Introduction
Globally, colorectal cancer (CC) is the third
leading cause of mortality associated with cancer (1). Worldwide, the increasing incidence of
CC is possibly caused by the modern life style which is
characterized by increased fat intake and reduced physical activity
(2). In CC, poor efficiency and lack
of effective methods for treating metastasis are the main causes
for mortality among patients (3).
For patients with local disease, the five-year survival rate can be
as high as 90.3%, but it declines to 70.4 and 12.5% for those with
regional and distant metastasis, respectively (3). Despite advances in the medical science
and technology area, the molecular mechanisms underlying CC
progression and pathogenesis remain unclear, which is important to
be elucidated.
The immune system is responsible for eliminating
cancerous cells and foreign infections (4). In particular, natural killer (NK) cells
are primarily responsible for eliminating tumor cells through
contact-dependent cytotoxicity and cytokine production (5). For instance, NK-92 cells attack cancer
cells and the tumors grown within the control of the organism
(6). One of those cytokines,
interferon gamma (IFN-γ), is secreted by NK cells and has been
previously reported to promote the apoptosis and cytolysis of
target tumor cells (4,7). IFN-γ has immunoregulatory, antiviral
and anti-tumor properties (8).
Additionally, in cancer cells, IFN-γ results in the inhibition of
cell proliferation (8). In cancer
cells, IFN-γ is expressed at higher levels and results in cell
death or growth inhibition (9).
Therefore, it is vital to study the molecular mechanisms behind the
NK cell-mediated killing of CC cells.
Cytokines produced during the process of the innate
immune response are important components linking inflammation with
cancer (10). IFN-γ has previously
been demonstrated to contribute to the antitumor activity of a
number of interleukins (ILs) (11).
IL-15 is a pleiotropic cytokine expressed and secreted by dendritic
cells, macrophages, fibroblasts and epithelial cells (12). IL-15 has demonstrated the ability to
suppress colitis-associated colon carcinogenesis through the
induction of antitumor immunity (13). However, the effects of IFN-γ on IL-15
in regulating tumor progression remain unknown. Since the
establishment of NK-92 cells in 1992, their anti-cancer activity
has been widely tested in mouse models (14). Therefore, pAb-IL-15R was used to
inhibit IL-15R signaling in NK-92 cells in the present study, we
aimed to investigate the role of NK-mediated IFN-γ in CC
progression and provide the potential molecular mechanism in this
process.
Materials and methods
Participants
For the present study, 21 patients with CC (aged
55±5 years old, 15 males and 6 females) and 21 healthy volunteers
(aged 53±7 years old, 15 males and 6 females) were enrolled in the
First Affiliated Hospital of Xi'an Jiaotong University between
February 2015 and October 2016. Patients who received any
radio/chemo-therapy are excluded from the study. All study
participants provided written informed consent and the present
study was approved by the Ethics Committee of the First Affiliated
Hospital of Xi'an Jiaotong University.
Peripheral blood mononuclear cells (PBMCs) were
obtained from the patients with CC and healthy volunteers using
Lymphocyte Separation medium (MP Biomedicals, LLC) as described: In
brief, venous blood (10 ml) was collected in the early morning.
This was anticoagulated using heparin (10 IU/ml; Merck KGaA), mixed
with Lymphocyte Separation medium and centrifuged at 1,600 × g for
20 min at 4°C. Cells were separated into 4 layers, namely; i)
plasma or tissue homogenate layer; ii) cyan milky lymphocyte or
monocyte layer; iii) transparent separation layer; and iv) a red
blood cell layer. Cells in the second layer were collected and
washed three times with normal saline and centrifuged at 1,600 × g
for 10 min at 4°C, followed by removal of the supernatant. The
cells were resuspended in 10% DMSO and 90% fetal calf serum (Gibco;
Thermo Fisher Scientific, Inc.) at a density of 5×107
cells/ml, and finally frozen and stored in liquid nitrogen. Cells
were counted using a Nexcelom Cellometer Auto 2000 (Nexcelom
Bioscience, LLC) by acridine orange (AO)/propidium iodide (PI)
staining (AO/PI cell viability kit, Nexcelom Bioscience, LLC.)
according to the manufacturer's protocol. PBMCs exhibited >95%
viability before and after freezing.
