and Children's Central Hospital
(Chengdu, China). A total of 52 pairs of cervical cancer tissues
and adjacent normal tissues were collected from patients with
primary cervical cancer at Chengdu Women and Children's Central
Hospital between March 2010 and September 2012. Written informed
consent was obtained from all participants. None of the patients
had received chemotherapy or radiotherapy prior to surgery. The
cervical cancer tissues and adjacent normal tissues were rapidly
frozen in liquid nitrogen shortly following surgical resection and
stored until use. The clinical characteristics of the patients are
summarized in Table I.
 | Table I.Association between lncRNA NCK1-AS1
expression and clinicopathological characteristics of patients with
cervical cancer. |
Table I.
Association between lncRNA NCK1-AS1
expression and clinicopathological characteristics of patients with
cervical cancer.
|
|
| NCK1-AS1 level |
|
|---|
|
|
|
|
|
|---|
| Variables | Cases (n=52) | Low (n=26) | High (n=26) | P-value |
|---|
| Age, years |
|
|
| 0.776 |
|
<55 | 20 | 9 | 11 |
|
| ≥55 | 32 | 17 | 15 |
|
| Tumor size, cm |
|
|
| 0.083 |
| ≤4 | 33 | 20 | 13 |
|
|
>4 | 19 | 6 | 13 |
|
| Differentiation |
|
|
| 0.173 |
|
Well-Moderate | 41 | 23 | 18 |
|
| Poor | 11 | 3 | 8 |
|
| Clinical stage |
|
|
| 0.003 |
| I–II | 33 | 22 | 11 |
|
|
III–IV | 19 | 4 | 15 |
|
| Lymph node
metastasis |
|
|
| 0.006 |
| No | 36 | 23 | 13 |
|
| Yes | 16 | 3 | 13 |
|
| Distant
metastasis |
|
|
| 0.051 |
| No | 47 | 26 | 21 |
|
| Yes | 5 | 0 | 5 |
|
Cell culture and transfection
A normal human cervix epithelial Ect1/E6E7 cell line
and cervical cancer SiHa, HeLa, C-33A and CaSki cell lines were
purchased from the American Type Culture Collection. The cells were
cultured in Dulbecco's modified Eagle's medium (DMEM; Thermo Fisher
Scientific, Inc.) containing 10% foetal bovine serum (Thermo Fisher
Scientific, Inc.) at 37°C in an incubator with 5% CO2.
Cell transfection was performed with 100 nM of negative control
(NC) small interfering RNA (siRNA; cat. no. 4390843; Thermo Fisher
Scientific, Inc.), NCK1-AS1 siRNA (cat. no. 4392420; Thermo Fisher
Scientific, Inc.), NC inhibitor (cat. no. 4464076; Thermo Fisher
Scientific, Inc.) or miR-134 inhibitor (cat. no. 4464084; Thermo
Fisher Scientific, Inc.) using Lipofectamine® 2000
(Invitrogen; Thermo Fisher Scientific, Inc.) according to the
manufacturer's protocol. The following experiments were conducted
48 h after transfection.
Reverse transcription quantitative
polymerase chain reaction (RT-qPCR)
Total RNA was extracted from tissues and cells using
TRIzol® reagent (Thermo Fisher Scientific, Inc.)
according to the manufacturer's protocol. RNA (1 µg) was then
reverse transcribed into cDNA using the miScript Reverse
Transcription kit (Qiagen GmbH), according to the manufacturer's
protocol. Then, qPCR was performed using cDNA with a
SYBR® Green Real-Time PCR Master Mix (Thermo Fisher
Scientific, Inc.), following the manufacturer's protocol. The qPCR
thermocycler conditions were as follows: 95°C for 3 min, followed
by 45 cycles of 95°C for 15 sec, 60°C for 15 sec and 72°C for 15
sec. GAPDH and U6 were used as internal references. The
2−ΔΔCq method was used to analyse the relative gene
expression (19). The following
primer sequences were used: GAPDH forward,
5′-CTGGGCTACACTGAGCACC-3′ and reverse, 5′-AAGTGGTCGTTGAGGGCAATG-3′;
U6 forward, 5′-CTCGCTTCGGCAGCACA-3′ and reverse,
5′-AACGCTTCACGAATTTGCGT-3′. The primers for NCK1-AS1 (cat. no.
