Introduction
Icariin (ICA), a flavonoid extract of epimedium, has
a wide range of pharmacological effects, such as anti-inflammatory,
anti-oxidant and antitumor effects in the cardio-cerebral vascular,
nervous and urogenital systems and malignant tumors (1-4).
Chinese medicine has been widely used to treat several types of
malignant tumors. ICA, as a vital active ingredient of epimedium in
Chinese medicine, serves an inhibitory role in the occurrence and
development of malignant tumors; therefore, it has been widely used
in the prevention and treatment of numerous types of cancer, such
as cervical, ovarian, colon and triple-negative breast cancer
(5-8).
Oral squamous cell carcinoma (OSCC) occurs in the
oral mucosa, and is characterized by strong local invasion and easy
metastasis to the cervical lymph nodes (9,10).
Furthermore, ICA is considered as a novel biological immune
modulator and inducer of differentiation, and it has been reported
to improve immune function, inhibit tumor cell proliferation, tumor
growth and angiogenesis, induce tumor cell apoptosis and alter
tumor cell cycle distribution (11,12).
Compared with other traditional antitumor drugs, ICA can regulate
tumor immunity and reduce the lethality of cells of the surrounding
normal tissues. Li et al (13) demonstrated that ICA could inhibit
hepatocellular carcinoma cell proliferation, promote apoptosis and
enhance the antitumor effects of arsenic trioxide in vitro
and in vivo. Additionally, Yang et al (14) revealed that ICA not only inhibited
the proliferation of glioblastoma cells in a dose-dependent manner,
but also enhanced the antitumor effect of temozolomide. Shi et
al (15) also found that ICA
attenuated the proliferation of colorectal cancer (CRC) cells and
inhibited the antitumor activity of 5-fluorouracil in CRC via
inhibiting the activity of nuclear factor-κB (NF-κB) in
vitro. The NF-κB signaling pathway is closely associated with
tumor cell apoptosis and plays a key role in the immune aging
process of tumor cells (16-18).
The aforementioned studies indicate that ICA has a wide range of
potential antitumor effects. However, its effects on OSCC remain
elusive.
The phosphatidylinositol-3-kinase (PI3K)/protein
kinase B (AKT) signaling pathway has been reported to play critical
roles in various types of cancer (19-21).
For example, the PI3K/AKT pathway is regarded as one of the key
mechanisms involved in lung cancer cell metastasis and the
epithelial-mesenchymal transition (19). Besides, the PI3K/AKT signaling
pathway is associated with renal cell carcinoma cell proliferation
and metastasis (20). Moreover, the
PI3K/AKT pathway plays important roles in the regulation of human
pharyngeal squamous carcinoma cell apoptosis (21). Activation of the PI3K/AKT signaling
pathway has been confirmed in OSCC, and inhibiting this signaling
pathway has been reported to attenuate the development of OSCC
(22-24).
Until now, whether ICA could affect PI3K/AKT signaling pathway
activation in OSCC remain unclear. Therefore, the present study
aimed to investigate the effects and underlying molecular
mechanisms of ICA on OSCC cells to provide a novel theoretical
basis for the treatment of OSCC.
Materials and methods
Cell culture and treatment
The OSCC cell lines Cal 27 and SCC9 were obtained
from the American Type Culture Collection, and human oral mucosa
fibroblasts were purchased from Shanghai Aiyan Biotechnology Co.,
Ltd. All cells were cultured in Dulbecco's modified Eagle's medium
supplemented with 10% fetal bovine serum (both Gibco; Thermo Fisher
Scientific, Inc.), 100 U/ml penicillin and 100 µg/ml streptomycin
at 37˚C in a humidified incubator containing 5% CO2.
Human normal oral keratinocytes (hNOKs) were obtained from
Lifeline® Cell Technology. hNOKs were cultured in
DermaLife® K Medium (Lifeline® Cell Technology). Cells
were treated with 0, 5, 10, 20 and 40 µM ICA (Shanghai Yuanye
Biotechnology Co., Ltd.) for 24, 48 and 72 h.
