Introduction
Breast cancer is one of the leading causes of
morbidity and mortality among women. The incidence of breast cancer
has been increasing annually and it poses a major threat to the
survival and quality of life of the patients (1). The three most common types of cancer
in women include breast, lung and colorectal cancer, accounting for
50% of all new cancer diagnoses. Breast cancer alone accounts for
30% of all cancers in women, and its incidence is rising at a
modest rate of ~0.3% annually (2).
In 2015, breast cancer was the most common cancer diagnosed among
Chinese women aged 30-59 years, and it was the main cause of
cancer-related mortality among women aged <45 years in China.
Furthermore, the morbidity and mortality rates of breast cancer in
China are continuously increasing (3). Therefore, continued research is
crucial for identifying effective therapeutic targets for patients
with breast cancer.
Nuclear factor of activated T cells (NFAT), a
transcription factor found in T lymphocytes, was originally
considered to be a gene regulating T-cell activation (4). The NFAT family consists of multiple
members, including NFATc1 (also known as NFAT2), NFATc2 (NFAT1),
NFATc3 (NFAT4) and NFATc4 (NFAT3), which are located in the
cytoplasm of immune cells in a highly phosphorylated state
(5). Once the cell is stimulated,
NFATc1 responds to Ca2+-calmodulin signaling by
dephosphorylation, and nuclear translocation occurs simultaneously
(6). Accumulating evidence has
shown that the activation of NFAT signaling does not only promote
the occurrence and development of hematological malignancies, but
can also accelerate the development of solid tumors. High
expression of NFATc1 has been observed in lung, liver and
pancreatic cancer (7,8). A recent study also reported that
NFATc1 was found to be highly expressed in breast cancer, and that
NFATc1 knockdown reduced the proliferation of tumor cells and
promoted their apoptosis, which may be due to the effect of the Ras
homolog family member A/Rho-associated protein kinase pathway
(9).
RNA-binding protein (RBP) is a general term for
ubiquitous proteins that can bind to RNA. RBPs bind specifically to
RNA and directly or indirectly regulate its function (10). The ENCORI database (http://starbase.sysu.edu.cn/index.php)
predicted that CUGBP Elav-like family member 2 (CELF2) is an RBP
targeting NFATc1 and that it may exert a regulatory effect on its
expression. The expression of CELF2 is low in breast cancer due to
epigenetic changes, and CELF2 has been shown to possess anticancer
properties and inhibit tumor progression (11). However, studies and experimental
verification of its relevant mechanisms of action are currently
lacking.
The aim of the present study was to investigate the
mechanism underlying the anticancer effects of CELF2, and
preliminarily evaluate the underlying mechanism by observing the
regulatory association between this RBP and the NFAT pathway in
breast cancer.
Materials and methods
Cell culture and transfection
The MCF-10A mammary epithelial cells, the BT-20,
T47D, MCF-7 and BT-549 breast cancer cell lines, and HUVECs, were
provided by the American Type Culture Collection.
MCF-10A, BT-20, T47D, MCF-7 and BT-549 cells were
cultured, digested and passaged in 90% RPMI-1640 medium
supplemented with 10% FBS (both from Thermo Fisher Scientific,
Inc.). MCF-7 cells at the logarithmic growth phase were uniformly
inoculated into a 6-well plate. When the cell confluence had
reached ~80%, cells were transfected with empty vector (negative
control; NC; 60 nM), OverExp-CELF2 (60 nM) and OverExp-NFATc1 (60
nM) (all from Shanghai GenePharma Co., Ltd.) using
Lipofectamine® 2000 (Thermo Fisher Scientific, Inc.) at
37˚C for 48 h, according to the manufacturer's instructions.
Serum-free RPMI-1640 culture medium was used during transfection.
Normal culture medium was replaced with RPMI-1640 containing 10%
FBS at 6 h after transfection, and cells were used for subsequent
experiments 48 h after transfection.
HUVECs were cultured in endothelial cell medium
(ECM; ScienCell Research Laboratories, Inc.). When the cell
confluence had reached ~80%, HUVECs were transfected with empty
vector (NC; 60 nM), OverExp-CELF2 (60 nM) and OverExp-NFATc1 (60
nM) using Lipofectamine® 2000 (Thermo Fisher Scientific,
Inc.) at 37˚C for 48 h, according to the manufacturer's
instructions.
