Introduction
Human hepatitis B virus (HBV) infection is strongly
associated with the development of hepatocellular carcinoma (HCC)
(1). It contains a unique open
reading frame, the sequence of which is highly conserved among
different mammalian hepadnaviruses and codes for a 16.5-kDa protein
known as hepatitis B virus X protein (HBx). The mechanisms through
which HBV causes malignant transformation remain unelucidated;
however, available evidence supports a pathogenetic role for the
product of the HBV X gene, HBx (2). A large number of putative mechanisms
through which HBx may contribute to the development of HCC have
been investigated. Studies on HBx-responsive elements have
indicated that HBx transactivates viral and cellular genes through
transcriptional regulatory factors, such as nuclear factor (NF)-κB,
activator protein (AP)-1, AP-2 and cAMP response element binding
protein (CREB). It also interacts with a group of transcriptional
co-activators that includes TATA-binding protein-associated factors
(3,4). However, the molecular mechanisms
responsible for the effects of HBx on transcription, cellular
proliferation and transformation are only partially defined. As HBx
does not have the ability to bind to double-stranded DNA (dsDNA),
protein-protein interaction is crucial for HBx functions. The
interactions of HBx with cellular proteins may trigger a cascade of
phosphorylation and dephosphorylation events that lead to a general
upregulation of gene expression (5). The identification of cellular
proteins that interact with HBx may provide insight into the
mechanisms through which HBV exerts its cellular effects. In our
previous study, using the Saccharomyces cerevisiae
two-hybrid system, we found a novel HBx-interacting protein that is
homologous to the Homo sapiens cytochrome c oxidase
III (COXIII) (6). In the present
study, once again using the yeast two-hybrid system, we took two
fragments of the HBV X gene mutants, that are translated as HBx X1
aa1-72 and X2 aa1-117, inserted them separately into plasmids, and
screened them to identify the fragment that interacts with COXIII.
The results indicated that the binding site for the interaction
between HBx and COXIII was located between aa72-117. The data
presented herein may form the basis for future studies on HBx
interactive elements and may provide insight into the mechanisms
through which HBx causes malignant transformation in HCC.
Materials and methods
Materials
The Saccharomyces cerevisiae AH109 yeast
strain was grown in YPD medium (10 g/l yeast extract, 20 g/l
peptone and 20 g/l dextrose) or synthetic minimal medium (0.67%
yeast nitrogen base, 2% dextrose and appropriate auxotrophic
supplements) following transfection. The media were from Clontech
Laboratories, Inc. (Mountain View, CA, USA). The yeast strain
carries the LacZ, HIS3 and ADE2 reporter genes
under the control of the Gal4 binding site and has deletions in the
trp and leu genes. Y187-pACT2-COXIII
(Saccharomyces cerevisiae Y187 yeast strain with a
LacZ reporter gene and deletions in the trp and
leu genes, carrying pACT2 expressing the full-length COXIII
gene) was obtained from our institution. The plasmids, pAS2-1
(which includes the binding domain for the detection of the fusion
protein), pCL1, pUCm-T, pCMV-HA and pLAM5′-1, were from Clontech
Laboratories, Inc.. pcDNA3-X, pAS2-1-X, pCMV-Myc-COXIII, pCMV-HA-X
and pUCm-T-X were constructed and maintained at our institution
(6). The binding domain and c-Myc
monoclonal (Cat. no. 631206) and hemagglutinin (HA) polyclonal
(Cat. no. 631207) antibodies were also provided by Clontech
Laboratories, Inc.. The mouse anti-human β-actin antibody was from
Santa Cruz Biotechnology, Inc. (Dallas, TX, USA). The EcoRI,
XhoI and PstI restriction enzymes, T4 DNA ligase, Taq
DNA polymerase, dNTP, the Plasmid DNA purification system and the
TransFast transfection reagent kit were from Promega Biosciences
LLC (San Luis Obispo, CA, USA). The DNA ladder was from Life
Technologies Corp. (Carlsbad, CA, USA). The glass beads were from
Sigma-Aldrich Corp. (St. Louis, MO, USA). The DNA gel extraction
kit was from Shanghai Sangon Biotech (Shanghai, China). The
electrochemiluminescence kit was from Boster Biotech (Wuhan,
China).
Methods
Plasmid construction
Escherichia coli (E. coli) DH5α cells
(Shenggong Corporation, Shanghai, China) containing pcDNA3-X were
cultured overnight on an LB plate containing 50 μg/ml
ampicillin. Single colonies were selected and inoculated into 3 ml
of LB broth containing 50 μg/ml ampicillin and incubated for
8 h at 37°C; subsequently, the broth was examined for cloudiness,
which signifies bacterial growth. The preparation and purification
of pcDNA3-X were carried out according to the manufacturer’s
instructions using the Plasmid DNA purification system (Promega
Biosciences LLC).