3-(4,5-Dimethylthiazol-2-yl)-2,5-diphenyl-2H-tetrazolium bromide
(MTT) assay
OSCC cell proliferation was evaluated using a MTT
assay. Briefly, Cal 27 and SCC9 cells were seeded (104
cells per well) into 96-well plates and cultured at 37˚C in a
humidified incubator containing 5% CO2. Subsequently,
cells were treated with 0, 5, 10, 20 and 40 µM ICA for 24, 48 or 72
h. Following treatment, cells were supplemented with 10 µl MTT
solution and incubated for an additional 4 h, according to the
manufacturer's instructions. The formazan crystals were dissolved
by the addition of dimethyl sulfoxide. Finally, optical density
values were measured at 570 nm using a multifunctional plate reader
(BD Biosciences).
Flow cytometric analysis
Cell apoptosis was assessed using an Annexin-V/PI
Apoptosis Detection kit (Beyotime Institute of Biotechnology). Cal
27 and SCC9 cells were treated with 0, 5, 10, 20 and 40 µM ICA for
48 h. Subsequently, cells in the logarithmic phase of growth were
digested with 0.25% trypsin solution without ethylene diamine tetra
acetic acid, centrifuged at 1,000 x g for 5 min at 4˚C and the
supernatant was discarded. The cell pellet was then washed twice
with pre-chilled PBS and re-suspended in 100 µl of pre-chilled 1X
Annexin V binding buffer (Beyotime Institute of Biotechnology). The
cells were then incubated with 5 µl Annexin V-FITC and 5 µl
propidium iodide (Beyotime Institute of Biotechnology) for 15 min
at room temperature in the dark. To detect apoptosis, a BD
FACSCalibur flow cytometer (BD Biosciences) was used, and data were
analyzed using the Cell Quest software (version 5.1; BD
Biosciences).
Western blot analysis
The expression levels of cleaved-caspase-3 (cat. no.
ab32042; dilution, 1:500; Cell Signaling Technology, Inc.),
pro-caspase-3 (cat. no. ab32150; dilution rate, 1:1,000; Abcam),
Bcl-2 (cat. no. 4223; dilution, 1:1,000; Cell Signaling Technology,
Inc.), Bax (cat. no. 5023; dilution, 1:1,000), phosphorylated
(p)-p65 (cat. no. 3033; dilution, 1:1,000), p65 (cat. no. 8242;
dilution, 1:1,000) and p-AKT (cat. no. 4060; dilution, 1:1,000) and
AKT (cat. no. 4685; dilution, 1:1,000) (all Cell Signaling
Technology, Inc.) were evaluated using western blot analysis. Total
proteins were extracted from OSCC cells using RIPA lysis buffer
(Beyotime Institute of Biotechnology), and the protein
concentration was determined using a bicinchoninic acid assay kit
(Sigma-Aldrich; Merck KGaA), according to the manufacturer's
protocol. Proteins (40 µg per lane) were separated by 10% SDS-PAGE
and then transferred onto a polyvinylidene fluoride (PVDF) membrane
(EMD Millipore). Following blocking with 5% skimmed milk for 1 h at
room temperature, the membrane was first incubated with primary
antibodies at 4˚C overnight and then with anti-rabbit IgG
horseradish peroxidase-linked antibodies (cat. no. 7074; dilution,
1:2,000; Cell Signaling Technology, Inc.) at room temperature for 1
h. Finally, the blot was developed with enhanced chemiluminescence
reagent (Cyvita) and visualized. The strips were evaluated by light
gray value analysis using ImageJ software (version 1.46; National
Institutes of Health).