Reverse transcription-quantitative
(RT-q) PCR analysis
A total of ~1x105 MCF-7 cells per
well were evenly inoculated into 6-well plates and cultured for 24
h. Total RNA was extracted from the cells using TRIzol®
reagent (Thermo Fisher Scientific, Inc.) and reverse-transcribed
into cDNA using a PrimeScript RT Reagent kit (Takara Biotechnology
Co., Ltd.) according to the manufacturer's instructions. Next, a
TaqMan Universal PCR Master Mix kit (Thermo Fisher Scientific,
Inc.) was used to conduct qPCR to determine the relative expression
of CELF2 and NFATc1. The thermocycling conditions were as follows:
Initial denaturation at 95˚C for 30 sec, followed by 40 cycles at
95˚C for 25 sec, 60˚C for 35 sec and 72˚C for 35 sec. The following
primer pairs were used for the qPCR: CELF2 forward,
5'-CTGGCGGGAAACAAACTCTG-3' and reverse, 5'-TCTAAGCCCTTGGCCTCCTC-3';
NFATc1 forward, 5'-CCACCGAGCCCACTACGAGA-3' and reverse,
5'-CAGGATTCCGGCACAGTCAAT-3'; GAPDH forward,
5'-AAGGTGAAGGTCGGAGTCAAC-3' and reverse,
5'-GGGGTCATTGATGGCAACAATA-3'. mRNA levels were quantified using the
2-ΔΔCq method and normalized to the internal reference
gene GAPDH (12).
Xenograft model
MCF-7 cells cultured for 6 days were collected and a
1x107/ml cell suspension was created using PBS. A total
of 15 female BALB/c nude mice (age, 6-8 weeks; weight, 18-20 g)
were housed in an environmentally controlled room (22±2˚C; 12-h
light/dark cycle; 50-65% humidity) and were given free access to
food and water. After being allowed to acclimate for 1 week, each
mouse was subcutaneously injected with 1x106 cells into
the right armpit. Mouse weight and tumor diameter were measured
once every 3 days after injection. The maximum tumor size obtained
was <1,000 mm3. The experiment finished after 3
weeks, and the mice were euthanized with intraperitoneal injection
of 150 mg/kg pentobarbital sodium and the tumors were resected. The
animal experimental protocols were approved by the Animal Ethics
Committee of Jilin Cancer Hospital (approval no.
JLCH2020-0052).
H&E staining
The tumor tissues were fixed in 10% neutral methylal
for 24 h at room temperature and washed fully with flowing water.
Following dehydration with 70, 80, 90, 95 and 100% ethanol, the
tissues were transparentized with xylene, embedded in paraffin, cut
into 4-µm sections and stained with H&E for 3 min at room
temperature. The sections were then observed and images were
captured using a light microscope (Olympus Corporation) at the
magnification of x400.
Western blot analysis
The lysis products were extracted from tumor tissues
and transfected cells using RIPA lysis buffer (Beyotime Institute
of Biotechnology), and the protein concentration was measured with
a BCA protein assay kit. After high-temperature denaturation, the
proteins (30 µg/lane) were loaded and separated via 12% SDS-PAGE
and transferred to a PVDF membrane. The membrane was then blocked
with 5% skimmed milk powder for 2 h at room temperature, and
incubated with antibodies against N-cadherin (N-cad; cat. no.
ab76011; dilution, 1:5,000; Abcam), CD34 (cat. no. ab81289;
dilution, 1:10,000; Abcam), NFATc1 (cat. no. ab124292; dilution,
1:5,000; Abcam) and GAPDH (cat. no. ab8245; dilution, 1:5,000;
Abcam) overnight at 4˚C. Subsequently, the membrane was incubated
with anti-rabbit HRP-linked IgG secondary antibody (1:1,000; cat.
no. 7074) or anti-mouse HRP-linked IgG secondary antibody (1:1,000;
cat. no. 7076) (both from Cell Signaling Technology, Inc.) at room
temperature for 2 h. ECL solution (MilliporeSigma) was used for
color development. Then, protein bands were examined using Image
Lab™ software (cat. no. 1709690; Bio-Rad Laboratories, Inc.) and
ImageJ software (v1.8.0.112; National Institutes of Health) was
used for semi-quantitative protein analysis.