NK cells were extracted from PBMCs using Human NK
Cell Isolation kit (negative selection procedure of magnetic
activated cell sorting; cat. no. 130-092-657; Miltenyi Biotec,
Inc.) according to the manufacturer's protocol. In brief, non-NK
cells including T cells, B cells, stem cells, dendritic cells,
monocytes, granulocytes and erythroid cells, were magnetically
labeled with the cocktail of biotin-conjugated antibodies and the
NK Cell MicroBead Cocktail. NK cells were isolated by depletion of
the magnetically labeled cells; NK cells with >90% purity at the
end was determined by flow cytometry.
Cell culture
NK-92 cells were (NantKwest, Inc.) cultured in
minimum essential medium Eagle (Sigma-Aldrich; Merck KGaA) with
Earle's salts and nonessential amino acids, supplemented with 12.5%
fetal bovine serum (FBS; Gibco; Thermo Fisher Scientific, Inc.),
12.5% horse serum (Gibco; Thermo Fisher Scientific, Inc.), 2 mM
L-glutamine (Gibco; Thermo Fisher Scientific, Inc.), 100 µM
2-mercaptoethanol (Sangon Biotech Co., Ltd.), 100 U/ml penicillin
(Sangon Biotech Co., Ltd.) and 100 U/ml streptomycin (Sangon
Biotech Co., Ltd.) at 37°C with 95% humidity and 5% CO2.
For the stimulation of NK-92 cells, they were incubated with IL-2
(100 U/ml; PeproTech, Inc.) for 24 h at 37°C. For the
neutralization of IL-15 bioactivity in NK-92 cells, NK-92 cells
were treated with anti-IL-15/IL-15R Complex Monoclonal Antibody
(pAb IL-15; clone GRW15PLZ; eBioscience; Thermo Fisher Scientific,
Inc.) for 12 h at 37°C.
Human colorectal carcinoma cell line SW480 was
purchased from American Type Culture Collection (ATCC). SW480 cells
were cultured in DMEM (Invitrogen; Thermo Fisher Scientific, Inc.)
with 10% FBS (Gibco; Thermo Fisher Scientific, Inc.), 100 U/ml
penicillin (Sangon Biotech Co., Ltd.), and 100 U/ml streptomycin
(Sangon Biotech Co., Ltd.) in an incubator at 37°C with a 95%
humidified atmosphere and 5% CO2.
Cytotoxicity assays
CytoTox 96® non-radioactive cytotoxicity
assay (Promega Corporation) was used for the detection of SW480
cell killing by NK-92 cells according to the manufacturer's
protocol as a previous report (15).
SW480 cells were first washed with PBS, re-suspended in fresh NK-92
culture medium (fully supplemented as previously described) and
seeded into 96-well plates (density, 5×103 cells/well).
NK-92 media alone did not affect the SW480 cells (data not shown).
Immediately, IL-2-induced NK-92 cells were subsequently added to
the SW480 cells at an effector-to-target ratio of 16:1 and
incubated at 37°C with 95% humidified atmosphere and 5%
CO2 for 4 h. The cytotoxic activity was measured by LDH
release from the supernatant which was obtained and analyzed using
the CytoTox 96 assay. The SW480 cell killing was calculated using
the following equation: % cytotoxicity=(Experimental-Effector
spontaneous-Target spontaneous)/(Target maximum-Target spontaneous)
×100, according to the manufacturer's protocol. Maximum and
spontaneous are both referring to LDH release.
Cell treatment
SW480 cells were randomly divided into 3 groups: i)
Control group, SW480 cells co-cultured with non-stimulated NK-92
cells; ii) IL-12 group, SW480 cells co-cultured with IL-2-activated
NK-92 cells; and iii) IL-12 + pAb IL-15 group, SW480 cells
co-cultured with IL-2 + anti-IL-15/IL-15R Complex Monoclonal
Antibody treated NK-92 cells.
Enzyme-linked immunosorbent assay
(ELISA)
Briefly, the NK-92 cells which were stimulated by
IL-12 for 24 h at 37°C were washed with PBS and seeded into 96-well
plates at a seeding density of 5×103 cells/well and
cultured for 4 h at 37°C with 5% CO2. They were then
centrifuged at 200 × g for 4 min at room temperature. Supernatants
were subsequently collected for the quantification of IFN-γ
concentration using the human IFN-γ ELISA kit (cat. no. EK0458;
Signalway Antibody LLC) in accordance with the manufacturer's
protocol. The absorbance at 450 nm was measured in each well using
a microplate reader (Synergy™ HT; BioTek Instruments, Inc.).
