Introduction
The expression of melanin, which produces
pigmentation (hair or skin color), is regulated by tyrosinase (TYR)
and tyrosinase-related proteins 1 and 2 (TYRP-1/2) (1,2). TYR
is regulated by microphthalmia-associated transcription factor
(MITF), which mediates the growth, proliferation, and
differentiation of melanocytes (3). MITF is regulated by several cell
signaling pathways during melanogenesis (4,5).
Stimulants such as isobutylmethylxanthine (IBMX), α-melanotropin,
and forskolin activate melanogenesis-related proteins by inducing
the expression of MITF and cyclic adenosine monophosphate (cAMP)
(6,7). Melanogenesis-related proteins are
activated by cAMP response element-binding protein (CREB), which is
phosphorylated by protein kinase A, thereby promoting MITF
transcription and inducing eumelanin synthesis (8-10).
MITF expression is regulated via the protein kinase B (AKT),
extracellular signal-regulated kinase (ERK), and phosphoinositide
3-kinase (PI3K) signaling pathways during melanogenesis (11). Phosphorylated ERK (p-ERK), AKT
(p-AKT), and PI3K (p-PI3K) inhibit melanin activity by promoting
the degradation and phosphorylation of MITF (12,13).
Nypa fruticans Wurmb (NF) is a plant found primarily in
tropical mangrove systems (14).
NF is considered a plant with low utility in the Araceae family.
Its roots, leaves, and stems have been traditionally used as
analgesics for liver disease, asthma, and sore throat (15). NF contains several substances (such
as flavonoids and polyphenols) that reportedly exhibit anticancer
and antioxidative effects (16).
In addition, flavonoids and polyphenols present in plants
reportedly inhibit melanogenesis (17-20).
Studies have indicated that NF has anti-nociceptive,
neuroprotective, and anti-inflammatory properties (21,22).
Therefore, we investigated the inhibitory effect of the ethyl
acetate fraction of N. fruticans (ENF) on melanogenesis and
cell signaling pathways via AKT/mammalian target of rapamycin
(mTOR)/CREB and mitogen-activated protein kinase (MAPK).
Materials and methods
Chemicals and reagents
HPLC-grade petroleum ether, methanol, dimethyl
sulfoxide (DMSO), and ethyl acetate were purchased from Merck
(Darmstadt, Germany). Antibiotics (streptomycin and penicillin),
0.25% trypsin with EDTA in HBSS, high-glucose Dulbecco's modified
Eagle medium (DMEM), and fetal bovine serum (FBS) were purchased
from Biowest (Nuaillé, France). Plasmocin prophylactic for
mycoplasma growth inhibition was purchased from InvivoGen (San
Diego, CA, USA). Antibodies for TYR (SC-7834), TYRP-1 (SC-10448),
TYRP-2 (SC-10451), and MITF (SC-10999) and anti-goat antibodies
(SC-2020) were purchased from Santa Cruz Biotechnology (Dallas, TX,
USA). p-ERK (9101S), ERK (4695S), p-CREB (9198S), CREB (4820S),
p-AKT (4060S), AKT (9272S), p-mTOR (5536S), mTOR (2972S), PI3K
(4292S), and anti-mouse (7076S) and anti-rabbit (7074S) antibodies
were purchased from Cell Signaling (Danvers, MA, USA).
Glyceraldehyde 3-phosphate dehydrogenase (GAPDH, ab8245), Alexa
Fluor® 568 (ab175471), p-PI3K (ab182651), and Alexa
Fluor® 488 (ab150113) were purchased from Abcam
(Cambridge, MA, USA). All standards for chromatography were
purchased from Sigma-Aldrich (St. Louis, MO, USA).
Sample preparation
NF was purchased from Nesta (Dongdaemun-gu, Seoul,
Korea). An NF sample (484.0 g) was extracted with 80% methanol (4.2
l) for 7 days. The extract and fraction were concentrated using a
vacuum evaporator (N-1110S, EYELA, Tokyo, Japan). The aqueous
residue was fractionated with petroleum ether and ethyl acetate.
The fractionated sample (ENF, 36.8 g) was refrigerated at 4˚C until
use. ENF used in the experiment was dissolved in DMSO (not
exceeding 0.1%) at a concentration of 50 mg/ml.
Analysis of compounds in ENF using
liquid chromatography-mass spectrometry (LC-MS)
Compounds in ENF (injection volume: 10 µl) were
analyzed using an e2695 system equipped with an ACQUITY QDa
detector (Waters, Milford, MA, USA) based on standards. The column
used for separation, a Sunfire (C18, 5 µm, 250x4.6 mm, Waters), was
maintained at 25˚C during analysis. 100% Acetonitrile (solvent A)
and 1.0% glacial acetic acid in deionized water (solvent B) were
used as solvents for the mobile phase (flow rate: 0.3 ml/min). The
proportions of solvent A were set as 1.0% at 0 min, 20% at 8 min,
30% at 40 min, 40.0% at 45 min, and 1.0% at 50 min. The
electrospray ionization mass spectrometer was operated in the
negative ion mode (mass range: m/z 100-600). The cone voltage was
set to 30 V and the capillary voltage was set to 0.8 kV.
Cell culture
B16F10 cells were purchased from the American Type
Culture Collection (CRL-6475, Manassas, VA, USA). The cells were
grown in an incubator (Thermo Fisher Scientific, Waltham, MA, USA)
under 5% CO2 in a humidified environment at 37˚C. The
medium for cell culture consisted of DMEM containing 1%
antibiotics, 1 ml Plasmocin prophylactic, and 10% FBS (complete
DMEM: cDMEM).
Cell viability using MTS assay
The protocol used in a previous study was adopted
(23). Briefly, cell viability was
measured using MTS reagent (Cell Titer 96® Aqueous One
Solution, Promega, Madison, WI, USA). B16F10 cells were cultured in
a 96-well plate using cDMEM for 24 h. The cells were treated with
ENF (12.5-400.0 µg/ml) for 24 h. Next, they were treated with 20 µl
MTS reagent for 2 h in an incubator maintained with 5%
CO2 in a humidified environment at 37˚C. The absorbance
of MTS reagent at a wavelength of 540 nm was measured using a
microplate reader (Biotek, Winooski, VT, USA) to determine cell
viability.