HBx primers were designed using sequences obtained
from the National Center for Biotechnology Information (NCBI)
containing the EcoRI and PstI sites. The sequences of
the primers with the restriction enzyme sites underlined are as
follows: P1, 5′-CGGAATTCGCCACCATGGCTGCTAGGCTGTGC-3′;
P2, 5′-CCCCTGCAGTTAGCGAAGTGCACACGGTCC-3′;
P3, 5′-CCCCTGCAGTTACCAGTCTTTAAACAAACAGTC-3′;
and P4, 5′-CCCCTGCAGAGATGATTAGGCAGAGGTGA-3′.
The primer pairs, P1/P2, P1/P3 and P1/P4, were used
to amplify HBx X1, HBx X2 and the full-length HBx gene,
respectively. The thermocycling conditions for polymerase chain
reaction (PCR) were as follows: initial denaturation at 94°C for 5
min; 30 cycles of denaturation at 94°C for 45 sec, annealing at
55°C for 45 sec and extension at 72°C for 1 min; followed by a
final extension at 72°C for 10 min. The PCR products were separated
by electrophoresis on a 1.3% agarose gel. Subsequently, 216-bp (X1)
and 352-bp (X2) fragments were collected and purified using the DNA
gel extraction kit following the manufacturer’s instructions
(Shanghai Sangon Biotech).
The pAS2-1 plasmid was extracted from E. coli
DH5α cells using the Plasmid DNA purification system, following the
manufacturer’s instructions (Promega Biosciences LLC). Following
double digestion with EcoRI and PstI, the pAS2-1
fragments were separated by electrophoresis in a 0.6% agarose gel.
Fragments with a size of 8,400 bp were collected and purified using
the DNA gel extraction kit.
The extracted 8400-bp fragment was ligated with the
X1 or X2 fragments using T4 DNA ligase overnight at 4°C. The
recombinant plasmids were transfected into the E. coli DH5α
cells and cultured overnight at 37°C before plating on LB medium
containing 50 μg/ml ampicillin and were again cultured
overnight at 37°C. Colonies on the LB plate were selected and
inoculated into 3 ml of LB broth containing 50 μg/ml
ampicillin, incubated for 8 h at 37°C and examined for cloudiness
in the broth, which signifies bacterial growth. The plasmids were
extracted, purified and sent to Life Technologies Corp., (Shanghai,
China), for sequencing. The recombinant plasmids were designated as
pAS2-1-X1 and pAS2-1-X2. The X mutant fragments were also cloned
into the pUCm-T vector, creating pUCm-T-X1 and pUCm-T-X2. The X
mutant fragments were excised from pUCm-T-X1 and pUCm-T-X2 using
the EcoRI and XhoI restriction enzymes and ligated
into the pCMV-HA vector that had been linearized with the same
enzymes.
Plasmid transfection and translation of
fusion proteins in yeast cells
The screening procedure used in this study was a
modification of the method previously described by Gietz et
al (7). The lithium acetate
method (7) was used to transfect
Saccharomyces cerevisiae AH109 with the pAS2-1 plasmids
(pAS2-1-X1, pAS2-1-X2, pAS2-1-X and pAS2-1), as well as pCL1 and
pLAM5′-1 plasmids into the cells. Transfectants were grown either
in YPD medium (10 g/l yeast extract, 20 g/l peptone and 20 g/l
dextrose) or synthetic minimal medium (0.67% yeast nitrogen base,
2% dextrose and appropriate auxotrophic supplements) at 30°C. The
plasmids, pCL1 and pLAM5′-1, were included as positive and negative
controls, respectively. The transfectants were plated on
SC/-trp-leu-his medium selective for histidine, leucine and
tryptophan prototrophy. The AH109 transfectants were designated as
AH109-pAS2-1-X1, AH109-pAS2-1-X2, AH109-pAS2-1-X, AH109-pAS2-1,
AH109-pLAM5′-1 and AH109-pCL1. The X1 genes in AH109-pAS2-1-X1 and
AH109-pAS2-1-X2 were detected by PCR. The X1 and X2 binding domain
(X1-BD and X2-BD) fusion proteins were detected in the AH109 strain
by western blot analysis. Cell lysates were blocked with non-fat
dried milk and exposed to a 1:3,000 dilution of binding domain (BD)
monoclonal antibody at 4°C overnight. After washing with
Tris-buffered saline/Tween-20, the secondary antibody, alkaline
phosphatase-conjugated goat anti-rabbit IgG, was added followed by
incubation for 2 h at room temperature. Proteins were visualized
with 5-bromo-4-chloro-3-indolyl phosphate and nitro blue
tetrazolium. In addition, freshly grown yeast transformant colonies
were assayed for β-galactosidase (β-gal) activity by replica
plating onto Whatman filter papers that were snap-frozen twice for
10 sec in liquid nitrogen and incubated at 30°C for 8 h in a buffer
containing 5-bromo-4-chloro-3-indolyl-β-D-galactopyranoside
solution. Positive interactions were detected by the appearance of
blue colonies.