Hs00606619_CE) and miR-134 (cat. no. Hs00500260_CE) were purchased
from Thermo Fisher Scientific, Inc.
Cell Counting Kit-8 (CCK-8) assay
The transfected HeLa and SiHa cells
(3×103 cells/well) were seeded into 96-well plates and
cultured at 37°C with 5% CO2 for 0, 24, 48 and 72 h.
Then, CCK-8 solution (Dojindo Molecular Technologies) was added,
and cells were incubated at 37°C for 2 h. Subsequently, the
absorbance value was determined at 450 nm using a microplate reader
(Bio-Rad Laboratories, Inc.).
Wound healing assay
The transfected HeLa and SiHa cells were seeded into
24-well plates and cultured at 37°C with 5% CO2 to 100%
confluence. Then, the cells were scratched with a sterile 200 µl
pipette tip and washed twice with PBS. Cells in DMEM were then
incubated at 37°C with 5% CO2 for 24 h. Images were
captured at 0 and 24 h using a light microscope (magnification,
×40).
Luciferase reporter gene assay
The target miRNAs of NCK1-AS1 were predicted using
RNAhybrid 2.12 software (http://bibiserv.techfak.uni-bielefeld.de/rnahybrid/).
miR-134 was predicted to have binding sites in the NCK1-AS1 gene.
To confirm this prediction, wild-type (WT) and mutant (Mut)
NCK1-AS1 sequences were produced by Shanghai GenePharma Co., Ltd.,
and inserted into the pGL3 luciferase vector (Promega Corporation).
HeLa and SiHa cells were transfected with miR-134 mimics or miR-NC,
together with WT or Mut pGL3-NCK1-AS1 reporter plasmids, using
Lipofectamine® 2000 according to the manufacturer's
protocol. At 48 h after transfection, luciferase activities were
determined using a Dual Luciferase Reporter Assay kit (Promega
Corporation) in accordance with the manufacturer's protocol.
Renilla luciferase activity was normalized to that of
firefly luciferase.
Western blot analysis
Transfected cells were lysed in cold
radioimmunoprecipitation assay lysis buffer (Thermo Fisher
Scientific, Inc.). The protein concentration was determined using
the BCA Protein Assay kit (Thermo Fisher Scientific, Inc.) in
accordance with the manufacturer's protocol. Proteins (50 µg/lane)
were separated with 10% SDS-PAGE and transferred to polyvinylidene
fluoride membranes (Thermo Fisher Scientific, Inc.). The membranes
were incubated in 5% non-fat dried milk (Yili Group) in PBS at 4°C
overnight. Following incubation with rabbit anti-human matrix
metalloproteinase (MMP)-2 antibody (1:500; cat. no. ab37150;
Abcam), rabbit anti-human MMP-9 antibody (1:200; cat. no. ab73734;
Abcam) or rabbit anti-human GAPDH antibody (1:500; cat. no. ab9485;
Abcam) at room temperature for 3 h, the membranes were incubated
with horseradish peroxidase-conjugated goat anti-rabbit secondary
antibody (1:5,000; cat. no. ab6721; Abcam) at room temperature for
1 h. An ECL Western Blotting kit (Thermo Fisher Scientific, Inc.)
was used to detect the immune complexes. Image-Pro Plus software
6.0 (Media Cybernetics, Inc.) was used to analyse the relative
protein expression.