Measurement of melanin content
The modified protocol used in a previous study was
adopted (24). Briefly, the
melanin content at the cellular level was investigated. B16F10
cells were treated with ENF and IBMX for 48 h. The melanin produced
was dissolved in 1 N NaOH containing 10% DMSO at 80˚C and the
absorbance of the solution was measured at 475 nm using a
UV/Visible spectrophotometer (X-ma 3000 (PC), Human Corp., Seoul,
Korea).
Immunoblotting
B16F10, the murine skin melanoma cell line, was
cultured at a density of 2x105 cells/well in a 6-well
plate for 24 h. Cells were pre-treated with ENF (25.0, 50.0, and
100.0 µg/ml) for 2 h and then treated with IBMX for 48 h. The cell
lysates prepared using RIPA buffer (supplemented with protease
inhibitor cocktail and 0.5 M EDTA solution, Thermo Fisher
Scientific) were centrifuged at 16,000 x g and 4˚C for 15 min.
Protein content in the cell lysates was quantified using Quick
Start Bradford 1X Dye Reagent (Bio-Rad, Hercules, CA, USA)
following the manufacturer's protocol. The proteins were
electrophoresed and then transferred to polyvinylidene fluoride
membranes (Bio-Rad) using the Trans-Blot® Turbo™
Transfer System (Bio-Rad). Blots were blocked using 5.0% BSA
(Bovine Serum Albumin, Bio-Sesang, Seoul, Korea) in Tris-buffered
saline supplemented with 0.1% Tween 20 (TBS-T, Bio-Sesang). Next, a
specific primary antibody was added to 3.0% BSA (1:2,000) and
incubated overnight at 4˚C. Subsequently, the blots were washed
with TBS-T and incubated with the HRP-conjugated secondary antibody
for 1 h. Chemiluminescence was detected using Clarity™ Western ECL
substrate (Bio-Rad) and visualized with a Chemi-Doc (Bio-Rad). The
intensity of the blots was analyzed using ImageJ software 1.51k
(developed at the National Institutes of Health, USA).
cDNA synthesis, reverse
transcription-quantitative polymerase chain reaction (RT-qPCR), and
reverse transcription-polymerase chain reaction (RT-PCR)
cDNA was synthesized from extracted RNA using
NucleoSpin® RNA Plus (Macherey-Nagel, Düren, Germany) in
a T100™ thermal cycler (Bio-Rad) according to the user manual.
RT-qPCR was performed using a Rotor-Gene Q (Qiagen, Hilden,
Germany) with a QuantiTect® SYBR Green PCR kit (Qiagen)
and an appropriate primer according to the user manual. Primer 3
software was used for primer design (Table I). Data analysis was conducted
using Rotor-Gene Q Series software 2.3.5 (Qiagen). Transcription
levels were normalized to those of the GAPDH gene. The
formula used to analyze mRNA expression was
2-ΔΔCt, where
ΔΔCt=(Cttarget-CtGAPDH)sample-(Cttarget-CtGAPDH)control.
RT-PCR was performed using a T100™ thermal cycler with Quick
Taq® HS DyeMix (Toyobo, Osaka, Japan) and synthesized
cDNA. The PCR product was subjected to DNA electrophoresis (2%
agarose gel with DNA SafeStain, LAMDA Biotech, Ballwin, MO, USA).
The intensity of the bands was analyzed using ImageJ software.
 | Table IPrimer sequences for RT-PCR and
RT-qPCR. |
Table I
Primer sequences for RT-PCR and
RT-qPCR.
| A, RT-PCR |
|---|
| Gene | Sequence,
5'-3' | Product size,
bp |
|---|
| TYR | F:
GAGAAGCGAGTCTTGATTAG | 176 |
| | R:
TGGTGCTTCATGGGCAAAATC | |
| TYRP-1 | F:
GCTGCAGGAGCCTTCTTTCTC | 268 |
| | R:
AAGACGCTGCACTGCTGGTCT | |
| TYRP-2 | F:
CCTGTCTCTCCAGAAGTTTG | 218 |
| | R:
CGTCTGTAAAAGAGTGGAGG | |
| MITF | F:
AGCGTGTATTTTCCCCACAG | 124 |
| | R:
TAGCTCCTTAATGCGGTCGT | |
| GAPDH | F:
AACTTTGGCATTGTGGAAGG | 130 |
| | R:
ATGCAGGGATGATGTTCTGG | |
| B, RT-qPCR |
| Gene | Sequence,
5'-3' | Product size,
bp |
| TYR | F:
ACAGCTACCTCCAAGAGTCA | 148 |
| | R:
TACTGCTAAGCCCAGAGAGA | |
| TYRP-1 | F:
CAAAGAGCAGCATAGGAGAC | 139 |
| | R:
ACACTCTCGTGGAAACTGAG | |
| TYRP-2 | F:
ATGAGGAGCTCTTCCTAACC | 106 |
| | R:
CCAATGACCACTGAGAGAGT | |
| MITF | F:
CTCAGCAGTCTCTTTTGGAC | 110 |
| | R:
AATGTCTACAGAGGCACCAC | |
| GAPDH | F:
CCTCCAAGGAGTAAGAAACC | 143 |
| | R:
CTAGGCCCCTCCTGTTATTA | |
Immunofluorescence (IF)
B16F10 cells were cultured on a glass coverslip and
incubated for 24 h. The cells were pre-treated with ENF (100 µg/ml)
for 2 h and then treated with IBMX for 24 h. Thereafter, the cells
were treated with 4% paraformaldehyde (Bio-Sesang) dissolved in
phosphate-buffered saline (PBS) and incubated at 25˚C for 15 min
for fixing. After incubation, the cells were washed with PBS and
blocked with 2% BSA in PBS (supplemented with 0.1% Triton-X, PBS-T,
Bio-Sesang) for 1 h. For IF analysis, the coverslips were treated
with anti-TYR and MITF antibodies (diluted 1:1,000) in 1% BSA/PBS-T
and incubated overnight at 4˚C. After incubation, anti-goat IgG
(Alexa Fluor® 568) and anti-mouse IgG (Alexa
Fluor® 488) were reacted with the primary antibody and
incubated for 1 h in the dark. 4',6-Diamidino-2-phenylindole
(Invitrogen, Waltham, MA, USA) was diluted in PBS-T and incubated
at 25˚C for 10 min. A fluorescence mounting solution (S3023, Dako,
Carpinteria, CA, USA) was added to the slide to mount the
coverslip. Images at x400 magnification were captured using a
fluorescence microscope (CKX53, Olympus, Tokyo, Japan) and DSLR
camera (DS126271, Canon, Tokyo, Japan), and the fluorescence
intensity was analyzed using ImageJ software.