Mating assays
Y187-pACT2-COXIII single colonies were streaked as
parallel lines on SC/-leu medium at 30°C for 2 days, as were
the AH109-pAS2-1-X1, AH109-pAS2-1-X2, AH109-pAS2-1-X, AH109-pAS2-1,
AH109-pLAM5′-1 and AH109-pCL1 colonies on SC/-trp medium.
The two types of transformants were then mated and incubated in YPD
medium at 30°C for 4 h. Finally, the transformants were subcultured
on SC/-leu-trp medium, incubated at 30°C for 2 days, and
assayed for LacZ activity. Freshly grown colonies on
SC/-leu-trp medium were selected and subcultured on the same
medium. β-gal activity was assayed by replica plating of the yeast
transformants onto Whatman filter papers. The filters were
snap-frozen twice for 10 sec in liquid nitrogen and incubated at
30°C for 8 h in buffer containing
5-bromo-4-chloro-3-indolyl-β-D-galactopyranoside solution. Positive
interactions were detected by the appearance of blue colonies.
Transient transfection,
co-immunoprecipitation and western blot analysis
The transient transfection of COS7 cells (Typical
Culture Preservation Commission Cell Bank, Chinese Academy of
Sciences, Shanghai, China) was used to detect the ectopic
expression of the HBV X1, HBV X2 and COXIII proteins. The COS7
cells were grown in DMEM (90% Dulbecco’s modified Eagle’s medium,
10% fetal bovine serum); 5×106 COS7 cells were
transfected with 10 μg of plasmid DNA using the liposome
TransFast transfection reagent kit (Promega Biosciences LLC).
Forty-eight hours after transfection, the cells were washed 3 times
with ice-cold phosphate-buffered saline (PBS) and lysed with lysis
buffer supplemented with the protease inhibitors provided with the
Protein G Immunoprecipitation kit (Promega Biosciences LLC).
Appropriate protein expression was verified by gel electrophoresis
of cellular lysates and immunoblotting with specific antibodies. To
prevent N-linked glycosylation during protein synthesis, the cells
were treated with tunicamycin for 2 h prior to transfection and the
tunicamycin level was maintained during the transient transfection
period. c-Myc monoclonal antibody was used to recognize the
expression of the tagged Myc-COXIII protein. For
co-immunoprecipitation, 1 ml of cell lysate was incubated for 3 h
at 4°C with 50 μl of a 50% suspension of protein G agarose
that had been pre-coated with 3 μg of c-Myc antibody. The
immunocomplex was washed 3 times with ice-cold lysis buffer and
resuspended in sodium dodecyl-sulfate polyacrylamide gel
electrophoresis (SDS-PAGE) sample buffer; electrophoresis was
performed, and the proteins were electroblotted onto a
nitrocellulose membrane. The blotting membrane was blocked with
non-fat dried milk and exposed to a 1:200 dilution of HA polyclonal
antibody for 1 h. After washing with Tris-buffered saline Tween-20,
the proteins on the nitrocellulose membrane were detected using an
electrochemiluminescence kit (Boster Biotech) that included a goat
anti-rabbit antibody, following the manufacturer’s instructions.
The COS7 cells transfected with the pCMV-Myc vector or with the
pCMV-HA-X vector and non-transfected cells were used as negative
controls; COS7 cells transfected with the pCMV-Myc-COXIII or
pCMV-HA-X vector were used as positive controls.
Results
Plasmid construction
The HBx X1 and X2 amplified fragments were ligated
with pAS2-1 following a double restriction enzyme digestion. The
recombinant plasmids were designated as pAS2-1-X1 and pAS2-1-X2.
