Introduction
Inflammatory bowel disease (IBD) is a group of
chronic inflammatory conditions that affect the gastrointestinal
tract and includes two main clinical forms: Crohn's disease (CD)
and ulcerative colitis (UC). Although the precise etiology of IBD
is not yet clearly understood, it has been suggested that an
abnormal immune response against intestinal microbes plays a role
in the pathogenesis of colitis (1).
5-Hydroxytryptamine (serotonin; 5-HT) modulates
intestinal inflammation in the pathogenesis of colitis. Changes in
the levels of 5-HT have been observed in experimental models of
colitis and in patients with IBD (2–4).
The knockdown 5-HT expression in tryptophan hydroxylase 1-deficient
(TPH1−/−) mice, has been shown to lead to a delayed
onset and to reduce the severity of dextran sodium sulfate
(DSS)-induced colitis (5).
Mucosal changes in IBD are characterized by ulcerative lesions with
the prominent infiltration of immune cells and alterations in
serotonin-producing enterochromaffin cells (6). 5-HT promotes the production of
pro-inflammatory cytokines from macrophages (7), and nuclear factor-κB (NF-κB)
signaling plays a critical role in the 5-HT-mediated activation of
immune cells (8). However, the
precise functions of 5-HT in intestinal inflammation are not yet
fully understood. This is partly due to the fact that a wide
variety of 5-HT receptors are expressed in the gut;
5-HT1A, 5-HT2A, 5-HT2B,
5-HT3, 5-HT4 and 5-HT7 receptors
are expressed in the intestines and some are known to have varying,
and even contrasting roles.
Ketanserin is a selective 5-HT2A receptor
(5-HT2AR) antag-antagonist with minor activity on the α1
adrenergic receptor (9). Previous
studies have suggested that ketanserin exerts anti-inflammatory
effects independent of the baroreflex (10,11). Ketanserin has been shown to
inhibit the expression of inducible nitric oxide (NO) synthase
(iNOS), NO, tumor necrosis factor-α (TNF-α), and interleukin (IL)-6
in lipopolysaccharide (LPS)-stimulated RAW264.7 cells (12). Moreover, the administration of
ketanserin has been shown to decrease the expression of iNOS
following endotoxic shock in mice (13). iNOS is primarily produced by
macrophages. Macrophages participate in the immune response to the
microenvironment in a polarized manner (14). Macrophages are often divided into
2 functional groups: 'inflammatory' M1 macrophages (high IL-12 and
iNOS expression) and 'wound-healing' M2 macrophages (low IL-12
expression, and high IL-10 expression) (15). M1 macrophages mainly produce
inflammatory cytokines (TNF-α and IL-12) and iNOS. The production
of these cytokines promotes naïve CD4+ T cells to
differentiate into Th1 and Th17 cells (16). M2 macrophages produce the
anti-inflammatory cytokine, IL-10, and are efficient at recruiting
regulatory T cells that promote tolerance to local immune responses
(16). In this study, we examined
the hypothesis that ketanserin may attenuate colitis by modulating
the function of macrophages, thus alleviating colitis.
Materials and methods
Human tissue samples
Intestinal specimens were obtained from 15 patients
with IBD. Normal tissue specimens were obtained from patients
receiving colonoscopy for colorectal cancer screening. The tissues
were frozen in liquid nitrogen and stored at −80°C for reverse
transcription-quantitative PCR (RT-qPCR) analyses, or were embedded
in optimum cutting temperature (OCT) compound for
immunofluorescence staining. All studies involving samples obtained
from human participants were approved by the Ethics Committee of
the Humanitas Clinical and Research Center of Shanghai East
Hospital, Shanghai, China. Written informed consent was obtained
from each patient or their family before initiating the study
protocol.
Mice
A total of 30 C57BL/6 male mice (8 to 10 weeks old,
weighing 20 g) were purchased from Slaccas Laboratory Animal Co.,
Ltd. (Shanghai, China) and all experiments were performed under
specific pathogen-free conditions at the Animal Resources
facilities of Shanghai East Hospital. All animal experiments were
performed following approval by the Animal Care and Use Committee
of Shanghai East Hospital.
Induction of colitis by DSS
The mice were randomly divided into 3 groups as
follows: the control group, DSS treatment group and the DSS plus
ketanserin treatment group (n=10 in each group). The mice were
administered 4% (wt/vol) DSS (MP Biomedicals, Solon, OH, USA)
dissolved in their drinking water for 7 days to induce acute
experimental colitis. Ketanserin (10 mg/kg; Janssen
Pharmaceuticals, Beijing, China) or phosphate-buffered saline (PBS;
vehicle control; administered to the mice in the DSS treatment
group) were administered intra-peritoneally once daily for 3 days
when the administration of DSS began. The weight of the mice was
recorded daily using an electronic weighing system (Practum
612-1CN; Sartorius AG, Goettingen, Germany). On day 7, the mice
were sacrificed by an intraperitoneal injection of 4% sodium
pentobarbital and the colons were obtained for the measurement of
colon length. For histological analysis, the colons were excised
and fixed in 4% paraformaldehyde. Subsequentoy, 1 cm of the distal
colon of each mouse was paraffin-embedded. The paraffin-embedded
sections were cut (4-μM-thick) using a microtome and stained
with hematoxylin and eosin. For gene and protein expression
analyses, the colons were immediately frozen in liquid
N2 and stored at −80°C.
RT-qPCR
Total RNA was extracted from the colon tissues or
lamina propria macrophages (LP-macrophages) and bone marrow
derived-macrophages (BMDMs) using TRIzol reagent (Invitrogen,
Carlsbad, CA, USA) according to the manufacturer's instructions.