Cell Counting Kit (CCK-8) assay
SW480 cells were randomly divided into 3 groups, the
control group, IL-12 group and IL-12 + pAb IL-15 group. Cell
proliferation was detected by CCK-8 (Dojindo Molecular
Technologies, Inc.) according to the manufacturer's protocol. In
brief, SW480 cells (seeding density, 1×105) were seeded
in Biocoat™ 24-well plates (BD Biosciences) in DMEM, 10% FBS, 100
U/ml penicillin (Sangon Biotech Co., Ltd.) and 100 U/ml
streptomycin (Sangon Biotech Co., Ltd.) in an incubator at 37°C
with a 95% humidified atmosphere and 5% CO2 for 24 h.
Afterwards, 10 µl of CCK-8 solution was added into each well and
incubated at 37°C for 2 h. The absorbance in each well was recorded
at 450 nm by a microplate reader.
Flow cytometry analysis
SW480 cells were randomly divided into 3 groups, the
control group, IL-12 group and IL-12 + pAb IL-15 group. To measure
apoptosis, SW480 cells were seeded into 12-well plates at a density
of 3×105 cells/well and cultured for 48 h at 37°C with
95% humidified atmosphere and 5% CO2. Thereafter, the
SW480 cells were collected using 0.025% trypsin (Thermo Fisher
Scientific, Inc.). After washing with PBS, 5 µl fluorescein
isothiocyanate (FITC)-labeled Annexin V and 5 µl PI from Annexin
V-FITC apoptosis detection kit (Sigma-Aldrich; Merck KGaA) were
added into each well and incubated in the dark for 15 min at 37°C
with 95% humidified atmosphere and 5% CO2. Cell
apoptosis was analyzed using flow cytometry (BD Biosciences) within
1 h, and the data were analyzed using the FlowJo software (version
10.2; FlowJo LLC).
Reverse transcription-quantitative PCR
(RT-qPCR)
Total RNA was extracted from cells using RNAiso
reagent (Takara Biotechnology Co., Ltd.) in accordance with the
manufacturer's protocol. The quality and quantity of RNA were
determined using a NanoDrop™ 1000 spectrophotometer (Thermo Fisher
Scientific, Inc.). cDNA was synthesized from RNA using PrimeScript™
II 1st Strand cDNA Synthesis kit (Takara Biotechnology Co., Ltd.)
in accordance with the manufacturer's protocol. qPCR was performed
using a SYBR-Green PCR kit (Takara Biotechnology Co., Ltd.) in a
StepOne™ Plus Real-Time PCR System (Thermo Fisher Scientific, Inc.)
for the measurement of the mRNA expression profiles. GAPDH was used
as an internal control. The primers used were as follows: IL-15
forward, 5′-ATGTTCATCAACACGTCCTGACT-3′ and reverse,
5′-GCAGCAGGTGGAGGTACCTTAA-3′; and GAPDH forward,
5′-ACACCCACTCCTCCACCTTT-3′ and reverse, 5′-TTACTCCTTGGAGGCCATGT-3′.
The thermocycling conditions used to perform the qPCR were as
listed: Initial denaturation at 95°C (30 sec), followed by 40
cycles of 95°C (5 sec) and 60°C (30 sec). Data were analyzed using
the 2−ΔΔcq method (16).
Western blot analysis
Cells were lysed using radioimmunoprecipitation
assay buffer (Beyotime Institute of Biotechnology). After that,
protein concentration was determined using the BCA method. Equal
amounts of protein (10 µg/lane) were separated by SDS-PAGE (10%
gel) and subsequently transferred onto polyvinylidene difluoride
membranes for 2 h at 4°C. The polyvinylidene difluoride membranes
were then blocked by 5% non-fat milk at 37°C for 1 h and incubated
with primary antibodies for IL-15 (cat. no. ab7213; 1:1,000;
Abcam), IFN-γ (cat. no. 8455; 1:1,000; Cell Signaling Technology,
Inc.), BAX (cat. no. 5023; 1:1,000; Cell Signaling Technology,
Inc.), Bcl-2 (cat. no. 4223; 1:1,000; Cell Signaling Technology,
Inc.), p-JAK2 (cat. no. 3776; 1:1,000; Cell Signaling Technology,
Inc.), JAK2 (cat. no. 3230; 1:1,000; Cell Signaling Technology,
Inc.), p-STAT3 (cat. no. 9145; 1:1,000; Cell Signaling Technology,
Inc.), STAT3 (cat. no. 12640; 1:1,000; Cell Signaling Technology,
Inc.) and GAPDH (cat. no. 5174; 1:1,000; Cell Signaling Technology,
Inc.) at 4°C overnight, where GAPDH served as the reference
protein. The membranes were incubated further with anti-rabbit IgG,
horseradish peroxidase-conjugated secondary antibodies (cat. no.