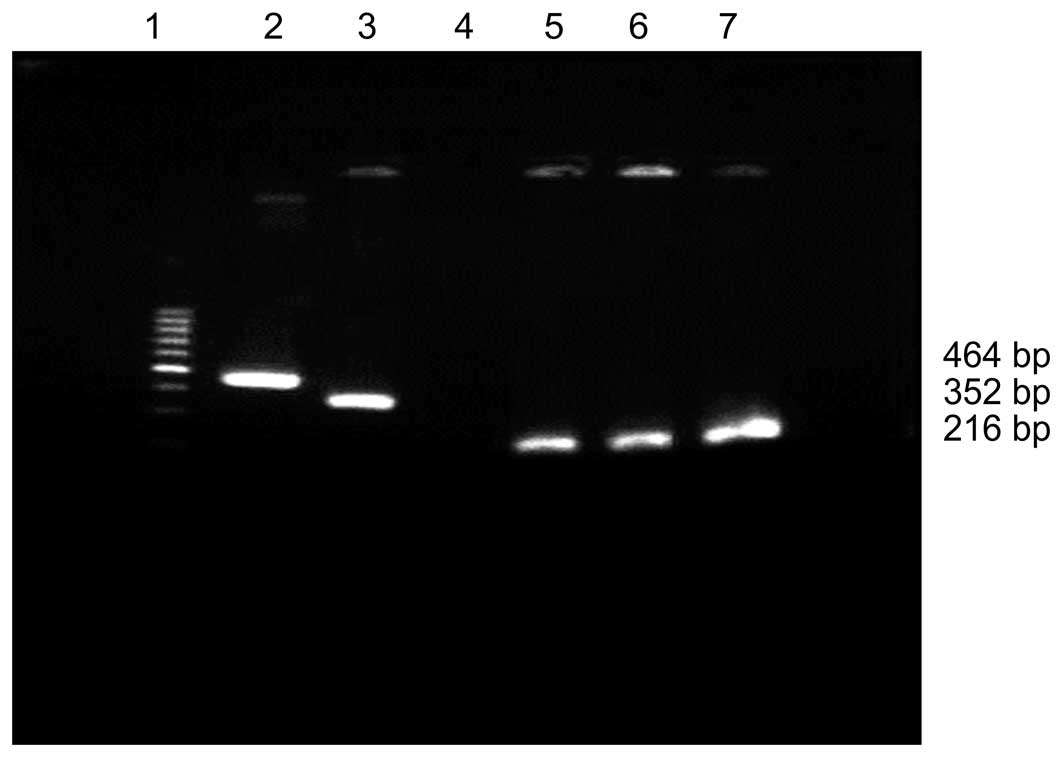
The PCR-amplified fragments from pAS2-1-X1, pAS2-1-X2 and pAS2-1-X
were separated electrophoretically on a 0.6% agarose gel (Fig. 1).
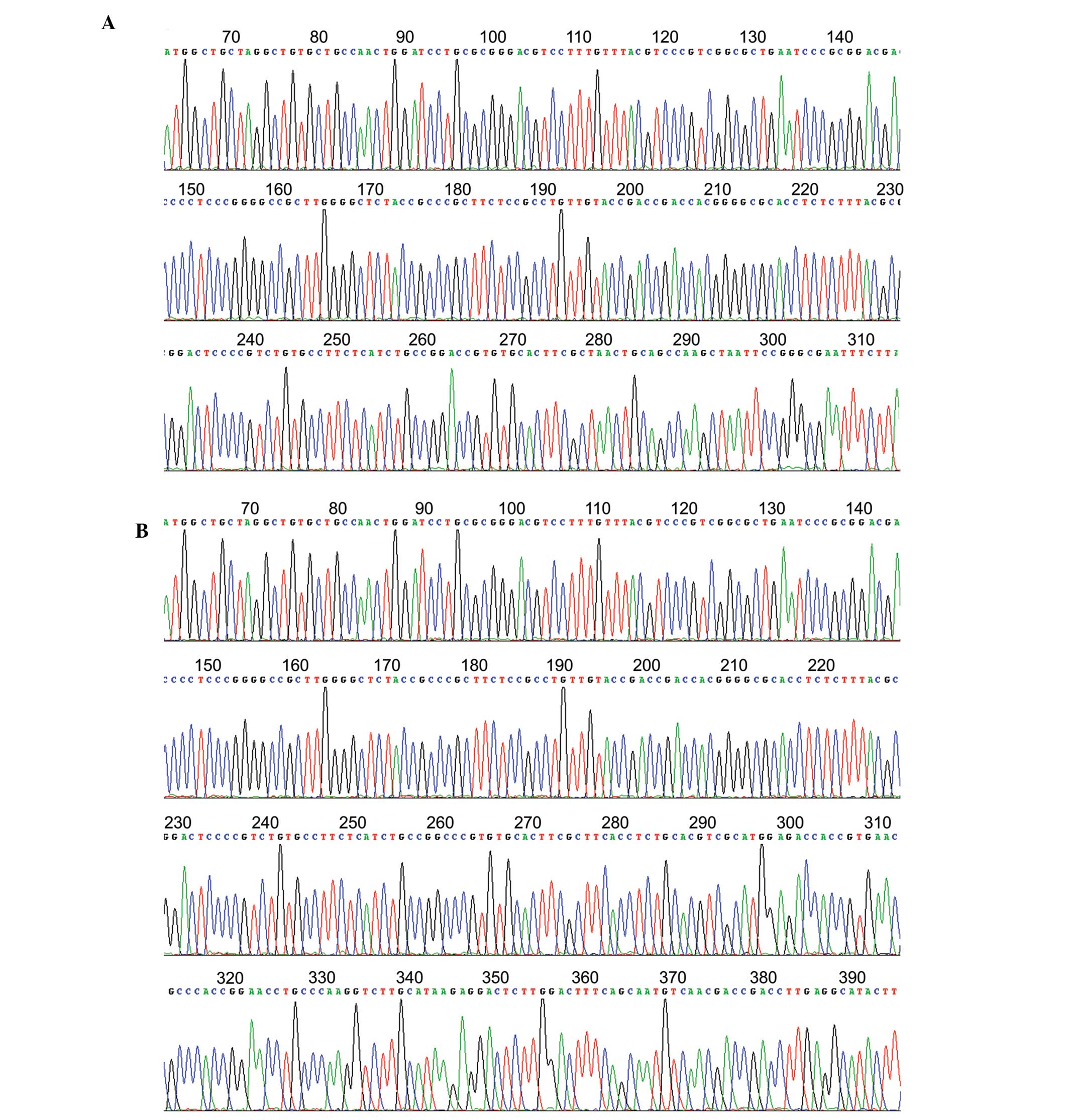
Sequencing of HBx X1 in pAS2-1-X1 and HBx
X2 in pAS2-1-X2
The amplified HBx X1 and X2 fragments were ligated
with pAS2-1 after a double restriction enzyme digestion. The
recombined plasmids were designated as pAS2-1-X1 and pAS2-1-X2.