Complementary DNA (cDNA) was synthesized using the PrimeScript™ RT
reagent kit (Takara, Shiga, Japan). Messenger RNA (mRNA)
transcripts were analyzed by quantitative PCR using
SYBR® Premix Ex Taq™ (Takara) with an Applied Biosystems
StepOne/StepOnePlus Real-Time PCR System. Gene expression was
normalized to the expression of glyceraldehyde 3-phosphate
dehydrogenase (GAPDH) and the relative expression levels were
quantified using the 2−ΔΔCT method. The primer pairs
used are listed in Table I.
 | Table ISequences of primers used for
RT-qPCR. |
Table I
Sequences of primers used for
RT-qPCR.
| Gene product | Forward primer | Reverse primer |
|---|
| mTNF-α |
5′-gtcgtagcaaaccaccaagtg-3′ |
5′-cagatttgtgttggtccttc-3′ |
| mIL-1β |
5′-atctcgcagcagcacatcaac-3′ |
5′-cacgggaaagacacaggtagc-3′ |
| mIL-6 |
5′-acaagtcggaggcttaattacacat-3′ |
5′-ttgccattgcacaactcttttc-3′ |
| mIL-10 |
5′-gctcttactgactggcatgag-3′ |
5′-cgcagctctaggagcatgtg-3′ |
| mIL-12 p40 |
5′-agacatggagtcataggctctg-3′ |
5′-ccattttccttcttgtggagca-3′ |
| miNOS |
5′-gccaccaacaatggcaaca-3′ |
5′-cgtaccggatgagctgtgaatt-3′ |
| mKC |
5′-cttgaaggtgttgccctcag-3′ |
5′-tggggacaccttttagcatc-3″ |
| mCXCL12 |
5′-aaaccagtcagcctgagctac-3′ |
5′-aagggcacagtttggagtgtt-3′ |
| mCD206 |
5′-ttcggtggactgtggacgagca-3′ |
5′-ataagccacctgccactccggt-3′ |
| mCD32 |
5′-aatcctgccgttcctactgatc-3′ |
5′-gtgtcaccgtgtcttccttgag-3′ |
|
m5-HT2A− |
5′-cgactcgaggctctacccta-3′ |
5′-ttcgcaggagaggttggttc-3′ |
|
h5-HT2AR |
5′-tgaagtcagcaaaacagaaacca-3′ |
5′-gactcgctgcatctctcaca-3′ |
Western blot analysis
Colon proteins were extracted using RIPA buffer
supplemented with protease inhibitors (Shanghai Shenggong Co.,
Ltd., Shanghai, China). A total of 40 μg protein was
separated by 10% sodium dodecyl sulfate-polyacrylamide gel
electrophoresis (SDS-PAGE), and the gels were then
electrotransferred onto nitrocellulose filter membranes (NC;
Whatman, Alameda, CA, USA). The membranes were incubated with
antibodies against total NF-κB (p65; Cat. no. 8424; 1:1,000),
phosphorylated (p-)NF-κB (p65; Cat. no. 3033; 1:1,000), or GAPDH
(Cat. no. 8884; 1:1,000) (all from Cell Signaling Technology,
Beverly, MA, USA) overnight at 4°C. The membranes were then
incubated with an IRDye 800CW-conjugated secondary antibody
(Rockland, Hamburg, Germany) for 1 h at room temperature. Images
were acquired using an Odyssey infrared imaging system (LI-COR
Biosciences, Inc., Lincoln, NE, USA).
Flow cytometric analysis
Briefly, the colons were longitudinally cut and
washed in PBS containing 1% fetal bovine serum and 1 mM DTT for 10
min at room temperature 3 times. Subsequently, the colon tissues
were digested in complete RPMI-1640 medium (Invitrogen)
supplemented with 0.5 mg/ml collagenase type VIII (Sigma-Aldrich,
St. Louis, MO, USA), 20 mg/ml DNase I (Roche Diagnostics, Mannheim,
Germany) for 60 min at 37°C in a shaking water bath. The cell
suspension was passed through a 100–70 μm filter and then
resuspended in 1.077 g/cm3 iso-osmotic metrizamide
medium (Accurate Chemical & Scientific Corp., Westbury, NY,
USA). Following centrifugation at 1,500 × g for 15 min at room
temperature, the low-density fraction was collected. For FACS
analysis of the colon samples, the cells were stained with
antibodies to CD45 (17-0451), CD11c (12-0114), MHC-II (56-5321),
F4/80 (15-4801), CD11b (11-0112), Ly-6G (Gr-1; 53-5931) (all from
eBioscience, San Diego, CA, USA) and CD206 (141711) (from
BioLegend, San Diego, CA, USA). The influx of neutrophils,
dendritic cells (DCs) and macrophages was determined by the
frequency of macrophages, neutrophils and DCs in CD45+
cells in the LP of colons.
Isolation and stimulation of murine
macrophages
To isolate the LP-macrophages from the lamina
propria of the mouse colons, the colons were washed with PBS and
cut into small sections. The latter were then incubated with 5 mM
EDTA and 3% FCS in Ca2+- and Mg2+-free Hanks
balanced salt solution for 30 min at 37°C with stirring. The small
sections were collected and were then digested with RPMI-1640
containing 5% FCS, 1 mg/ml collagenase type IV and 0.1 mg/ml DNase
for 1 h at 37°C. The liberated cells were collected through a
stainless steel sieve. Following centrifugation (500 × g, 4°C) and
washing with PBS 3 times, the intestinal macrophages were enriched
by positive selection with anti-CD11b Dynabeads following the
manufacturer's instructions (Miltenyi Biotec GmbH, Bergisch
Gladbach, Germany). The cells obtained were tested for the
expression of macrophage markers by flow cytometry
(CD45+CD11b+CD11c−F4/80+);
cells with a purity >85% were used in the experiments.
