Introduction
Germline mutations in the tumor suppressor liver
kinase B1 (LKB1), a serine/threonine kinase, lead to Peutz-Jeghers
syndrome, which predisposes patients to gastrointestinal tumors
(1). The loss of LKB1 owing to
mutations is an important cause of tumorigenesis, and the rate of
LKB1 sporadic mutation is high in many types of malignant tumors,
including non-small cell lung cancer (90%) (2), cervical cancer (20%) (3), hepatic cancer (22%) (4) and pancreatic adenocarcinoma (26%)
(5). By contrast, high-throughput
mutation profiling has shown that sporadic LKB1 mutations are rare
in metastatic (3.4%) (6),
advanced (2.1%) (7), and early
(2/15) (8) gastric cancer (GC).
Therefore, the aberrant LKB1-induced dysregulation of signaling
pathways, rather than sporadic mutations in LKB1, is associated
with GC initiation and progression.
As a critical regulator of a variety of signaling
pathways (9), LKB1 phosphorylates
and thus activates adenosine monophosphate (AMP)-activated protein
kinase (AMPK). AMPK suppresses the mammalian target of rapamycin
(mTOR), and thereby inhibits proliferation, polarity and energy
metabolism of cancer cells (10–13). LKBI is also an essential tumor
suppressor in the Wnt/β-catenin (14), Hedgehog (15) and Hippo (10) signaling pathways, where it blocks
the growth, migration, and epithelial-mesenchymal transition of
cancer cells.
In the Hippo signaling pathway, LKB1 phosphorylates
the Yes-associated protein (Yap) indirectly, by phosphorylating
cytoplasmic LATS1/2, which is a pivotal mediator of Hippo
signaling. Phosphorylated (p-)Yap is sequestrated outside the
nucleus; however, in the absence of phosphorylation, it
translocates to the nucleus, where it binds to TEAD and promotes
the transcription of genes involved in the proliferation and
metastasis of cancer cells (10,16,17). However, Nguyen et al showed
that LKB1 inhibits the proliferation of cancer cells by directly
phosphorylating Yap, independently of either large tumor suppressor
(LATS) phosphorylation or AMPK/mTOR signaling [see ref. 10 in the study by Nguyen et al
(11)]. In lung cancer cells,
LKB1 binds directly to the adenomatous polyposis coli
(APC)/glycogen synthase kinase-3β (GSK3β) destruction complex,
inducing the degradation of β-catenin (14), and thus preventing Wnt/β-catenin
signaling. Yap was also reported to protect β-catenin from
degradation in the cytoplasm, by recruiting β-transducin repeat
containing E3 ubiquitin protein ligase (β-TrCP) ubiquitin ligase
away from the Axin/GSK3β destruction complex (18) in cancer cells. However, Imajo
et al (19) reported that
Yap suppresses Wnt/β-catenin signaling by its direct binding to
β-catenin in the cytoplasm, which prevents its nuclear
translocation. Therefore, the underlying mechanism of LKB1 in its
role as a tumor suppressor in cancer cells is uncertain.
A positive correlation was demonstrated between LKB1
expression levels and prognosis of patients with colorectal
(20), liver (21), lung (22) and pancreatic (23) cancer, whereas high LKB1 expression
is associated with poor overall and disease-free survival in
patients with breast cancer (24). By contrast, the association
between LKB1 and the clinicopathological characters of patients
with GC has not been clearly established. In the present study,
using immunohistochemical (IHC) staining, we found a negative
correlation between LKB1 expression levels and the nuclear staining
of Yap and β-catenin in GC tissues. A clinicopathological analysis
revealed that LKB1 correlated negatively with lymph node
metastasis, tumor infiltration, and TNM stage in GC and positively
with the longer overall survival of GC these patients. We then
found that LKB1 suppresses the proliferation and cell cycle
progression of GC cells, most likely by inhibiting the nuclear
translocation of Yap and β-catenin.