7074; 1:2,000; Cell Signaling Technology, Inc.) for 2 h at 37°C.
All blots were visualized using an ECL reagent (Beyotime Institute
of Biotechnology). Densitometry was performed using ImageJ software
(version. 1.8.0; National Institutes of Health).
Statistical analysis
Each experiment was performed at least 3 times.
Values are expressed as the mean ± SD. GraphPad Prism (GraphPad
Software Inc.) was used for the analysis of experimental data.
Statistical comparisons between 2 groups were performed using
Student's t-test. Statistical comparisons between 3 groups were
performed using one-way ANOVA followed by Dunnett's t-test.
P<0.05 was considered to indicate a statistically significant
difference.
Results
IL-15 expression in NK cells isolated
from patients
To measure IL-15 expression in NK cells from
patients with CC compared with healthy volunteers, RT-qPCR and
western blot analysis were performed to detect the levels of IL-15
mRNA and protein, respectively. IL-15 expression was significantly
reduced in NK cells from the CC group compared with the healthy
volunteer group (Fig. 1A). Likewise,
IL-15 protein levels were markedly lower in CC group compared with
in the healthy volunteer group (Fig.
1B).
IL-15 expression in NK-92 cells
Cells were randomly divided into two treatment
groups: The control group and the IL-2 treatment group. IL-15
expression was measured in the NK-92 cell line in response to IL-2
stimulation. IL-15 mRNA (Fig. 2A)
and protein (Fig. 2B) expression was
markedly increased in NK-92 cells following IL-2 treatment compared
with untreated control cells.
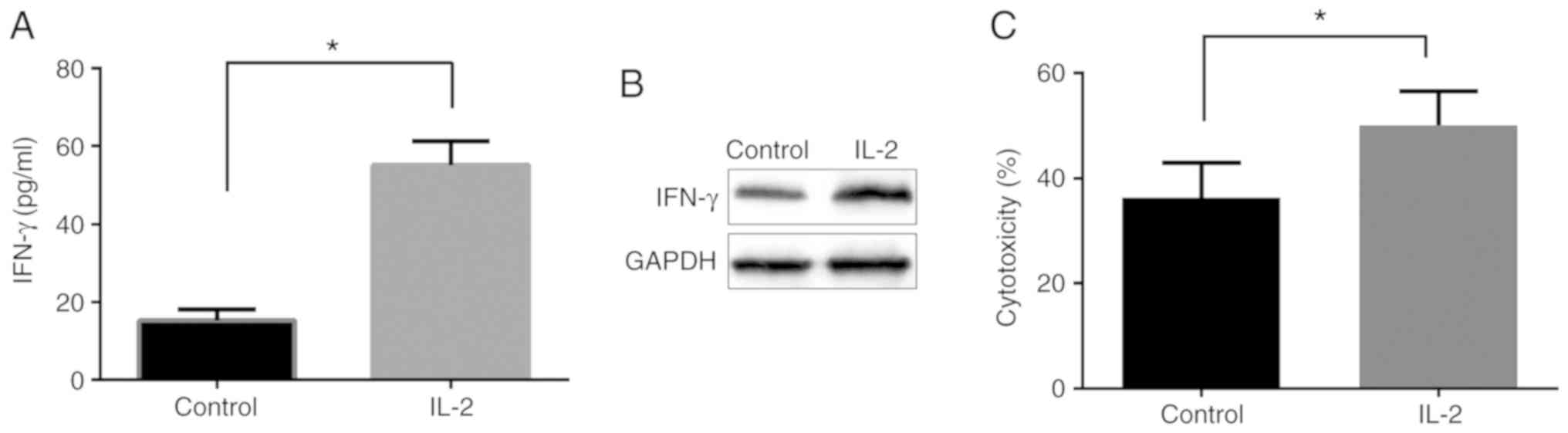
Activation of NK-92 cells by IL-2
affects the killing effect
In the present study, cells were randomly separated
into two groups, including the control and IL-2 groups. It was
found that IFN-γ secretion (Fig. 3A)
and expression (Fig. 3B) following
IL-2 stimulation were markedly higher compared with that of the
control group, suggesting the enhancement of the activation of
NK-92 cells.