LP-macrophages (5×104) were cultured in
complete RPMI-1640 medium and treated with 100 ng/ml LPS for 6 h.
Supernatants were collected for the measurement of cytokine levels
and the attached cells were lysed for RNA extraction. Briefly, the
levels of the cytokines, IL-1β, IL-6, IL-10 and TNF-α, were
evaluated in the supernatant from the macrophage cultures by
enzyme-linked immunosorbent assay (ELISA), following the
manufacturer's instructions (R&D Systems, Inc., Minneapolis,
MN, USA).
Migration and phagocytosis of
LP-macrophages
The migration of the LP-macrophages induced by C-X-C
motif chemokine 12 [CXCL12, also known as stromal cell-derived
factor 1 (SDF-1)] was assessed using a 48-well microchemotaxis
chamber (Neuro Probe, Gaithersburg, MD, USA). CXCL12 (50 nM;
250-20B, PeproTech, Rocky Hill, NJ, USA) was placed in wells (30
μl) of the lower compartment, and LP-macrophages (50
μl of a 5×10*4/ml suspension) were seeded in the
wells of the upper compartment. The chamber was incubated in a
humidified environment at 37°C with 5% CO2 for 4 h. The
membrane was removed, fixed in methanol, and stained with crystal
violet solution for 30 min followed by destaining with water. The
cells that had migrated across the membrane were counted using a
microscope (Leica TCS SP8 CARS confocal microscope, Leica
Microsystems GmbH, Wetzlar, Germany). The chemotaxis index was
calculated as the ratio of the number of cells that had migrated
towards the chemoattractant divided by the number of cells that had
migrated towards the medium.
The LP-macrophages (5×10*4) were infected
with E. coli at an MOI of 1:10 for 1 h in complete RPMI-1640
medium without antibiotics, and then incubated in complete medium
containing 40 μg/ml gentamycin for 120 min. The cells were
then lysed and seeded on LB agar plates. Following overnight
incubation at 37°C, bacterial colonies (CFUs) were counted as a
measure of intracellular bacteria.
Immunofluorescence staining
For the human tissues, 4-μm-thick frozen
sections of intestinal specimens were fixed in cold acetone for 10
min at −20°C and blocked with 5% BSA for 1 h at room temperature,
then incubated with a primary rabbit anti-human 5-HT2AR
antibody (ab66049; rabbit polyclonal to 5-HT2AR; reacts
to mouse, rat, human; Abcam, Cambridge, UK; 1:500) overnight at
4°C. Mouse anti-human CD68 antibody (MCA5709; mouse anti-human
CD68, monoclonal antibody; AbD Serotec, Kidlington, UK; 1:500)
(overnight at 4°C) was subsequently used to detect the macrophages.
For the murine colon tissues, 5-HT2AR expression was
detected by overnight incubation at 4°C with rabbit anti-mouse
5-HT2AR antibody (ab66049; rabbit polyclonal to
5-HT2AR; reacts to mouse, rat, human; Abcam; 1:300).
Subsquently, rat anti-mouse CD68 antibody (MCA1957GA; rat
anti-mouse CD68, monoclonal antibody; AbD Serotec; 1:500) was used
overnight at 4°C. For both analyses, Alexa-Fluor 488- and
Alexa-Fluor 555-conjugated antibodies [goat anti-rabbit IgG
(ab150077, goat anti-rat IgG (ab150158) and goat anti-mouse IgG
(ab150118); all from Abcam, Cambridge, UK; 1:1,000; 1 h at room
temperature] were used as secondary antibodies, followed by
incubation with 1 μg/ml DAPI (20 min, room temperature). The
sections were finally visualized under a confocal microscope
(Olympus, Tokyo, Japan). Images were captured using FluoView
software.
Transfection with small interfering RNA
(siRNA)
siRNA targeting the 5-HT2AR (Genepharm
Biotech, Shanghai, China) was synthesized (GACAACUGUCGUGAUUAUUTT)
and control siRNA (UUCUCCGAACGUGUCACGUTT) was also used. BMDMs were
isolated from bone marrow cells obtained from the mice. Three
C57BL/6 male mice (8 to 10 weeks old, weighing 20 g) were
sacrificed as described for the mice in the other experiments, and
bone marrow cells were isolated from femurs and cultured with
RPMI-1640 supplemented with 10% FBS, 1% penicillin and streptomycin
(Invitrogen) and 10% L929 conditioned medium. The culture fluid was
exchanged for fresh culture medium every 4 days. Under these
conditions, adherent macrophage monolayers were obtained within
8–10 days. The BMDMs were then transfected with the siRNAs as
previously described (17).
Following transfection, the BMDMs (1×106/ml) were
pre-incubated with ketanserin (10 μM) for 10 min and then
stimulated with LPS (100 ng/ml) and 5-HT (100 nmol/l) for 12 h. RNA
was then extracted for PCR.
Statistical analysis
The statistical significance of the differences
between the treatment and control groups was determined using a
Student's t-test. Data were analyzed with one-way analysis of
variance (ANOVA), followed by the Student's t-test for experiments
involving only 2 groups, and Dunnett's t-test for experiments
involving >2 groups. All data are expressed as the means ±
standard deviation (SD). Statistical significance was set at
P<0.05.