RNA pull-down assay
Briefly, HUVECs were lysed with 500 µl RIPA lysis
buffer (Beyotime Institute of Biotechnology) and incubated with
biotinylated NFATc1 probes (Shanghai GenePharma Co., Ltd.) at 4˚C
for 2 h. Then, 50 µl streptavidin magnetic beads (Thermo Fisher
Scientific, Inc.) were added to each sample, which was incubated at
4˚C for 2 h. The beads were washed with lysis buffer and the
binding proteins in the pull-down products were collected for use
in the western blot analysis as aforementioned (13).
Cell Counting Kit (CCK)-8 assay
MCF-7 cells (4,000 cells/well) and HUVECs
(1x105 cells/well) were inoculated into a 96-well plate
and cultured for 24 h after transfection. Then, 48 h after culture,
10 µl CCK-8 solution (Beyotime Institute of Biotechnology) was
added to each well and incubated at 37˚C for 1 h. The optical
density at 450 nm was determined using a microplate reader (Thermo
Fisher Scientific, Inc.).
Transwell assay
The matrix glue was solidified in a cell incubator
for 1 h after diluting with 50 µl basic glue in a small chamber.
After transfection for 24 h, MCF-7 cells at the logarithmic growth
phase were evenly inoculated into the upper chamber of a 24-well
Transwell chamber with RPMI-1640 medium at a density of
5x104 cells/well and cultured for 24 h at 37˚C. The
lower chamber contained RPMI-1640 medium with 10% FBS. The cells in
the upper surface of the filter (8-µm pore size) were removed with
a cotton swab, and the cells invading to the lower surface of the
filter were fixed with 4% formaldehyde for 30 min at room
temperature and stained with 0.2% crystal violet solution for 60
min at room temperature and counted at least six random microscopic
fields under a light microscope (Olympus Corporation;
magnification, x100).
HUVEC tube formation assay
HUVECs (5x104/well) were added into 300
µl ECM and seeded on a 48-well plate coated with 300 µl
Matrigel/well. After 2 days of incubation at 37˚C, tube formation
in the Matrigel was observed under a phase-contrast microscope
(magnification, x4).
Statistical analysis
SPSS 20.0 software (IBM Corp.) was used for
analysis, and the quantitative data of normal distribution are
presented as the mean ± SD from three experimental repeats. An
unpaired t-test and one-way ANOVA with Tukey's post hoc test were
used for comparison between two and multiple groups, respectively.
P<0.05 was considered to indicate a statistically significant
difference.
Results
CELF2 expression is decreased in
breast cancer cells
The protein expression level of CELF2 was found to
be decreased in BT-20, MCF-7 and BT-549 cells, but was not notably
different in T47D cells, compared with MCF-10A cells. Moreover,
CELF2 expression was the lowest in MCF-7 cells; therefore, MCF-7
cells were selected for the subsequent experiments (Fig. 1A). After MCF-7 cells were
transfected with OverExp-CELF2-#1/2, CELF2 expression markedly
increased. It was found that CELF2 protein expression (Fig. 1B) and CELF2 mRNA expression
(Fig. 1C) were both higher in the
OverExp-CELF2-#1 group; thus, OverExp-CELF2-#1 was selected for
subsequent experiments.
CELF2 overexpression inhibits tumor
growth and angiogenesis
Macroscopic images of the mice and tumors in the
three groups are presented in Fig.
2A. CELF2 overexpression led to weight loss in the mice
(Fig. 2B) and decrease in tumor
volume from day 0 to day 21 (Fig.
2C). At the end of the experiment, tumor weight (Fig. 2D) and tumor volume (Fig. 2E) were decreased in the
OverExp-CELF2 group. Furthermore, CELF2 overexpression decreased
the number of microvessels in tumor tissues (Fig. 2F). The expression levels of N-cad,
CD34 (angiogenesis-promoting factor) and NFATc1 were significantly
reduced in the OverExp-CELF2 group (Fig. 2G).
