Introduction
Periodontitis is a highly prevalent inflammatory
oral disease. Delayed treatment for aggressive periodontitis could
result in the loss of soft connective tissues and bone that
surrounds the teeth and is a major cause of tooth loss in adults
(1). The ideal aim of periodontal
therapy is achieving simultaneous regeneration of the lost alveolar
bone, cementum and periodontal soft tissues (2). Current treatment options, such as
professional tooth cleaning/scaling/polishing, root planning,
periodontal debridement and guided tissue regeneration, used either
alone or in combination, only have a limited success in impeding
periodontitis, particularly at the advanced stage of the disease
(2–4). Together with the recent advances in
periodontal tissue engineering, novel treatment methods utilizing
signaling molecules and/or cell-based therapies have been developed
as new strategies to regenerate periodontal tissue damaged by
periodontitis (2,5,6).
Human periodontal ligament cells (hPDLCs) are
believed to be key players during periodontal tissue regeneration.
Numerous studies have shown that hPDLCs can be committed to several
cell lineages, including osteoblastic, fibroblastic, and
cementoblastic (7–10). In addition, the aforementioned
studies indicate that the organic osteogenic inductive factors,
dexamethasone (DEX) and β-glycerophosphate (β-GP), are efficient
for stimulating human osteoblasts (HOBs) and mesenchymal stem cells
(MSCs) in vitro, but are not believed to be able to achieve
similar stimulation in vivo. Additionally, the exact
phenotype of hPDLCs remains a topic of controversy, in particular,
concerning whether these cells are similar to a terminally
differentiated cell type with a fibroblastic nature or a progenitor
cell that potentially can be induced into a fibroblastic or
osteoblastic phenotype by certain growth factors (11).
Basic fibroblast growth factor (FGF-2) is reported
to be a strong mitogen and acts on various cell types, including
HOBs, MSCs and hPDLCs, in addition to being effective in
accelerating the proliferation of fibroblasts and osteoblasts, and
enhancing angiogenesis. These biological actions are directly
associated with periodontal tissue regeneration (12–14).
The optimum aims of periodontal treatment are to
regenerate the destroyed cementum, alveolar bone and periodontal
ligament, while also preventing epithelization during wound healing
(2,15,16). Numerous studies have demonstrated
that the topical application of FGF-2 promotes the healing of
destructive periodontal tissue without ankylosis, root resorption
or epithelial downgrowth in experimental alveolar bone defects in
the beagle dog and primate models (16–22). However, FGF-2 is considered a
multifunctional growth factor that has diversified actions and its
effect in the differentiation of hPDLCs remains controversial
(12). hPDLCs are a heterogeneous
population of cells and the exact function of FGF-2 in stabilizing
the fibroblastic phenotype and maintaining the human periodontal
ligament (hPDL) structural integrity remains largely unknown. Based
on the above factors and our previous study (23), we hypothesize that in the present
experimental system (with or without osteogenic inducers), FGF-2
may be able to maintain the fibroblastic phenotype of hPDLCs and
prevent them from differentiating into mature osteoblasts.
Thus, the aim of the present study was to reveal the
molecular and cellular mechanisms by which FGF-2 affects the
osteoblastic and fibroblastic phenotypes of hPDLCs in the presence
and absence of osteogenic inducers. The knowledge gained from this
study may facilitate the development of purposeful strategies to
influence hPDLC reparative capacities.
Materials and methods
Cell culture
hPDLCs were isolated from healthy premolar teeth
using a previously described method (23). Healthy subjects <20 years old
undergoing orthodontic treatment were recruited with the
understanding and consent for the isolation of hPDLCs. All the
experimental protocols used were approved by the Ethics Committee
of Sun Yat-sen University (Guangdong, China). Briefly, fresh PDL
tissues were collected rapidly following the tooth extraction.
Under sterile conditions, the PDL tissue was washed with sterile
phosphate-buffered saline (PBS) and scraped from the middle-third
of the root with a scalpel. The tissue was subsequently chopped
into smaller pieces and soaked in fresh Dulbecco's modified Eagle's
medium [DMEM/high glucose; Hyclone Biochemical Product (Beijing)
Co., Ltd., Beijing, China] containing 20% fetal bovine serum (FBS)
(Biological Industries Israel Beit-Haemek, Ltd., Kibbutz
Beit-Haemek, Israel) and 2% (v/v) penicillin/streptomycin
(Invitrogen Life Technologies, Carlsbad, CA, USA). An explant
culture method was used in the primary culture. The medium was
changed every 3 days. Cells typically emerged from the tissues
within 1–2 weeks of being explanted in a humidified atmosphere of
5% CO2 at 37°C. When the cells growing from the explants
reached ~80% confluency, they were subcultured at a 1:3 ratio
through trypsinization (trypsin/EDTA; Invitrogen Life
Technologies). Only the fifth passage of hPDLCs was used in
subsequent experiments.