Results
5-HT2AR expression is elevated
in both patients with IBD and in mice with experimental colitis,
and is specifically enhanced in macrophages
We first examined the expression of
5-HT2AR in patients with IBD and in mice with
DSS-induced experimental colitis. The 5-HT2AR mRNA level
was increased in the patients with IBD (Fig. 1A) compared to the normal tissue
specimens, and 5-HT2AR protein expression was also
elevated in the patients with IBD, as detected by
immunofluorescence staining (Fig.
1B). 5-HT2AR is known to be expressed on the surface
of lymphocytes, natural killer (NK) cells and
monocytes/macrophages/DCs (18,19). We observed a specific
co-localization of 5-HT2AR with the CD68 macro-with the
CD68 macrophage marker (Fig. 1B),
reflecting that 5-HT2AR expression is enhanced in
macrophages. The 5-HT2AR expression level was also
upregulated in the mice with DSS-induced experimental colitis
(Fig. 1C and D) and was
co-localized with CD68 postivity (Fig. 1E). These data suggest that the
synthesis of 5-HT2AR is induced in the inflamed colon
and that it is mostly expressed in macrophages.
The administration of ketanserin
attenuates the development of DSS-induced acute experimental
colitis in mice
To investigate whether ketanserin reduces
susceptibility to colitis, we induced colitis in mice using DSS.
The mice with DSS-induced colitis exhibited a continuous decrease
in body weight from day 4 to day 7 and shortened colon lengths. By
contrast, the administration of ketanserin during the induction of
colitis significantly prevented the decrease in body weight
(Fig. 2A) and colon shortening
(Fig. 2B). A histological
examination of the colons of the mice with DSS-induced colitis
revealed severe inflammation with ulcerative lesions, loss of
crypts and the infiltration of inflammatory cells, whereas
treatment with ketanserin alleviated these histological changes and
damage to the colon, characterized by a decrease in the loss of
architecture, fewer ulcerative lesions, and a decrease in
inflammatory cell infiltration into the inflamed mucosa (Fig. 2C and D). Our data therefore
suggest that ketanserin exerts a potent therapeutic effect,
ameliorating DSS-induced colitis.
DSS is directly toxic to the colonic epithelium and
triggers inflammation in the gut with high levels of inflammatory
mediators, such as iNOS, IL-1β and IL-6 (20,21). We thus evaluated the effects of
ketanserin on the production of inflammatory mediators in the
colonic mucosa by RT-qPCR. Compared with the vehicle-treated mice
with colitis (DSS group), the ketanserin-treated mice exhibited
significantly lower mRNA levels of keratinocyte-derived chemokine
(KC; a major chemoattractant for neutrophils), iNOS, TNF-α, IL-1β
and IL-6 (Fig. 2E). Moreover, the
transcript levels of IL-10 were found to be higher in the
ketanserin-treated mice. The levels of CXCL12, the major
chemoattractant for DCs, remained unaltered.
Ketanserin reduces the infiltration of
neutrophils and macrophages into the intestinal mucosa in
colitis
Innate immune cells are the major sources of
inflammatory mediators in DSS-induced colitis (22,23). We thus investigated whether the