Reverse transcription-quantitative PCR
(RT-qPCR) analysis
Total RNA was extracted from OSCC cells using
TRIzol® reagent (Invitrogen; Thermo Fisher Scientific,
Inc.) according to the manufacturer's protocol. Total RNA was then
reverse transcribed into cDNA using the HiScript™ II qRT SuperMix
(Vazyme Biotech Co., Ltd.). Reverse transcription reaction
condition was as following: 25˚C For 5 min, 42˚C for 60 min and
80˚C for 2 min. cDNA was analyzed by qPCR using the ChamQ™
Universal SYBR qPCR Master mix (Vazyme Biotech Co., Ltd.) according
to the manufacturer's instructions. The amplification conditions
were as follows: Pre-denaturation at 95˚C for 10 min; followed by
40 cycles of denaturation at 95˚C for 10 sec, annealing at 60˚C for
20 sec and extension at 72˚C for 34 sec. Primer sequences were
listed as following: GAPDH, forward: 5'-CTTTGGTATCGTGGAAGGACTC-3'
and reverse: 5'-GTAGAGGCAGGGATGATGTTCT-3'; Bcl-2, forward:
5'-GATCCTCGAGATGGCGCACGCTGGGAGAAC-3' and reverse:
5'-GATCGGATCCTCATGGCTGAGCGCAG-3'; Bax, forward:
5'-GGACGAACTGGACAGTAACATGG-3' and reverse:
5'-GCAAAGTAGAA-AAGGGCGACAAC-3'; p65, forward:
5'-ACAACAACCCCTTCCAAGAAGA-3' and reverse:
5'-CAGCCTGGTCCCGTGAAATA-3'; AKT, forward:
5'-TAAAGAAGGAGGTCATCGTGG-3' and reverse: 5'-CGGGACAGGTGGAAGAAAA-3'.
The relative gene expression levels were analyzed using the
2-ΔΔCq method (25). All
experiments were performed in triplicate.
Statistical analysis
The data from three independent experiments are
expressed as the mean ± standard deviation. Statistical analysis
was carried out using GraphPad Prism 5 software (GraphPad Software,
Inc.). The statistical differences among different groups were
analyzed by one-way ANOVA followed by Tukey's post hoc tests.
P<0.05 was considered to indicate a statistically significant
difference.
Results
Effect of ICA on the OSCC cell lines,
Cal 27 and SCC9
The chemical molecular structure of ICA is shown in
Fig. 1A. First, the effect of ICA
on the proliferation of human normal oral keratinocytes (hNOKs) was
determined, and the results indicated that there was no significant
effect of ICA on the viability of hNOKs (Fig. 1B). Then, the effect of ICA on OSCC
cells investigated and Cal 27 and SCC9 cell lines were treated with
0, 5, 10, 20 or 40 µM ICA for 48 h. The effect of ICA on cell
viability was determined using a MTT assay. The results
demonstrated that ICA inhibited Cal 27 and SCC9 cell viability in a
dose-dependent manner (Fig. 1C and
D).
 | Figure 1Effect of ICA on OSCC cell viability.
(A) Chemical molecular structure of ICA. (B) hNOKs were treated
with 0, 5, 10, 20 or 40 µM ICA for 48 h, and then a MTT assay was
performed to measure cell viability. (C and D) OSCC cell lines, Cal
27 and SCC9, were treated with 0, 5, 10, 20 or 40 µM ICA for 48 h,
and then a MTT assay was performed to measure cell viability.