Statistical analysis
All experimental data were statistically analyzed
using a statistics program (GraphPad Prism 5.02, GraphPad Software,
San Diego, CA, USA). Each data point was analyzed using one-way
analysis of variance. Dunnett's post-hoc test was used to compare
mean values between groups. P<0.05 was considered to indicate a
statistically significant difference.
Results
Compound analysis
The compounds in ENF were identified using LC-MS
chromatograms and quantified by comparison with the quantitative
linear equations of the corresponding standards. The quantitative
linear equation and its linearity for catechin (y=12434x+276633,
R2=0.9917), epicatechin (y=11765x+412542,
R2=0.9865), and isoquercitrin (y=21534x-700748,
R2=0.9945) were determined using various concentrations
of the standards (Fig. 1). Thus,
it was found that ENF contained 130.1 mg/g of catechin, 16.1 mg/g
of epicatechin, and 85.7 mg/g of isoquercitrin (Table II).
| Table IICatechin, epicatechin, and
isoquercitrin content of ethyl acetate fraction of Nypa
fruticans Wurmb as determined by liquid chromatography-mass
spectrometry analysis. |
Table II
Catechin, epicatechin, and
isoquercitrin content of ethyl acetate fraction of Nypa
fruticans Wurmb as determined by liquid chromatography-mass
spectrometry analysis.
| Compound | Molecular
weight | Retention time,
min | Content, mg/g |
|---|
| Catechin | 290.26 | 24.751 | 130.1 |
| Epicatechin | 290.26 | 26.991 | 16.1 |
| Isoquercitrin | 464.10 | 35.164 | 85.7 |
Effects of ENF on cell viability
To investigate the cytotoxicity of ENF, B16F10 cells
were treated with 0-400 µg/ml ENF and incubated for 24 h. After
incubation, MTS reagent was used to analyze cell viability
following the manufacturer's protocol. ENF did not significantly
inhibit cell growth and exhibited no cytotoxicity in B16F10 cells
up to a concentration of 400 µg/ml (Fig. 2).
Effects of ENF on the expression of
TYR, TYRP-1, TYRP-2, and MITF
To evaluate whether ENF inhibits melanogenesis, we
measured intracellular melanin contents after the application of
ENF to B16F10 cells in the presence of IBMX. As shown in Fig. 3A, IBMX treatment increased
intracellular melanin contents compared with those of the control.
However, ENF treatment in the presence of IBMX decreased
intracellular melanin contents compared with those of the
IBMX-treated group. B16F10 cells stimulated by IBMX exhibited an
increase in the expression of TYRP-1 (1.86-fold), TYRP-2
(1.62-fold), TYR (3.03-fold), and MITF (2.96-fold) compared with
the untreated group (1.00-fold). In contrast, ENF treatment alone
at 50 and 100 µg/ml significantly decreased the expression of
TYRP-1 (1.69 and 0.83-fold), TYRP-2 (0.86 and 0.44-fold), TYR (1.73
and 1.04-fold), and MITF (1.73 and 1.04-fold) (Fig. 3B-F). The inhibitory effect of ENF
was also confirmed at the mRNA level (Fig. 4A-E). B16F10 cells stimulated by
IBMX exhibited an increase in the expression of TYRP-1 (3.62-fold),
TYRP-2 (3.07-fold), TYR (2.45-fold), and MITF (3.02-fold) mRNA
compared with the untreated group (1.00-fold). ENF treatment at 100
µg/ml suppressed the mRNA levels of TYRP-1 (2.14-fold), TYRP-2
(2.40-fold), TYR (1.80-fold), and MITF (1.80-fold). The
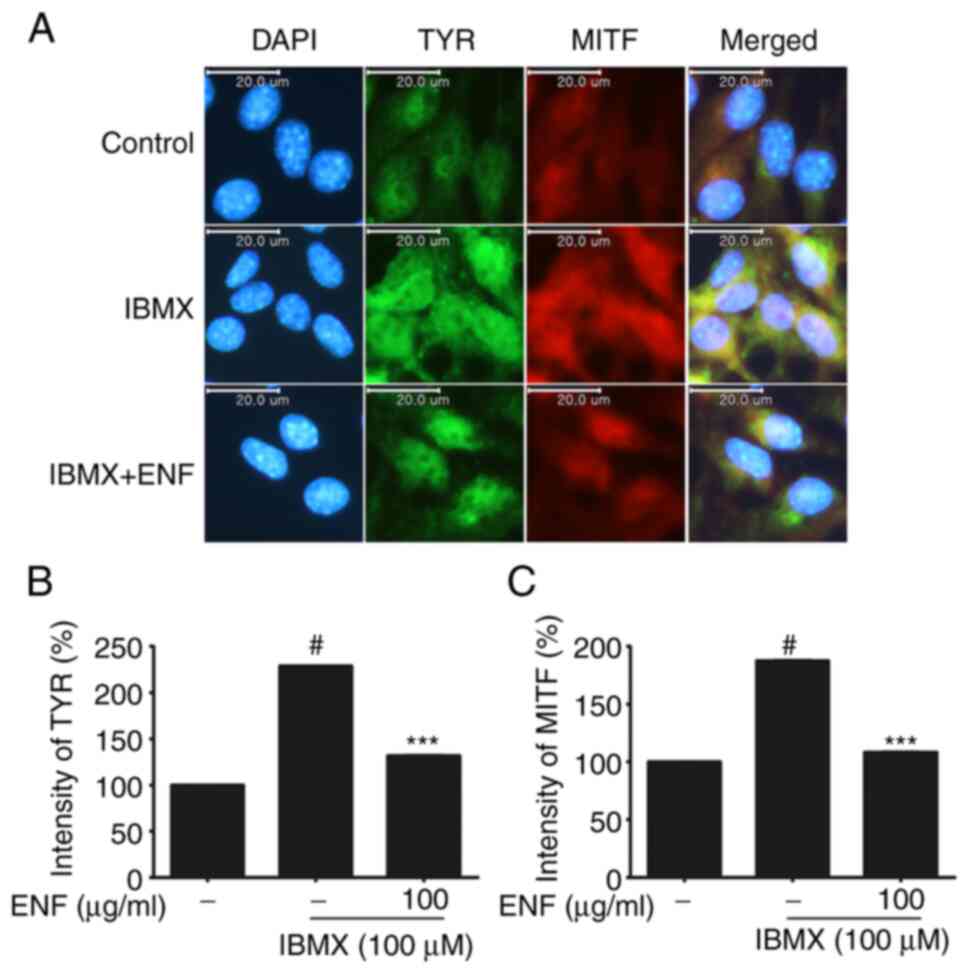
accumulation of TYR and MITF in live cells was confirmed using
immunofluorescence (Fig. 5A-C).