effects of ketanserin alter the infiltration of innate immune
cells, thus influencing the inflammatory response in the colon. In
the mice with DSS-induced colitis, the percentage of neutrophils,
macrophages and DCs in the CD45+ cells was significantly
increased in the colonic lamina propria on day 7 compared with the
control mice (Fig. 3A).
Consistently, treatment with ketanserin resulted in a decrease in
the influx of neutrophils and macrophages into the inflamed colons
of the mice (Fig. 3B). Minor
differences in the relative percentage of DCs in the
CD4+ T cell population were observed. We also found that
the colonic macrophages in the ketanserin-treated group expressed
higher levels of CD206, a marker of M2 macrophages, compared to the
vehicle-treated mice with colitis (DSS group; Fig. 3A and C).
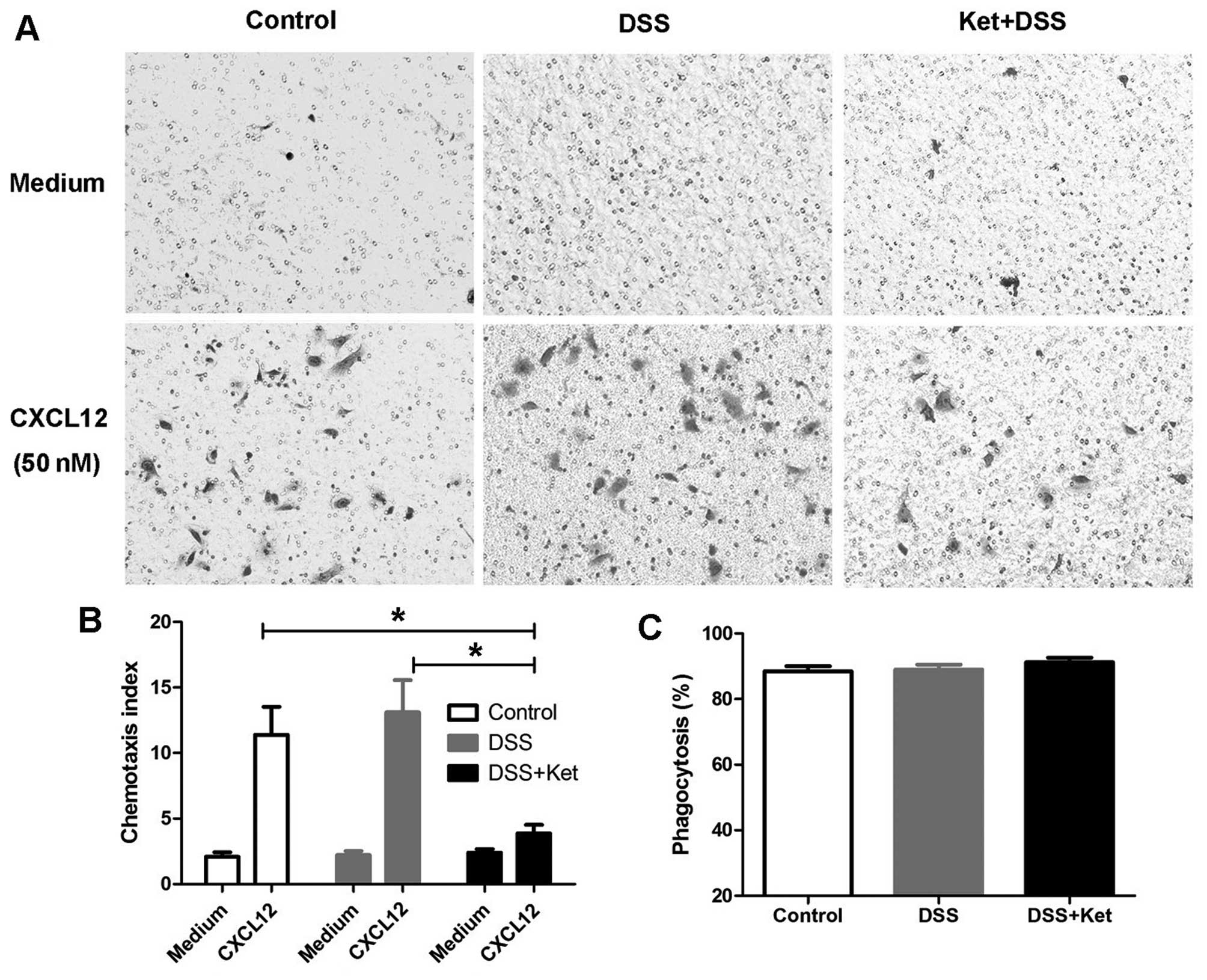
Ketanserin inhibits the migration of
LP-macrophages without affecting the ability of bacterial
phagocytosis
As the ketanserin-treated mice exhibited a reduced
influx of macrophages into the colonic lamina propria, we then
determined whether ketanserin suppresses the chemotaxis of
macrophages. As CXCL12 is a critical regulator of macrophage
migration (24,25), the migration of LP-macrophages
through an 8-μm filter towards CXCL12 (50 nM) placed in the
lower chamber was examined. The chemotaxis index indicated that
ketanserin significantly reduced the migration of macrophages
towards CXCL12 (Fig. 4A and
B).
It has been shown that 5-HT modulates the activity
of the phagocytosis of bacteria by macrophages through 5-HT
receptors (26). Thus, to
determine whether ketanserin affects the phagocytic ability of
macrophages, LP-macrophages were infected with E. coli and
viable intracellular bacteria CFUs were counted in order to
evaluate bacterial phagocytosis. We found that the ability of the
LP-macrophages to phagocytize E. coli in the
ketanserin-treated mice with colitis did not differ from that in
the vehicle-treated mice with colitis (DSS group; Fig. 4C).
Ketanserin inhibits the expression of
pro-inflammatory mediators and influences the polarization of
LP-macrophages
We then evaluated the release of cytokines in the
supernatants of LP-macrophages obtained from the inflamed colon. In
line with our hypothesis, the secretion of TNF-α, IL-1β and IL-6
was significantly decreased in the ketanserin-treated mice
(Fig. 5A–C). In addition, the
ketanserin-treated mice produced higher amounts of IL-10 (Fig. 5D).
Since the LP-macrophages from the ketanserin-treated
mice with colitis exhibited a high level of CD206, a marker of M2
macrophages (as shown by FACS analysis), we therefore measured the
mRNA levels of iNOS, CD32 and IL-12 p40, as M1 polarization
markers, and the levels of CD206 and IL-10, as M2 polarization
markers in the LP-macrophages in all 3 groups. Of note, the
LP-macrophages from the ketanserin-treated mice exhibited a notably
decreased expression of iNOS, CD32 and IL-12 p40 (Fig. 5E), but elevated levels of CD206
and IL-10 (Fig. 5F). Taken
together, these data indicate that ketanserin modulates
LP-macrophage function by reducing the production of
pro-inflammatory cytokines and promoting M2 as opposed to M1
polarization.