Patients and methods
Patients and specimens
Gastric adenocarcinoma (GAC) tissues were obtained
from 109 patients with GC who underwent radical tumor resection at
the Chinese People's Liberation Army (PLA) General Hospital
(Beijing, China) between 2008 and 2014. Normal controls consisted
of 54 adjacent non-cancerous (NC) tissues from the same patients.
Of these, 54 pairs of GAC and NC tissues were stored in a
refrigerator at −80°C for later use; all 109 GAC and 54 NC tissues
were fixed in formalin and embedded in paraffin. The patients
ranged in age from 31 to 84 years (median, 57 years). The
pathological TNM stages of their tumors were determined based on
the revised 7th edition of the American Joint Committee on Cancer
staging system. Thus, 25 tumors were classified as stage I, 47 as
stage II, 28 as stage III, and 9 as stage IV. The follow-up period
was calculated from the date of surgery until December 15, 2014.
This study was conducted with the approval of the Ethics Committee
of the Chinese PLA General Hospital. All experiments were performed
in accordance with internationally recognized guidelines and the
Helsinki Declaration.
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR) assays
Total RNA was extracted from GAC tissues and GC
cells cultured with TRIzol (Thermo Fisher Scientific, Inc.,
Waltham, MA, USA) according to the manufacturer's instructions. For
mRNA detection, total RNA extracted from cells was reverse
transcribed using the PrimeScript™ RT reagent kit, and qPCR was
then performed using SYBR Premix Ex Taq™ II (both from Takara Bio
Inc., Shiga, Japan) and the following primers: LKB1 forward,
5′-GCCGGGACTGACGTGTAGA-3′ and reverse, 5′-CCCAAAAGGAAGGGAAAAACC-3′;
Yap forward, 5′-GCAACTCCAACCAGCAGCAACA-3′ and reverse, 5′-CGCAGCCTC
TCCTTCTCCATCTG-3′; β-catenin forward,
5′-ATGGGTAGGGCAAATCAGTAAGAGGT-3′ and reverse,
5′-AAGCATCGTATCACAGCAGGTTAC-3′. Relative mRNA levels were analyzed
using the comparative Ct method (2−ΔΔCt). All samples
were normalized to GAPDH levels and the fold-change between samples
was calculated using the comparative Ct method. All experiments
were performed at least 3 times.
Western blotting and antibodies
GAC tissues and GC cells were collected and total
protein was extracted with RIPA Lysis and Extraction Buffer (Thermo
Fisher Scientific, Inc.). For subcellular fractionation, protein
was extracted using NE-PER™ Nuclear and Cytoplasmic Extraction
reagents (Thermo Fisher Scientific, Inc.). Halt™ Phosphatase
Inhibitor Cocktail (Thermo Fisher Scientific, Inc.) was added to
preserve protein phosphorylation as required. Proteins were
separated by electrophoresis on 6 or 12% SDS-PAGE gels, transferred
to PVDF membranes overnight at 4°C and probed with the primary
antibodies against LKB1 (1:1,000, AV41743; Sigma-Aldrich, St.
Louis, MO, USA), Yap (1:1,000, #12395), p-Yap (1:1,000, #13008)
(both from Cell Signaling Technology, Danvers, MA, USA), β-catenin
(1:4,000, ab6302), cyclin D1 (1:1,000, ab131475), CTGF (1:5,000,
ab6992), Histone H3 (1:5,000, ab1791) (all from Abcam, Cambridge,
UK), GAPDH (1:1,000, #5174; Cell Signaling Technology). GAPDH bands
were used as the loading control for cytoplasmic or total proteins,
and histone H3 bands were used as the loading control for nuclear
proteins. Proteins were visualized using an ECL kit (Merck
Millipore Corp., Bedford, MA, USA) according to the manufacturer's
instructions.
