Introduction
Cadmium (Cd) is a heavy metal with a wide occurrence
in the environment, classified as group 1 human carcinogen since
1993 by the International Agency for Research on Cancer (IARC)
(1,2). Cadmium was described as a cumulative
toxin due to its long half-life (up to 1-3 decades) and its very
low rate of excretion (3),
parameters that are directly linked to the noxious effects on key
organs such as kidneys, liver, heart, thyroid, bones and
reproductive system (1,4-6).
In addition, long-term exposure to Cd was associated with an
increased risk for breast, lung, genitourinary, prostate, hepatic
and colon cancer (1,7,8).
Although important progress has been made towards the elucidation
of mechanisms involved in cadmium toxicity (such as oxidative
stress, endocrine disruption, interference with sulfhydryl groups,
impairment of zinc-linked cellular processes, weak mutagenicity and
poor DNA binding capacity), the molecular mechanisms underlying Cd
carcinogenic properties are not fully known and understood
(3,6,9,10). It
was fairly recently found that Cd has the ability to trigger
genomic instability through epigenetic mechanisms, involving DNA
methylation machinery (DNMT-DNA methyltransferases) (9-12).
Colorectal cancer is the fourth cause of
cancer-related mortality globally (13-15).
This disease presents a heterogeneous profile characterized by
chromosomal instability (most cases-over 85%) and high-frequency
microsatellite instability (15% of the cases). Its occurrence is
prevalent in persons with no apparent genetic predisposition or
hereditary colorectal pathologies (65%). The remaining cases are
associated with familial inherited susceptibility (13). Besides the genetic risk factors, age
and health status, environmental factors also play an important
role in the pathogenesis of colorectal cancer (14). Previous studies supported a possible
link between exposure to heavy metals such as cadmium, chromium,
mercury and arsenic and the risk for colorectal cancer (16,17). A
recent study asserted a potential molecular mechanism for the
implication of cadmium in colorectal tumor pathogenesis (1).
Copper is one of the essential metals involved in
the activity of different enzymes, either as a structural
component, or as a cofactor. Nevertheless, at high concentrations
it is a toxic compound (18).
Copper also interferes with key processes related to tumorigenesis
(progression, metastasis and reluctance to conventional treatments)
by direct or indirect mechanisms such as activation of the
oncogenic MAPK pathway (promotion), impairment of cancer cell redox
status, angiogenic activity (19),
and aberrant DNA methylation and changes in expression patterns of
DNMT genes (20,21). Moreover, elevated copper
concentrations in serum of patients represent a reliable marker for
the diagnosis of a malignancy (including colorectal cancer) in
advanced stage, with a low response to therapy. The link between
copper and colon cancer is even stronger, since copper modulates
BRAF signaling and BRAFV600E mutation, which represents
90% of all mutations encountered in colon cancer patients (18,19).
Epigenetic changes can be defined as inherited
modifications in DNA with no alterations within the sequence. They
include DNA methylation, miRNA regulation, and histone
deacetylation (22). DNA
methylation is fundamental for embryogenesis and maintenance of the
specificity of cell-lineage gene expression for life (9). Dysregulated DNA methylation induces
improper organism development, chronic pathologies and even
tumorigenesis (9,20-22).
A feature of colorectal cancer and other types of malignancies in
terms of epigenetic changes, is the decreased global DNA
methylation and hypermethylation of locus specific gene promoters
(23-25).
In light of the data mentioned above, the present
study aimed at characterizing the impact of CdCl2 and
CuSO4 aqueous solutions on DNA methylation in human
colorectal carcinoma HT-29 cells by quantifying DNA
methyltransferase (DNMT1, DNMT3A and DNMT3B) mRNA expression. In
addition, studies on cell viability and morphology, and migratory
capacity were performed.
Materials and methods
Reagents
CuCl2 and CuSO4 were acquired
from Sigma- Aldrich; Merck KGaA, as powders of analytical grade
purity. The cell culture media: McCoy's 5a Medium Modified and
supplements-fetal bovine serum (FBS), penicillin/streptomycin
antibiotic mixture were from ATCC (American Type Cell Collection),
Thermo Fisher Scientific, Inc., and Sigma-Aldrich; Merck KGaA. The
other reagents used in the present experimental design: Phosphate
saline buffer (PBS), Trypan blue, and Alamar blue were acquired
from Sigma-Aldrich; Merck KGaA, and applied as recommended by the
manufacturers.
Cell line
The in vitro experimental part of the present
study was conducted on a human colorectal carcinoma cell line-HT-29
(ATCC® HTB-38™), that was acquired as frozen vial from
ATCC.
Cell culture protocol
HT-29 cells were grown in specific cell culture
medium, McCoy's 5a modified medium (ATCC® 30-2007™)
supplemented with 10% fetal bovine serum (FBS) purchased from
Thermo Fisher Scientific, Inc., and 1% solution of antibiotic
mixture (Penicillin and Streptomycin; Sigma-Aldrich; Merck KGaA)
added to minimize the risk of contamination in the culture.
Cell viability assessment-Alamar blue
test
In order to assess the impact of the test compounds
(CdCl2 and CuSO4) on HT-29 cell viability,
the Alamar blue assay was performed. HT-29 cells were seeded in
96-well plates, 1x104 cells/well/200 µl culture medium
and were allowed to adhere to the plate for 24 h. When the cells
reached the optimal confluence, the old culture medium was replaced
with fresh culture medium 200 µl/well that contained different
concentrations of the test solutions (CdCl2 or
CuSO4) 0.05; 0.2; 1; 10 and 100 µg/ml for 24 h. The 24 h
exposure period was followed by addition of 20 µl/well of Alamar
blue reagent, incubation for 3 h at 37˚C and reading of the
absorbance at 570 and 600 nm by a xMark™ Microplate
spectrophotometer (Bio-rad). The percentage of viable cells (%) was
calculated according to the formula presented in a previous study
(26), as follows:
Percentage of viable cells
(%)={[(εOX)λ2
Aλ1-(εOX)λ1 Aλ2 of test
agent dilution]/[(εOX)λ2
A°λ1-(εOX)λ1 A°λ2 of
untreated positive growth control]} x100,
Where: εOX=molar extinction coefficient
of Alamar blue oxidized form (BLUE)
A=absorbance of test wells
A°=absorbance of positive growth control well (cells
without tested compounds)
λ1=570 nm and λ2=600 nm.
Cell morphology microscopic
evaluation
Changes in cell morphology and shape induced by
different compounds represent specific signs of cytotoxicity.
Therefore, the effect of the tested compounds on HT-29 cell
morphology was evaluated as a part of the toxicological profile.
The cells were photographed under bright field illumination of the
Olympus IX73 inverted microscope (Olympus) at 24 h post-stimulation
with the test compounds (CdCl2 or CuSO4,
0.05; 0.2; 1; 10 and 100 µg/ml) and the photos were analyzed by the
cellSens Dimensions v.1.8. software (Olympus).
Migration rate evaluated by the
Scratch assay
To evaluate the impact of the test compounds
(CdCl2 and CuSO4) on migratory capacity of
HT-29 cells, the scratch assay (also known as wound healing assay)
was performed. The protocol applied was similar to the one
described in our previous articles (27,28)
and was adapted to the present experimental conditions. A number of
2x105 HT-29 cells/well/1.5 ml culture medium were
cultured in 12-well plates. When confluence reached 90-95%, the old
medium was removed, a scratch was drawn manually using a pipette
tip (2-200 µl) and the detached cells were washed with PBS. Fresh
culture medium containing the test compounds (0.05, 0.2 and 1
µg/ml) was added to cells for a period of 24 h. Pictures were taken
at 0 and 24 h following addition of the medium containing the test
compounds, using an Olympus IX73 inverted microscope equipped with
a DP74 camera (Olympus). The scratch widths were measured at 0 and
24 h by the means of CellSense Dimension 1.17. software (Olympus).