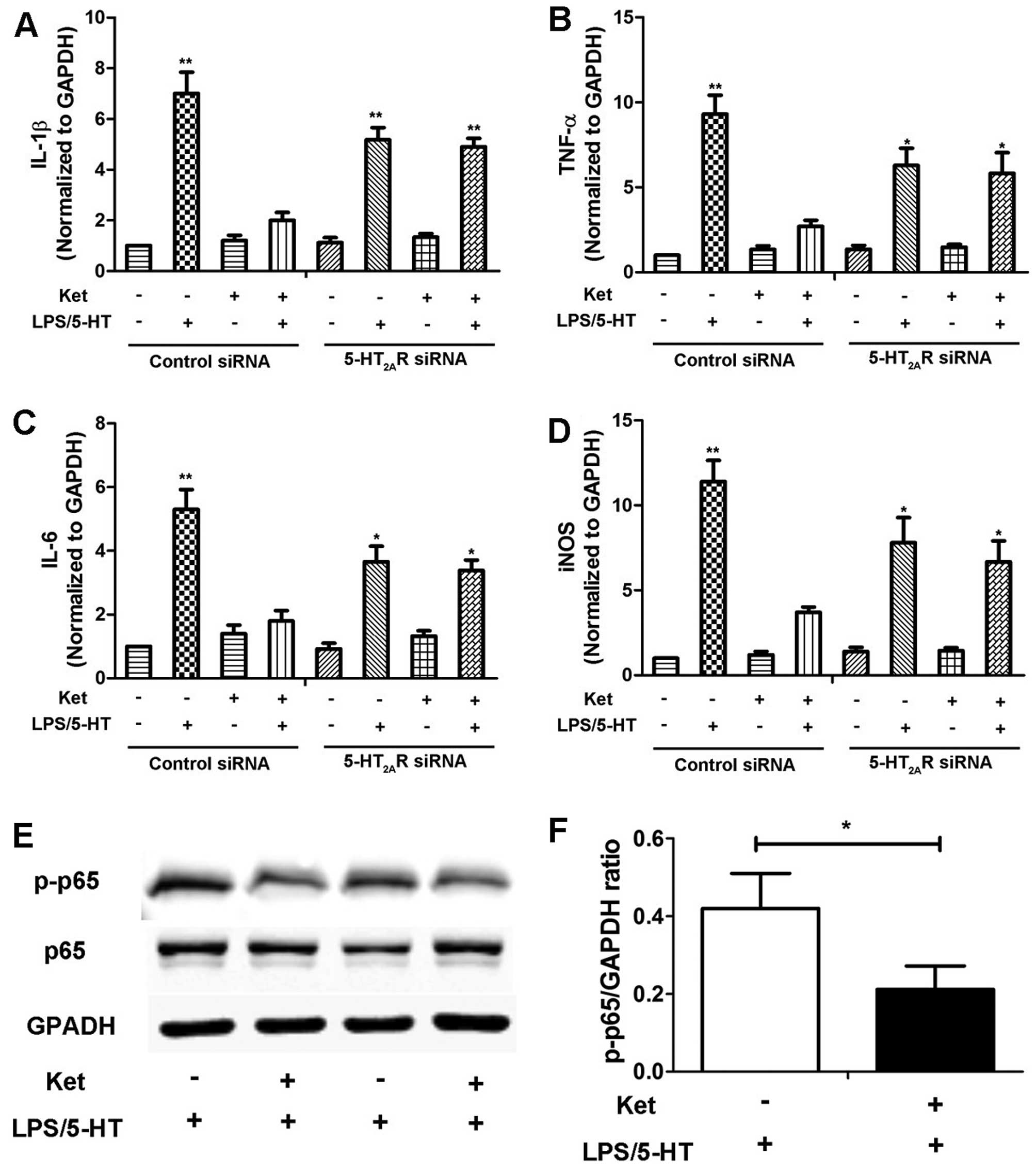
Ketanserin inhibits the expression of
pro-inflammatory cytokines in BMDMs through
5-HT2AR/NF-κB
In order to determine whether 5-HT2AR
plays a role in the effects of ketanserin, we used
5-HT2AR siRNA to downregulate the expression of
5-HT2AR in BMDMs. The LPS/5HT-induced expression of the
pro-inflammatory cytokines, IL-1β (Fig. 6A), TNF-α (Fig. 6B), IL-6 (Fig. 6C) and iNOS (Fig. 6D), was inhibited by treatment with
ketanserin in the BMDMs. The knockdown of 5-HT2AR by
siRNA partly abolished the inhibitory effects of ketanserin on the
expression of these pro-inflammatory cytokines in the BMDMs
stimulated with LPS plus 5-HT (Fig.
6). In a previous study, it was suggested that the
anti-inflammatory effects of ketanserin are partly dependent on the
arterial baroreflex (13). In the
present study, we found that ketanserin decreased the levels of
inflammatory cytokines in BMDMs, suggesting that the
anti-inflammatory effects of ketanserin are not entirely dependent
on the baroreflex. Moreover, these effects were dimished by
transfection with siRNA targeting 5-HT2AR, thus
demonstrating that the inhibitory effects of ketanserin are
partly-dependent on 5-HT2AR.
NF-κB is a critical transcription factor for the
inflammatory response (27). It
functions as a pro-inflammatory factor and participates in the
pathophysiology of IBD (28).
Thus, in order to elucidate the mechanisms responsible for the
anti-inflammatory effects of ketanserin, we examined its effects on
the activation of the NF-κB pathway in BMDMS. Changes in the levels
of p-NF-κB p65 in the BMDMs treated with or without ketanserin were
evaluated by western blot analysis. As demonstrated in Fig. 6E and F, ketanserin markedly
decreased the protein level of p-p65 compared with the BMDMs not
treated with ketanserin. These data indicate that
5-HT2AR/NF-κB may play a role in the anti-inflammatory
effects of ketanserin in macrophages.
Discussion
The unresolved inflammatory state in the intestinal
tract, which was caused by failing to switch from a
pro-inflammatory response to an inflammation-resolving
anti-inflammatory response, is a characteristic of IBD (29). Both macrophages and DCs actively
promote the transition from an inflammatory state to a homeostatic
state following immune system activation, and unresolved
inflammation is associated with IBD (30). Macrophages are a highly
heterogeneous population of cells with a continuum of activation
states (31). Evidence indicates
that of the macrophages in the lamina propria, the pro-inflammatory
M1 subtype is more prominent in patients with CD and less prominent
in patients with UC (32).
In the present study, we found that
5-HT2AR, which was elevated in colitis, was specifically
enhanced in CD68+ macrophages. Treatment with
ketanserin, a selective 5-HT2AR antagonist,
significantly decreased the DSS-induced production of iNOS and
pro-inflammatory cytokines (TNF-α and IL-6) in the colons of mice
with experimental colitis, with a marked reduction in the
infiltration of neutrophils and macrophages in the intestinal
mucosa, without a significant difference in the frequency of DCs.