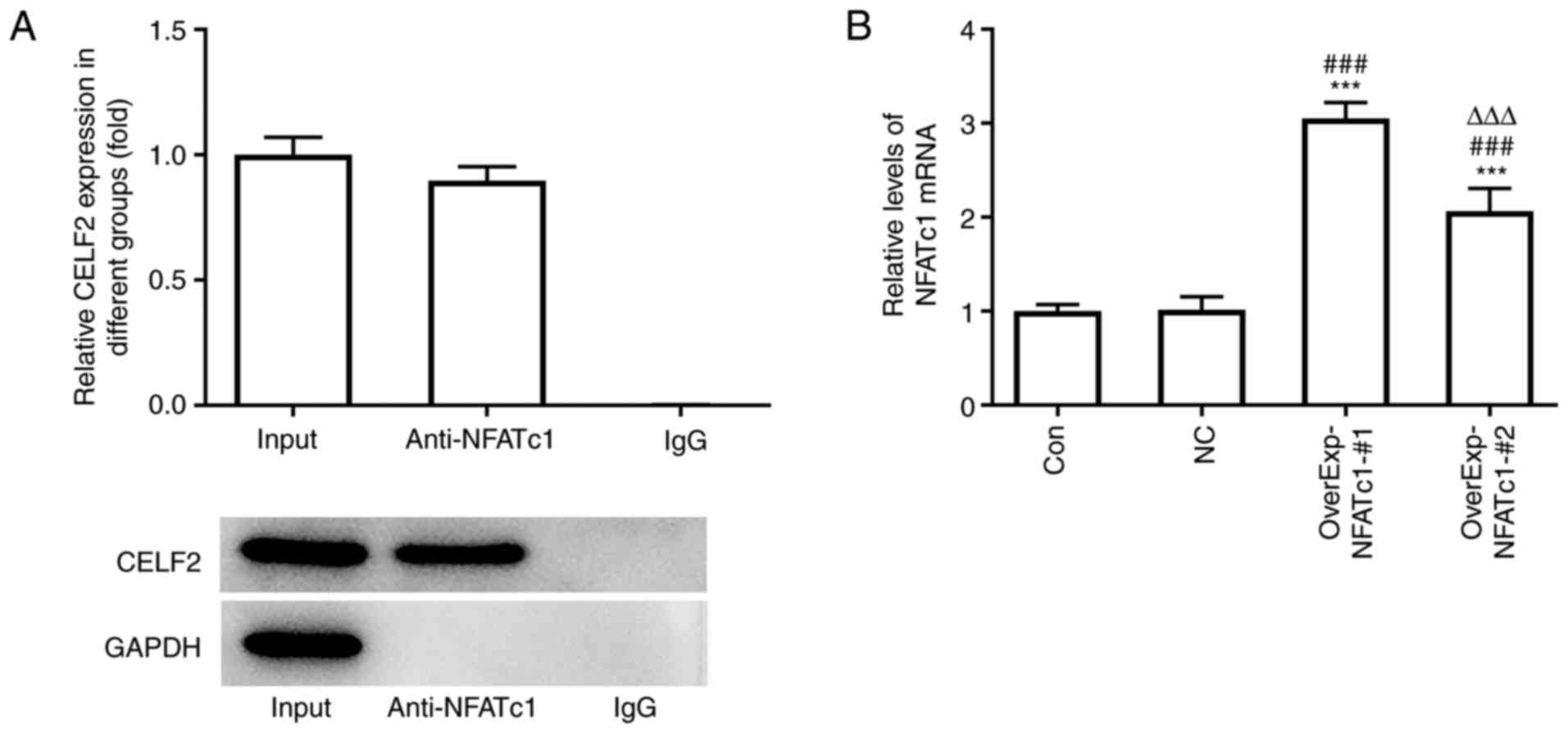
CELF2 is combined with NFATc1
The RNA pull-down assay results demonstrated the
association between CELF2 and NFATc1 (Fig. 3A). NFATc1 expression was increased
in MCF-7 cells transfected with OverExp-NFATc1-#1/2, and NFATc1
expression was highest in the OverExp-NFATc1-#1 group. Therefore,
OverExp-NFATc1-#1 was selected for the subsequent experiments
(Fig. 3B).