TYR (green) over-accumulated in the IBMX-treated group. Its
accumulation was suppressed in the 100 µg/ml ENF-treated group.
MITF (red) also showed excessive accumulation in the IBMX-treated
group; its accumulation was also suppressed in the 100 µg/ml
ENF-treated group.
 | Figure 3Inhibitory effects of ENF on the
expression of melanin, TYRP-1, TYRP-2, TYR, and MITF during
melanogenesis. (A) Effect of ENF on melanin content in B16F10
cells. (B) Expression of TYRP-1, TYRP-2, TYR, and MITF based on
immunoblotting. (C) Bar graph of TYRP-1 expression. (D) Bar graph
of TYRP-2 expression. (E) Bar graph of TYR expression. (F) Bar
graph of MITF expression. All results are expressed as means ±
standard deviations (n≥3). #P<0.001 vs. untreated
group; *P<0.05, **P<0.01,
***P<0.001 vs. IBMX-stimulated group. ENF, ethyl
acetate fraction of Nypa fruticans Wurmb; IBMX,
isobutylmethylxanthine; MITF, microphthalmia-associated
transcription factor; TYR, tyrosinase; TYRP, TYR-related
protein. |
 | Figure 4Inhibitory effects of ENF on
expression of TYRP-1, TYRP-2, TYR, and MITF mRNA during
melanogenesis. (A) Expression of TYRP-1, TYRP-2, TYR, and MITF mRNA
based on RT-PCR. (B) Bar graph of the expression of TYRP-1 mRNA
based on RT-qPCR. (C) Bar graph of the expression of TYRP-2 mRNA
based on RT-qPCR. (D) Bar graph of the expression of TYR mRNA based
on RT-qPCR. (E) Bar graph of the expression of MITF mRNA based on
RT-qPCR. All results are expressed as means ± standard deviations
(n≥3). #P<0.001 vs. untreated group;
*P<0.05, ***P<0.001 vs. IBMX-stimulated
group. ENF, ethyl acetate fraction of Nypa fruticans Wurmb;
IBMX, isobutylmethylxanthine; MITF, microphthalmia-associated
transcription factor; RT-PCR, reverse transcription-polymerase
chain reaction; RT-qPCR, RT-quantitative PCR; TYR, tyrosinase;
TYRP, TYR-related protein. |
Effects of ENF on CREB and
PI3K/AKT/mTOR signaling pathways
The CREB and PI3K/AKT/mTOR signaling pathways
reportedly regulate MITF expression (25-29).
B16F10 cells stimulated by IBMX showed an increase in the
expression of p-CREB (1.68-fold) compared with the untreated group
(1.00-fold). The p-CREB levels were lower in the ENF-treated groups
(0.08-fold at 100 µg/ml) than in the IBMX-stimulated group
(Fig. 6A and B). In IBMX-induced B16F10 cells, the
levels of mTOR, AKT, and PI3K phosphorylation were 3.42, 4.58, and
1.29-fold higher than those in the untreated group (1.00-fold). In
contrast, mTOR, AKT, and PI3K showed lower phosphorylation levels
in the ENF-treated group (0.16, 0.75, and 0.10-fold at 100 µg/ml)
than in the IBMX-stimulated group (Fig. 6A-E). Finally, ERK phosphorylation
was significantly inhibited in the ENF-treated group (0.67-fold at
100 µg/ml) compared with that in the IBMX-stimulated group
(1.26-fold) (Fig. 6A and F).
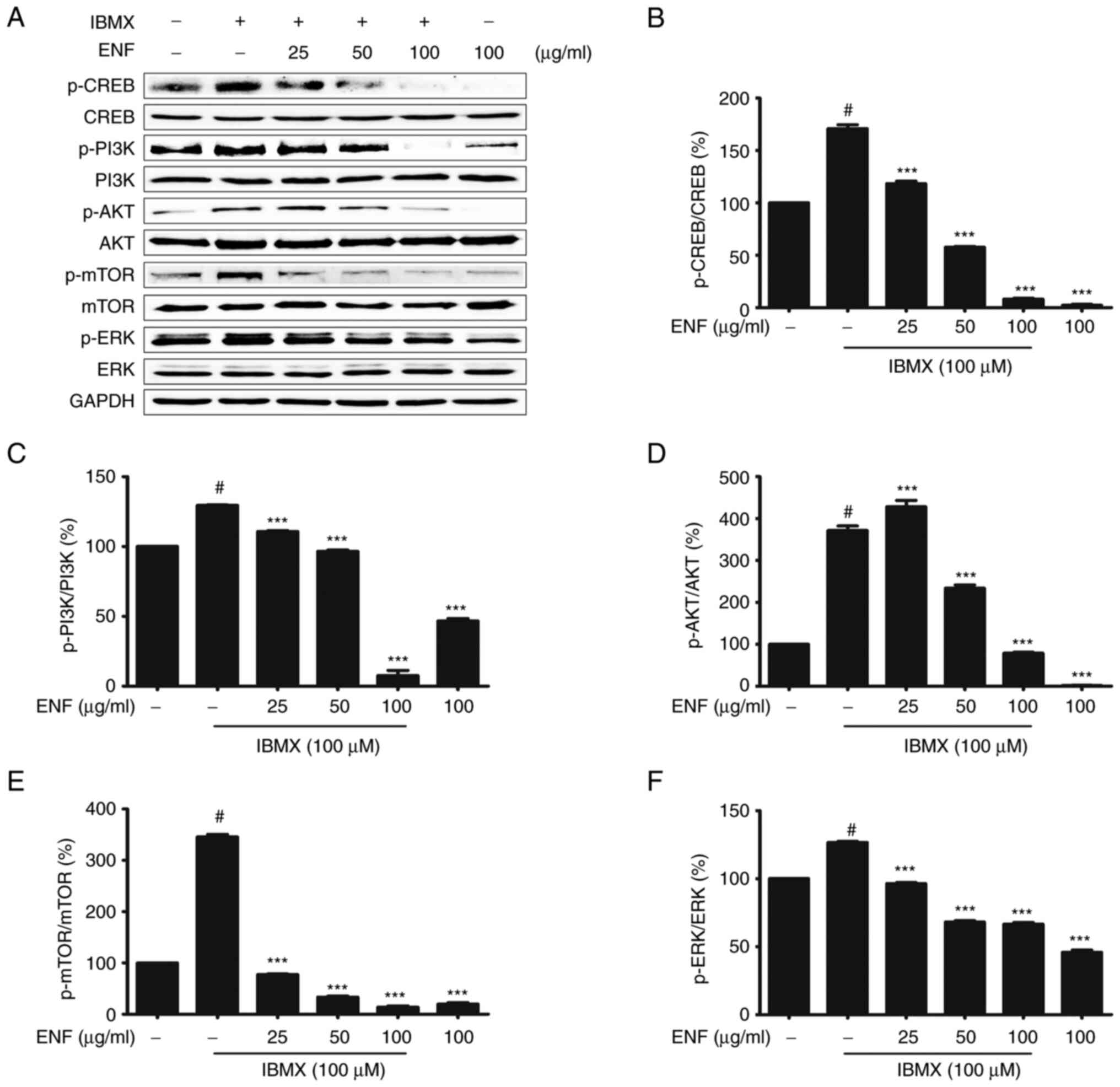 | Figure 6Inhibitory effects of ENF on the
expression of p-CREB, p-PI3K, p-AKT, p-mTOR, and p-ERK during
melanogenesis. (A) Expression of p-CREB, p-PI3K, p-AKT, p-mTOR, and
p-ERK based on immunoblotting. (B) Bar graph of p-CREB expression.