Statistical analysis
Statistical analysis was performed using SPSS 19.0
software (IBM Corp.). The data are expressed as the mean ± standard
deviation. An unpaired Student's t-test was used to analyse
differences between two groups. A one-way analysis of variance and
Turkey's post-hoc test was used for multiple comparisons. The
χ2 test was performed to analyse the association between
NCK1-AS1 expression and the clinical features of patients. A
Kaplan-Meier survival curve and a log-rank test were applied for
survival analysis. Spearman's correlation analysis was conducted to
evaluate the correlation between NCK1-AS1 and miR-134 expression in
cervical cancer tissues. P<0.05 was considered to indicate a
statistically significant difference.
Results
Upregulation of NCK1-AS1 in cervical
cancer
In the present study, NCK1-AS1 expression was first
analysed in several human cervical cancer cell lines and normal
cells using the qPCR analysis. As demonstrated in Fig. 1A, NCK1-AS1 was significantly
upregulated in cervical cancer cells compared with the NCK1-AS1
levels in Ect1/E6E7 cells. To additionally confirm the upregulation
of NCK1-AS1 in cervical cancer, a total of 52 paired cervical
cancer tissues and adjacent normal tissues were analysed. The qPCR
analysis was then performed to examine NCK1-AS1 expression. As
indicated in Fig. 1B, the expression
levels of NCK1-AS1 were increased in cervical cancer tissues
compared with the expression levels in adjacent normal tissues.
Then, the clinical significance of NCK1-AS1 expression in cervical
cancer was studied. Patients were then divided into high and low