*P<0.05 and **P<0.01 vs. 0 µM ICA
control group. ICA, icariin; OSCC, oral squamous cell carcinoma;
hNOKs, human normal oral keratinocytes. |
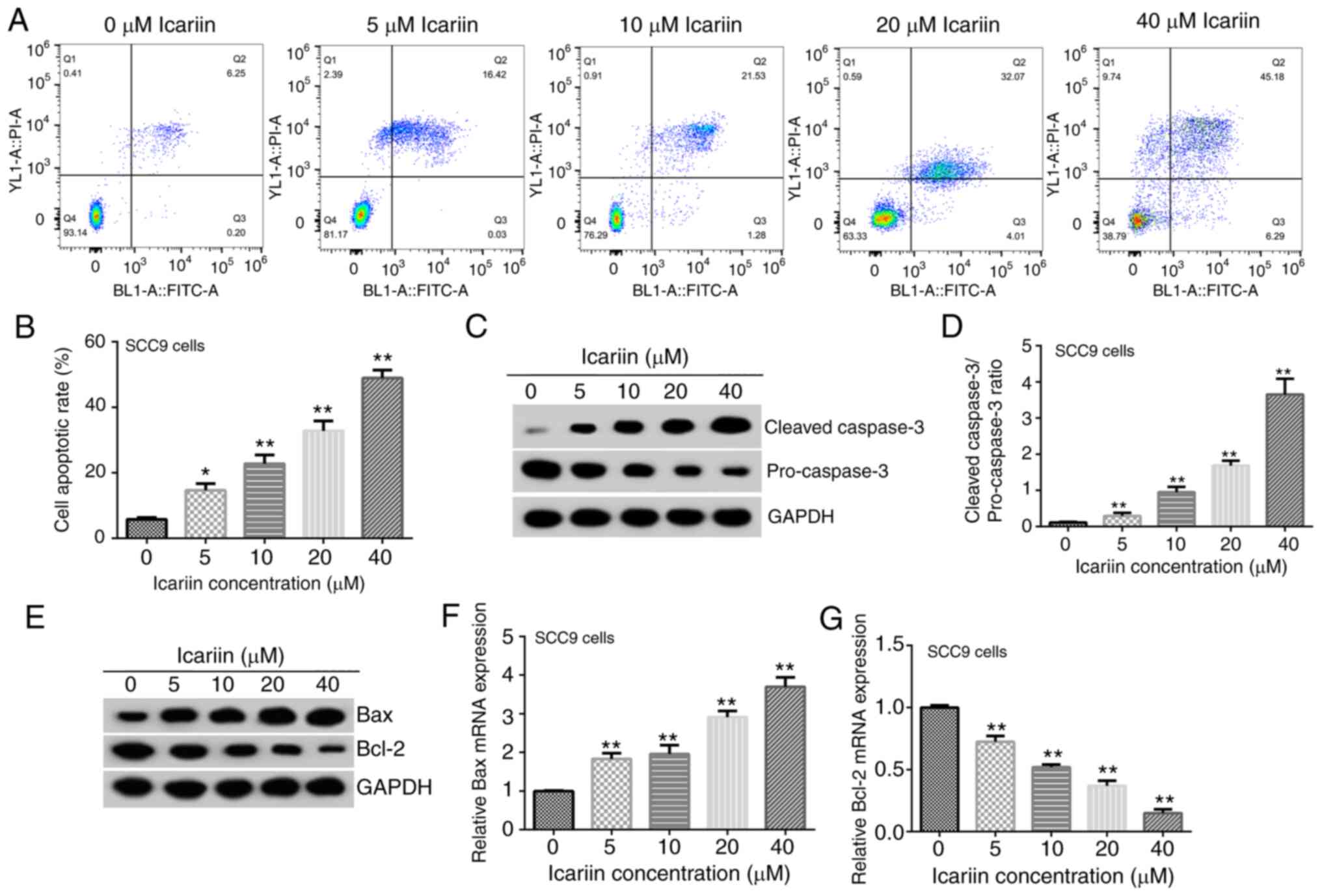
Effect of ICA on OSCC cell apoptosis
and expression of apoptosis-related proteins
The results revealed that following treatment of Cal
27 cells with ICA (0, 5, 10, 20 or 40 µM) for 48 h, the apoptosis
rate was significantly increased with increasing concentrations of
ICA (Fig. 2A and B). Furthermore, the protein expression
levels of cleaved-caspase-3 in Cal 27 cells were upregulated, those
of pro-caspase-3 were downregulated and the ratio of
cleaved-caspase-3/pro-caspase-3 was increased, in a dose-dependent
manner (Fig. 2C and D). Besides, it was demonstrated that the
protein and mRNA expression levels of Bax (Fig. 2E and F) in Cal 27 cells were upregulated, and
those of Bcl-2 (Fig. 2E and
G) were downregulated in a
dose-dependent manner. Additionally, the apoptosis rate results
(Fig. 3A and B) and the expression levels of the
apoptosis-related proteins (Fig.