FGF-2 treatment
Recombinant Human FGF-basic (FGF-2) (Peprotech,
Inc., Rocky Hill, NJ, USA) was added to the defined growth culture
medium and osteogenic inductive culture medium. There were four
treatment groups: i) hPDLCs + growth medium (GM) (DMEM containing
10% FBS and 2% penicillin/streptomycin); ii) hPDLCs + GM + 20 ng/ml
FGF-2 (GM + FGF-2); iii) hPDLCs + osteogenic medium (OM) [DMEM with
10% FBS, 2% (v/v) penicillin/streptomycin, 10−8 M DEX,
50 µg/ml ascorbic acid and 8 mM β-GP]; and iv) hPDLCs + OM +
20 ng/ml FGF-2 (OM + FGF-2).
Cell proliferation assay
hPDLC viability was monitored using a Cell Counting
kit-8 (CCK-8) (Beyotime, Shanghai, China). Briefly, hPDLCs at
passage 5 were seeded in 96-well plates at an initial density of
3×103 cells/well in medium, with five replicates for
each group. The medium was replaced every 3 days. After 1, 2, 3, 5
and 7 days of subculture in medium treated differently, 20
µl of CCK-8 solution was added to every well and incubated
with the cells for 4 h in a humidified atmosphere of 5%
CO2 at 37°C. The absorbance of each sample was
determined at the wavelength of 450 nm on a 96-well plate
reader.
Alkaline phosphatase (ALP) activity
assay
hPDLCs were seeded in 24-well plates in triplicate
at a density of 2×104 cells/well and cultured in defined
media subsequent to reaching ~80% confluency. Following 3, 7, 14
and 21 days in culture, an ALP activity assay kit (Nanjing
Jiancheng Biotechnology Co., Ltd., Jiangsu, China) was used to
detect ALP activity of hPDLCs, following the manufacturer's
instructions. In brief, the cell layers in plates were washed three
times with 10 mM sterile PBS, and subsequently a small amount (1
ml) of cold 10 mM Tris-HCl buffer (pH 7.4) containing 0.1% Triton
X-100 was added to the culture well, and the mixture was lysed
through two freeze (−20°C)/thaw cycles. The supernatant (50
µl/well) was placed into 24-well plates for the ALP activity
assay using 50 µl of an ALP substrate solution (16 mM
p-nitrophenyl phosphate and 2 mM MgCl2). After a
30-min incubation at 37°C, the reaction was stopped by the addition
of 50 µl of 0.2 M NaOH, and the absorbance of the reaction
product (p-nitrophenol) was read at 520 nm.
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR) analysis
After 7 and 14 days of culture, the expression
levels of selected osteoblastic and fibroblastic genes were
determined in triplicate by RT-qPCR. Total RNA was extracted from
hPDLCs using TRIzol Reagent (Invitrogen Life Technologies) and was
quantified in a spectrophotometer by absorbance readings at 260 nm.
The extracted total RNA (2 µg) was treated with 1 unit DNase
I for cDNA synthesis in each RT-qPCR using the RevertAid™ First
Strand cDNA Synthesis kit (K1622; MBI Fermentas, Inc., Burlington,
ON, Canada). PCR was carried out using a combination of 2.5
µl of diluted cDNA and 12.5 µl of SYBR-Green
Real-time PCR Master mix (QPK-201; Toyobo Co., Ltd., Osaka, Japan)
in a Chromo4 System (Bio-Rad, Hercules, CA, USA). The sequential
reaction conditions were as follows: 95°C for 1 min denaturation
followed by 40 cycles of denaturation at 95°C for 15 sec, annealing
at 60°C for 15 sec and extension at 72°C for 30 sec. The
specificity of the PCR products was confirmed by melting curve
analysis (60–93°C, read every 0.4°C, hold 1 sec). GAPDH was used as
the reference gene for data normalization. The number of cDNA
copies was calculated with the apparent cycle threshold (CT). The
ΔCT value expresses the difference between the CT of the target
gene and the CT of GAPDH from the same sample: ΔCT = the target
gene CT - GAPDH CT, which can be expressed as a percentage of
GAPDH. The relative expression level of the target gene (fold of
the reference gene) was obtained by transforming the logarithmic
values to absolute values using 2−ΔCT. The primer
sequences are shown in Table
I.
 | Table IPrimer sequences used for
RT-qPCR. |
Table I
Primer sequences used for
RT-qPCR.