(C) Bar graph of p-PI3K expression. (D) Bar graph of p-AKT
expression. (E) Bar graph of p-mTOR expression. (F) Bar graph of
p-ERK expression. All results are expressed as means ± standard
deviations (n≥3). #P<0.001 vs. untreated group;
***P<0.001 vs. IBMX-stimulated group. AKT, protein
kinase B; CREB, cAMP response element-binding protein; ENF, ethyl
acetate fraction of Nypa fruticans Wurmb; ERK, extracellular
signal-regulated kinase; IBMX, isobutylmethylxanthine; mTOR,
mammalian target of rapamycin; p-, phosphorylated; PI3K,
phosphoinositide 3-kinase. |
Discussion
Melanin protects the dermis, hypodermis, and
epidermis from external stimuli; however, when produced in excess,
it causes various problems, such as hyperpigmentation, freckles,
and skin cancer (30). This study
revealed that ENF inhibits melanogenesis in B16F10 cells. ENF
treatment (25-100 µg/ml) decreased the melanin content at the
cellular level in a dose-dependent manner. TYR is known as a
rate-limiting enzyme essential for melanogenesis; a decrease in TYR
levels leads to the inhibition of melanin production (31,32).
Catechin, epicatechin, and isoquercitrin, which are phenolic
compounds, regulate melanogenesis by inhibiting TYR (33-35).
LC-MS analysis showed that ENF contains catechin (130 mg/g),
epicatechin (16.1 mg/g), and isoquercitrin (85.7 mg/g). A previous
study on catechin revealed its inhibitory effects on cell
proliferation in melanoma and TYR expression (above 5 µM) (36). Moreover, catechin and epicatechin
(each above 1.2 mg/ml) are known to inhibit the activities of
enzymes related to melanin biosynthesis (37). ENF suppresses melanin production by
regulating the expression of the proteins involved in melanogenesis
(TYR, TYRP-1, TYRP-2, and MITF). This inhibition of
melanogenesis-related factors can be attributed to the activities
of phenolic compounds, such as isoquercitrin, catechin, and
epicatechin. Isoquercitrin acts as a mediator that strongly
inhibits melanogenesis (half-maximal inhibitory concentration: 21.7
µM) by suppressing TYR expression at the cellular level (38). These studies suggest that the
levels of melanin, which is synthesized by TYR, are decreased by
the activity of the phytochemicals in ENF, such as catechin,
epicatechin, and isoquercitrin. In studies of other plants
(Pinellia pedatisecta, P. ternata, and Colocasia
affinis) belonging to the Araceae family, the bioactivity
derived from the phytochemicals in these plants were found to
regulate melanogenesis, inhibit cancer, and alleviate inflammation
(39-41).
The inhibitory effect of ENF on melanogenesis is related to
cellular signaling pathways associated with MITF, which is
controlled via PI3K/AKT/mTOR, ERK, and CREB (42). The inhibitory effect of ENF on
phosphorylation in the signaling pathways induced by IBMX was
confirmed based on the downregulation of the pathways by the
phytochemicals in NF (40). It is
assumed that this decrease in phosphorylation levels downregulates
MITF expression via cell signaling transduction and inhibits
melanin biosynthesis. Collectively, these findings indicated that
PI3K/AKT/mTOR and CREB phosphorylation were inhibited. Hence, MITF
inhibition via the downregulation of various signaling pathways is
considered the mechanism by which ENF suppresses melanogenesis
(Fig. 7). Although it is known
that there are more factors (43-46)
that exist in the mechanism of inhibiting melanogenesis, not
clarifying some factors for eliciting effects on melanogenesis is
considered a limitation of this study. However, by confirming the
mechanism by which ENF inhibits MITF, the potential seen
possibility that natural resources can be used academically and
industrially as materials for anti-melanogenesis agents, cosmetics,
food, and pharmaceuticals.