The scratch closure/migration rate (%) was calculated with the
following formula:
Scratch closure %=[(At=0 h-At=24
h)/At=0 h] X100,
Where: At=0 h is the width of the wound
measured immediately after scratching at 0 h.
At=24 h is the width of the wound
measured after 24 h (29,30).
RNA extraction and
reverse-transcription polymerase chain reaction (RT-PCR)
Total RNA was isolated from HT-29 cells using the
Trizol reagent (Thermo Fisher Scientific, Inc.) and the Quick-RNA™
purification kit (Zymo Research). cDNA was generated by reverse
transcription with the Maxima® First Strand cDNA
Synthesis Kit (Fermentas). Quantitative real-time PCR analysis was
performed in 20 µl reactions containing Power SYBR-Green PCR Master
Mix (Thermo Fisher Scientific, Inc.)
Cycling conditions in a Quant Studio 5 real-time PCR
system (Thermo Fisher Scientific, Inc.) were: 95˚C for 10 sec
followed by 40 cycles of denaturing at 95˚C for 15 sec and
annealing and extension at 55˚C for 1 min. The following primer
pairs (Eurogentec) were used:
18S (as housekeeping gene): F:
5'GTAC-CCGT-TGAA-CCCC-ATT3', R: 5'CCAT-CCAA-TCGG-TAGT-AGCG3',
DNMT1: F: 5'ACCG-CTTC-TACT-TCCT-CGAG-GCCTA3' R:
5'GTTG-CAGT-CCTC-GTGA-ACAC-TGTGG3', DNMT3A: F:
5'CACA-CAGA-AGCA-TATC-CAGG-AGTG3' R:
5'AGTG-GACT-GGGA-AACC-AAAT-ACCC3', DNMT3B: F:
5'AATG-TGAA-TCCA-GTCA-GGAA-AGGC3' R:
5'ACTG-GATT-ACAC-TCCA-GGAA-CCGT3'.
Statistical analysis
Graph Pad Prism 8 was used for the statistical
interpretation of the data. The results were expressed as the mean
± standard deviation (SD). One-way ANOVA was applied to determine
the statistical differences followed by Dunnett's post-test
(*P<0.05; ****P<0.0001).
Results
CdCl2 and CuSO4
reduced viability of HT-29 cells in a concentration-dependent
manner
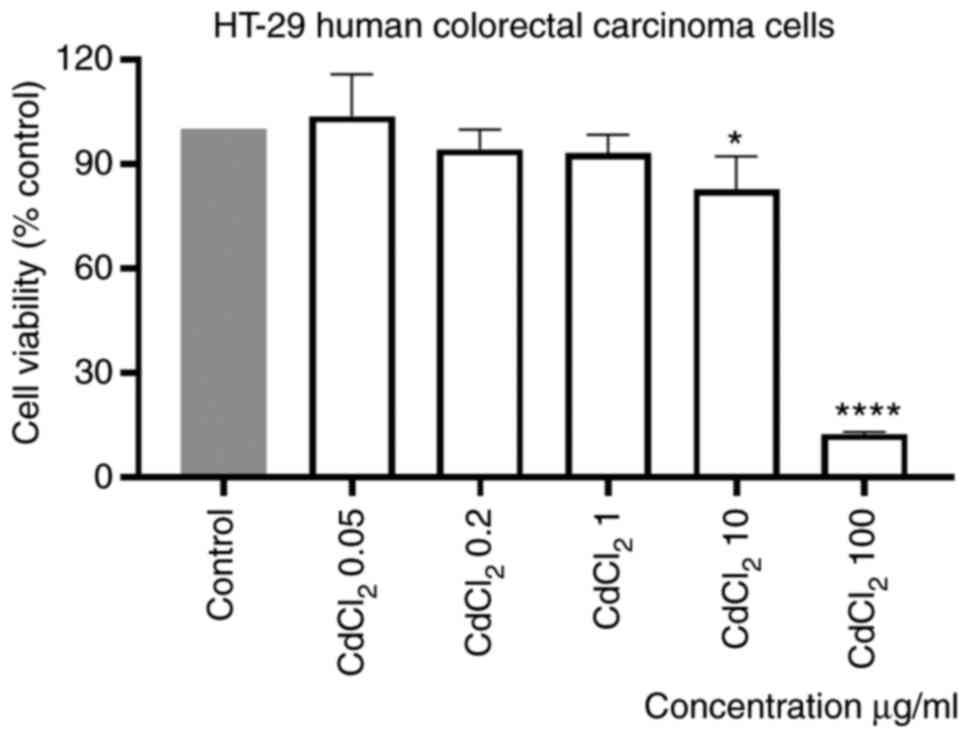
The effect of CdCl2 on the viability of
human colorectal HT-29 cells was evaluated after stimulation of
cells with different concentrations (0.05; 0.2; 1; 10 and 100
µg/ml) of the tested compound for 24 h. Our results showed that the
lowest concentration tested, 0.05 µg/ml, had a stimulatory effect
on cell viability. On the contrary, a dose-dependent cytotoxicity
was observed when the other concentrations were tested. The effect
was statistically significant only at the two highest
concentrations, 10 and 100 µg/ml, the calculated percentages of
viable cells were 83 and 13%, respectively (Fig. 1). The calculated IC50
value was 16.21 µg/ml.
Stimulation of the cells with the same volume of
sterile water did not influence the viability of the cells as
compared with control cells (unstimulated cells).
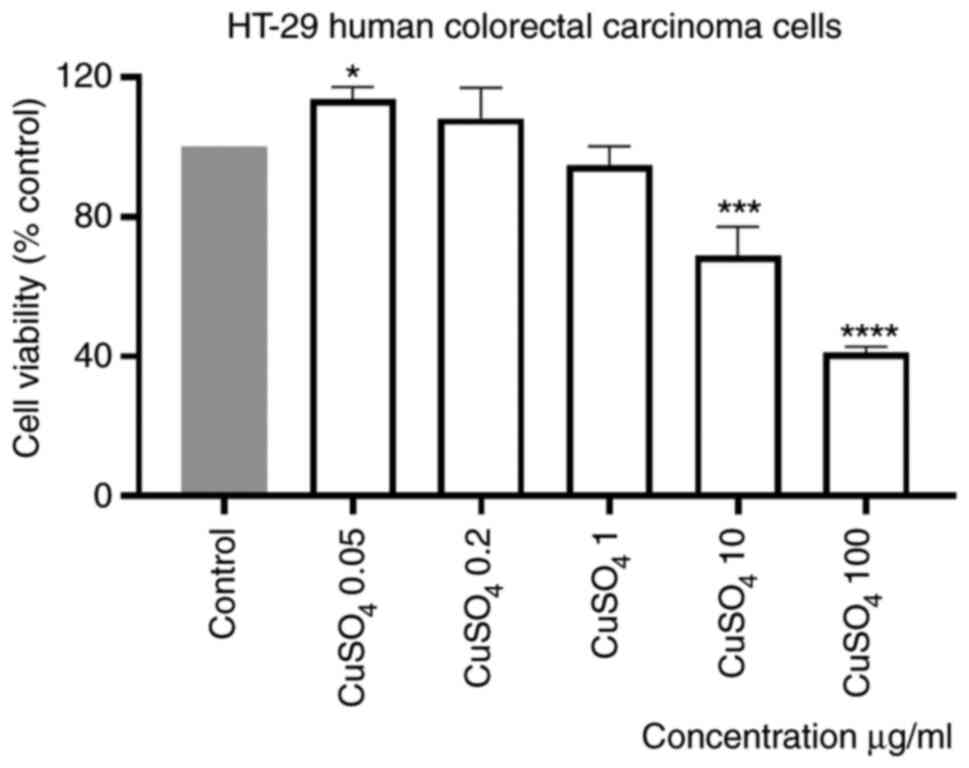
Stimulation of HT-29 cells with a CuSO4
aqueous solution (prepared at the same concentrations as for
CdCl2), induced a dose-dependent decrease of human
colorectal carcinoma cell viability percentage, compared with
control (unstimulated) cells. The effect was statistically
significant only at the two highest concentrations tested, 10 and
100 µg/ml (Fig. 2). The calculated
percentages of viable cells were 68.83 and 41.02%, respectively.