CELF2 overexpression suppresses the
viability and invasion of breast cancer cells by downregulating
NFATc1
CELF2 overexpression decreased the viability
(Fig. 4A) and invasion ability
(Fig. 4B and C) of MCF-7 cells, and these effects were
reversed by NEAFc1 overexpression. Moreover, CELF2 overexpression
decreased the expression levels of N-cad and NFATc1, which was
reversed by NEAFc1 overexpression (Fig.
4D).
CELF2 overexpression suppresses HUVEC
tube formation by downregulating NFATc1
CELF2 overexpression suppressed the viability and
angiogenesis of HUVECs, and NEAFc1 overexpression reversed the
effects of CELF2 overexpression (Fig.
5A and B). It was also found
that the expression levels of CD34 and NFATc1 in HUVECs were
suppressed by CELF2 overexpression, which was reversed by NEAFc1
overexpression (Fig. 5C).
Discussion
Breast cancer is a heterogeneous disease and is one
of the leading causes of mortality among women worldwide (14). It has been reported that there are
1.67 million new cases of breast cancer annually, accounting for
11.88% of all cancer cases (15).
Although a variety of risk factors, such as family history,
exogenous hormones, obesity and reproductive factors, have been
identified for breast cancer, early cancer detection and further
improvement of treatment outcomes remain significant challenges for
clinicians (16-18).
The RBP CELF2 was initially identified in
colchicine-induced neuroblastoma cells (19). CELF2 expression is downregulated in
colon tumor tissues and may be a potential tumor suppressor protein
(20), and restoration of CELF2
expression inhibits the proliferation of breast cancer cells
(11). MicroRNA (miR)-615-3p, which
promotes gastric cancer cell proliferation and migration by
downregulating CELF2 expression, has been found to be highly
expressed, while CELF2 is underexpressed, in gastric cancer cells
and tumor tissues (21). It has
also been reported that CELF2 expression is decreased in non-small
cell lung carcinoma (NSCLC) tissues, and that CELF2 overexpression
suppresses both Akt phosphorylation and cell proliferation in NSCLC
(22). In the present study, it was
observed that CELF2 expression was decreased in MCF-7 cells, and
CELF2 overexpression suppressed the viability and invasion of MCF-7
cells, as well as tumor growth and angiogenesis.
As a transcription factor, NFATc1 serves an
important role in tumors. Overexpression of NFATC1 promotes the
proliferation of ovarian cancer cells and the occurrence of tumors
by regulating the ERK1/2/P38 MAPK signaling pathway (23). Furthermore, activation of NFATc1 may
upregulate cyclooxygenase-2 expression, thus promoting the invasion
of human glioma cells (24). It has
also been demonstrated that inhibition of NFATc1 may suppress the
proliferative, migratory and invasive abilities of prostate cancer
cells, possibly by decreasing the expression of c-myc and pyruvate
kinase isozymes M1/M2(25). NFATc1
is highly expressed in human serous/mucinous ovarian cancer and
downregulation of NFATc1 suppresses cell cycle progression,
invasion and migration, and promotes the apoptosis of ovarian
cancer cells (26). In the present
study, after MCF-7 cells were co-transfected with OverExp-CELF2 and
OverExp-NFATc1, it was observed that NFATc1 overexpression weakened
the effect of CELF2 overexpression and promoted the viability and
invasion of MCF-7 cells. In addition, after HUVECs were
co-transfected with OverExp-CELF2 and OverExp-NFATc1, it was
demonstrated that NFATc1 overexpression weakened the effect of
CELF2 overexpression and promoted the viability and angiogenesis of
HUVECs.
In conclusion, the results of the present revealed
that CELF2 expression is downregulated in breast cancer cells and
that CELF2 may inhibit breast cancer cell invasion and angiogenesis
by downregulating the expression of NFATc1. Moreover, NFATc1
overexpression may partially reverse the effects of CELF2
overexpression on breast cancer cells.
Acknowledgements
Not applicable.
Funding
Funding: No funding was received.
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
XX conceived the study and revised the manuscript.