| Genes | Primers | Size, bp |
|---|
| GAPDH | F:
CATGTTCCAATATGATTCCACC
R: GATGGGATTTCCATTGATGAC | 88 |
| Runx2 | F:
CCAACCCACGAATGCACTATC
R: TAGTGAGTGGTGGCGGACATAC | 91 |
| Col1a1 | F:
GAACGCGTGTCATCCCTTGT
R: GAACGAGGTAGTCTTTCAGCAACA | 91 |
| OCN | F:
CCTGAAAGCCGATGTGGT
R: GGCAGCGAGGTAGTGAAGA | 148 |
| EGFR | F:
GGAGAACTGCCAGAAACTGACC
R: GCCTGCAGCACACTGGTTG | 106 |
Western blotting
After 7 and 14 days of culture, Runx2 and
osteocalcin (OCN) protein expression was measured using western
blotting. In brief, hPDLCs were washed three times with pre-cooling
PBS. The constructs were subsequently homogenized in sodium dodecyl
sulfate polyacrylamide gel electrophoresis (SDS-PAGE) sample
buffer, and cellular protein was extracted in lysis buffer [50 mM
Tris-HCl, 0.5% Triton X-100, 2 mM EDTA and 150 mM NaCl (pH 7.5)]
containing phenylmethylsulfonyl fluoride. The protein samples (30
µg) were electrophoresed through 10% SDS-PAGE and
transferred to a polyvinylidene fluoride (PVDF) membrane
(Millipore, Bedford, MA, USA) in a wet blotting apparatus for 1.5 h
at 300 mA. The PVDF membrane was subsequently blocked with 5%
bovine serum albumin (Wuhan Boster Biological Technology, Ltd.,
Hubei, China) for 1 h at room temperature, and the blot was
incubated with 1:1,000 dilution of anti-human Runx2 antibody (Cat.
no. AF2006; R&D Systems, Minneapolis, MN, USA) and 1:500 of
anti-human OCN (Cat. no. ab10911; Millipore, Billerica, MA, USA)
for 2 h, washed with Tris-buffered saline Tween 20 (TBST) four
times and incubated with a 1:2,500 dilution of horseradish
peroxidase (HRP) AffiniPure goat anti-rabbit secondary antibody
(Santa Cruz Biotechnology, Inc., Dallas, TX, USA) for 1 h, and
further washed four times with TBST. HRP detection was accomplished
via chemical luminescence exposed to X-ray film with an ECL western
blotting detection system.
Calcium content quantitative assay
The calcium content of the cells was measured by a
colorimetric quantitative method using a Calcium Assay kit
(Shanghai Genmed Gene Pharmaceutical Technology Co., Ltd.,
Shanghai, China) following the manufacturer's instructions. Cells
were seeded at a cell density of 5×104 cells/well in
12-well tissue culture plates. After 14 and 21 days of culture,
deposition of calcium in the cell layer was performed in
triplicate. The absorbances of dyes were read at a wavelength of
595 nm using a UV/Visible light Spectrophotometer (Varian Cary 50;
Varian Australia Pty, Ltd., Mulgrave, VIC, Australia).
Alizarin Red S staining
After 7, 14 and 21 days of culture, the mineralized
matrix nodules were detected using an Alizarin Red S Staining kit