Moreover, the smallest concentrations appeared to have a
stimulatory effect (Fig. 2). The
IC50 value obtained was 4.4 µg/ml.
Effect of test compounds
(CdCl2 and CuSO4) on morphology of HT-29
cells
As shown in Fig. 3,
the solvent used for the two test solutions, sterile water, had no
impact on HT-29 cell morphology as compared with control cells
(unstimulated cells): The cells stimulated with sterile water
present the same shape as control cells, are well-attached to the
culture plate and their confluence is not affected (Fig. 3). These data are in agreement with
the results obtained for viability experiments.
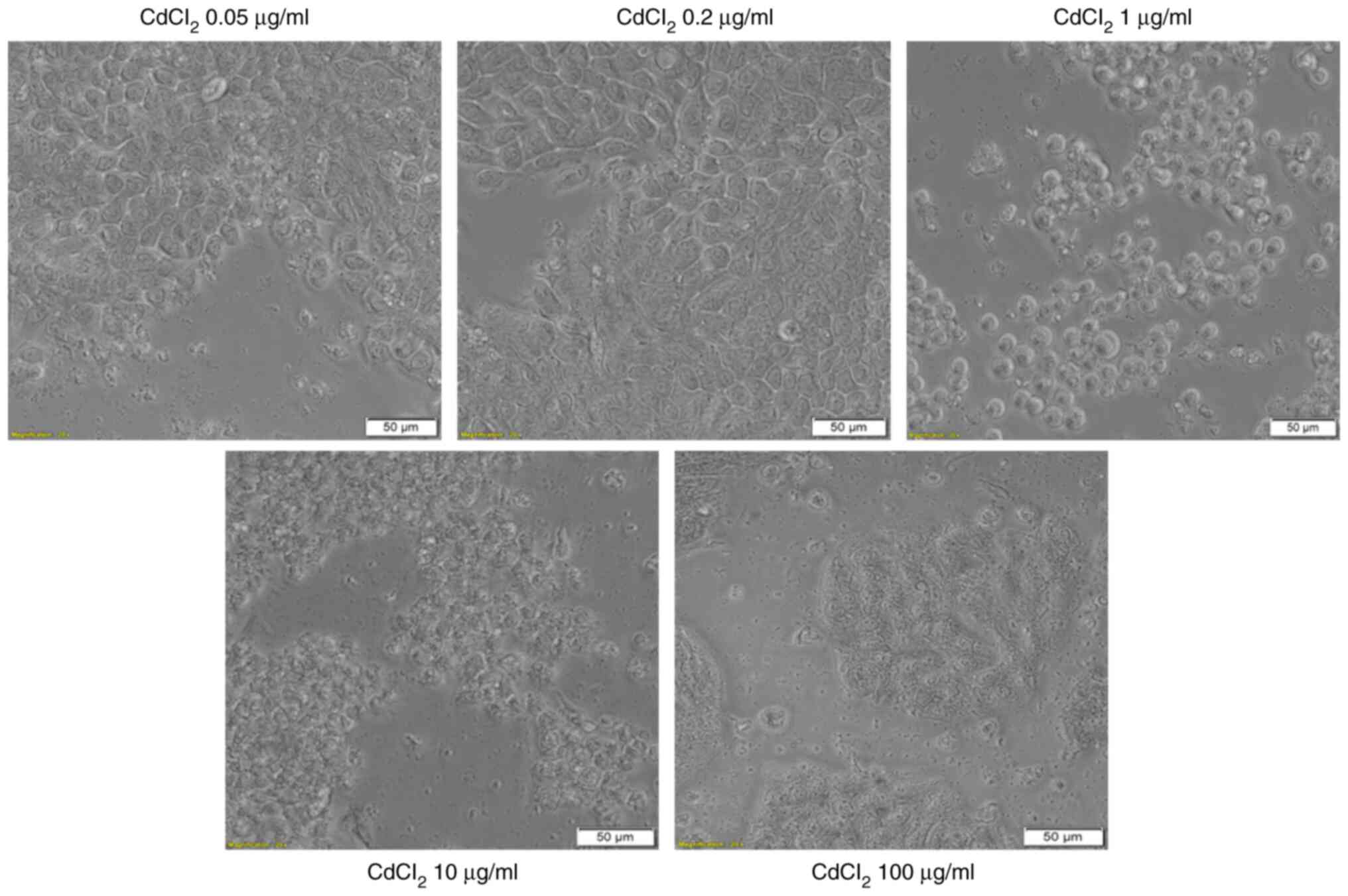
The addition of CdCl2 into the medium of
HT-29 cells and exposure of the cells to this compound for 24 h was
associated with several changes in cell morphology, compared with
control (unstimulated) cells and solvent (sterile water) treated
cells. The lowest concentrations, 0.05 and 0.2 µg/ml had no impact
on the cell shape, confluence or adherence to the culture plate. At
the concentration of 1 µg/ml the cells become round and were
floating in the medium. At the highest concentration tested, 100
µg/ml, the cells appeared disintegrated (Fig. 4-lower panel). These results
reinforce the viability data and indicate a cytotoxic effect.
CuSO4 stimulation (at concentrations of
0.05; 0.2 and 1 µg/ml) for 24 h had no impact on HT-29 cells
morphology in terms of shape, adherence or confluence as compared
with control cells or solvent-stimulated cells (Fig. 5). Several changes were observed in
the cells exposed to the highest concentrations of 10 and 100
µg/ml. These cells presented different morphology related to the
other groups of cells, round cells that floated (Fig. 5).
CdCl2 and CuSO4
stimulation interferes with the migratory capacity of HT-29
cells
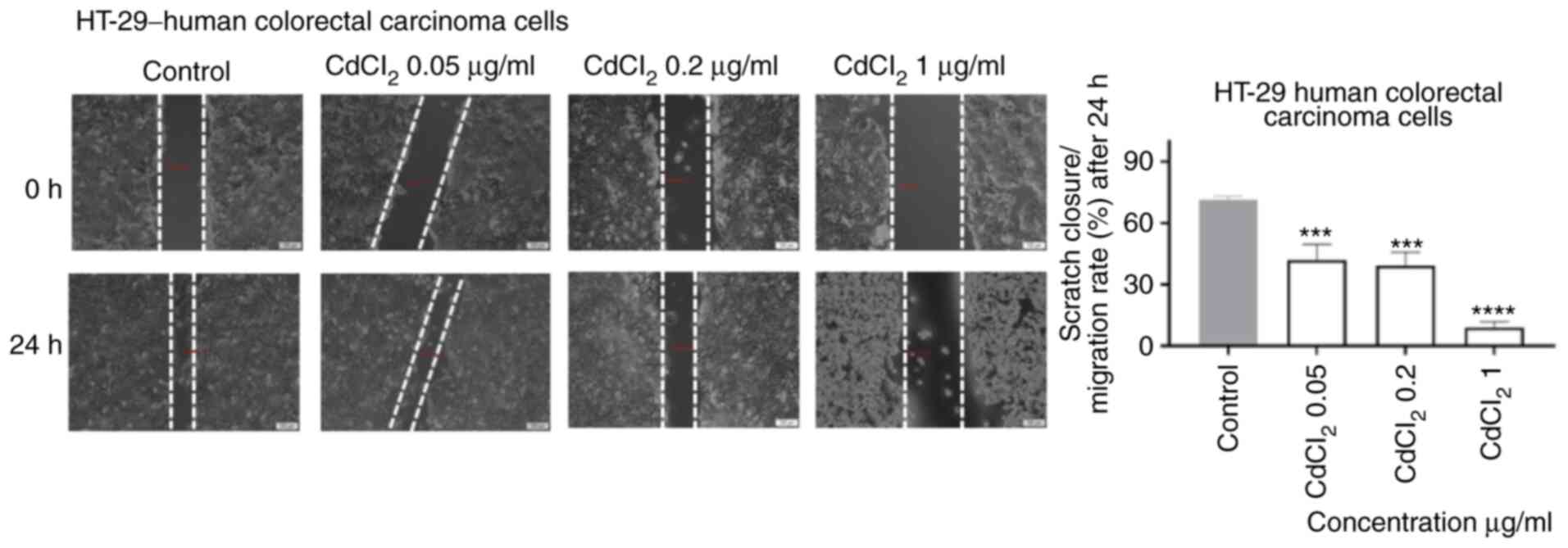
As shown in Fig. 6,
1 µg/ml of CdCl2 induced a significant inhibition of
cell migration following 24 h of exposure. Furthermore, the shape
of the cells was also different (round) as compared with control
cells. An inhibitory effect, less pronounced, was also recorded for
the other concentrations tested (Fig.