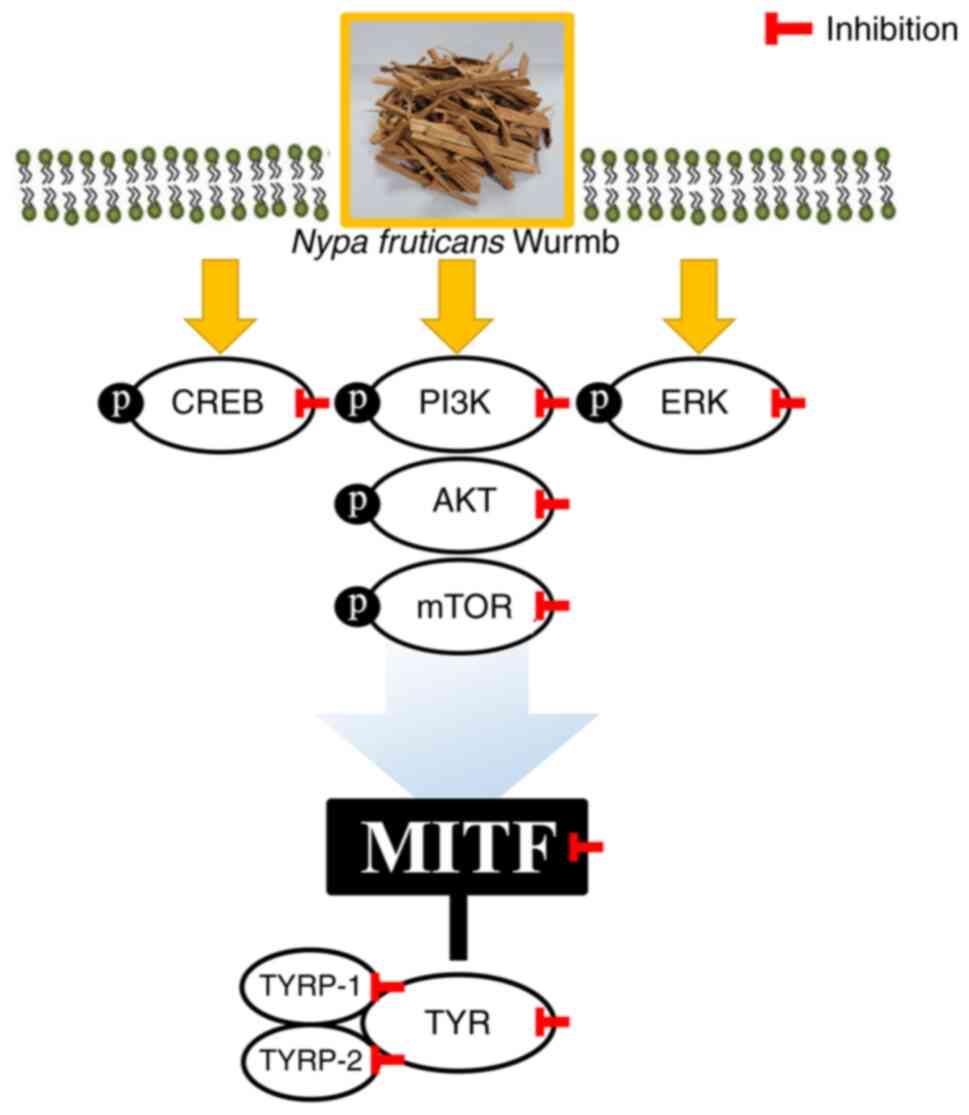 | Figure 7Effects of ENF on MITF inhibition.
MITF was inhibited through various signaling pathways. ENF, ethyl
acetate fraction of Nypa fruticans Wurmb; MITF,
microphthalmia-associated transcription factor; p-CREB,
phosphorylated cAMP response element-binding protein; p-PI3K,
phosphorylated phosphoinositide 3-kinase; p-ERK, phosphorylated
extracellular signal-regulated kinase; p-AKT, phosphorylated
protein kinase B; p-mTOR, phosphorylated mammalian target of
rapamycin; TYRP-1, TYR-related protein-1; TYRP-2, TYR-related
protein-2; TYR, tyrosinase. |
Acknowledgements
Not applicable.
Funding
Funding: No funding was received.
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
SYH and TWJ conceived the study and wrote the
manuscript. SYH, TWJ and HJP performed the experiments. SYH, TWJ,
JBL, SSO, SMM and JHP carried out the data collection and data
analysis. SYH, TWJ, JHP, HJP, JBL, SSO and SMM confirm the
authenticity of all the raw data. SYH, TWJ, JHP, HJP, JBL, SSO and
SMM reviewed the results. All authors have read and approved the
final manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Gilchrest BA and Eller MS: DNA photodamage
stimulates melanogenesis and other photoprotective responses. J
Investig Dermatol Symp Proc. 4:35–40. 1999.PubMed/NCBI View Article : Google Scholar
|
|
2
|
Swalwell H, Latimer J, Haywood RM and
Birch-Machin MA: Investigating the role of melanin in UVA/UVB-and
hydrogen peroxide-induced cellular and mitochondrial ROS production
and mitochondrial DNA damage in human melanoma cells. Free Radic
Biol Med. 52:626–634. 2012.PubMed/NCBI View Article : Google Scholar
|
|
3
|
Buscà R and Ballotti R: Cyclic AMP a key
messenger in the regulation of skin pigmentation. Pigment Cell
Melanoma Res. 13:60–69. 2000.PubMed/NCBI View Article : Google Scholar
|
|
4
|
Jiang Z, Xu J, Long M, Tu Z, Yang G and He
G: 2,3,5,4'-Tetrahyd-roxystilbene-2-O-β-d-glucoside (THSG) induces
melanogenesis in B16 cells by MAP kinase activation and tyrosinase
upregulation. Life Sci. 85:345–350. 2009.PubMed/NCBI View Article : Google Scholar
|
|
5
|
Ye Y, Chu JH, Wang H, Xu H, Chou GX, Leung
AK, Fong WF and Yu ZL: Involvement of p38 MAPK signaling pathway in
the anti-melanogenic effect of San-bai-tang, a Chinese herbal
formula, in B16 cells. J Ethnopharmacol. 132:533–535.
2010.PubMed/NCBI View Article : Google Scholar
|
|
6
|
Jung E, Lee J, Huh S, Lee J, Kim YS, Kim G
and Park D: Phloridzin-induced melanogenesis is mediated by the
cAMP signaling pathway. Food Chem Toxicol. 47:2436–2440.
2009.PubMed/NCBI View Article : Google Scholar
|
|
7
|
Han HJ, Park SK, Kang JY, Kim JM, Yoo SK
and Heo HJ: Anti-melanogenic effect of ethanolic extract of
Sorghum bicolor on IBMX-induced melanogenesis in B16/F10
melanoma cells. Nutrients. 12(832)2020.PubMed/NCBI View Article : Google Scholar
|
|
8
|
Bertolotto C, Abbe P, Hemesath TJ, Bille
K, Fisher DE, Ortonne JP and Bauotti R: Microphthalmia gene product
as a signal transducer in cAMP-induced differentiation of
melanocytes. J Cell Biol. 142:827–835. 1998.PubMed/NCBI View Article : Google Scholar
|
|
9
|
Park WS, Kwon O, Yoon TJ and Chung JH:
Anti-graying effect of the extract of Pueraria thunbergiana
via upregulation of cAMP/MITF-M signaling pathway. J Dermatol Sci.