3C-G) in SCC9 cells were consistent with those observed in the
Cal 27 cell line.
Effect of ICA on the NF-κB and
PI3K/AKT signaling pathways in OSCC cell lines
Cal 27 cells were treated with ICA (0, 5, 10, 20 or
40 µM) for 48 h, and the protein expression levels of p-p65, p65,
p-AKT and AKT were detected using western blot analysis. The
protein expression levels of p-p65 and p-AKT in Cal 27 cells were
decreased with increasing concentrations of ICA (Fig. 4A). In addition, the p-p65/p65 and
p-AKT/AKT protein ratios were reduced in Cal 27 cells treated with
ICA (Fig. 4B and C). The RT-qPCR results revealed no
significant changes in the mRNA expression levels of p65 and AKT
(Fig. 4D and E). Similar results were obtained in SCC9
cells (Fig. 5). The aforementioned
findings indicated that treatment of OSCC cells with ICA could
attenuate cell proliferation and induce apoptosis via inhibiting
the NF-κB and PI3K/AKT signaling pathways.
Discussion
The present study aimed to investigate the effects
of ICA in OSCC cells. The results showed that treatment with ICA
significantly increased the apoptosis rate in a dose- and
time-dependent manner. In OSCC cells treated with different
concentrations of ICA, the protein expression of cleaved-caspase-3
and pro-caspase-3 were up- and downregulated, respectively, in a
dose-dependent manner. Furthermore, ICA could attenuate OSCC cell
proliferation via inhibiting the NF-κB and PI3K/AKT signaling
pathways. These findings provided novel insights into the role of
ICA and potential for the management and clinical treatment of
OSCC.
With increasing research in tumor biology and
immunology, it has been gradually considered that the occurrence
and development of malignant tumors depend not only on the tumor
cells themselves, but also on their regulation by the immune system
(26). Therefore, ICA can regulate
the function of immune organs and cells, and enhance the activity
of immune cytokines, thus improving tumor immune function (27). It has been reported that the
Bax/Bcl-2 ratio is increased with increasing concentrations of ICA
(3). In addition, a study
demonstrated that ICA is involved in the human immune regulation
via increasing the activity of NF-κB in anti-inflammatory- and
antiviral-related pathways, inducing the expression of p65,
accelerating lymphocyte apoptosis and delaying immune aging
(28). Furthermore, ICA could
inhibit the proliferation of acute promyelocytic leukemia cells in
a dose-dependent manner, thereby promoting their apoptosis
(4). Additionally, a study revealed
that ICA could block the NF-κB signaling pathway, which in turn
could hinder the mRNA expression of the inflammation-related factor
inducible nitric oxide synthase and delay the onset of inflammatory
diseases, thus suggesting that ICA has strong anti-inflammatory
effects (2). Therefore, it was
hypothesized that ICA could also inhibit the NF-κB and PI3K/AKT
signaling pathways in OSCC, thereby attenuating OSCC cell
proliferation and inducing apoptosis.