NCK1-AS1 expression groups, using its mean expression value as the
cut-off (2.44). As indicated in Table
I, a high expression NCK1-AS1 level was significantly
associated with positive lymph node metastasis and advanced
clinical stage. In addition, it was identified that the patients
with cervical cancer with high NCK1-AS1 expression exhibited a
shorter survival times compared with patients with low NCK1-AS1
expression levels (Fig. 1C). These
data suggest that NCK1-AS1 may be involved in the malignant
progression of cervical cancer.
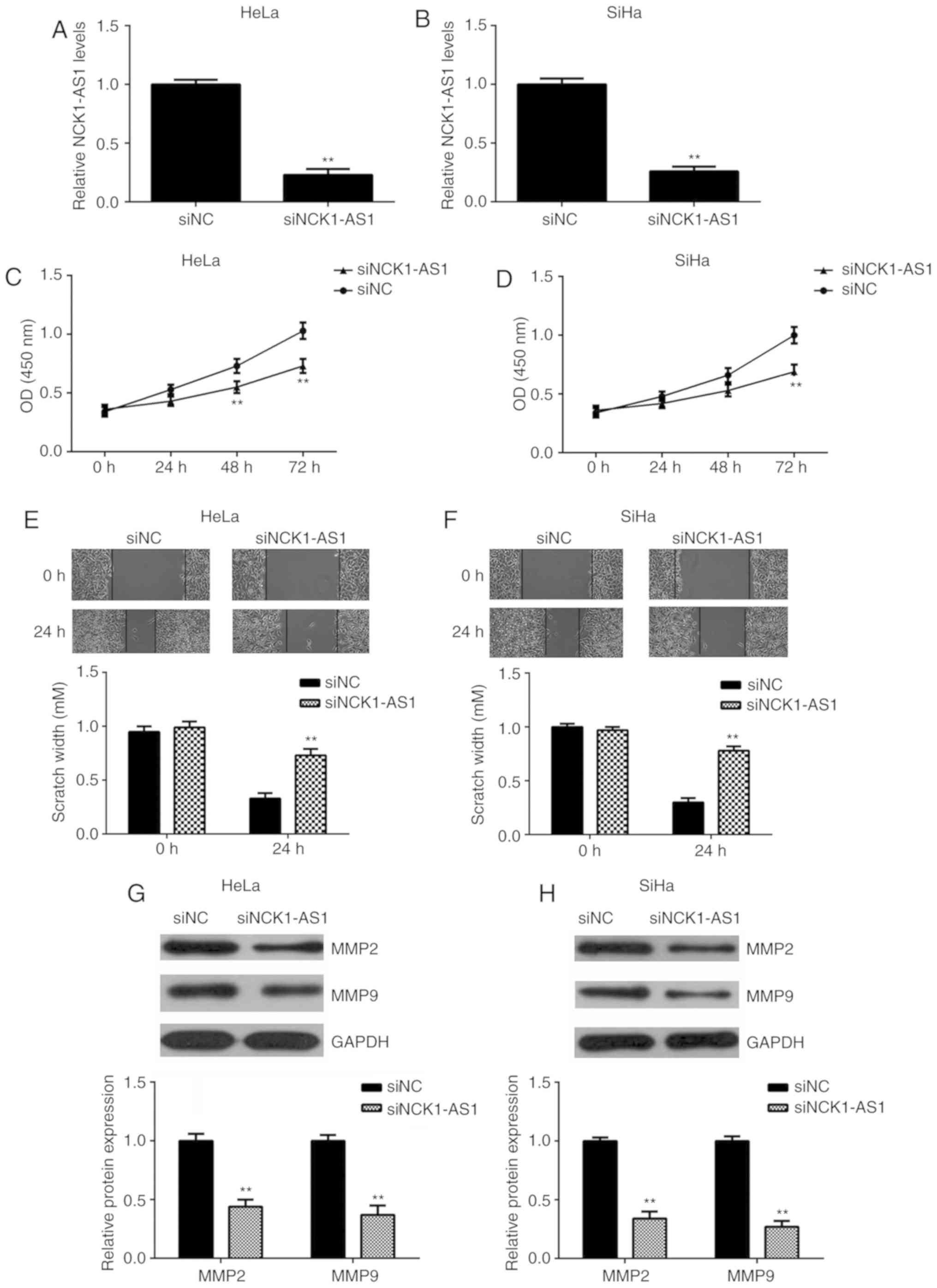
Silencing of NCK1-AS1 expression
inhibits the proliferation and migration of cervical cancer
cells
The function of NCK1-AS1 in cervical cancer was
studied using HeLa and SiHa cells. These 2 cell lines were
transfected with NCK1-AS1 siRNA to knock down the expression of
NCK1-AS1. Transfection with NC siRNA was performed as a control.
qPCR data indicated that the expression of NCK1-AS1 was
significantly downregulated following transfection with NCK1-AS1
siRNA compared with the expression in the NC siRNA group (Fig. 2A and B). The CCK-8 and wound healing
assays additionally indicated that the knockdown of NCK1-AS1
significantly decreased the proliferation and migration of HeLa and
SiHa cells (Fig. 2C-F), indicating
that NCK1-AS1 may serve a promotive role in cervical cancer cell
proliferation and migration. In addition, it was observed that
silencing of NCK1-AS1 significantly decreased the protein
expression of MMP-2 and MMP-9, 2 important markers for tumor cell
migration and invasion (Fig. 2G and
H).