6).
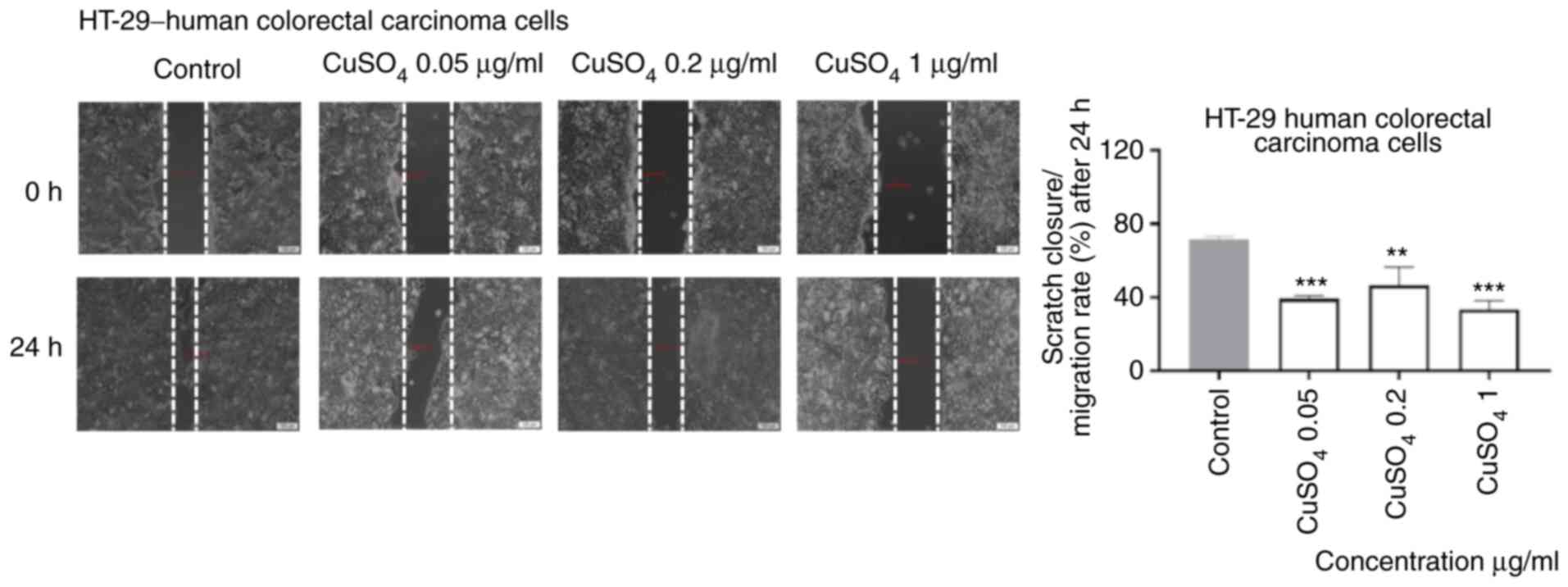
Exposure to CuSO4 (0.05, 0.2 and 1 µg/ml)
for 24 h induced a similar effect on the migratory capacity of
HT-29 cells as the one described for CdCl2, namely an
inhibitory effect even at the lowest concentration tested (Fig. 7).
CdCl2 and CuSO4
stimulation interferes with DNMT1, DNMT3A and DMNT3B gene
expression in HT-29 human colorectal carcinoma cells Cadmium
chloride
There was a significant effect on the expression of
DNMT1, DNMT3A and DNMT3B for all cadmium concentrations tested in
the present experiment (ANOVA, P<0.05).
The DNMT3A expression was significantly increased at
the two lowest concentrations tested, as compared with the
expression of DNMT1 and DNMT3B (Newman-Keuls tests, P<0.05). The
increase in CdCl2 concentration resulted in significant
changes in the expression of DNMT1, DNMT3A and DNMT3B (ANOVA,
P<0.05). Expression of DNMT1 and DNMT3B showed a similar pattern
of evolution as the concentration of CdCl2 increased.
The determined values decreased from the lowest cadmium
concentration to the concentration of 10 µg/ml and increased at the
concentration of 100 µg/ml (Table
I).
 | Table IRelative expression of DNMT1, DNMT3A
and DNMT3B in human colorectal carcinoma cells (HT-29) exposed to
CdCl2 and CuSO4 normalized to control
(unstimulated cells). |
Table I
Relative expression of DNMT1, DNMT3A
and DNMT3B in human colorectal carcinoma cells (HT-29) exposed to
CdCl2 and CuSO4 normalized to control
(unstimulated cells).
| | CdCl2
(µg/ml) | CuSO4
(µg/ml) |
|---|
| Gene | 0.05 | 1 | 10 | 100 | 0.05 | 1 | 10 | 100 |
|---|
| DNMT1 | -3.72 | -5.67 | -8.69 | -1.03 | 18.58 | 16.81 | 17.3 | 11.69 |
| | (-0.24) | (-0.64) | (-0.54) | (-0.23) | (3.83) | (3.20) | (2.93) | (1.25) |
| DNMT3A | 5.72 | 1.03 | -17.12 | -7.19 | 0 | 0 | 0 | 0 |
| | (0.45) | (0.13) | (-1.98) | (-0.87) | - | - | - | - |
| DNMT3B | -2.85 | -5.82 | -13.7 | -8.7 | 22.19 | 12.64 | 25.53 | 21.02 |
| | (-0.45) | (-0.38) | (-1.67) | (-0.63) | (2.54) | (1.45) | (2.86) | (3.45) |
Copper sulphate
In the case of copper, the expression of DNMT1
remained relatively constant up to the concentration of 10 µg/ml
but showed a significant decrease for the highest dose treatment
(ANOVA P<0.05). There was no detectable expression of DNMT3A.
However, for DNMT3B there were significant differences (ANOVA,
P<0.05). The expression decreased significantly at the second
lowest dose, compared with the lowest dose (Table I).