The killing effect of NK-92 cells was subsequently
detected using CytoTox 96 non-radioactive cytotoxicity assay.
Compared with the control group, IL-2 stimulation enhanced the
killing effect of NK cells on SW480 cells (Fig. 3C).
Activated NK-92 cells regulate SW480
cell apoptosis
In the present study, following the stimulation of
NK-92 cells using IL-2, the effects of NK-92 cells on SW480 cell
apoptosis were determined using flow cytometry. Cells were randomly
separated into three groups, including control, IL-2 and IL-2 + pAb
IL-15 groups. In the IL-2 + pAb IL-15 group, SW480 cells were
co-cultured with NK-92 cells that were pre-treated with pAb IL-15
prior to IL-2 stimulation. Compared with the control group, NK-92
cells treated with IL-2 significantly increased SW480 cell
apoptosis, but this effect was not observed in the IL-2 + pAb IL-15
group (Fig. 4A and B).
Activated NK-92 cells regulate SW480
cell proliferation
Therefore, the effects of NK-92 cells on SW480 cell
proliferation were measured using CCK-8 assay. Accordingly, cells
were randomly separated into three groups, including the control,
IL-2 and IL-2 + pAb IL-15 groups. Compared with the control group,
NK-92 cells treated with IL-2 significantly reduced SW480 cell
proliferation, whilst no significant effects on SW480 cell
proliferation were observed following co-culture with NK-92 cells
co-treated with of IL-2 and pAb IL-15 (Fig. 5).
Activated NK-92 cells regulate the
expression of proteins associated with apoptosis in SW480
cells
Compared with control group, IL-2 treated NK92 cells
significantly increased BAX expression in SW480 cells, whilst
significantly reducing Bcl-2 expression (Fig. 6A-C). IL-2 treated NK92 cells also
significantly reduced the ratios of p-JAK2/JAK2 and p-STAT1/STAT1.
The effects of IL-2 treated NK92 cells on SW480 cells were reversed
when pAb IL-15 was included alongside IL-2 (Fig. 6A, D and E).
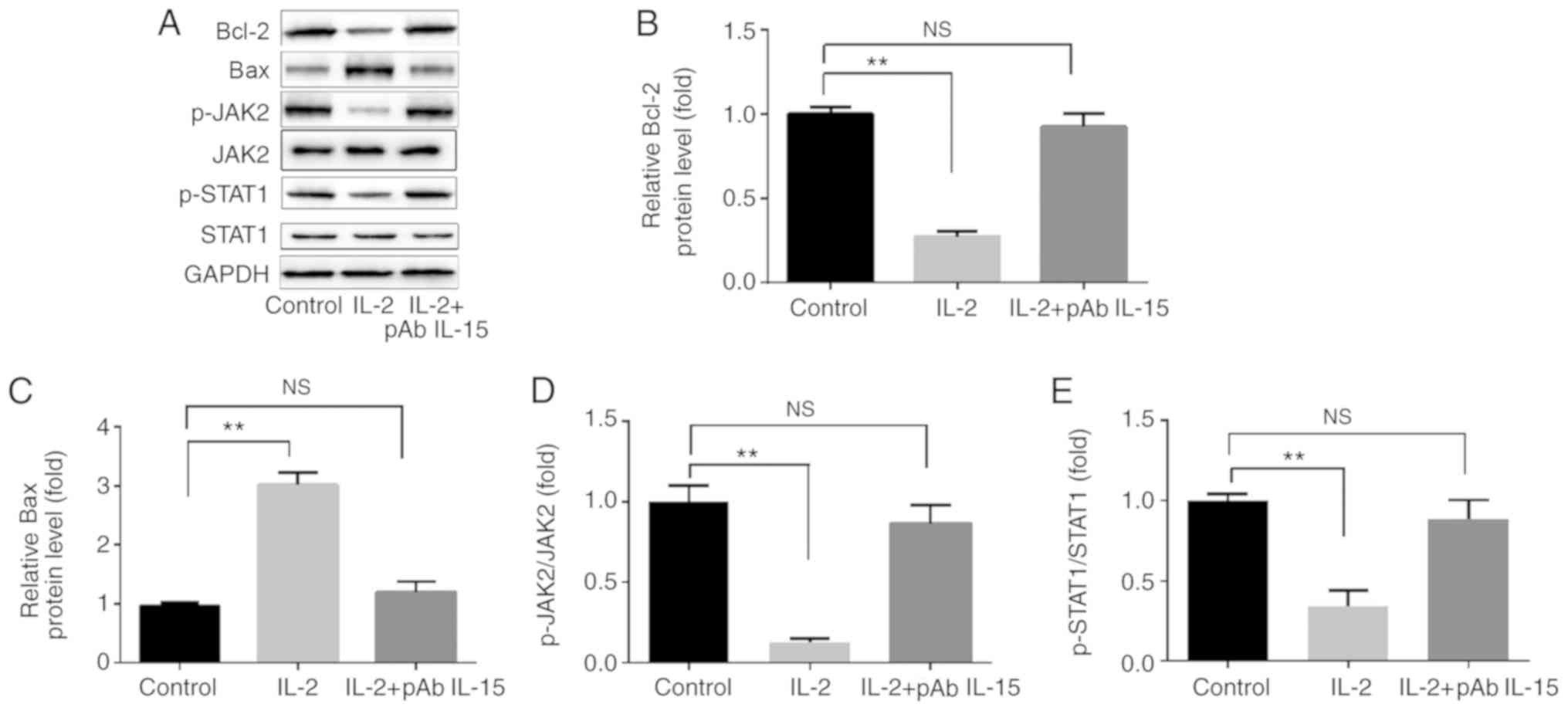 | Figure 6.Activated NK-92 cells regulate the
expression of proteins associated with apoptosis. (A)
Representative western blots comparing the expression of Bax and
Bcl-2 in addition to JAK2 and STAT1 phosphorylation in control,
IL-2 and IL-2 + pAb IL-15 groups. Compared with the control group,