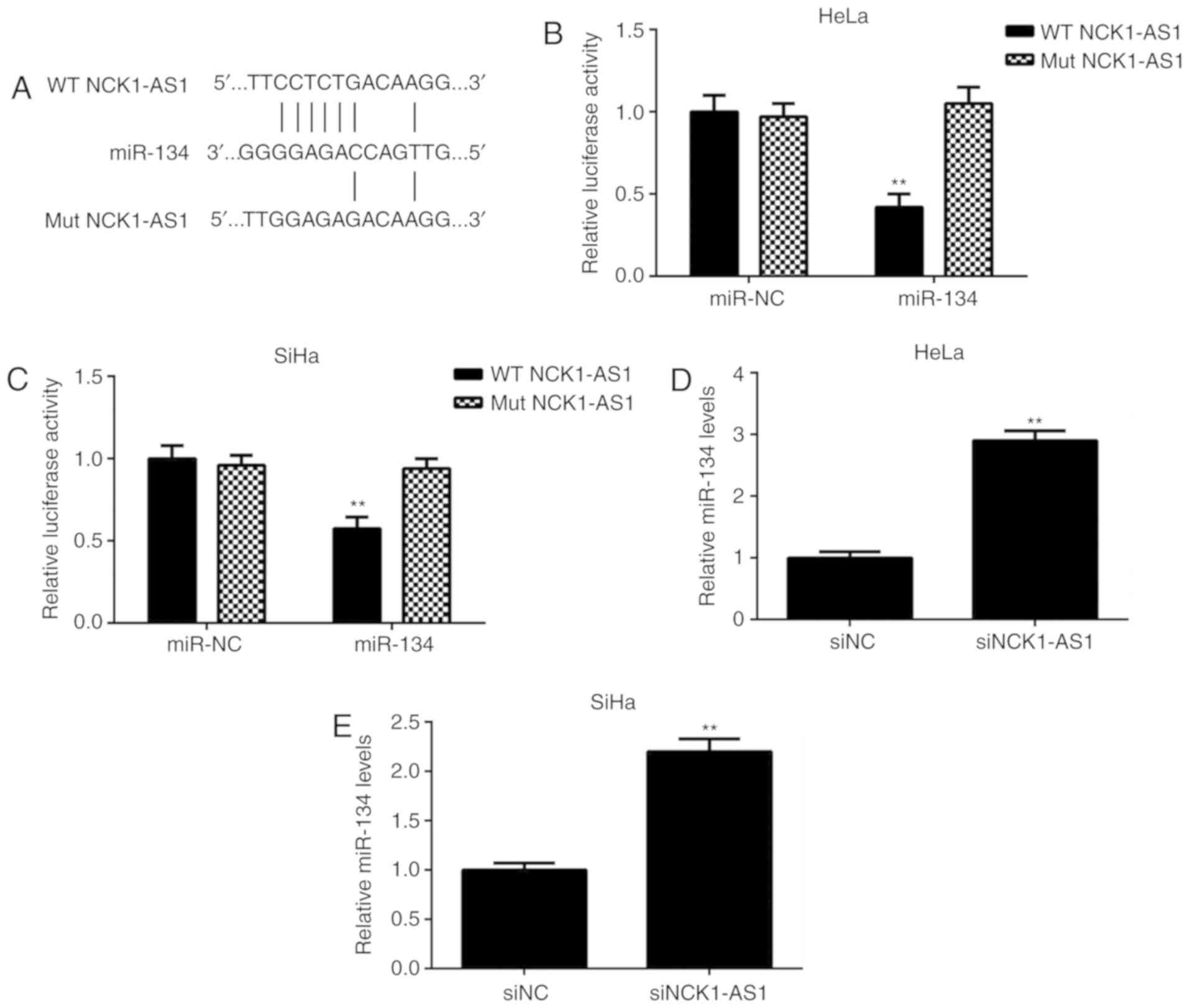
NCK1-AS1 gene contains an miR-134
binding site in cervical cancer cells
Bioinformatic analysis was then performed to study
the potential NCK1-AS1-miR interactions. As indicated in Fig. 3A, miR-134 had a potential binding
site in NCK1-AS1. To clarify this prediction, a luciferase reporter
gene assay was performed using cervical cancer cells. The data
demonstrated that transfection with miR-134 mimics led to a
significant decrease in the luciferase activity of the WT NCK1-AS1
luciferase reporter gene plasmid but did not affect the luciferase
activity of the Mut NCK1-AS1 luciferase reporter gene plasmid
(Fig. 3B and C). In addition, it was
identified that silencing of NCK1-AS1 expression caused a
significant upregulation of miR-134 in cervical cancer cells
(Fig. 3D and E). These results
suggested that NCK1-AS1 may directly target miR-134 in cervical
cancer cells.
Downregulation of miR-134 is inversely
correlated with the upregulation of NCK1-AS1 in cervical cancer
tissues
It was identified that the expression levels of
miR-134 were significantly decreased in cervical cancer cells
compared with the expression levels in Ect1/E6E7 cells (Fig. 4A). Consistently, miR-134 expression
levels were also significantly downregulated in cervical cancer
tissues compared with the expression levels in adjacent tissues
(Fig. 4B). Notably, an inverse
correlation between miR-134 and NCK1-AS1 expression was observed in
cervical cancer tissues (Fig. 4C).