LZ performed the experiments, data analysis, organized the figures
and was responsible for the draft manuscript. LZ and XX confirm the
authenticity of all the raw data. Both authors have read and
approved the final manuscript.
Ethics approval and consent to
participate
The animal experimental protocols were approved by
the Animal Ethics Committee of Jilin Cancer Hospital (approval no.
JLCH2020-0052).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Harbeck N and Gnant M: Breast cancer.
Lancet. 389:1134–1150. 2017.PubMed/NCBI View Article : Google Scholar
|
|
2
|
Siegel RL, Miller KD and Jemal A: Cancer
statistics, 2020. CA Cancer J Clin. 70:7–30. 2020.PubMed/NCBI View Article : Google Scholar
|
|
3
|
Chen W, Zheng R, Baade P, Zhang S, Zeng H,
Bray F, Jemal A, Yu X and He J: Cancer statistics in China, 2015.
CA Cancer J Clin. 66:115–132. 2016.PubMed/NCBI View Article : Google Scholar
|
|
4
|
Irnaten M, Zhdanov A, Brennan D, Crotty T,
Clark A, Papkovsky D and O'Brien C: Activation of the NFAT-calcium
signaling pathway in human lamina cribrosa cells in glaucoma.
Invest Ophthalmol Vis Sci. 59:831–842. 2018.PubMed/NCBI View Article : Google Scholar
|
|
5
|
Lu WC, Xie H, Tie XX, Wang R, Wu AH and
Shan FP: NFAT-1 hyper-activation by methionine enkephalin (MENK)
significantly induces cell apoptosis of rats C6 glioma in vivo and
in vitro. Int Immunopharmacol. 56:1–8. 2018.PubMed/NCBI View Article : Google Scholar
|
|
6
|
He RL, Wu ZJ, Liu XR, Gui LX, Wang RX and
Lin MJ: Calcineurin/NFAT signaling modulates pulmonary artery
smooth muscle cell proliferation, migration and apoptosis in
monocrotaline-induced pulmonary arterial hypertension rats. Cell
Physiol Biochem. 49:172–189. 2018.PubMed/NCBI View Article : Google Scholar
|
|
7
|
Müller MR and Rao A: NFAT, immunity and
cancer: A transcription factor comes of age. Nat Rev Immunol.
10:645–656. 2010.PubMed/NCBI View
Article : Google Scholar
|
|
8
|
Ouyang Z, Guo X, Chen X, Liu B, Zhang Q,
Yin Z, Zhai Z, Qu X, Liu X, Peng D, et al: Hypericin targets
osteoclast and prevents breast cancer-induced bone metastasis via
NFATc1 signaling pathway. Oncotarget. 9:1868–1884. 2018.PubMed/NCBI View Article : Google Scholar
|
|
9
|
Liu M, Wei L, Wang Y, Liu P, Liu X and
Zhang M: Effects of NFATc1 on proliferation and apoptosis of breast
cancer cells by regulating RhoA /ROCK signaling pathway. Mod Oncol.
28:562–568. 2020.
|
|
10
|
He Y and Ding M: Research progress on the
interaction between RNA binding proteins and RNA-proteins. Sci
Technol Innovation. 10–13. 2020.
|
|
11
|
Piqué L, Martinez de Paz A, Piñeyro D,
Martínez-Cardús A, Castro de Moura M, Llinàs-Arias P, Setien F,
Gomez-Miragaya J, Gonzalez-Suarez E, Sigurdsson S, et al:
Epigenetic inactivation of the splicing RNA-binding protein CELF2
in human breast cancer. Oncogene. 38:7106–7112. 2019.PubMed/NCBI View Article : Google Scholar
|
|
12
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408.
2001.PubMed/NCBI View Article : Google Scholar
|
|
13
|
Dou YQ, Kong P, Li CL, Sun HX, Li WW, Yu
Y, Nie L, Zhao LL, Miao SB, Li XK, et al: Smooth muscle SIRT1
reprograms endothelial cells to suppress angiogenesis after
ischemia. Theranostics. 10:1197–1212. 2020.PubMed/NCBI View Article : Google Scholar
|
|
14
|
Jemal A, Bray F, Center M, Ferlay J, Ward
E and Forman D: Global cancer statistics. CA Cancer J Clin.