(Shanghai Genmed Gene Pharmaceutical Technology Co., Ltd.)
following the manufacturer's instructions. Prior to staining, the
cell cultures were washed five times with 10 mM sterile PBS and
fixed using 10% (v/v) neutral buffered formalin for 30 min.
Subsequently, the samples were stained for 5 min and the excess
stain was rinsed with PBS. An Inverted Phase Contrast Microscope
(Olympus IX41; Olympus Corp., Tokyo, Japan) was used to observe the
stained cells.
Statistical analysis
All the values are reported as the mean ± standard
deviation. Statistical analysis of the data was performed using the
SPSS 17.0 software package (SPSS, Inc., Chicago, IL, USA). One-way
analysis of variance and the least significant difference test were
used for parameter estimation and hypothesis testing. P<0.05 was
considered to indicate a statistically significant difference.
Results
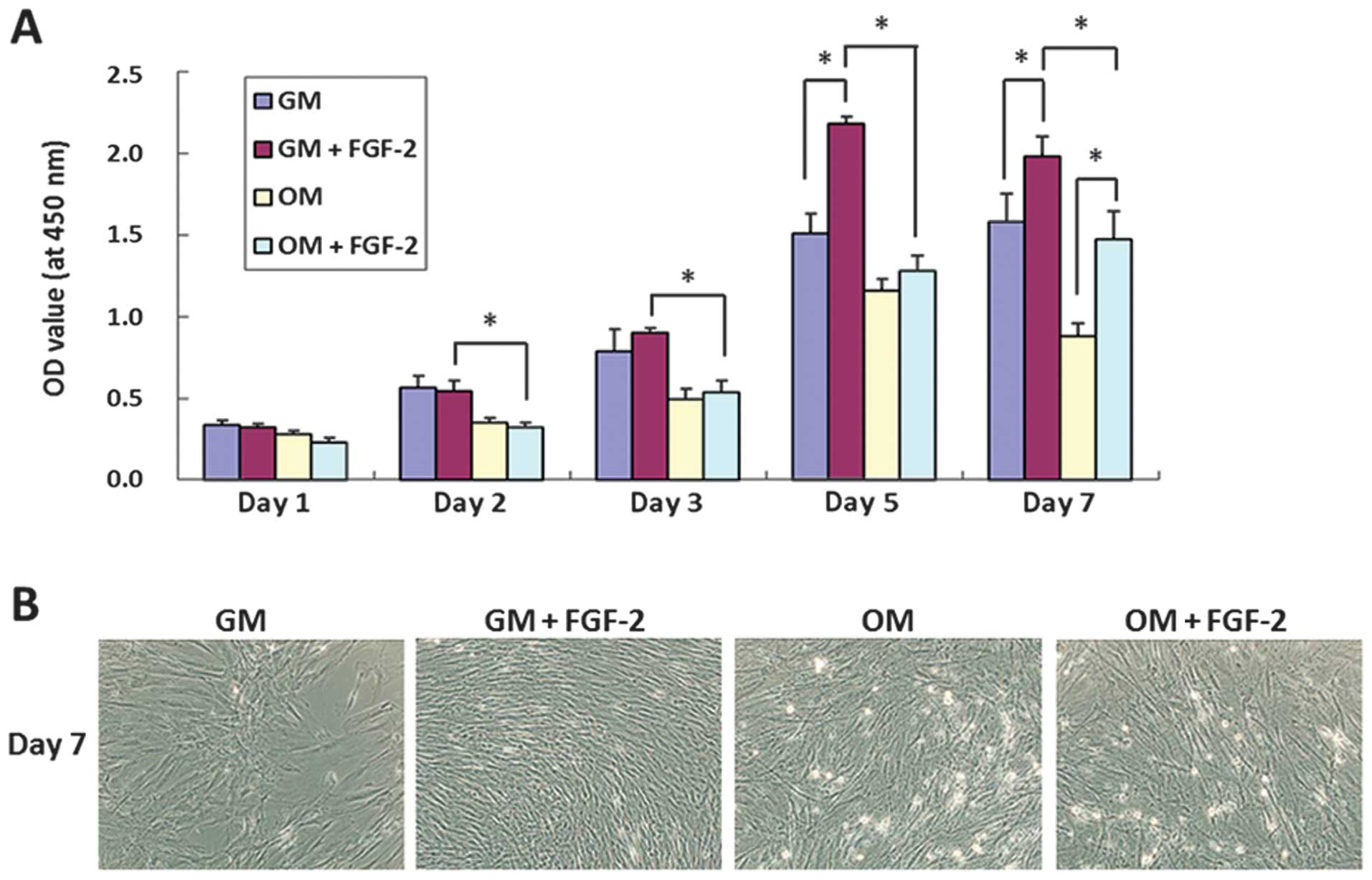
FGF-2 promotes the proliferation of
hPDLCs
At days 1 and 2, FGF-2 had no significant effect on
the cell proliferation rate (P>0.05). From day 3 onwards, hPDLCs
cultured in the GM with added FGF-2 exhibited the maximal levels of
proliferation, and those in the OM exhibited minimum levels
(P<0.05) (Fig. 1). This
suggested that supplementing growth media with FGF-2 enhanced
proliferation of hPDLCs in the presence and absence of osteogenic
inducers.
FGF-2 does not affect ALP activity
The specific ALP activity progressively increased
with the duration of culture time in all the groups. At days 3 and
7 of culture, FGF-2 inhibited the ALP activity. However, FGF-2
promoted the ALP activity at days 14 and 21. There were, however,
no significant differences between FGF-2-treated and -untreated
groups at any time-points (P>0.05) (Fig. 2).
FGF-2 affects the mRNA and protein
expression of Runx2, but not the OCN protein level
In the cultures without osteogenic inducers,
Runx2 mRNA expression was significantly upregulated under
the stimulation of FGF-2 at days 7 and 14 (P<0.05). However, in
the OM, there was no evident effect on Runx2 expression
(P>0.05) (Fig. 3A).
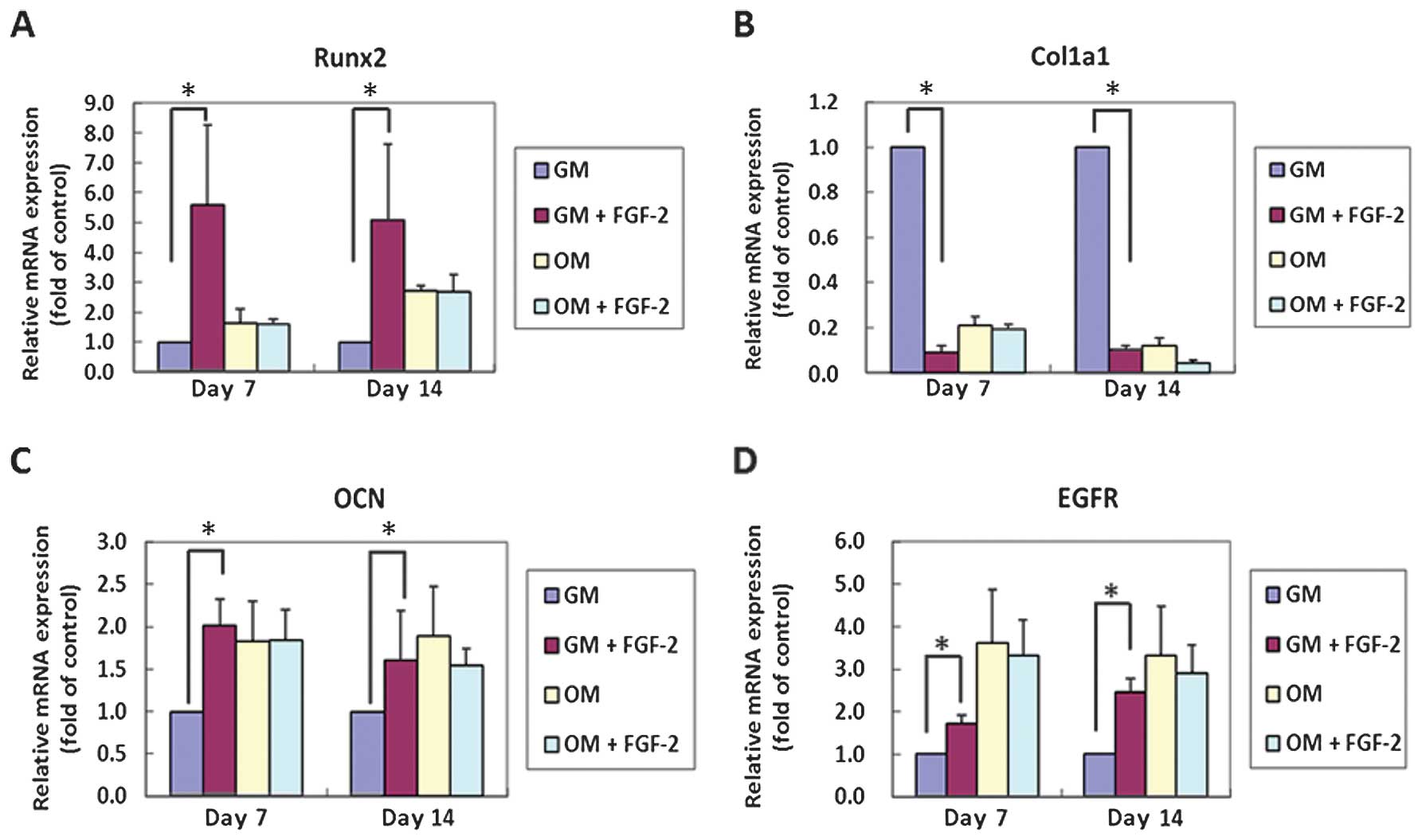 | Figure 3Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR) analysis of osteogenic and
fibroblastic differentiation markers in human periodontal ligament
cells (hPDLCs). The results are standardized to the reference gene
GAPDH, and expressed as relative mRNA levels. (A)
Runx2 mRNA. In the cultures lacking osteogenic inducers
[growth medium (GM)], the Runx2 mRNA expression was
upregulated significantly at days 7 and 14. However, in the