Furthermore, ketanserin inhibited the production of iNOS, CD32 and
IL-12 in the LP-macrophages upon challenge with LPS, whereas the
production of IL-10 and CD206 increased. This indicates that
ketanserin may influence macrophages by promoting their
anti-inflammatory function and alterating the polarization from M1
to M2.
The change in the cytokine profile in the
ketanserin-treated macrophages is consistent with that of other
studies. Ahonen et al observed significantly higher levels
of IL-1 and IL-6 in LPS-stimulated macrophages following incubation
with 5-HT in their study (33).
In the study by Liu et al, ketanserin markedly reduced the
expression of iNOS, TNF-α and IL-6, as well as that of reactive
oxygen species in RAW264.7 cells upon challenge with LPS (12).
The murine chemokines, KC and CXCL12, are required
for neutrophil and macrophage recruitment, respectively to the
sites of inflammation. Our results indicated that ketanserin
induced a reduction in the KC level along with a considerable
decrease in the number of macrophages in the colonic mucosa,
suggesting that the efficacy of ketanserin may be related to a
lower neutrophil and macrophage influx to the site of injury. In a
previous study, there was also a significant decrease in macrophage
infiltration in TPH1−/− mice with DSS-induced colitis
(5). In addition, we demonstrated
that ketanserin plays a direct inhibit role in the migration of
macrophages, without affecting macrophage phagocytosis.
We also examined 5-HT2AR expression in
colitis and found that macrophages expressed high levels of
5-HT2AR in the inflamed colon. 5-HT2AR
expression has been also shown to be increased in other
inflammatory conditions, such as allergic contact dermatitis
(34). Furthermore, we
demonstrated that 5-HT2AR siRNA partly reversed the
anti-inflammatory effects of ketanserin in BMDMs, thus
demonstrating that the inhibitory effects of ketanserin are
partly-dependent on 5-HT2AR. In addition, we
demonstrated that ketanserin inhibits NF-κB activation in BMDMs.
These findings reveal an essential role of 5-HT2AR/NF-κB
in the effects of ketanserin on the release of cytokines.
In conclusion, the results presented herein
demonstrate that treatment with ketanserin attenuated the
progression of DSS-induced colitis in mice. The mechanisms
responsible for these effects include the inhibition of the release
of inflammatory cytokines and the inhibition of the migration of
macrophages, but not of their phagocytic ability. Ketanserin also
promoted the M2 polarization of colonic macrophages. Moreover,
5-HT2AR/NF-κB may play a role in the therapeutic effects
of ketanserin in colitis.
References
|
1
|
Ravi A, Garg P and Sitaraman SV: Matrix
metalloproteinases in inflammatory bowel disease: Boon or a bane?
Inflamm Bowel Dis. 13:97–107. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Linden DR, Chen JX, Gershon MD, Sharkey KA
and Mawe GM: Serotonin availability is increased in mucosa of
guinea pigs with TNBS-induced colitis. Am J Physiol-Gastr L.
285:G207–G216. 2003.
|
|
3
|
Oshima S, Fujimura M and Fujimiya M:
Changes in number of serotonin-containing cells and serotonin
levels in the intestinal mucosa of rats with colitis induced by
dextran sodium sulfate. Histochem Cell Biol. 112:257–263. 1999.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Bishop AE, Pietroletti R, Taat CW,
Brummelkamp WH and Polak JM: Increased populations of endocrine
cells in Crohn's ileitis. Virchows Arch A Pathol Anat Histopathol.
410:391–396. 1987. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Ghia JE, Li N, Wang HQ, Collins M, Deng
YK, El-Sharkawy RT, Cote F, Mallet J and Khan WI: Serotonin has a
key role in pathogenesis of experimental colitis. Gastroenterology.
137:1649–1660. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Coates MD, Mahoney CR, Linden DR, Sampson
JE, Chen J, Blaszyk H, Crowell MD, Sharkey KA, Gershon MD and Mawe
GM: Molecular defects in mucosal serotonin content and decreased
serotonin reuptake transporter in ulcerative colitis and irritable
bowel syndrome. Gastroenterology. 126:1657–1664. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Bischoff SC, Mailer R, Pabst O, Weier G,
Sedlik W, Li Z, Chen JJ, Murphy DL and Gershon MD: Role of
serotonin in intestinal inflammation: Knockout of serotonin
reuptake transporter exacerbates 2,4,6-trinitrobenzene sulfonic
acid colitis in mice. Am J Physiol Gastrointest Liver Physiol.
296:G685–G695. 2009. View Article : Google Scholar
|
|
8
|
Regmi SC, Park SY, Ku SK and Kim JA:
Serotonin regulates innate immune responses of colon epithelial
cells through Nox2-derived reactive oxygen species. Free Radic Biol
Med. 69:377–389. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Wang D, Chen T, Gao Y, Quirion R and Hong
Y: Inhibition of SNL-induced upregulation of CGRP and NPY in the
spinal cord and dorsal root ganglia by the 5-HT(2A) receptor
antagonist ketanserin in rats. Pharmacol Biochem Behav.
101:379–386. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Ito T, Ikeda U, Shimpo M, Yamamoto K and
Shimada K: Serotonin increases interleukin-6 synthesis in human
vascular smooth muscle cells. Circulation. 102:2522–2527. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Lau WK, Chan SC, Law AC, Ip MS and Mak JC:
The role of MAPK and Nrf2 pathways in ketanserin-elicited
attenuation of cigarette smoke-induced IL-8 production in human
bronchial epithelial cells. Toxicol Sci. 125:569–577. 2012.