Discussion
Colorectal cancer presents lower mortality rates
compared with the past, due to the efficient measures implemented
(screening methods, early detection and intervention, optimized
treatments) (14). Nevertheless, it
is characterized by a poor survival rate of 13% if metastasis
occurs, with an overall median survival period of 24 months
(19,31). Several innovative methods were
developed in recent years, to gather insights into the cancer
development, diagnostic, and finding novel therapeutic approaches,
such as: Cellomics (32),
proteomics (33), and QSAR machine
learning-based models (34). The
various risk factors for colorectal cancer include heredity, age,
health status (different pathologies, obesity, inflammatory Bowel
disease and ulcerative colitis), as well as lifestyle. Similarly,
risk factors such as lifestyle (nutrition, smoking, alcohol
consumption), long-term use of contraceptives, exposure to
environmental toxicants were described in the tumorigenesis related
to human papilloma virus infection (35,36). A
novel identified risk factor for colorectal cancer is microbial
dysbiosis (biofilms) in gut microbiome (37), biofilms being also responsible for
most types of infections (~65% nosocomial infections and ~80% of
all microbial infections) (38).
Furthermore, environmental toxicants such as heavy metals,
including cadmium, play an asserted role in the pathogenesis of the
disease, according to recent studies (14,39).
The wide use of Cd in industry (refining industries,
metal mining, construction, Cd-containing batteries, and shipyard)
and its environmental dissemination (7,40-42)
led to the ranking of this metal among the most toxic ones (the
seventh most toxic heavy metal) (16). It is responsible for multiple
noxious effects such as nephrotoxicity, cardiotoxicity, bone
diseases, reproductive toxicity, inflammatory disorders and
tumorigenesis. To reduce the toxicity of the metals used for
biomedical applications, there were proposed multiple coatings,
diamond-like carbon films being a promising tool (43). The underlying molecular pathways
involved in the carcinogenic properties of cadmium are not fully
understood. The possible mechanisms include: i) Induction of
reactive oxygen species (ROS) via activation of the mitogen
activated protein kinases (MAPKs) pathway with oxidative impairment
of proteins, lipids, and DNA; ii) the estrogen-like properties that
were linked to the development of endometrial, breast and prostate
tumors; iii) initiation of inflammation via upregulation of COX-2
(cyclooxygenase-2) expression, a key player in colorectal cancer
via its major metabolite PGE2-prostaglandin
E2, iv) triggering of malignant transformation in normal
cells (human bronchial epithelial cells) by inducing apoptosis
followed by DNA impairment, reduced DNA repair capacity and genomic
instability through accumulation of mutations in DNA repair genes,
creating a suitable ground for cancer development, v) epigenetic
changes by interfering in DNA methylation, and vi) mitochondrial
toxicity both in healthy (HPNE) and tumor (AsPC-1) pancreatic cells
by altering mitochondrial function (1,2,7,9,10,44-46).
Tumor cell migration represents an essential
condition for invasion and metastasis, and numerous studies have
focused lately to identify the factors that regulate the migration
process that is directly connected to signal biomolecules of the
immune and neuroendocrine systems (47).
In a recent experimental study, Naji and colleagues
showed that exposure to low levels of CdCl2 (100 and
1,000 nM equivalent to 0.01833 and 0.1833 µg/ml, respectively) for
9 and 12 h induced an increased migratory effect on HT-29 cells by
activating several signaling pathways, such as the
ROS-p38-COX-2-PGE2 and the ROS-Akt (1). These data are in accordance with our
results that showed an increased percentage of viable cells at the
lowest concentrations tested (0.05 and 0.2 µg/ml), compared with
control cells (Fig. 1). Regarding
the effect of Cd on the migratory capacity of HT-29 cells in our
study, the longer exposure time (24 h), as compared with 9 and 12 h
reported by Naji and colleagues, induced an inhibitory effect on
HT-29 cell migration rate as compared with control cells (Fig. 6). These results are also confirmed
by the study of Naji and colleagues who found that no further
increase of wound closure was observed after 12 h (1).
An interesting finding of this work is the cytotoxic
effect of CdCl2 on human colorectal carcinoma HT-29
cells which was observed at concentrations ≥1 µg/ml. This effect
was characterized by a decrease in the viable cell percentage
(Fig. 1) and changes in cell
morphology (Fig. 4).
This novel cytotoxic property of Cd on cancer cells
was also reported by Hajrezaie et al in a study where colon
cancer HT-29 cells were exposed for 72 h to a complex of Cd, a
Schiff based complex,
CdCl2(C14H21N3O2)
that induced apoptosis via activation of mitochondrial pathway
(31). Another study that supports
our data on the cytotoxic effect of Cd on cancer cells, was
published by Guo et al who showed the antiproliferative and
proapoptotic effects of CdCl2 as a single agent, as well
as in combination with hSmac, in hepatocellular carcinoma cells
(48).
Zhou et al showed that Cd exerted its
cytotoxic effects on multiple epithelial-like cells, both healthy,
as well as of tumor origin (hepatoma cell line FLC-4, breast
carcinoma MCF7, gastric adenocarcinoma AGS, colon carcinoma HCT116,
esophageal carcinoma TE4 and embryonic kidney cell line HEK293) in
a cell-type and dose-dependent manner (49). A dose- and time-dependent cytotoxic
effect of CdCl2 was shown on MDA-MB468 breast carcinoma
cells (50), data that are in
agreement with our results.
The connection between colorectal cancer and copper
could be considered stronger, compared with cadmium, since elevated
serum concentrations of copper in patients diagnosed with
colorectal cancer indicate an advanced disease with a high risk of
mortality (19).
The role of copper in the modulation of
tumorigenesis via direct or indirect mechanisms (angiogenic
cofactor, a redox-active metal and activation of MAPK pathway) is
established. Moreover, it was shown that depletion of copper
determines apoptosis of cancer cells (19). To highlight the function of Cu in
colon cancer, different copper chelators were tested as potential
anticancer therapies (18,19).
A strong connection also lies between Cd and Cu,
since Cd is responsible for a disrupted balance of essential metals
(Zn, Cu, Ca) leading to depletion of these metals and inducing
noxious effects on human health including cancer of the intestine
(39). In our previous study,
conducted on land snails-Cantareus aspersus, we showed that
low doses of dietary cadmium interfere with hepatopancreas copper
deposition (51).
Our results on the effect of Cu on HT-29 cell
viability showed a dose-dependent cytotoxic effect and are
supported by the findings of another study that tested a
CuCl2 solution (1-1,000 µM) on MDA-MB458 breast
carcinoma cells (50). The
cytotoxic effect of a CuSO4 solution was also studied on
a human glioma cell line-U87-MG (52) and HeLa cells (53). The findings of both studies are
consistent with our results. Moreover, our experimental data
indicate an anti-migratory effect of copper on HT-29 cells
(Fig. 7).
Among the multiple mechanisms of toxicity linked to
cadmium and copper exposure, the interference with DNA methylation
has also been described (3,9-11,20,21).
DNA methylation is prevalent in the mammalian genome and is
responsible for genomic stability, chromatin structure modulation,
and transcriptional regulation of specific genes (22,25).
This process occurs at the cytosines in specific regions of CpG
dinucleotides and is catalyzed by DNA methyltransferases (DNMTs),
DNMT1, DNMT3A and DNMT3B (22,25,54).
DNMT1 is responsible for maintaining the patterns of DNA
methylation during cell division and shows an affinity for
hemi-methylated DNA strands, whereas DNMT3A and DNMT3B are
described as de novo methyltransferases with different
target sites from DNMT1 (22,25,54).
Impaired DNA methylation has been associated with carcinogenesis
(including colorectal cancer) and with the carcinogenic potential
of cadmium (3,24).
The expression of DNMT1 and DNMT3B, key players in
establishing and maintaining the DNA methylation patterns, are
known to be disrupted in the HT-29 cell line. It was found that
these genes are generally silenced in different colorectal cancer
cell lines, including the HT-29 cell line, and the consequent
disruptions in the methylation pattern contribute directly to the
process of carcinogenesis (55).