osteogenic medium (OM), there were no clear trends in the levels of
Runx2. (B) Collagen type I, α1 (Col1a1) mRNA. In the
GM, that is, in the absence of osteogenic inducers, basic
fibroblast growth factor (FGF-2) suppressed Col1a1 mRNA
levels; however, no no significant effect was shown in the OM (in
the presence of osteogenic inducers). (C) Osteocalcin (OCN) mRNA.
In the GM, lacking osteogenic inducers, FGF-2 upregulated the gene
expression of OCN significantly at day 7. However, in the OM
supplemented with osteogenic inducers, there were no clear trends.
(D) Epidermal growth factor receptor (EGFR) mRNA. FGF-2
promoted the expression of the EGFR gene in the absence of
osteogenic inducers; however, there was inhibited EGFR gene
expression in the presence of osteogenic inducers. |
In the western blot analysis, Runx2 expression was
increased at days 7 and 14 in the medium without osteogenic
inducers under the stimulation of FGF-2. In the OM samples, there
was no increase in Runx2 protein levels at day 7, whereas at day 14
there was a significantly increased protein expression stimulated
by FGF-2 (Fig. 4A). However, the
protein expression of OCN showed no evident changes at day 7 and 14
(Fig. 4B).
FGF-2 affects collagen type I, α1
(Col1a1), OCN and epidermal growth factor receptor (EGFR) gene
expression of hPDLCs in the absence of osteogenic inducers
There was a significant decrease in Col1a1
mRNA levels in the GM + FGF-2 samples (P<0.05), whereas in the
OM samples the gene expression was downregulated at day 7
(P>0.05) under the stimulation of FGF-2 (Fig. 3B). OCN and EGFR expression
was upregulated in GM + FGF-2 (P<0.05); but downregulated to
various degrees in the OM + FGF-2 samples (P>0.05) (Fig. 3C and D). Compared with the GM
condition, OM culture stimulated OCN and EGFR expression in
the absence of FGF-2.
FGF-2 suppresses the mineralization of
hPDLCs in the presence of osteogenic inducers
After 14 and 21 days of culture, the calcium content
in the cultures was measured. The calcium content in the
FGF-2-treated groups was higher than the non-FGF-2 groups
(P>0.05). However, in the OM + FGF-2 group there was a
significant reduction of the calcium content (P<0.05) (Fig. 5A).
Alizarin Red S staining indicated that mineralized
nodes were observed only in the OM, and mineralized matrix
deposition gradually increased with time in culture. The addition
of FGF-2 resulted in reduced calcium deposition as evaluated at
days 7, 14 and 21 (Fig. 5B).
Discussion
Cell proliferation and differentiation are two
crucial aspects of cell-based bone regeneration.