IL-2 stimulation significantly (B) increased BAX expression, (C)
reduced Bcl-2 expression, and reduced the ratios of (D) p-JAK2/JAK2
and (E) p-STAT3/STAT3. All of the aforementioned effects were
reversed when pAb IL-15 was co-administered with IL-2. **P<0.01.
NS, not significant; IL, interleukin; JAK2, Janus kinase 2; p-JAK2,
phosphorylated JAK2; p-STAT3, phosphorylated STAT3; pAb, polyclonal
antibody. |
Discussion
Globally, CC is the third leading cause of mortality
associated with cancer (1).
Therefore, it is of upmost importance to investigate the molecular
mechanism underlying the pathogenesis of CC.
NK cells are primarily responsible for the
elimination of tumor cells (5). In
particular, IL-15 is a pleiotropic cytokine secreted and expressed
by dendritic cells, macrophages, fibroblasts and epithelial cells
(12), in addition to being
upregulated in macrophages during inflammation (17). In preclinical studies, IL-15 has been
demonstrated to promote humoral and cell-mediated immune responses
to inhibit tumor growth (18,19).
Indeed, IL-15 inhibits colitis-associated colon carcinogenesis by
inducing antitumor immunity (13) in
addition to upregulating IFN-γ secreted by CD8+ T-cells,
and prolongs the survival of tumor-bearing animals (20). IL-15 has attracted increasing
attention as a possible antitumor therapeutic agent in the
immunotherapy of malignancies due to the reported enhancement of NK
cell-mediated cytotoxicity (21).
Consistently, in the present study, it was found that levels of
IL-15 were significantly reduced in NK cells isolated from patients
with CC compared with the healthy volunteer group. IL-15 expression
was significantly increased in NK-92 cells following IL-2
stimulation compared with the control group.
IFNs are cytokines produced by host immune cells to
non-specifically repress viral replication (22,23).
Among the list of commercialized enzymes, vaccines, antibodies and
antibiotics, IFNs exhibit remarkable therapeutic efficacy against a
large number of diseases (24). In
line with these previous reports, the present study found that IL-2
stimulation enhanced the production/secretions of IFN-γ by NK-92
cells which enhanced SW480 cell killing by NK-92 cells, suggesting
a therapeutic potential of IFN-γ in CC patients.