Purified recombined plasmids were sent to Life Technologies Corp.,
Shanghai Branch for sequencing. The HBx X1 and X2 genes were 98%
homologous with the X gene from GenBank (Fig. 2).
Expression of fusion proteins in yeast
cells
The pAS2-1-X1 and pAS2-1-X2 recombinant plasmids
were transfected into the AH109 yeast cells, creating
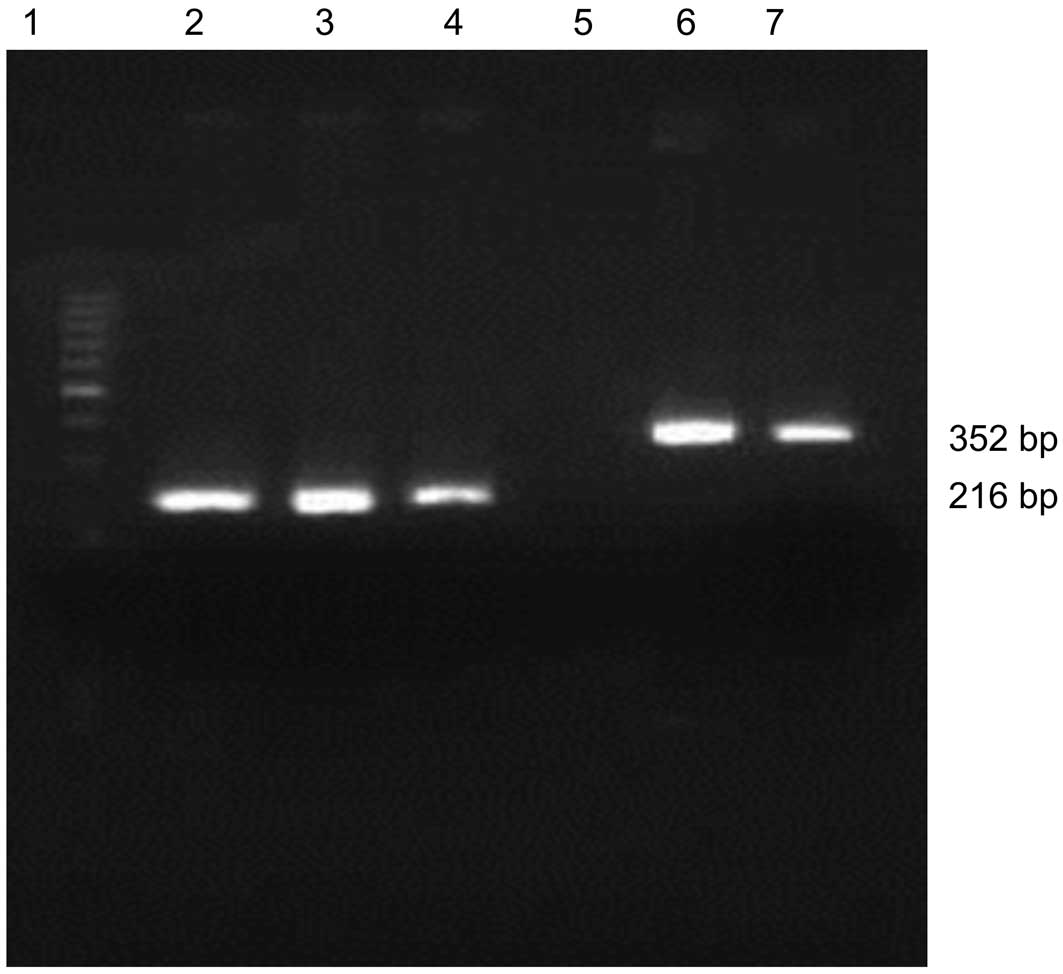
AH109-pAS2-1-X1 and AH109-pAS2-1-X2, respectively. The presence of
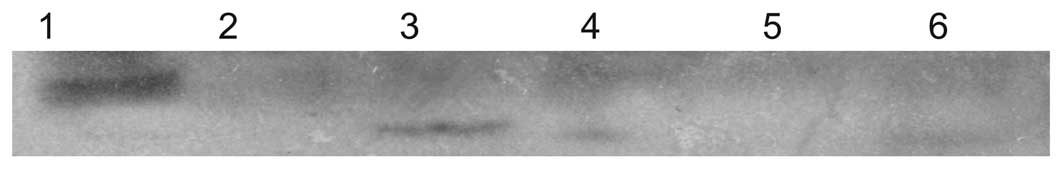
HBx X1 and X2 was detected by PCR (Fig. 3). The expression of the two fusion
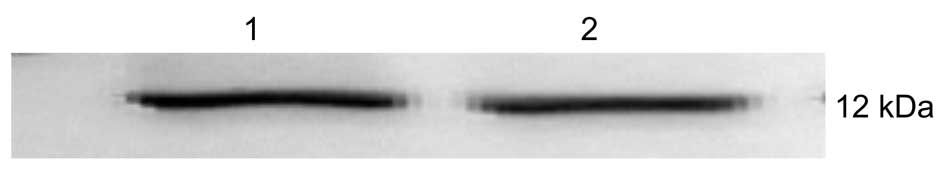
proteins and the X1 and X2 binding domains (X1-BD and X2-BD) in the
AH109 strain was detected by western blot analysis (Fig. 4).