Therefore, the upregulation of NCK1-AS1 may contribute to the
downregulation of miR-134 in cervical cancer.
Silencing of miR-134 expression
impairs the suppressive effects of NCK1-AS1 inhibition on cervical
cancer cell proliferation and migration
Based on the aforementioned data, we hypothesized
that miR-134 may be involved in the NCK1-AS1-mediated proliferation
and migration of cervical cancer cells. To confirm this hypothesis,
HeLa and SiHa cells were co-transfected with NCK1-AS1 siRNA
together with an NC inhibitor or a miR-134 inhibitor. As indicated
in Fig. 5A, miR-134 was
significantly downregulated in the siNCK1-AS1 + anti-miR-134 group
compared with the miR-134 levels in the siNCK1-AS1 + anti-NC group.
The CCK-8 and wound healing assays additionally demonstrated that
the proliferation and migration of cervical cancer cells were
significantly enhanced in the siNCK1-AS1 + anti-miR-134 group
compared with the siNCK1-AS1 + anti-NC group (Fig. 5B-E), indicating that the knockdown of
miR-134 impaired the inhibitory effects on cervical cancer cell
proliferation and migration induced by NCK1-AS1 downregulation.
Taken together, these data suggested that miR-134 was involved in
the NCK1-AS1-mediated proliferation and migration of cervical
cancer cells.
Discussion
The regulatory mechanism underlying the effect of
NCK1-AS1 in cervical cancer progression remains largely unclear. In
the present study, it was identified that NCK1-AS1 was
significantly upregulated in cervical cancer tissues compared with
the NCK1-AS1 expression in adjacent non-tumour tissues. High
expression levels of NCK1-AS1 were associated with tumour
progression and poor prognosis in patients with cervical cancer.
Silencing of NCK1-AS1 expression decreased cervical cancer cell
proliferation and migration, and decreased the protein expression
levels of MMP-2 and MMP-9. The results of the luciferase reporter
gene assay indicated that an miR-134 binding site was contained
within the NCK1-AS1 gene. miR-134 was significantly downregulated
in cervical cancer tissues compared with the miR-134 levels in
adjacent non-tumour tissues, and its expression levels were
inversely correlated with the expression levels of NCK1-AS1 in
cervical cancer tissues. Knockdown of miR-134 attenuated the
inhibitory effects on the proliferation and migration of cervical
cancer cells induced by NCK1-AS1 downregulation.
Recently, a number of lncRNAs have been suggested to
be significantly deregulated in cervical cancer; some of these
lncRNAs are associated with tumour progression and poor prognosis
(20,21). For example, lncRNA HOX transcription
antisense RNA is significantly upregulated in cervical cancer and
promotes cancer progression by increasing the expression of Bcl-2
via the inhibition of miR-143-3p (22). The lncRNA Pvt-1 oncogene facilitates
the proliferation, migration and invasion of cervical cancer cells
by negatively regulating miR-424 (21). In the present study, an upregulation
of NCK1-AS1 was observed in cervical cancer, and high expression
levels of NCK1-AS1 were associated with lymph node metastasis,
advanced clinical stage and poor prognosis of patients. These data
suggested that NCK1-AS1 upregulation may contribute to cervical
cancer progression. The role of NCK1-AS1 in cervical cancer growth
and metastasis in vitro was also studied. To knock down the
expression of NCK1-AS1, cervical cancer cells were transfected with
NCK1-AS1-specific siRNA. The results demonstrated that the
silencing of NCK1-AS1 significantly inhibited the proliferation and
migration of cervical cancer cells, suggesting that NCK1-AS1 may
serve a promotive role in cervical cancer growth and
metastasis.