In cancer cells, IFN-γ is secreted by NK cells to
induce cell apoptosis (7) and growth
inhibition (9). Compared with the
control group, IL-2 stimulation significantly increased cell
apoptosis whilst reducing cell viability, both of which were
reversed when co-administered with pAb IL-15, suggesting that IL-15
is indispensable for the function of IL-2. However, the molecular
mechanism involved in apoptotic progression require further
study.
The Bcl-2 protein family involved in the process of
apoptosis includes Bax, which is pro-apoptotic, and Bcl-2, which is
anti-apoptotic (25). Compared with
the control group, IL-2 stimulation significantly increased BAX
expression whilst reducing Bcl-2 expression. The effect of IL-2 was
reversed in the presence of pAb IL-15; however, the components
lying upstream of BAX and Bcl-2 remain unclear.
During signal transduction, IFN-γ receptor binds to
JAK2 binding domains (26), which is
followed by the phosphorylation of STAT3 and transcription of
IFN-stimulated genes (ISG) (24) or
IFN-regulated factor 1 (27). In the
current study, compared with the control group, IL-2 treatment
significantly reduced p-JAK2 and p-STAT1 levels, which was negated
by the presence of pAb IL-15.
The present study investigated the role of
NK-mediated IFN-γ in CC progression. However, the relationship
between IFN-γ and IL-15, the molecular pathway linking JAK2/STAT1
with BAX/Bcl-2 and changes in the expression of ISGs remain
unclear, and should be explored further in the future.
Acknowledgements
Not applicable.
Funding
No funding was received.
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
FC and KN designed the study. FC, DQ, RS and MZ
performed experiments. FC, DQ and KN interpreted the data. KN
prepared the manuscript. All authors read and approved the final
manuscript.
Ethics approval and consent to
participate
The present study was approved by the Ethics
Committee of Ethics Committee of the First Affiliated Hospital of
Xi'an Jiaotong University (Shaanxi, China). All patients signed
written informed consent.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Brenner H, Kloor M and Pox CP: Colorectal
cancer. Lancet. 383:1490–1502. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Hughes LAE, Simons CCJM, van den Brandt
PA, van Engeland M and Weijenberg MP: Lifestyle, Diet, and
colorectal cancer risk according to (Epi)genetic instability:
Current evidence and future directions of molecular pathological
epidemiology. Curr Colorectal Cancer Rep. 13:455–469. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Siegel R, DeSantis C and Jemal A:
Colorectal cancer statistics, 2014. CA Cancer J Clin. 64:104–117.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Gòmez-Lomelí P, Bravo-Cuellar A,
Hernández-Flores G, Jave-Suárez LF, Aguilar-Lemarroy A, Lerma-Díaz
JM, Domínguez-Rodríguez JR, Sánchez-Reyes K and Ortiz-Lazareno PC:
Increase of IFN-γ and TNF-α production in CD107a+NK-92 cells
co-cultured with cervical cancer cell lines pre-treated with the
HO-1 inhibitor. Cancer Cell Int. 14:1002014. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Kim TD, Lee SU, Yun S, Sun HN, Lee SH, Kim
JW, Kim HM, Park SK, Lee CW, Yoon SR, et al: Human microRNA-27a*
targets Prf1 and GzmB expression to regulate NK-cell cytotoxicity.
Blood. 118:5476–5486. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Gong J, Maki G and Klingemann H:
Characterization of a human cell line (NK-92) with phenotypical and
functional characteristics of activated natural killer cells.
Leukemia. 8:652–658. 1994.PubMed/NCBI
|
|
7
|
Wang R, Jaw JJ, Stutzman NC, Zou Z and Sun
PD: Natural killer cell-produced IFN-γ and TNF-α induce target cell
cytolysis through up-regulation of ICAM-1. J Leukocyte Biol.
91:299–309. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Schroder K, Hertzog PJ, Ravasi T and Hume
DA: Interferon-gamma: An overview of signals, mechanisms and
functions. J Leukoc Biol. 75:163–189. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Razaghi A, Owens L and Heimann K: Review
of the recombinant human interferon gamma as an immunotherapeutic:
Impacts of production platforms and glycosylation. J Biotechnol.