Osteogenic-differentiated cells generally exhibit low proliferation
rates (24) and may explain why
hPDLCs cultured in medium with added osteogenic inducers exhibited
the lowest proliferation rate. hPDLCs in groups without osteogenic
inducers had higher EGFR mRNA levels, which resulted in
increased cell proliferation most possibly via activation of its
downstream targets, ERK1/2 and Akt (25,26). Thus, the cell proliferation rates
in this group are higher compared to the OM group. CCK-8 analysis
indica ted that FGF-2 was able to increase the proliferation rate
of hPDLCs in the presence and absence of osteogenic inducers. These
results are similar to those of a study that identified that
co-stimulation with fetal calf serum synergistically enhanced
FGF-2-induced periodontal ligament cell proliferation, but
inhibited FGF-2-induced proliferation of gingival epithelial cells
(27). We speculate that this
enhancement could provide an alternate route to limit the
downgrowth of junctional epithelium and increase the relatively low
number of cells adhering to the scaffold surface in periodontal
tissue engineering.
ALP activity was a well-defined marker for
osteogenic differentiation. Previous studies indicate that the
addition of FGF-2 significantly decreased ALP activity and
calcified nodule formation in hPDLCs in a dose-dependent manner
(12). The concentration of FGF-2
chosen in the present study at 20 ng/ml was based on the previous
studies (28,29). In these studies, it was reported
that FGF-2 promoted the proliferation of hPDLCs in a dose-dependent
manner and that the action plateaued c at >100 ng/ml, whereas
ALP activities of hPDLCs were completely prohibited when the cells
were treated with >10 ng/ml of FGF-2. The present data indicated
that FGF-2 inhibited calcified nodule formation; however, it did
not affect the ALP activity of hPDLCs. This difference may be
associated with the concentration of FGF-2 and the experimental
conditions.
Endogenous Runx2 is expressed in
pre-osteoblasts, immature osteoblasts, early mature osteoblasts and
pre-odontoblasts, and has been served as a master regulator in
osteoblastic differentiation and bone formation (30). The present results showed that
there was no observable effect of FGF-2 on Runx2 gene
expression regardless of FGF-2-induced Runx2 protein expression on
day 14. We hypothesize that this phenomenon may be explained by the
study of Shui et al (31),
which reported that osteoblastic differentiation of human
mesenchymal stem cells (hMSCs) is associated primarily with
increases of Runx2 activity through a post-translational mechanism
without a change in mRNA.
Col1a1 is the most abundant organic component of
dentin, hPDL and bone, and is an essential factor in the formation
of calcified nodules (32). The
overexpression of Runx2 inhibits osteoblast terminal
differentiation and alters their expression of extracellular matrix
protein genes, which results in down-regulation of Col1a1
mRNA (30,33). In the present study, FGF-2
inhibited Col1a1 mRNA levels to various degrees in the
absence of osteogenic inducers. These results may be associated
with the increase of Runx2 mRNA and protein expression
triggered by FGF-2 and are similar to previously reported studies,
which identified that the addition of FGF-2 to culture media
suppressed the expression of Col1a1 mRNA at all time-points
<14 days (29,34,35). Therefore, in the present culture
system, FGF-2 induced Runx2 protein expression but suppressed the
mineralization of hPDLCs by affecting Col1a1 mRNA
expression. Additionally, DEX has been demonstrated to inhibit the
expression of Col1a1 mRNA in immortalized hMSCs during
osteogenic differentiation (36).
This function-suppressing differentiation of hPDLCs into
mineralized tissue may aid in the stabilization of the fibroblastic
phenotype and maintenance of hPDL structural integrity.
EGFR is highly upregulated in periodontal disease
and may have a pivotal role in regulating cell migration,
proliferation and epithelial wound healing (37,38). A previous study has reported that
the EGF/EGFR system regulates the phenotype of different cell
populations (39), and
pre-osteoblasts and prechondrocytes have high levels of EGFR in
vivo. However, the amount diminishes significantly as
differentiation progresses. Mature osteoblasts and chondrocytes do
not express EGFR (40,41), and in addition, EGFR on hPDLCs
functions as a negative regulator of their differentiation into
mineralized tissue-forming cells (12). In the present study, the gene
expression of EGFR was significantly upregulated by FGF-2
and osteogenic inducers compared with hPDLCs cultured in the GM,
but downregulated by FGF-2 in the presence of osteogenic inducers.
Thus, we hypothesize that the upregulation of the EGFR gene
may have an important role in maintaining the hPDLC phenotype,
balancing the population of fibroblasts in the hPDL by inhibiting
their differentiation into mineralized tissue-forming cells.
In conclusion, the results of the present study
demonstrate that FGF-2 facilitates hPDLC proliferation in the
absence and presence of osteogenic inducers, while modulating the
balance between their osteoblastic and fibroblastic phenotypes by
affecting the gene expression of Runx2, OCN, Col1a1
and EGFR in the absence of ostoegenic inducers, in addition
to inhibiting the mineralization of hPDLCs in the presence of
osteogenic inducers. These results suggested that FGF-2 induces the
growth of immature hPDLCs, which is a competitive inhibitor of
epithelial downgrowth, and suppresses their differentiation into
mineralized tissue by affecting Runx2 expression. This may lead to
the acceleration of periodontal regeneration.