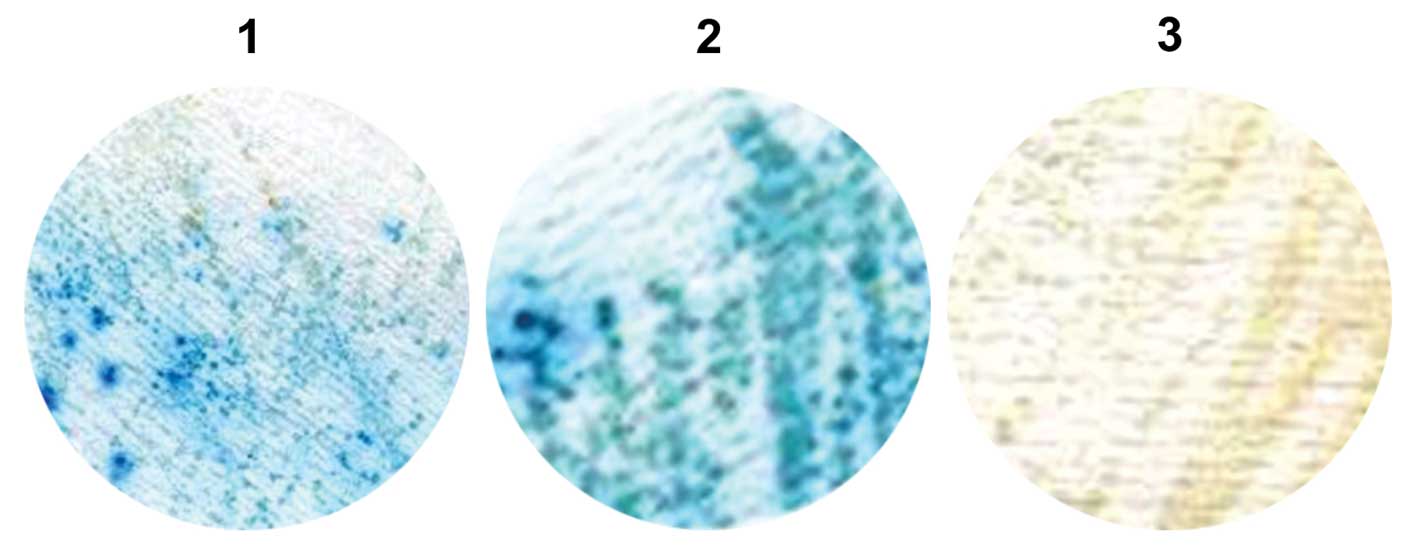
Detection of β-gal activity
Following the elimination of the auto-activation of
X1-BD and X2-BD in AH109, freshly grown clones on an
SC/-leu-trp medium were selected and subcultured on the same
medium. β-gal activity was assayed by replica plating of the yeast
transformants onto Whatman filter papers. Positive interactions
were detected by the appearance of blue colonies (Fig. 5).