In the current study, the effect of ICA on Cal 27
and SCC9 cell viability and apoptosis was assessed by MTT and
western blot assays, respectively. The results revealed that ICA
could attenuate cell viability and induce apoptosis in the OSCC
cell lines in a dose-dependent manner. Chen et al (2) demonstrated that ICA could upregulate
the protein expression of sirtuin 6 (SIRT6) and downregulate that
of NF-κB (p65) in animal tissues and cell models. These results
showed that the ICA-mediated SIRT6 upregulation had an inhibitory
effect on the NF-κB inflammatory signaling pathway, since treatment
with ICA decreased the mRNA expression levels of the NF-κB
downstream target genes, TNF-α, intercellular adhesion molecule 1,
IL-2 and IL-6. Herein, the protein expression levels of p-p65 and
p-AKT were decreased, following cell treatment with ICA, in a
dose-dependent manner. No obvious changes were observed regarding
the mRNA expression levels of p65 and AKT in both OSCC cell lines.
In addition, the p-p65/p65 and p-AKT/AKT ratios were decreased in
OSCC cells in a dose-dependent manner. The aforementioned findings
suggested that ICA could attenuate OSCC cell proliferation via
inhibiting the NF-κB and PI3K/AKT signaling pathways.
ICA is a novel biological immunomodulator and
inducer of differentiation, thus improving immunity function, and
inhibiting tumor cell proliferation, tumor growth and tumor
angiogenesis (27,29,30).
Herein, ICA inhibited the NF-κB and PI3K/AKT signaling pathways to
promote OSCC cell proliferation, suggesting that this drug should
be considered as a potential monomer for treating OSCC. However,
the current study still had some limitations. For example, the
effect of ICA was only studied in two OSCC cell lines, and so more
cell lines need to be used for verification. Furthermore, the
effects of only four concentrations of ICA on OSCC cell lines were
studied, thus more concentrations of ICA should be explored. In
addition, no in vivo studies were conducted. In the future,
these issues will be studied in more depth.
Acknowledgements
Not applicable.
Funding
Funding: No funding was received.
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
LS contributed to study design, data collection,
statistical analysis, data interpretation and manuscript
preparation. JZ contributed to data collection, statistical
analysis and manuscript preparation. All authors read and approved
the final manuscript. LS and JZ confirm the authenticity of all the
raw data.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Qin L, Zhang G, Sheng H, Wang XL, Wang YX,
Yeung KW, Griffith JF, Li ZR, Leung KS and Yao XS: Phytoestrogenic
compounds for prevention of steroid-associated osteonecrosis. J
Musculoskelet Neuronal Interact. 8:18–21. 2008.PubMed/NCBI
|
|
2
|
Chen Y, Sun T, Wu J, Kalionis B, Zhang C,
Yuan D, Huang J, Cai W, Fang H and Xia S: Icariin intervenes in
cardiac inflammaging through upregulation of SIRT6 enzyme activity
and inhibition of the NF-kappa B pathway. Biomed Res Int.
2015(895976)2015.PubMed/NCBI View Article : Google Scholar
|
|
3
|
Carlsten M and Childs RW: Genetic
manipulation of NK cells for cancer immunotherapy: Techniques and
clinical implications. Front Inmunol. 6(266)2015.PubMed/NCBI View Article : Google Scholar
|
|
4
|
Wang Z, Zhang H, Dai L, Song T, Li P, Liu
Y and Wang L: Arsenic trioxide and icariin show synergistic
anti-leukemic activity. Cell Biochem Biophys. 73:213–219.
2015.PubMed/NCBI View Article : Google Scholar
|
|
5
|
Huang S, Xie T and Liu W: Icariin inhibits
the growth of human cervical cancer cells by inducing apoptosis and
autophagy by targeting mTOR/PI3K/AKT signalling pathway. J BUON.
24:990–996. 2019.PubMed/NCBI
|
|
6
|
Wang P, Zhang J, Xiong X, Yuan W, Qin S,
Cao W, Dai L, Xie F, Li A and Liu Z: Icariin suppresses cell cycle
transition and cell migration in ovarian cancer cells. Oncol Rep.
41:2321–2328. 2019.PubMed/NCBI View Article : Google Scholar
|
|
7
|
Kim B, Seo JH, Lee KY and Park B: Icariin
sensitizes human colon cancer cells to TRAIL-induced apoptosis via
ERK-mediated upregulation of death receptors. Int J Oncol.