Acknowledgments
The present study was supported by the Guangdong
Natural Science Foundation (grant no. S2012040007041), and
Guangdong Science & Technology Planning Program Fund (grant no.
2013B022000039).
References
|
1
|
Pihlstrom BL, Michalowicz BS and Johnson
NW: Periodontal diseases. Lancet. 366:1809–1820. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Ishikawa I, Iwata T, Washio K, Okano T,
Nagasawa T, Iwasaki K and Ando T: Cell sheet engineering and other
novel cell-based approaches to periodontal regeneration.
Periodontol 2000. 51:220–238. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Lin NH, Gronthos S and Bartold PM: Stem
cells and future periodontal regeneration. Periodontol 2000.
51:239–251. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Chen FM, Sun HH, Lu H and Yu Q: Stem
cell-delivery therapeutics for periodontal tissue regeneration.
Biomaterials. 33:6320–6344. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Taba M Jr, Jin Q, Sugai JV and Giannobile
WV: Current concepts in periodontal bioengineering. Orthod
Craniofac Res. 8:292–302. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Hynes K, Menicanin D, Gronthos S and
Bartold PM: Clinical utility of stem cells for periodontal
regeneration. Periodontol 2000. 59:203–227. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Seo BM, Miura M, Gronthos S, Bartold PM,
Batouli S, Brahim J, Young M, Robey PG, Wang CY and Shi S:
Investigation of multi-potent postnatal stem cells from human
periodontal ligament. Lancet. 364:149–155. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Lekic P, Rojas J, Birek C, Tenenbaum H and
McCulloch CA: Phenotypic comparison of periodontal ligament cells
in vivo and in vitro. J Periodontal Res. 36:71–79. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Ballini A, De Frenza G, Cantore S, Papa F,
Grano M, Mastrangelo F, Tetè S and Grassi FR: In vitro stem cell
cultures from human dental pulp and periodontal ligament: New
prospects in dentistry. Int J Immunopathol Pharmacol. 20:9–16.
2007.PubMed/NCBI
|
|
10
|
Nagatomo K, Komaki M, Sekiya I, Sakaguchi
Y, Noguchi K, Oda S, Muneta T and Ishikawa I: Stem cell properties
of human periodontal ligament cells. J Periodontal Res. 41:303–310.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Lossdörfer S, Fiekens D, Salik MI, Götz W
and Jäger A: Subculture affects the phenotypic expression of human
periodontal ligament cells and their response to fibroblast growth
factor-2 and bone morphogenetic protein-7 in vitro. J Periodontal
Res. 43:563–569. 2008.PubMed/NCBI
|
|
12
|
Murakami S: Periodontal tissue
regeneration by signaling molecule(s): What role does basic
fibroblast growth factor (FGF-2) have in periodontal therapy?
Periodontol 2000. 56:188–208. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Ramseier CA, Rasperini G, Batia S and
Giannobile WV: Advanced reconstructive technologies for periodontal
tissue repair. Periodontol 2000. 59:185–202. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Stavropoulos A and Wikesjö UM: Growth and
differentiation factors for periodontal regeneration: A review on
factors with clinical testing. J Periodontal Res. 47:545–553. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Ivanovski S, Gronthos S, Shi S and Bartold
PM: Stem cells in the periodontal ligament. Oral Dis. 12:358–363.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Bartold PM, Shi S and Gronthos S: Stem
cells and periodontal regeneration. Periodontol 2000. 40:164–172.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Murakami S, Takayama S, Ikezawa K,
Shimabukuro Y, Kitamura M, Nozaki T, Terashima A, Asano T and Okada
H: Regeneration of periodontal tissues by basic fibroblast growth
factor. J Periodontal Res. 34:425–430. 1999. View Article : Google Scholar
|
|
18
|
Seshima F, Ota M, Kinumatsu T, Shibukawa Y
and Yamada S: Effect of recombinant basic fibroblast growth factor
on reim-planted teeth in beagle dogs. Oral Surg Oral Med Oral
Pathol Oral Radiol Endod. 109:142–148. 2010. View Article : Google Scholar
|
|
19
|
Takayama S, Murakami S, Shimabukuro Y,
Kitamura M and Okada H: Periodontal regeneration by FGF-2 (bFGF) in
primate models. J Dent Res. 80:2075–2079. 2001. View Article : Google Scholar
|
|
20
|
Chen FM and Jin Y: Periodontal tissue
engineering and regeneration: Current approaches and expanding
opportunities. Tissue Eng Part B Rev. 16:219–255. 2010. View Article : Google Scholar
|
|
21
|
Shiratani S, Ota M, Fujita T, Seshima F,
Yamada S and Saito A: Effect of basic fibroblast growth factor on
root resorption after delayed autotransplantation of tooth in dogs.