Co-immunoprecipitation and western blot
analysis for detecting interactions between HBx X1 or HBx X2
proteins with COXIII in COS7 cells
COS7 cells co-transformed using the plasmids
pCMV-HA-X, pCMV-HA-X1, pCMV-HA-X2 or pCMV-Myc-COXIII or with the
pCMV-HA-X and pCMV-Myc plasmids, were extracted and
immunoprecipitated with Myc antibody. A protein of approximately 17
kDa (the predicted size of HA-X) was co-immunoprecipitated with Myc
antibody from cells containing HA-X and Myc-COXIII fusion protein,
and a protein of approximately 14 kDa (the predicted size of HA-X2)
was co-immunoprecipitated with Myc antibody from cells containing
HA-X2 and Myc-COXIII fusion protein, but no co-immunoprecipitation
was observed from the cells co-transformed with pCMV-HA-X1 and
pCMV-Myc-COXIII or with pCMV-HA-X and pCMV-Myc. The HA-X and HA-X 2
proteins were recognized by immunoblotting with HA antibody. These
results confirm that aa72-117 of HBx include the key peptide(s) for
the combination of HBx and COXIII (Fig. 6).
Discussion
The HBV genome contains four functionally distinct
open reading frames on one strand in the same transcriptional
polarity. The X gene is the smallest, coding for a regulatory
protein of 154 aa, and is known to exhibit a transcriptional
transactivation function through the X-responsive elements of many
viral and cellular genes implicated in the carcinogenicity of this
virus (8,9). One of the basic properties of HBx is
that it has no DNA binding activity; therefore, its transactivation
and/or oncogenic function is possibly exerted through
interaction(s) with cellular protein(s) (2). HBx is predominantly localized in the
cytoplasm and its presence there is required for it to function
(2,10); therefore, the direct cellular
target of HBx may also be located in the cytoplasm. The
identification of this target represents a major goal in defining
the functions of HBx in HBV replication and HCC. In our previous
study (6), we detected and cloned
a novel HBx-interactive protein from a normal human liver cDNA
library. Sequence analysis revealed that it was a homolog of
Homo sapiens COXIII. In addition, using confocal microscopy,
our laboratory has recently demonstrated that HBx co-localizes with
inner mitochondrial protein COXIII in HL7702 cells (11).
There is considerable evidence that HBx is involved
in the dysfunction of cytochrome c oxidase (10,12), the terminal enzyme of the
mitochondrial respiratory chain that catalyzes the transfer of
electrons from reduced cytochrome c to molecular oxygen.
COXIII, which is encoded by mitochondrial DNA and is synthesized
within the mitochondria, is a large COX subunit and acts as the
catalytic core of the enzyme (13). COXIII is a 7 transmembrane helix
protein and binds to subunit I. The N-terminal region of subunit
III is adjacent to D132 of subunit I, the initial proton acceptor
of the D pathway that transfers protons from the protein surface to
the buried active site located 30 Å away (14). One of the roles of subunit III in
the normal oxidase is to maintain rapid proton uptake through the D
pathway at a physiological pH (14). Single-turnover experiments have
indicated that proton uptake through the D pathway at pH 8.0 is
reduced from >10,000 s−1 in the presence of subunit
III to 350 s−1 in its absence. Part of a proton antenna
for the D pathway may be lost upon the removal of subunit III
(15). Based on these facts, when
the proton-transferring function of COXIII is inhibited, the
electrochemical gradient across the inner mitochondrial membrane
decreases and triggers the opening of the mitochondrial
permeability transition pore (MPTP), which results in the elevation
of cytosolic calcium levels. This causes calcium overload, leading
to mitochondrial disruption and hepatic cell injury (16,17). Since there is a long time gap
(chronic infection) between HBV infection and liver carcinogenesis,
the chronic hepatic cell injury caused by HBx should be focused on
to a greater extent in future studies.
Identification of the precise location for the
interaction between HBx and COXIII is crucial for clarifying the
precise mechanisms involved in HBx co-localization with the
mitochondria. A genetic approach, the yeast two-hybrid system, is a
powerful method for mapping the essential binding sites of proteins
of interest. In the present study, we successfully constructed two
plasmids that produce mutants of HBx X1 (aa1-72) or HBx X2
(aa1-117). The interaction of HBx with COXIII was identified using
the yeast two-hybrid system. That this binding is specific for HBx
as supported by the histidine-independent growth and blue-colony
formation in the β-gal assay by yeast cells harboring both
pAS-2-1-X2 and pACT2-COXIII recombinant plasmids, as well as by the
behavior of the cells in false-positive elimination tests.
Furthermore, the outcome of co-immunoprecipitation experiments
confirmed the specificity of the interaction between HBx X2 mutant
proteins and COXIII. From these data, it is reasonable to conclude
that aa72-117 of HBx encode the peptides essential for HBx to bind
with COXIII.
Among mammalian hepadnavirus genomes, there are many
conserved HBx regions, and these seem to play an important role in
HBx function. For example, Misra et al (18) demonstrated that the conserved
amino-terminal region (aa1-20) of HBx has a transrepression
function, and Kumar et al (19) demonstrated that a truncated mutant
(residues, aa 58-140) of HBx effecientlyh stimulates the Rous
sarcoma virus long terminal repeat (RSV-LTR). It has also been
demonstrated that aa68-117 in HBx are sufficient for HBx
localization in the mitochondria and cell death induction, which is
consistent with our conclusions, and this effect is independent of
its transactivation activity (12). In studies investigating HBx
function, it is important to map the site of its interaction with
the mitochondria. In the present study, our findings support the
view that aa72-117 of HBx form the key domain for the combination
of HBV with COXIII, which may play a crucial role in the function
of HBV in the mitochondria.