75:153–155. 2014.PubMed/NCBI View Article : Google Scholar
|
|
10
|
Kang YG, Choi EJ, Choi Y and Hwang JK:
5,7-Dimethoxyflavone induces melanogenesis in B16F10 melanoma cells
through cAMP-dependent signalling. Exp Dermatol. 20:445–447.
2011.PubMed/NCBI View Article : Google Scholar
|
|
11
|
Jang JY, Lee JH, Kang BW, Chung KT, Choi
YH and Choi BT: Dichloromethane fraction of Cimicifuga
heracleifolia decreases the level of melanin synthesis by
activating the ERK or AKT signaling pathway in B16F10 cells. Exp
Dermatol. 18:232–237. 2009.PubMed/NCBI View Article : Google Scholar
|
|
12
|
Jang JY, Kim HN, Kim YR, Choi WY, Choi YH,
Shin HK and Choi BT: Partially purified components of
Nardostachys chinensis suppress melanin synthesis through
ERK and Akt signaling pathway with cAMP down-regulation in B16F10
cells. J Ethnopharmacol. 137:1207–1214. 2011.PubMed/NCBI View Article : Google Scholar
|
|
13
|
Kim DS, Jeong YM, Park IK, Hahn HG, Lee
HK, Kwon SB, Jeong JH, Yang SJ, Sohn UD and Park KC: A new
2-imino-1,3-thiazoline derivative, KHG22394, inhibits melanin
synthesis in mouse B16 melanoma cells. Biol Pharm Bull. 30:180–183.
2007.PubMed/NCBI View Article : Google Scholar
|
|
14
|
Cho JH, Robinson JP, Arave RA, Burnett WJ,
Kircher DA, Chen G, Davies MA, Grossmann AH, VanBrocklin MW,
McMahon M and Holmen SL: AKT1 activation promotes development of
melanoma metastases. Cell Rep. 13:898–905. 2015.PubMed/NCBI View Article : Google Scholar
|
|
15
|
Mantiquilla JA, Shiao MS, Lu HY, Sridith
K, Sidique SN, Liyanage WK, Chu YL, Shih HC and Chiang YC: Deep
structured populations of geographically isolated nipa (Nypa
fruticans Wurmb.) in the Indo-West Pacific revealed using
microsatellite markers. Frontiers in Plant Science 13, 2022.
|
|
16
|
Prasad N, Yang B, Kong KW, Sun J, Azlan A,
Ismail A and Romi ZB: Phytochemicals and antioxidant capacity from
Nypa fruticans Wurmb. fruit. Evid Based Complementary Altern
Med: Article ID 154606, 9 pages: 2013.
|
|
17
|
Kim D, Park J, Kim J, Han C, Yoon J, Kim
N, Seo J and Lee C: Flavonoids as mushroom tyrosinase inhibitors: A
fluorescence quenching study. J Agric Food Chem. 54:935–941.
2006.PubMed/NCBI View Article : Google Scholar
|
|
18
|
Kubo I and Kinst-Hori I: Flavonols from
saffron flower: Tyrosinase inhibitory activity and inhibition
mechanism. J Agric Food Chem. 47:4121–4125. 1999.PubMed/NCBI View Article : Google Scholar
|
|
19
|
Itoh K, Hirata N, Masuda M, Naruto S,
Murata K, Wakabayashi K and Matsuda H: Inhibitory effects of
Citrus hassaku extract and its flavanone glycosides on
melanogenesis. Biol Pharm Bull. 32:410–415. 2009.PubMed/NCBI View Article : Google Scholar
|
|
20
|
Rodboon T, Okada S and Suwannalert P:
Germinated riceberry rice enhanced protocatechuic acid and vanillic
acid to suppress melanogenesis through cellular oxidant-related
tyrosinase activity in B16 cells. Antioxidants.
9(247)2020.PubMed/NCBI View Article : Google Scholar
|
|
21
|
Kang MS, Lee GH, Choi GE, Yoon HG and Hyun
KY: Neuroprotective effect of Nypa fruticans wurmb by
suppressing TRPV1 following sciatic nerve crush injury in a rat.
Nutrients. 12(2618)2020.PubMed/NCBI View Article : Google Scholar
|
|
22
|
Kang MS and Hyun KY: Antinociceptive and
anti-inflammatory effects of Nypa fruticans wurmb by
suppressing TRPV1 in the sciatic neuropathies. Nutrients.
12(135)2020.PubMed/NCBI View Article : Google Scholar
|
|
23
|
Jang TW and Park JH: Anti-inflammatory
effects of Abeliophyllum distichum nakai (Cultivar Okhwang
1) callus through inhibition of PI3K/Akt, NF-κB and MAPK signaling
pathways in lipopolysaccharide-induced macrophages. Processes.
9(1071)2021.
|
|
24
|
Son KH, Baek JK, Park SB, Kin HN, Park GH,
Son HJ, Eo HJ, Song JH, Jeong HJ and Jeong JB: Enhancement of
melanin synthesis by the branch extracts of Vaccinium
oldhamii through activating tyrosinase activity in B16F10
melanoma cells. Korean J Plant Res. 31:547–553. 2018.
|
|
25
|
Jin KS, Oh YN, Hyun SK, Kwon HJ and Kim
BW: Betulinic acid isolated from Vitis amurensis root
inhibits 3-isobutyl-1-methylxanthine induced melanogenesis via the
regulation of MEK/ERK and PI3K/Akt pathways in B16F10 cells. Food
Chem Toxicol. 68:38–43. 2014.PubMed/NCBI View Article : Google Scholar
|
|
26
|
Xie X, White EP and Mehnert JM: Coordinate
autophagy and mTOR pathway inhibition enhances cell death in
melanoma. PLoS One. 8(e55096)2013.PubMed/NCBI View Article : Google Scholar
|
|
27
|
Werzowa J, Koehrer S, Strommer S, Cejka D,
Fuereder T, Zebedin E and Wacheck V: Vertical inhibition of the
mTORC1/mTORC2/PI3K pathway shows synergistic effects against
melanoma in vitro and in vivo. J Invest Dermatol. 131:495–503.