56:821–834. 2020.PubMed/NCBI View Article : Google Scholar
|
|
8
|
Song L, Chen X, Mi L, Liu C, Zhu S, Yang
T, Luo X, Zhang Q, Lu H and Liang X: Icariin-induced inhibition of
SIRT6/NF-κB triggers redox mediated apoptosis and enhances
anti-tumor immunity in triple-negative breast cancer. Cancer Sci.
111:4242–4256. 2020.PubMed/NCBI View Article : Google Scholar
|
|
9
|
Ju ZY, Tang QM, Chen LL, et al: Axl
inhibitor R428 induces autophagy of oral squamous cell carcinoma
Cal27 cells. J Pract Stomatol. 35:2019.
|
|
10
|
Hu XG, Qiu ZL, Zen JC, et al: Differential
expression of microRNAs in human oral squamous cell carcinoma stem
cells. J Pract Stomatol. 3:398–402. 2019.
|
|
11
|
Schneider P, Schon M, Pletz N, Seitz CS,
Liu N, Ziegelbauer K, Zachmann K, Emmert S and Schön MP: The novel
PI3 kinase inhibitor, BAY80-6946, impairs melanoma growth in vivo
and in vitro. Exp Dermatol. 23:579–584. 2014.PubMed/NCBI View Article : Google Scholar
|
|
12
|
Liu YJ, Huang LL, Hao BH, Li H, Zhu S,
Wang Q, Li R, Xu Y and Zhang X: Use of an osteoblast overload
damage model to probe the effect of icariin on the proliferation,
differentiation and mineralization of MC3T3-E1cells through the
Wnt/β-catenin signalling pathway. Cell Physiol Biochem.
41:1605–1615. 2017.PubMed/NCBI View Article : Google Scholar
|
|
13
|
Li W, Wang M, Wang LY, Ji S, Zhang J and
Zhang C: Icariin synergizes with arsenic trioxide to suppress human
hepatocellular carcinoma. Cell Biochem Biophys. 68:427–436.
2014.PubMed/NCBI View Article : Google Scholar
|
|
14
|
Yang LJ, Wang YX, Guo H and Guo M:
Synergistic anti-cancer effects of icariin and temozolomide in
glioblastoma. Cell Biochem Biophys. 71:1379–1385. 2015.PubMed/NCBI View Article : Google Scholar
|
|
15
|
Shi DB, Li XX, Zheng HT, Li DW, Cai GX,
Peng JJ, Gu WL, Guan ZQ, Xu Y and Cai SJ: Icariin-mediated
inhibition of NF-κB activity enhances the in vitro and in vivo
antitu-mour effect of 5-fluorouracil in colorectal cancer. Cell
Biochem Biophys. 69:523–530. 2014.PubMed/NCBI View Article : Google Scholar
|
|
16
|
Xu W, Tao H and Zhang Q: Effect of Six1
gene on As2O3-induced oral squamous cell
carcinoma cell apoptosis and ROS levels. J Clin Exp Medicine.
(15):2019.
|
|
17
|
Peng M and Pang C: MicroRNA-140-5p
inhibits the tumorigenesis of oral squamous cell carcinoma by
targeting p21-activated kinase 4. Cell Biol Int: Aug 8, 2019 (Epub
ahead of print).
|
|
18
|
Wang XM, Liu CM, Zhang CR, Xu XG and Fu
SB: Functional significance of TGF-beta1 signal transduction
pathway in oral squamous cell carcinoma. Zhonghua Zhong Liu Za Zhi.