Other studies indicate that DNMT1, DNMT3A and DNMT3B are highly
expressed in colon cancer (56,57),
and the inhibition of their expression determines a reduction of
tumorigenesis process (58). With
respect to copper, scientific evidence on humans does not support
an effect of this trace metal on genomic or gene-specific DNA
methylation levels (59). However,
our results show that copper has an effect on the expression of
DNMT1 and DNMT3B, and subsequently on DNA methylation in the HT-29
cell line. Our data show that the expression of these genes is
increased in response to copper exposure over a broad range of
concentrations. This is most probably related to the dysregulated
methylation control cycle in these tumor cells, which favors their
uncontrolled growth and spreading (60). The relationship between the
concentrations of CuSO4 and changes in the expression of
the three enzymes were strikingly different; DNMT1 expression in
copper exposed cells showed a significant decrease and approached
the expression levels in control (unstimulated cells) only at the
highest CuSO4 concentration (100 µg/ml). DNMT3B
exhibited the same trend at a significantly lower CuSO4
concentration (1 µg/ml). These patterns might be related to their
different role in establishing and maintaining the DNA methylation
pattern.
The effect of cadmium on DNMT1 and DNMT3B expression
showed a similar pattern. The expression of both enzymes was
suppressed by cadmium. This finding is consistent with the results
of a previous study that showed an association between acute
cadmium exposure and decreased DNA methylation in the TRL1215 rat
liver cells. It showed that the decrease in genomic DNA methylation
levels is caused by noncompetitive inhibition of DNMT activity, a
mechanism which could also explain our results (61). In this context, it is noteworthy to
highlight the differential effect of Cu and Cd on these two genes
in terms of direction and magnitude in the changes in DNMT1 and
DNMT3B expression. Based on our results, it appears that for acute
exposure 24 h of Cu increases expression and acts as a
hypermethylating agent, while Cd suppresses expression and acts as
a hypomethylating agent in the HT-29 cells.
We have currently shown that both CdCl2
and CuSO4 solutions induced cytotoxicity on HT-29 cells
in a dose-dependent manner. The toxic effect included a decrease of
cell viability, changes in cell morphology, and anti-migratory
effects even at the lowest concentrations tested (0.05; 0.2 and 1
µg/ml). Moreover, cadmium acts as a hypomethylating agent by
suppressing DNMT expression, whereas, copper acts as a
hypermethylating compound by increasing DNMT expression. A
limitation of this study could be considered the selection of a
single time point of 24 h, but further studies are in progress to
determine the mechanistic insights of a possible connection between
the cytotoxic effects and the epigenetic changes observed.
Acknowledgements
The in vitro experiments were conducted
within the Center of Pharmaco-toxicological evaluations from the
Faculty of Pharmacy, ‘Victor Babes’ University of Medicine and
Pharmacy (Timisoara, Romania).
Funding
The present study was supported by the Executive
Agency for Higher Education, Research, Development and Innovation
Funding Institution [grant no. PN-III-P1-1.1-PD-2016-1982] and the
grant was awarded to DEC.
Availability of data and materials
All data generated or analyzed during the study are
included in this published article.
Authors' contributions
GAD, AI, DEC, CD, AMT and OC conceived and designed
the study. IoM, IaM, DEC, GAD and AT acquired the data and drafted
the manuscript. DEC, GAD, RD and LK analysed the data. DEC, AT, AI
and GAD wrote the manuscript. LK, AMT, OC and CD revised the
manuscript. All authors read and approved the final version of the
manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Naji S, Issa K, Eid A, Iratni R and Eid
AH: Cadmium induces migration of colon cancer cells: Roles of
reactive oxygen species, P38 and cyclooxygenase-2. Cell Physiol
Biochem. 52:1517–1534. 2019.PubMed/NCBI View Article : Google Scholar
|
|
2
|
Zhou Z, Wang C, Liu H, Huang Q, Wang M and
Lei Y: Cadmium induced cell apoptosis, DNA damage, decreased DNA
repair capacity, and genomic instability during malignant
transformation of human bronchial epithelial cells. Int J Med Sci.
10:1485–1496. 2013.PubMed/NCBI View Article : Google Scholar
|
|
3
|
Martinez-Zamudio R and Ha HC:
Environmental epigenetics in metal exposure. Epigenetics.
6:820–827. 2011.PubMed/NCBI View Article : Google Scholar
|
|
4
|
Reyes-Hinojosa D, Lozada-Pérez CA, Zamudio
Cuevas Y, López-Reyes A, Martínez-Nava G, Fernández-Torres J,
Olivos-Meza A, Landa-Solis C, Gutiérrez-Ruiz MC, Rojas Del Castillo
E and Martínez-Flores K: Toxicity of cadmium in musculoskeletal
diseases. Environ Toxicol Pharmacol. 72(103219)2019.PubMed/NCBI View Article : Google Scholar
|
|
5
|
Geng HX and Wang L: Cadmium: Toxic effects
on placental and embryonic development. Environ Toxicol Pharmacol.
67:102–107. 2019.PubMed/NCBI View Article : Google Scholar
|
|
6
|
Buha A, Matovic V, Antonijevic B, Bulat Z,
Curcic M, Renieri EA, Tsatsakis AM, Schweitzer A and Wallace D:
Overview of cadmium thyroid disrupting effects and mechanisms. Int
J Mol Sci. 19(1501)2018.PubMed/NCBI View Article : Google Scholar
|
|
7
|
Luevano J and Damodaran C: A review of
molecular events of cadmium-induced carcinogenesis. J Environ
Pathol Toxicol Oncol. 33:183–194. 2014.PubMed/NCBI View Article : Google Scholar
|
|
8
|
Pal D, Suman S, Kolluru V, Sears S, Das
TP, Alatassi H, Ankem MK, Freedman JH and Damodaran C: Inhibition
of autophagy prevents cadmium-induced prostate carcinogenesis. Br J
Cancer. 117:56–64. 2017.PubMed/NCBI View Article : Google Scholar
|
|
9
|
Hossain MB, Vahter M, Concha G and Broberg
K: Low-level environmental cadmium exposure is associated with DNA
hypomethylation in Argentinean women. Environ Health Perspect.
120:879–884. 2012.PubMed/NCBI View Article : Google Scholar
|
|
10
|
Zhou ZH, Lei YX and Wang CX: Analysis of
aberrant methylation in DNA repair genes during malignant
transformation of human bronchial epithelial cells induced by
cadmium. Toxicol Sci. 125:412–417. 2012.PubMed/NCBI View Article : Google Scholar
|
|
11
|
Šrut M, Drechsel V and Höckner M: Low
levels of Cd induce persisting epigenetic modifications and
acclimation mechanisms in the earthworm Lumbricus terrestris. PLoS
One. 12(e0176047)2017.PubMed/NCBI View Article : Google Scholar
|
|
12
|
Kimura T, Hosaka T, Nakanishi T and Aozasa
O: Long-term cadmium exposure enhances metallothionein-1 induction
after subsequent exposure to high concentrations of cadmium in
P1798 mouse lymphosarcoma cells. J Toxicol Sci. 44:309–316.
2019.PubMed/NCBI View Article : Google Scholar
|
|
13
|
Nguyen HT and Duong HQ: The molecular
characteristics of colorectal cancer: Implications for diagnosis
and therapy. Oncol Lett. 16:9–18. 2018.PubMed/NCBI View Article : Google Scholar
|
|
14
|
Thanikachalam K and Khan G: Colorectal
cancer and nutrition. Nutrients. 11(164)2019.PubMed/NCBI View Article : Google Scholar
|
|
15
|
Krasanakis T, Nikolouzakis TK, Sgantzos M,
Mariolis-Sapsakos T, Souglakos J, Spandidos DA, Tsitsimpikou C,
Tsatsakis A and Tsiaoussis J: Role of anabolic agents in colorectal
carcinogenesis: Myths and realities (Review). Oncol Rep.