2011.PubMed/NCBI View Article : Google Scholar
|
|
28
|
Karbowniczek M, Spittle CS, Morrison T, Wu
H and Henske EP: mTOR is activated in the majority of malignant
melanomas. J Invest Dermatol. 128:980–987. 2008.PubMed/NCBI View Article : Google Scholar
|
|
29
|
Meier F, Schittek B, Busch S, Garbe C,
Smalley K, Stayamoorthy K, Li G and Herdyn M: The RAS/RAF/MEK/ERK
and PI3K/AKT signaling pathways present molecular targets for the
effective treatment of advanced melanoma. Front Biosci.
10:2986–3001. 2005.PubMed/NCBI View
Article : Google Scholar
|
|
30
|
Costin GE and Hearing VJ: Human skin
pigmentation: Melanocytes modulate skin color in response to
stress. FASEB J. 21:976–994. 2007.PubMed/NCBI View Article : Google Scholar
|
|
31
|
D'Mello SA, Finlay GJ, Baguley BC and
Askarian-Amiri ME: Signaling pathways in melanogenesis. Int J Mol
Sci. 17(1144)2016.PubMed/NCBI View Article : Google Scholar
|
|
32
|
Boissy RE, Visscher M and DeLong MA:
DeoxyArbutin: A novel reversible tyrosinase inhibitor with
effective in vivo skin lightening potency. Exp Dermatol.
14:601–608. 2005.PubMed/NCBI View Article : Google Scholar
|
|
33
|
Bloom van Staden A, Oosthuizen CB and Lall
N: The effect of Aspalathus linearis (Burm. f.) R. Dahlgren
and its compounds on tyrosinase and melanogenesis. Sci Rep.
11(7020)2021.PubMed/NCBI View Article : Google Scholar
|
|
34
|
Zhang X, Li J, Li Y, Liu Z, Lin Y and
Huang JA: Anti-melanogenic effects of epigallocatechin-3-gallate
(EGCG), epicatechin-3-gallate (ECG) and gallocatechin-3-gallate
(GCG) via down-regulation of cAMP/CREB/MITF signaling pathway in
B16F10 melanoma cells. Fitoterapia. 145(104634)2020.PubMed/NCBI View Article : Google Scholar
|
|
35
|
Ramli S and Ruangrungsi N: Tyrosinase
inhibition, antioxidant activity and total phenolic content of
selected mimosaceae pericarps ethanolic extracts. Int J Pharm.
4:47–57. 2021.
|
|
36
|
Sato K and Toriyama M: Depigmenting effect
of catechins. Molecules. 14:4425–4432. 2009.PubMed/NCBI View Article : Google Scholar
|
|
37
|
Chen Z, Liang J, Zhang C and Rodrigues CJ
Jr: Epicatechin and catechin may prevent coffee berry disease by
inhibition of appressorial melanization of Colletotrichum
kahawae. Biotechnol Lett. 28:1637–1640. 2006.PubMed/NCBI View Article : Google Scholar
|
|
38
|
Ohguchi K, Nakajima C, Oyama M, Iinuma M,
Itoh T, Akao Y, Nozawa Y and Ito M: Inhibitory effects of flavonoid
glycosides isolated from the peel of Japanese persimmon
(Diospyros kaki ‘Fuyu’) on melanin biosynthesis. Biol Pharm
Bull. 33:122–124. 2010.PubMed/NCBI View Article : Google Scholar
|
|
39
|
Balogun TA, Ipinloju N, Abdullateef OT,
Moses SI, Omoboyowa DA, James AC, Saibu OA, Akinyemi WF and Oni EA:
Computational evaluation of bioactive compounds from Colocasia
affinis schott as a novel EGFR inhibitor for cancer treatment.
Cancer Inform. 20(11769351211049244)2021.PubMed/NCBI View Article : Google Scholar
|
|
40
|
Wang W, Mao S, Yu H, Wu H, Shan X, Zhang
X, Cui G and Liu X: Pinellia pedatisecta lectin exerts a
proinflammatory activity correlated with ROS-MAPKs/NF-κB pathways
and the NLRP3 inflammasome in RAW264.7 cells accompanied by cell
pyroptosis. Int Immunopharmacol. 66:1–12. 2019.PubMed/NCBI View Article : Google Scholar
|
|
41
|
An JH, Won HJ, Seo SK, Kim DY, Ku CS, Oh
SK and Ryu HW: Utilization of [6]-gingerol as an origin
discriminant marker influencing melanin inhibitory activity
relative to its content in Pinellia ternata. J Appl Biol Chem.
59:323–330. 2016.
|
|
42
|
Byun EB, Song HY, Mushtaq S, Kim HM, Kang
JA, Yang MS, Sung NY, Jang BS and Byung EH: Gamma-irradiated
luteolin inhibits 3-isobutyl-1-methylxanthine-induced melanogenesis
through the regulation of CREB/MITF, PI3K/Akt and ERK pathways in
B16BL6 melanoma cells. J Med Food. 20:812–819. 2017.PubMed/NCBI View Article : Google Scholar
|
|
43
|
Hsiao JJ and Fisher DE: The roles of
microphthalmia-associated transcription factor and pigmentation in
melanoma. Arch Biochem Biophys. 563:28–34. 2014.PubMed/NCBI View Article : Google Scholar
|
|
44
|
Du J, Widlund HR, Horstmann MA, Ramaswamy
S, Ross K, Huber WE, Nishimura EK, Golub TR and Fisher DE: Critical
role of CDK2 for melanoma growth linked to its melanocyte-specific
transcriptional regulation by MITF. Cancer Cell Int. 6:565–576.
2004.PubMed/NCBI View Article : Google Scholar
|
|
45
|
Buscà R, Berra E, Gaggioli C, Khaled M,
Bille K, Marchetti B, Thyss R, Fitsialos G, Larribère L,
Bertolotto C, et al: Hypoxia-inducible factor 1α is a new target of
microphthalmia-associated transcription factor (MITF) in melanoma
cells. J Cell Biol. 170:49–59. 2005.PubMed/NCBI View Article : Google Scholar
|
|
46
|
McGill GG, Haq R, Nishimura EK and Fisher
DE: c-Met expression is regulated by Mitf in the melanocyte
lineage. J Biol Chem. 281:10365–10373. 2006.PubMed/NCBI View Article : Google Scholar
|