61:69–90. 2011.PubMed/NCBI View Article : Google Scholar
|
|
15
|
Ferlay J, Soerjomataram I, Dikshit R, Eser
S, Rebelo M, Parkin D, Forman D and Bray F: Cancer incidence and
mortality worldwide: Sources, methods and major patterns in
GLOBOCAN 2012. Int J Cancer. 136:E359–E386. 2014.PubMed/NCBI View Article : Google Scholar
|
|
16
|
Snietura M and Lange D: Current ASCO/CAP
2013 recommendations for testing for HER2 status in breast cancer.
Pol J Pathol. 65 (Suppl 2):S32–S41. 2014.PubMed/NCBI(In Polish).
|
|
17
|
Naja F, Fadel RA, Alameddine M, Aridi Y,
Zarif A, Hariri D, Mugharbel A, Khalil M, Nahleh Z and Tfayli A:
Complementary and alternative medicine use and its association with
quality of life among Lebanese breast cancer patients: A
cross-sectional study. BMC Complement Altern Med.
15(444)2015.PubMed/NCBI View Article : Google Scholar
|
|
18
|
Węsierska-Gądek J and Mauritz M: Why
(multi)targeting of cyclin-dependent kinases is a promising
therapeutic option for hormone-positive breast cancer and beyond.
Future Med Chem. 8:55–72. 2016.PubMed/NCBI View Article : Google Scholar
|
|
19
|
Li D, Bachinski LL and Roberts R: Genomic
organization and isoform-specific tissue expression of human NAPOR
(CUGBP2) as a candidate gene for familial arrhythmogenic right
ventricular dysplasia. Genomics. 74:396–401. 2001.PubMed/NCBI View Article : Google Scholar
|
|
20
|
Ramalingam S, Ramamoorthy P, Subramaniam D
and Anant S: Reduced expression of RNA binding protein CELF2, a
putative tumor suppressor gene in colon cancer.
Immunogastroenterology. 1:27–33. 2012.PubMed/NCBI View
Article : Google Scholar
|
|
21
|
Wang J, Liu L, Sun Y, Xue Y, Qu J, Pan S,
Li H, Qu H, Wang J and Zhang J: miR-615-3p promotes proliferation
and migration and inhibits apoptosis through its potential target
CELF2 in gastric cancer. Biomed Pharmacother. 101:406–413.
2018.PubMed/NCBI View Article : Google Scholar
|
|
22
|
Yeung YT, Fan S, Lu B, Yin S, Yang S, Nie
W, Wang M, Zhou L, Li T, Li X, et al: CELF2 suppresses non-small
cell lung carcinoma growth by inhibiting the PREX2-PTEN
interaction. Carcinogenesis. 41:377–389. 2020.PubMed/NCBI View Article : Google Scholar
|
|
23
|
Xu W, Gu J, Ren Q, Shi Y, Xia Q and Wang
J, Wang S, Wang Y and Wang J: NFATC1 promotes cell growth and
tumorigenesis in ovarian cancer up-regulating c-Myc through
ERK1/2/p38 MAPK signal pathway. Tumour Biol. 37:4493–4500.
2016.PubMed/NCBI View Article : Google Scholar
|
|
24
|
Wang L, Wang Z, Li J, Zhang W, Ren F and
Yue W: NFATc1 activation promotes the invasion of U251 human
glioblastoma multiforme cells through COX-2. Int J Mol Med.
35:1333–1340. 2015.PubMed/NCBI View Article : Google Scholar
|
|
25
|
Liu Y, Liang T, Qiu X, Ye X, Li Z, Tian B
and Yan D: Down-regulation of Nfatc1 suppresses proliferation,
migration, invasion, and warburg effect in prostate cancer cells.
Med Sci Monit. 25:1572–1581. 2019.PubMed/NCBI View Article : Google Scholar
|
|
26
|
Li L, Duan Z, Yu J and Dang HX: NFATc1
regulates cell proliferation, migration, and invasion of ovarian
cancer SKOV3 cells in vitro and in vivo. Oncol Rep. 36:918–928.
2016.PubMed/NCBI View Article : Google Scholar
|