View Article : Google Scholar
|
|
12
|
Liu C, Zhang X, Zhou JX, Wei W, Liu DH, Ke
P, Zhang GF, Cai GJ and Su DF: The protective action of ketanserin
against lipopolysaccharide-induced shock in mice is mediated by
inhibiting inducible NO synthase expression via the MEK/ERK
pathway. Free Radic Biol Med. 65:658–666. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Liu C, Zhang GF, Song SW, Cai GJ, Liu WH,
Miao CY and Su DF: Effects of ketanserin on endotoxic shock and
baroreflex function in rodents. J Infect Dis. 204:1605–1612. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Martinez FO, Sica A, Mantovani A and
Locati M: Macrophage activation and polarization. Front Biosci.
13:453–461. 2008. View
Article : Google Scholar
|
|
15
|
Mosser DM and Edwards JP: Exploring the
full spectrum of macrophage activation. Nat Rev Immunol. 8:958–969.
2008. View
Article : Google Scholar : PubMed/NCBI
|
|
16
|
Bettelli E, Carrier Y, Gao W, Korn T,
Strom TB, Oukka M, Weiner HL and Kuchroo VK: Reciprocal
developmental pathways for the generation of pathogenic effector
TH17 and regulatory T cells. Nature. 441:235–238. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Xie N, Cui H, Banerjee S, Tan Z, Salomao
R, Fu M, Abraham E, Thannickal VJ and Liu G: miR-27a regulates
inflammatory response of macrophages by targeting IL-10. J Immunol.
193:327–334. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Hernandez ME, Martinez-Fong D, Perez-Tapia
M, Estrada-Garcia I, Estrada-Parra S and Pavon L: Evaluation of the
effect of selective serotonin-reuptake inhibitors on lymphocyte
subsets in patients with a major depressive disorder. Eur
Neuropsychopharmacol. 20:88–95. 2010. View Article : Google Scholar
|
|
19
|
Young MR and Matthews JP: Serotonin
regulation of T-cell subpopulations and of macrophage accessory
function. Immunology. 84:148–152. 1995.PubMed/NCBI
|
|
20
|
Berndt BE, Zhang M, Chen GH, Huffnagle GB
and Kao JY: The role of dendritic cells in the development of acute
dextran sulfate sodium colitis. J Immunol. 179:6255–6262. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Kitajima S, Takuma S and Morimoto M:
Changes in colonic mucosal permeability in mouse colitis induced
with dextran sulfate sodium. Exp Anim. 48:137–143. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Hunter MM, Wang A, Parhar KS, Johnston MJ,
Van Rooijen N, Beck PL and McKay DM: In vitro-derived alternatively
activated macrophages reduce colonic inflammation in mice.
Gastroenterology. 138:1395–1405. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Qualls JE, Kaplan AM, van Rooijen N and
Cohen DA: Suppression of experimental colitis by intestinal
mononuclear phagocytes. J Leukoc Biol. 80:802–815. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Beider K, Bitner H, Leiba M, Gutwein O,
Koren-Michowitz M, Ostrovsky O, Abraham M, Wald H, Galun E, Peled A
and Nagler A: Multiple myeloma cells recruit tumor-supportive
macrophages through the CXCR4/CXCL12 axis and promote their
polarization toward the M2 phenotype. Oncotarget. 5:11283–11296.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Kim D, Kim J, Yoon JH, Ghim J, Yea K, Song
P, Park S, Lee A, Hong CP, Jang MS, et al: CXCL12 secreted from
adipose tissue recruits macrophages and induces insulin resistance
in mice. Diabetologia. 57:1456–1465. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Maneglier B, Guillemin G, Clayette P,
Rogez-Kreuz C, Brew B, Dormont D, Advenier C, Therond P and
Spreux-Varoquaux O: Serotonin decreases HIV-1 replication in
primary cultures of human macrophages through 5-HT(1A) receptors.
Br J Pharmacol. 154:174–182. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Hayden MS, West AP and Ghosh S: NF-kappaB
and the immune response. Oncogene. 25:6758–6780. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Atreya I, Atreya R and Neurath MF:
NF-kappaB in inflammatory bowel disease. J Intern Med. 263:591–596.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Steinbach EC and Plevy SE: The role of
macrophages and dendritic cells in the initiation of inflammation
in IBD. Inflamm Bowel Dis. 20:166–175. 2014. View Article : Google Scholar :
|
|
30
|
Nathan C and Ding AH: Nonresolving
Inflammation. Cell. 140:871–882. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Murray PJ and Wynn TA: Protective and
pathogenic functions of macrophage subsets. Nat Rev Immunol.
11:723–737. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Smith AM, Rahman FZ, Hayee B, Graham SJ,
Marks DJ, Sewell GW, Palmer CD, Wilde J, Foxwell BM, Gloger IS, et
al: Disordered macrophage cytokine secretion underlies impaired
acute inflammation and bacterial clearance in Crohn's disease. J
Exp Med. 206:1883–1897. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Ahonen A, Kyosola K and Penttila O:
Enterochromaffin cells in macrophages in ulcerative colitis and
irritable colon. Ann Clin Res. 8:1–7. 1976.PubMed/NCBI
|
|
34
|
Wetterberg J, Taher C, Azmitia EC and
El-Nour H: Time-dependent modulation of serotonin and its receptors
1A and 2A expression in allergic contact dermatitis. J Eur Acad
Dermatol. 25:1200–1205. 2011. View Article : Google Scholar
|