The downstream miRNAs of NCK1-AS1 was then studied
in cervical cancer cells, and bioinformatics analysis predicted
that miR-134 and NCK1-AS1 had binding sites in cervical cancer
cells. Previous studies have demonstrated that miR-134 serves an
oncogenic or a tumour suppressive role in different types of human
cancer: For example, Peng et al (7) revealed that miR-134 targeted PDCD7 to
decrease epithelial cadherin expression and enhance oral cancer
progression. By contrast, However, to the best of our knowledge,
the expression pattern and the detailed role of miR-134 in cervical
cancer has not been studied. In the present study, it was
identified that the expression of miR-134 was significantly
decreased in cervical cancer tissues and cell lines compared with
that in adjacent tissues and normal cells. In addition, it was
demonstrated that the expression of miR-134 was inversely
correlated with NCK1-AS1 expression in cervical cancer tissues, and
that silencing of NCK1-AS1 expression increased miR-134 expression
in cervical cancer cells. These data suggested that the
downregulation of miR-134 may be due to the upregulation of
NCK1-AS1 in cervical cancer. It was then revealed that the
inhibition of miR-134 impaired the suppressive effects on the
proliferation and migration of cervical cancer cells induced by
NCK1-AS1 knockdown, which suggested that miR-134 may serve as a
downstream effecter of NCK1-AS1 in cervical cancer cells.
In addition to NCK1-AS1, several other lncRNAs have
been identified to directly target miR-134 in other cancer types.
For example, through the inhibition of miR-134-5p, the lncRNA
prostate cancer associated transcript 7 regulated E74 like ETS
transcription factor 2 signalling in nasopharyngeal carcinoma
(23), and induced poor prognosis
and promoted tumourigenesis in non-small-cell lung cancer (24). Therefore, the results of the present
study improve the understanding of the LncRNA/miRNA axis in human
cancer.
To the best of our knowledge, this is the first
study to demonstrate that silencing of NCK1-AS1 expression
repressed the proliferation and migration of cervical cancer cells
via miR-134, suggesting that NCK1-AS1 may be used as a potential
therapeutic target for cervical cancer.
Acknowledgements
Not applicable.
Funding
No funding was received.
Availability of data and materials
All data generated or analysed during this study are
included in this published article.
Authors' contributions
JY and LHu designed the study and wrote the
manuscript. XG collected clinical tissues and performed statistical
analysis. LHu, LHe and LW performed all experiments. All authors
read and approved the final manuscript.
Ethics approval and consent to
participate
This study was approved by the Ethics Committee of
Chengdu Women and Children's Central Hospital. Written informed
consent was obtained from all patients involved.
Patient consent for publication
All patients provided written informed consent.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Jemal A, Siegel R, Ward E, Murray T, Xu J,
Smigal C and Thun MJ: Cancer statistics, 2006. CA Cancer J Clin.
56:106–130. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Torre LA, Bray F, Siegel RL, Ferlay J,
Lortet-Tieulent J and Jemal A: Global cancer statistics, 2012. CA
Cancer J Clin. 65:87–108. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
King T: HPV, cervical cancer, and you. J
Midwifery Womens Health. 53:263–264. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Yang WT and Zheng PS: Kruppel-like factor
4 functions as a tumor suppressor in cervical carcinoma. Cancer.
118:3691–3702. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Shen Y, Zhou J, Li Y, Ye F, Wan X, Lu W,
Xie X and Cheng X: miR-375 mediated acquired chemo-resistance in
cervical cancer by facilitating EMT. PLoS One. 9:e1092992014.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Zhou S, Yu L, Xiong M and Dai G: LncRNA
SNHG12 promotes tumorigenesis and metastasis in osteosarcoma by
upregulating Notch2 by sponging miR-195-5p. Biochem Biophys Res
Commun. 495:1822–1832. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Peng Z, Liu C and Wu M: New insights into
long noncoding RNAs and their roles in glioma. Mol Cancer.