Oral Surg Oral Med Oral Pathol Oral Radiol. 114:e14–e21. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Kitamura M, Akamatsu M, Machigashira M, et
al: FGF-2 stimulates periodontal regeneration: Results of a
multi-center randomized clinical trial. J Dent Res. 90:35–40. 2011.
View Article : Google Scholar
|
|
23
|
An S, Ling J, Gao Y and Xiao Y: Effects of
varied ionic calcium and phosphate on the proliferation, osteogenic
differentiation and mineralization of human periodontal ligament
cells in vitro. J Periodontal Res. 47:374–382. 2012. View Article : Google Scholar
|
|
24
|
Lu H, Kawazoe N, Tateishi T, Chen G, Jin X
and Chang J: In vitro proliferation and osteogenic differentiation
of human bone marrow-derived mesenchymal stem cells cultured with
hardystonite (Ca2ZnSi2O7) and
β-TCP ceramics. J Biomater Appl. 25:39–56. 2010. View Article : Google Scholar
|
|
25
|
Kasaj A, Willershausen B, Reichert C,
Gortan-Kasaj A, Zafiropoulos GG and Schmidt M: Human periodontal
fibroblast response to a nanostructured hydroxyapatite bone
replacement graft in vitro. Arch Oral Biol. 53:683–689. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Kasaj A, Willershausen B, Reichert C,
Röhrig B, Smeets R and Schmidt M: Ability of nanocrystalline
hydroxyapatite paste to promote human periodontal ligament cell
proliferation. J Oral Sci. 50:279–285. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Takayama S, Yoshida J, Hirano H, Okada H
and Murakami S: Effects of basic fibroblast growth factor on human
gingival epithelial cells. J Periodontol. 73:1467–1473. 2002.
View Article : Google Scholar
|
|
28
|
Hidaka T, Nagasawa T, Shirai K, Kado T and
Furuichi Y: FGF-2 induces proliferation of human periodontal
ligament cells and maintains differentiation potentials of
STRO-1(+)/CD146(+) periodontal ligament cells. Arch Oral Biol.
57:830–840. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Takayama S, Murakami S, Miki Y, Ikezawa K,
Tasaka S, Terashima A, Asano T and Okada H: Effects of basic
fibroblast growth factor on human periodontal ligament cells. J
Periodontal Res. 32:667–675. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Komori T: Regulation of bone development
and extracellular matrix protein genes by RUNX2. Cell Tissue Res.
339:189–195. 2010. View Article : Google Scholar
|
|
31
|
Shui C, Spelsberg TC, Riggs BL and Khosla
S: Changes in Runx2/Cbfa1 expression and activity during
osteoblastic differentiation of human bone marrow stromal cells. J
Bone Miner Res. 18:213–221. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Ohi T, Uehara Y, Takatsu M, Watanabe M and
Ono T: Hypermethylation of CpGs in the promoter of the COL1A1 gene
in the aged periodontal ligament. J Dent Res. 85:245–250. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Miyazaki T, Kanatani N, Rokutanda S,
Yoshida C, Toyosawa S, Nakamura R, Takada S and Komori T:
Inhibition of the terminal differentiation of odontoblasts and
their transdifferentiation into osteoblasts in Runx2 transgenic
mice. Arch Histol Cytol. 71:131–146. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Palmon A, Roos H, Edel J, Zax B, Savion N,
Grosskop A and Pitaru S: Inverse dose- and time-dependent effect of
basic fibroblast growth factor on the gene expression of collagen
type I and matrix metalloproteinase-1 by periodontal ligament cells
in culture. J Periodontol. 71:974–980. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Lee JH, Um S, Jang JH and Seo BM: Effects
of VEGF and FGF-2 on proliferation and differentiation of human
periodontal ligament stem cells. Cell Tissue Res. 348:475–484.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Ogston N, Harrison AJ, Cheung HF, Ashton
BA and Hampson G: Dexamethasone and retinoic acid differentially
regulate growth and differentiation in an immortalised human clonal
bone marrow stromal cell line with osteoblastic characteristics.
Steroids. 67:895–906. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Ekuni D, Firth JD and Putnins EE:
Regulation of epithelial cell growth factor receptor protein and
gene expression using a rat periodontitis model. J Periodontal Res.
41:340–349. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Kim JM, Bak EJ, Chang JY, Kim ST, Park WS,
Yoo YJ and Cha JH: Effects of HB-EGF and epiregulin on wound
healing of gingival cells in vitro. Oral Dis. 17:785–793. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Wei N, Yu H, Yang S, Yang X, Yuan Q, Man Y
and Gong P: Effect of FDC-SP on the phenotype expression of
cultured periodontal ligament cells. Arch Med Sci. 7:235–241. 2011.
View Article : Google Scholar
|
|
40
|
Cho MI and Garant PR: Expression and role
of epidermal growth factor receptors during differentiation of
cementoblasts, osteoblasts, and periodontal ligament fibroblasts in
the rat. Anat Rec. 245:342–360. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Dereka XE, Markopoulou CE and Vrotsos IA:
Role of growth factors on periodontal repair. Growth Factors.
24:260–267. 2006. View Article : Google Scholar
|