42:2228–2244. 2019.PubMed/NCBI View Article : Google Scholar
|
|
16
|
Jaishankar M, Tseten T, Anbalagan N,
Mathew BB and Beeregowda KN: Toxicity, mechanism and health effects
of some heavy metals. Interdiscip Toxicol. 7:60–72. 2014.PubMed/NCBI View Article : Google Scholar
|
|
17
|
Tchounwou PB, Yedjou CG, Patlolla AK and
Sutton DJ: Heavy metals toxicity and the environment. Exp Suppl.
101:133–164. 2012.PubMed/NCBI View Article : Google Scholar
|
|
18
|
Fatfat M, Merhi RA, Rahal O, Stoyanovsky
DA, Zaki A, Haidar H, Kagan VE, Gali-Muhtasib H and Machaca K:
Copper chelation selectively kills colon cancer cells through redox
cycling and generation of reactive oxygen species. BMC Cancer.
14(527)2014.PubMed/NCBI View Article : Google Scholar
|
|
19
|
Baldari S, Di Rocco G, Heffern MC, Su TA,
Chang CJ and Toietta G: Effects of copper chelation on BRAFV600E
positive colon carcinoma cells. Cancers (Basel).
11(659)2019.PubMed/NCBI View Article : Google Scholar
|
|
20
|
Sun Y, Liu C, Liu Y, Hosokawa T, Saito T
and Kurasaki M: Changes in the expression of epigenetic factors
during copper-induced apoptosis in PC12 cells. J Environ Sci Health
A Tox Hazard Subst Environ Eng. 49:1023–1028. 2014.PubMed/NCBI View Article : Google Scholar
|
|
21
|
Dorts J, Falisse E, Schoofs E, Flamion E,
Kestemont P and Silvestre F: DNA methyltransferases and
stress-related genes expression in zebrafish larvae after exposure
to heat and copper during reprogramming of DNA methylation. Sci
Rep. 6(34254)2016.PubMed/NCBI View Article : Google Scholar
|
|
22
|
Wong KK, Lawrie CH and Green TM: Oncogenic
roles and inhibitors of DNMT1, DNMT3A, and DNMT3B in acute myeloid
leukaemia. Biomark Insights. 14(1177271919846454)2019.PubMed/NCBI View Article : Google Scholar
|
|
23
|
Sarabi MM and Naghibalhossaini F:
Association of DNA methyltransferases expression with global and
gene-specific DNA methylation in colorectal cancer cells. Cell
Biochem Funct. 33:427–433. 2015.PubMed/NCBI View Article : Google Scholar
|
|
24
|
Wu W, Ye S, Tan W, Zhou Y and Quan J:
Analysis of promoter methylation and epigenetic regulation of
miR-32 in colorectal cancer cells. Exp Ther Med. 17:3209–3214.
2019.PubMed/NCBI View Article : Google Scholar
|
|
25
|
Honeywell RJ, Sarkisjan D, Kristensen MH,
de Klerk DJ and Peters GJ: DNA methyltransferases expression in
normal tissues and various human cancer cell lines, xenografts and
tumors. Nucleosides Nucleotides Nucleic Acids. 37:696–708.
2018.PubMed/NCBI View Article : Google Scholar
|
|
26
|
Soica C, Oprean C, Borcan F, Danciu C,
Trandafirescu C, Coricovac D, Crăiniceanu Z, Dehelean CA and
Munteanu M: The synergistic biologic activity of oleanolic and
ursolic acids in complex with hydroxypropyl-γ-cyclodextrin.
Molecules. 19:4924–4940. 2014.PubMed/NCBI View Article : Google Scholar
|
|
27
|
Gheorgheosu D, Jung M, Ören B, Schmid T,
Dehelean C, Muntean D and Bruene B: Betulinic acid suppresses
NGAL-induced epithelial-to-mesenchymal transition in melanoma. Biol
Chem. 394:773–781. 2013.PubMed/NCBI View Article : Google Scholar
|
|
28
|
Coricovac D, Farcas C, Nica C, Pinzaru I,
Simu S, Stoian D, Soica C, Proks M, Avram S, Navolan D, et al:
Ethinylestradiol and Levonorgestrel as active agents in normal
skin, and pathological conditions induced by UVB exposure: In vitro
and in ovo assessments. Int J Mol Sci. 19(3600)2018.PubMed/NCBI View Article : Google Scholar
|
|
29
|
Felice F, Zambito Y, Belardinelli E,
Fabiano A, Santoni T and Di Stefano R: Effect of different chitosan
derivatives on in vitro scratch wound assay: A comparative study.
Int J Biol Macromol. 76:236–241. 2015.PubMed/NCBI View Article : Google Scholar
|
|
30
|
Grada A, Otero-Vinas M, Prieto-Castrillo
F, Obagi Z and Falanga V: Research techniques made simple: Analysis
of collective cell migration using the wound healing assay. J
Invest Dermatol. 137:e11–e16. 2017.PubMed/NCBI View Article : Google Scholar
|
|
31
|
Hajrezaie M, Paydar M, Looi CY,
Moghadamtousi SZ, Hassandarvish P, Salga MS, Karimian H, Shams K,
Zahedifard M, Majid NA, et al: Apoptotic effect of novel Schiff
based
CdCl2(C14H21N3O2)
complex is mediated via activation of the mitochondrial pathway in
colon cancer cells. Sci Rep. 5(9097)2015.PubMed/NCBI View Article : Google Scholar
|
|
32
|
Boda D: Cellomics as integrative omics for
cancer. Current Proteomics. 10:237–245. 2013.
|
|
33
|
Ion A, Popa IM, Papagheorghe LM, Lisievici
C, Lupu M, Voiculescu V, Caruntu C and Boda D: Proteomic approaches
to biomarker discovery in cutaneous T-cell lymphoma. Dis Markers.
2016(9602472)2016.PubMed/NCBI View Article : Google Scholar
|
|
34
|
Ancuceanu R, Dinu M, Neaga I, Laszlo FG
and Boda D: Development of QSAR machine learning-based models to
forecast the effect of substances on malignant melanoma cells.
Oncol Lett. 17:4188–4196. 2019.PubMed/NCBI View Article : Google Scholar
|
|
35
|
Boda D, Docea AO, Calina D, Ilie MA,
Caruntu C, Zurac S, Neagu M, Constantin C, Branisteanu DE,
Voiculescu V, et al: Human papilloma virus: Apprehending the link
with carcinogenesis and unveiling new research avenues (Review).
Int J Oncol. 52:637–655. 2018.PubMed/NCBI View Article : Google Scholar
|
|
36
|
Boda D, Neagu M, Constantin C, Voinescu
RN, Caruntu C, Zurac S, Spandidos DA, Drakoulis N, Tsoukalas D and
Tsatsakis AM: HPV strain distribution in patients with genital
warts in a female population sample. Oncol Lett. 12:1779–1782.
2016.PubMed/NCBI View Article : Google Scholar
|
|
37
|
Drewes JL, White JR, Dejea CM, Fathi P,
Iyadorai T, Vadivelu J, Roslani AC, Wick EC, Mongodin EF, Loke MF,
et al: High-resolution bacterial 16S rRNA gene profile
meta-analysis and biofilm status reveal common colorectal cancer
consortia. NPJ Biofilms Microbiomes. 3(34)2017.PubMed/NCBI View Article : Google Scholar
|
|
38
|
Andor B, Tucuina AAT, Berceanu-Vaduva D,
Lazureanu V, Cheveresan A and Poenaru M: Antimicrobial activity and
cytotoxic effect on gingival cells of silver nanoparticles obtained
by biosynthesis. Rev Chim (Bucharest). 70:781–783. 2019.
|
|
39
|
Klimczak M, Dziki A, Kilanowicz A, Sapota
A, Duda-Szymańska J and Daragó A: Concentrations of cadmium and
selected essential elements in malignant large intestine tissue.