17:612018. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Xu R, Zhu X, Chen F, Huang C, Ai K, Wu H,
Zhang L and Zhao X: LncRNA XIST/miR-200c regulates the stemness
properties and tumourigenicity of human bladder cancer stem
cell-like cells. Cancer Cell Int. 18:412018. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Xiong W, Huang C, Deng H, Jian C, Zen C,
Ye K, Zhong Z, Zhao X and Zhu L: Oncogenic non-coding RNA NEAT1
promotes the prostate cancer cell growth through the SRC3/IGF1R/AKT
pathway. Int J Biochem Cell Biol. 94:125–132. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Xiao Z, Qu Z, Chen Z, Fang Z, Zhou K,
Huang Z, Guo X and Zhang Y: LncRNA HOTAIR is a prognostic biomarker
for the proliferation and chemoresistance of colorectal cancer via
miR-203a-3p-mediated Wnt/β-catenin signaling pathway. Cell Physiol
Biochem. 46:1275–1285. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Sun Y, Jin JG, Mi WY, Zhang SR, Meng Q and
Zhang ST: Long noncoding RNA UCA1 targets miR-122 to promote
proliferation, migration, and invasion of glioma cells. Oncol Res.
26:103–110. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Lv L, Jia JQ and Chen J: The lncRNA CCAT1
upregulates proliferation and invasion in melanoma cells via
suppressing miR-33a. Oncol Res. 26:201–208. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Zhu H, Zheng T, Yu J, Zhou L and Wang L:
LncRNA XIST accelerates cervical cancer progression via
upregulating Fus through competitively binding with miR-200a.
Biomed Pharmacother. 105:789–797. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Zhu J, Shi H, Liu H, Wang X and Li F: Long
non-coding RNA TUG1 promotes cervical cancer progression by
regulating the miR-138-5p-SIRT1 axis. Oncotarget. 8:65253–65264.
2017.PubMed/NCBI
|
|
15
|
Li H, Jia Y, Cheng J, Liu G and Song F:
LncRNA NCK1-AS1 promotes proliferation and induces cell cycle
progression by crosstalk NCK1-AS1/miR-6857/CDK1 pathway. Cell Death
Dis. 9:1982018. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Bartel DP: MicroRNAs: Genomics,
biogenesis, mechanism, and function. Cell. 116:281–297. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Pan JY, Zhang F, Sun CC, Li SJ, Li G, Gong
FY, Bo T, He J, Hua RX, Hu WD, et al: miR-134: A human cancer
suppressor? Mol Ther Nucleic Acids. 6:140–149. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Liu Y, Sun Y and Zhao A: MicroRNA-134
suppresses cell proliferation in gastric cancer cells via targeting
of GOLPH3. Oncol Rep. 37:2441–2448. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Zhang JJ, Wang DD, Du CX and Wang Y: Long
noncoding RNA ANRIL promotes cervical cancer development by acting
as a sponge of miR-186. Oncol Res. 26:345–352. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Gao YL, Zhao ZS, Zhang MY, Han LJ, Dong YJ
and Xu B: Long Non-coding RNA PVT1 facilitates cervical cancer
progression via negative regulating of miR-424. Oncol Res.
25:1391–1398. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Liu M, Jia J, Wang X, Liu Y, Wang C and
Fan R: Long non-coding RNA HOTAIR promotes cervical cancer
progression through regulating BCL2 via targeting miR-143-3p.
Cancer Biol Ther. 19:391–399. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Liu Y, Tao Z, Qu J, Zhou X and Zhang C:
Long non-coding RNA PCAT7 regulates ELF2 signaling through
inhibition of miR-134-5p in nasopharyngeal carcinoma. Biochem
Biophys Res Commun. 491:374–381. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Liu Q, Wu Y, Xiao J and Zou J: Long
Non-coding RNA prostate cancer-associated transcript 7 (PCAT7)
induces poor prognosis and promotes tumorigenesis by inhibiting
mir-134-5p in non-small-cell lung (NSCLC). Med Sci Monit.
23:6089–6098. 2017. View Article : Google Scholar : PubMed/NCBI
|