In conclusion, our data led us to the hypothesis
that HBx may impair the mitochondrial respiration chain and energy
metabolism through the association between HBx and COXIII. Future
investigation are required to focus on the physiological
significance of HBx interactions with COXIII and make the peptides
in aa72-117 of HBx a novel target for further research on and in
the treatment of tumorigenesis in HCC.
Acknowledgments
The present study was supported by grants from the
National Natural Science Foundation of China (81300321), the Key
Discipline Foundation of Fujian Province (2012–149) and Young and
Middle-Aged Personnel Training Project of Fujian Province Health
Department (2014-ZQN-ZD-9).
References
|
1
|
Hakami A, Ali A and Hakami A: Effects of
hepatitis B virus mutations on its replication and liver disease
severity. Open Virol J. 7:12–18. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Koike K: Hepatitis B virus X gene is
implicated in liver carcinogenesis. Cancer Lett. 286:60–68. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Zhou Y, Wang S, Ma JW, et al: Hepatitis B
virus protein X-induced expression of the CXC chemokine IP-10 is
mediated through activation of NF-kappaB and increases migration of
leukocytes. J Biol Chem. 285:12159–12168. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Shamay M, Barak O, Doitsh G, et al:
Hepatitis B virus pX interacts with HBXAP, a PHD finger protein to
coactivate transcription. J Biol Chem. 277:9982–9988. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Yeh CT: Hepatitis B virus X protein:
searching for a role in hepatocarcinogenesis. J Gastroenterol
Hepatol. 15:339–341. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Wang XZ, Li D, Tao QM, et al: A novel
hepatitis B virus X-interactive protein: cytochrome C oxidase III.
J Gastroenterol Hepatol. 21:711–715. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Gietz RD, Triggs-Raine B, Robbins A, et
al: Identification of proteins that interact with a protein of
interest: applications of the yeast two-hybrid system. Mol Cell
Biochem. 172:67–79. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Aufiero B and Schneider RJ: The hepatitis
B virus X-gene product trans-activates both RNA polymerase II and
III promoters. EMBO J. 9:497–504. 1990.PubMed/NCBI
|
|
9
|
Spandau DF and Lee CH: Trans-activation of
viral enhancers by the hepatitis B virus X protein. J Virol.
62:427–434. 1988.PubMed/NCBI
|
|
10
|
Takada S, Shirakata Y, Kaneniwa N, et al:
Association of hepatitis B virus X protein with mitochondria causes
mitochondrial aggregation at the nuclear periphery, leading to cell
death. Oncogene. 18:6965–6973. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Zheng BY, Fang XF, Zou LY, Huang YH, Chen
ZX, Li D, Zhou LY, Chen H and Wang XZ: The co-localization of HBx
and COXIII upregulates COX-2 promoting HepG2 cell growth. Int J
Oncol. 45:1143–1150. 2014.PubMed/NCBI
|
|
12
|
Shirakata Y and Koike K: Hepatitis B virus
X protein induces cell death by causing loss of mitochondrial
membrane potential. J Biol Chem. 278:22071–22078. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Pelicano H, Lu W, Zhou Y, et al:
Mitochondrial dysfunction and reactive oxygen species imbalance
promote breast cancer cell motility through a CXCL14-mediated
mechanism. Cancer Res. 69:2375–2383. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Schäfer E, Seelert H, Reifschneider NH, et
al: Architecture of active mammalian respiratory chain
supercomplexes. J Biol Chem. 281:15370–15375. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Gilderson G, Salomonsson L, Aagaard A, et
al: Subunit III of cytochrome c oxidase of Rhodobacter sphaeroides
is required to maintain rapid proton uptake through the D pathway
at physiologic pH. Biochemistry. 42:7400–7409. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Bouchard MJ, Puro RJ, Wang L, et al:
Activation and inhibition of cellular calcium and tyrosine kinase
signaling pathways identify targets of the HBx protein involved in
hepatitis B virus replication. J Virol. 77:7713–7719. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
McClain SL, Clippinger AJ, Lizzano R, et
al: Hepatitis B virus replication is associated with an
HBx-dependent mitochondrion-regulated increase in cytosolic calcium
levels. J Virol. 81:12061–12065. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Misra KP, Mukherji A and Kumar V: The
conserved amino-terminal region (amino acids 1–20) of the hepatitis
B virus X protein shows a transrepression function. Virus Res.
105:157–165. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Kumar V, Jayasuryan N and Kumar R: A
truncated mutant (residues 58–140) of the hepatitis B virus X
protein retains trans-activation function. Proc Natl Acad Sci USA.
93:5647–652. 1996. View Article : Google Scholar
|