Prz Gastroenterol. 11:24–29. 2016.PubMed/NCBI View Article : Google Scholar
|
|
40
|
Nedelescu M, Baconi D, Neagoe A, Iordache
V, Stan M, Constantinescu P, Ciobanu AM, Vardas AI, Vinceti M and
Tsatsakis AM: Environmental metal contamination and health impact
assessment in two industrial regions of Romania. Sci Total Environ.
580:984–995. 2017.PubMed/NCBI View Article : Google Scholar
|
|
41
|
Renieri EA, Alegakis AK, Kiriakakis M,
Vinceti M, Ozcagli E, Wilks MF and Tsatsakis AM: Cd, Pb and Hg
Biomonitoring in fish of the Mediterranean region and risk
estimations on fish consumption. Toxics. 2:417–442. 2014.
|
|
42
|
Renieri EA, Safenkova IV, Alegakis AΚ,
Slutskaya ES, Kokaraki V, Kentouri M, Dzantiev BD and Tsatsakis AM:
Cadmium, lead and mercury in muscle tissue of gilthead seabream and
seabass: Risk evaluation for consumers. Food Chem Toxicol.
124:439–449. 2019.PubMed/NCBI View Article : Google Scholar
|
|
43
|
Calenic B, Greabu M, Caruntu C, Nicolescu
MI, Moraru L, Surdu-Bob CC, Badulescu M, Anghel A, Logofatu C and
Boda D: Oral keratinocyte stem cells behavior on diamond like
carbon films. Romanian Biotechnological Lett. 21:11914–11922.
2016.
|
|
44
|
Nair AR, Degheselle O, Smeets K, Van
Kerkhove E and Cuypers A: Cadmium-induced pathologies: Where is the
oxidative balance lost (or Not)? Int J Mol Sci. 14:6116–6143.
2013.PubMed/NCBI View Article : Google Scholar
|
|
45
|
Wallace DR, Spandidos DA, Tsatsakis A,
Schweitzer A, Djordjevic V and Djordjevic AB: Potential interaction
of cadmium chloride with pancreatic mitochondria: Implications for
pancreatic cancer. Int J Mol Med. 44:145–156. 2019.PubMed/NCBI View Article : Google Scholar
|
|
46
|
Yao M, Kargman S, Lam EC, Kelly CR, Zheng
Y, Luk P, Kwong E, Evans JF and Wolfe MM: Inhibition of
Cyclooxygenase-2 by Rofecoxib attenuates the growth and metastatic
potential of colorectal carcinoma in mice. Cancer Res. 63:586–592.
2003.PubMed/NCBI
|
|
47
|
Solomon I, Voiculescu VM, Caruntu C, Lupu
M, Popa A, Ilie MA, Albulescu R, Caruntu A, Tanase C, Constantin C,
et al: Neuroendocrine factors and head and neck squamous cell
carcinoma: An affair to remember. Dis Markers.
2018(9787831)2018.PubMed/NCBI View Article : Google Scholar
|
|
48
|
Guo C, Li Y, Zhang H, Wang Z, Jin M, Zhang
L, An L, Hu G, Liu X, Liu Y, et al: Enhancement of
antiproliferative and proapoptotic effects of cadmium chloride
combined with hSmac in hepatocellular carcinoma cells.
Chemotherapy. 57:27–34. 2011.PubMed/NCBI View Article : Google Scholar
|
|
49
|
Zhou X, Koizumi Y, Zhang M, Natsui M,
Koyota S, Yamada M, Kondo Y, Hamada F and Sugiyama T:
Cadmium-coordinated supramolecule suppresses tumor growth of T-cell
leukemia in mice. Cancer Sci. 106:635–641. 2015.PubMed/NCBI View Article : Google Scholar
|
|
50
|
Panjehpour M, Taher MA and Bayesteh M: The
growth inhibitory effects of cadmium and copper on the MDA-MB468
human breast cancer cells. J Res Med Sci. 15:279–286.
2010.PubMed/NCBI
|
|
51
|
Nica DV, Draghici GA, Andrica FM, Popescu
S, Coricovac DE, Dehelean CA, Gergen II, Kovatsi L, Coleman MD and
Tsatsakis AM: Short-term effects of very low dose cadmium feeding
on copper, manganese and iron homeostasis: A gastropod perspective.
Environ Toxicol Pharmacol. 65:9–13. 2019.PubMed/NCBI View Article : Google Scholar
|
|
52
|
Li Y, Hu J, Guan F, Song L, Fan R, Zhu H,
Hu X, Shen E and Yang B: Copper induces cellular senescence in
human glioblastoma multiforme cells through downregulation of
Bmi-1. Oncol Rep. 29:1805–1810. 2013.PubMed/NCBI View Article : Google Scholar
|
|
53
|
Chen SY, Liu ST, Lin WR, Lin CK and Huang
SM: The mechanisms underlying the cytotoxic effects of copper via
differentiated embryonic chondrocyte gene 1. Int J Mol Sci.
20(5225)2019.PubMed/NCBI View Article : Google Scholar
|
|
54
|
Fragou D, Fragou A, Kouidou S, Njau S and
Kovatsi L: Epigenetic mechanisms in metal toxicity. Toxicol Mech
Methods. 21:343–352. 2011.PubMed/NCBI View Article : Google Scholar
|
|
55
|
Jin B, Yao B, Li JL, Fields CR, Delmas AL,
Liu C and Robertson D: DNMT1 and DNMT3B modulate distinct
polycomb-mediated histone modifications in colon cancer. Cancer
Res. 69:7412–7421. 2009.PubMed/NCBI View Article : Google Scholar
|
|
56
|
Subramaniam D, Thombre R, Dhar A and Anant
S: DNA methyltransferases: A novel target for prevention and
therapy. Front Oncol. 4(80)2014.PubMed/NCBI View Article : Google Scholar
|
|
57
|
Fouad MA, Salem SE, Hussein MM, Zekri ARN,
Hafez HF, El Desouky ED and Shouman SA: Impact of global DNA
methylation in treatment outcome of colorectal cancer patients.
Front Pharmacol. 9(1173)2018.PubMed/NCBI View Article : Google Scholar
|
|
58
|
Li S, Han Z, Zhao N, Zhu B, Zhang Q, Yang
X, Sheng D, Hou J, Guo S, Wei L and Zhang L: Inhibition of DNMT
suppresses the stemness of colorectal cancer cells through
down-regulating Wnt signaling pathway. Cell Signal. 47:79–87.
2018.PubMed/NCBI View Article : Google Scholar
|
|
59
|
Ryu HW, Lee DH, Won HR, Kim KH, Seong YJ
and Kwon SH: Influence of toxicologically relevant metals on human
epigenetic regulation. Toxicol Res. 31:1–9. 2015.PubMed/NCBI View Article : Google Scholar
|
|
60
|
Ogoshi K, Hashimoto S, Nakatani Y, Qu W,
Oshima K, Tokunaga K, Sugano S, Hattori M, Morishita S and
Matsushima K: Genome-wide profiling of DNA methylation in human
cancer cells. Genomics. 98:280–287. 2011.PubMed/NCBI View Article : Google Scholar
|
|
61
|
Takiguchi M, Achanzar WE, Qu W, Li G and
Waalkes MP: Effects of cadmium on DNA-(Cytosine-5)
methyltransferase activity and DNA methylation status during
cadmium-induced cellular transformation. Exp Cell Res. 286:355–365.
2003.PubMed/NCBI View Article : Google Scholar
|