31:28–32. 2009.PubMed/NCBI(In Chinese).
|
|
19
|
Chen Z, He J, Xing X, Li P, Zhang W, Tong
Z, Jing X, Li L, Liu D, Wu Q and Ju H: Mn12Ac inhibits the
migration, invasion and epithelial-mesenchymal transition of lung
cancer cells by downregulating the Wnt/β-catenin and PI3K/AKT
signaling pathways. Oncol Lett. 16:3943–3948. 2018.PubMed/NCBI View Article : Google Scholar
|
|
20
|
Xie J, Lin W, Huang L, Xu N, Xu A, Chen B,
Watanabe M, Liu C and Huang P: Bufalin suppresses the proliferation
and metastasis of renal cell carcinoma by inhibiting the
PI3K/Akt/mTOR signaling pathway. Oncol Lett. 16:3867–3873.
2018.PubMed/NCBI View Article : Google Scholar
|
|
21
|
Choi MS, Moon SM, Lee SA, Park BR, Kim JS,
Kim DK, Kim YH and Kim CS: Adenosine induces intrinsic apoptosis
via the PI3K/Akt/mTOR signaling pathway in human pharyngeal
squamous carcinoma FaDu cells. Oncol Lett. 15:6489–6496.
2018.PubMed/NCBI View Article : Google Scholar
|
|
22
|
Hao Y, Zhang C, Sun Y and Xu H:
Licochalcone A inhibits cell proliferation, migration, and invasion
through regulating the PI3K/AKT signaling pathway in oral squamous
cell carcinoma. Onco Targets Ther. 12:4427–4435. 2019.PubMed/NCBI View Article : Google Scholar
|
|
23
|
Fan QC, Tian H, Wang Y and Liu XB:
Integrin-alpha5 promoted the progression of oral squamous cell
carcinoma and modulated PI3K/AKT signaling pathway. Arch Oral Biol.
101:85–91. 2019.PubMed/NCBI View Article : Google Scholar
|
|
24
|
Velu P, Vijayalakshmi A and Vinothkumar V:
Inhibiting the PI3K/Akt, NF-kB signalling pathways with syringic
acid for attenuating the development of oral squamous cell
carcinoma cells SCC131. J Pharm Pharmacol. 72:1595–1606.
2020.PubMed/NCBI View Article : Google Scholar
|
|
25
|
Ferlay J, Soerjomataram I, Dikshit R, Eser
S, Mathers C, Rebelo M, Parkin DM, Forman D and Bray F: Cancer
incidence and mortality worldwide: Sources, methods and major
patterns in GLOBOCAN 2012. Int J Cancer. 136:E359–E386.
2015.PubMed/NCBI View Article : Google Scholar
|
|
26
|
Galluzzi L and Rudqvist NP: Preface: More
than two decades of modern tumor immunology. Methods Enzymol.
629:xxii–xlii. 2019.PubMed/NCBI View Article : Google Scholar
|
|
27
|
Zhang X, Kang Z, Li Q, Zhang J, Cheng S,
Chang H, Wang S, Cao S, Li T, Li J, et al: Antigen-adjuvant effects
of icariin in enhancing tumor-specific immunity in
mastocytoma-bearing DBA/2J mice. Biomed Pharmacother. 99:810–816.
2018.PubMed/NCBI View Article : Google Scholar
|
|
28
|
Gao M and Li W: In vitro study on the
inhibitory effect of icariin on human bladder cancer T24 cells.
Shijiazhuang: Hebei Medical University 2010.
|
|
29
|
Hao H, Zhang Q, Zhu H, Wen Y, Qiu D, Xiong
J, Fu X, Wu Y, Meng K and Li J: Icaritin promotes tumor T-cell
infiltration and induces antitumor immunity in mice. Eur J Immunol.
49:2235–2244. 2019.PubMed/NCBI View Article : Google Scholar
|
|
30
|
Tan HL, Chan KG, Pusparajah P, Saokaew S,
Duangjai A, Lee LH and Goh BH: Anti-cancer properties of the
naturally occurring aphrodisiacs: Icariin and its derivatives.
Front Pharmacol. 7(191)2016.PubMed/NCBI View Article : Google Scholar
|