240:48–60. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Lin WW and Karin M: A cytokine-mediated
link between innate immunity, inflammation, and cancer. J Clin
Invest. 117:1175–1183. 2007. View
Article : Google Scholar : PubMed/NCBI
|
|
11
|
Uemura A, Takehara T, Miyagi T, Suzuki T,
Tatsumi T, Ohkawa K, Kanto T, Hiramatsu N and Hayashi N: Natural
killer cell is a major producer of interferon gamma that is
critical for the IL-12-induced-tumor effect in mice. Cancer Immunol
Immunother. 59:453–463. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Van Heel DA: Interleukin 15: Its role in
intestinal inflammation. Gut. 55:444–445. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Bahri R, Pateras IS, D'Orlando O,
Goyeneche-Patino DA, Campbell M, Polansky JK, Sandig H, Papaioannou
M, Evangelou K, Foukas PG, et al: IL-15 suppresses
colitis-associated colon carcinogenesis by inducing antitumor
immunity. Oncoimmunology. 4:e10027212015. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Tam YK, Miyagawa B, Ho VC and Klingemann
HG: Immunotherapy of malignant melanoma in a SCID mouse model using
the highly cytotoxic natural killer cell line NK-92. J Hematother.
8:281–290. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Xu LJ, Ma Q, Zhu J, Li J, Xue BX, Gao J,
Sun CY, Zang YC, Zhou YB, Yang DR and Shan YX: Combined inhibition
of JAK1,2/Stat3-PD-L1 signaling pathway suppresses the immune
escape of castration-resistant prostate cancer to NK cells in
hypoxia. Mol Med Rep. 17:8111–8120. 2018.PubMed/NCBI
|
|
16
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Doherty TM, Seder RA and Sher A: Induction
and regulation of IL-15 expression in murine macrophages. J
Immunol. 156:735–741. 1996.PubMed/NCBI
|
|
18
|
Kobayashi H, Dubois S, Sato N, Sabzevari
H, Sakai Y, Waldmann TA and Tagaya Y: Role of trans-cellular IL-15
presentation in the activation of NK cell-mediated killing, which
leads to enhanced tumor immunosurveillance. Blood. 105:721–727.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Zhang M, Yao Z, Dubois S, Ju W, Müller JR
and Waldmann TA: Interleukin-15 combined with an anti-CD40 antibody
provides enhanced therapeutic efficacy for murine models of colon
cancer. ProcNatl Acad Sci USA. 106:7513–7518. 2009. View Article : Google Scholar
|
|
20
|
Yu P, Steel J, Zhang M, Morris J and
Waldmann T: Simultaneous blockade of multiple immune system
inhibitory checkpoints enhances anti-tumor activity mediated by
interleukin-15 in a murine metastatic colon carcinoma model. Clin
Cancer Res. 16:6019–6028. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Jakobisiak M, Golab J and Lasek W:
Interleukin 15 as a promising candidate for tumor immunotherapy.
Cytokine Growth Factor Rev. 22:99–108. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Qi Z, Nie P, Secombes CJ and Zou J:
Intron-containing type I and type III IFN coexist in amphibians:
Refuting the concept that a retroposition event gave rise to type I
IFNs. J Immunol. 184:5038–5046. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Takaoka A and Yanai H: Interferon
signalling network in innate defence. Cell Microbiol. 8:907–922.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Borden EC, Sen GC, Uze G, Silverman RH,
Ransohoff RM, Foster GR and Stark GR: Interferons at age 50: Past,
current and future impact on biomedicine. Nat Rev Drug Discov.
6:975–990. 2007. View
Article : Google Scholar : PubMed/NCBI
|
|
25
|
Danial NN and Korsmeyer SJ: Cell death:
Critical control points. Cell. 116:205–219. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Crisafulli S, Pandya Y, Moolchan K and
Lavoie T: Interferon gamma: Activity and ELISA detection
comparisons. Biotechniques. 45:101–102. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Li P, Du Q, Cao Z, Guo Z, Evankovich J,
Yan W, Chang Y, Shao L, Stolz DB, Tsung A and Geller DA:
Interferon-gamma induces autophagy with growth inhibition and cell
death in human hepatocellular carcinoma (HCC) cells through
interferon-regulatory factor-1 (IRF-1). Cancer Lett. 314:213–222.
2012. View Article : Google Scholar : PubMed/NCBI
|