Immunohistochemical analysis
Antigen retrieval was performed by incubating the
tumor sections at 95°C for 30 min. They were then incubated
overnight at 4°C with a primary antibody against LKB1 (1:500), Yap
(1:400,), or β-catenin (1:500). Sections were washed, and incubated
with the appropriate biotinylated secondary antibodies [goat
anti-rabbit IgG (H+L) HRP or goat anti-mouse IgG (H+L) HRP,
dilution 1:10,000; Abcam] for 30 min, at 20°C. The reactions were
visualized with 3, 3′-diaminobenzidine/hydrogen peroxide. The
sections were then counterstained with haematoxylin, and washed and
mounted with mounting medium. Antigen expression was scored based
on the staining intensity (0, no staining; 1+, weak
positive staining; 2+, moderate positive staining; and
3+, intense positive staining) and proportion (0, no
cells stained; 1+, <10% positive staining;
2+, 10–50% positive staining; and 3+, >50%
positive staining). The scores obtained from the IHC images were
then calculated and defined as negative (≤1) or positive (2–6),
as previously described (25,26). IHC images were examined by two
experienced pathologists blinded to clinicopathological data. The
final score was evaluated twice.
Culture of GC cells
The human GC cell line AGS was purchased from the
Cell Bank of Shanghai, Institutes for Biological Sciences, China.
The AGS cell line was cultured in RPMI-1640 medium supplemented
with 10% fetal bovine serum (FBS; Gibco, Grand Island, NY, USA) in
a humidified incubator at 37°C and 5% CO2.
Construction of expression vectors and
cell transfection
The vector for the overexpression of LKB1 was
constructed by synthesizing full-length LKB1-DNA and cloning it
into a lentiviral vector-GV341 (pGCMV/MCS/Puro) for mammalian
expression. The GV341-LKB1 vector and the control GV341 empty
vector were produced in HEK293T cells (GenePharma, Shanghai, China)
using the second generation packaging vectors, psPAX2 (Addgene
plasmid 12260) and pMD2.G (Addgene plasmid 12259). The shRNA
sequences targeting LKB1 and Yap were as follows: LKB1,
5′-GCTGAAAGGGATGCTTGAGTATTCAAGAGATACTCAAGCATCCCTTTCAGCTT-3′; Yap,
5′-CCGGGCCACCAAGCTAGATAAAGAACTCGAGTTCTTTATCTAGCTTGGTGGCTTT TTG-3′.
Control shRNAs were constructed using scrambled sequences.
Transfections were performed using Lipofectamine 2000 (Invitrogen,
Carlsbad, CA, USA).
Human GAC AGS cells were seeded in 6-well plates and
incubated for 24 h prior to transfection, at which time they
reached sub-confluence (70–90%). They were then transfected with
GV341-LKB1 or GV341-vector (8 µg) and Lipofectamine 2000 (20
µl) in Opti-MEM medium (Invitrogen). Five hours
post-transfection, the medium was replaced with RPMI-1640 medium
containing 10% FBS. The ability of GV341-LKB1, LKB1-shRNA and
Yap-shRNA to regulate the expression of their targets was confirmed
by western blotting. Geneticin (Gibco) (400 µg/ml) was added
to GV341-LKB1 or GV341 vector-transfected AGS cells to select those
stably expressing LKB1 (LKB1 cells) or the empty vector (Vec
cells). For the simultaneous silencing of LKB1 or Yap, 10 µg
of the respective shRNAs and 25 µl Lipofectamine 2000 were
used. GC cells were harvested for downstream analysis 24–72 h
post-transfection.
In vitro cell proliferation assay
The in vitro proliferation of GC cells was
quantitated using flow cytometric analysis to determine the number
of cells. GC cells were seeded in 6-well plates, transfected with
GV341-LKB1 or GV341-vector, and incubated for 2 h at 37°C for 5
days. The number of cells were determined based on the absorbance
at 450 nm, measured using a BD FACSCalibur (BD Biosciences,
Franklin Lake, NJ, USA). Cell growth in vitro was also
evaluated by clonogenic assays, using a Multiskan spectrum
spectrophotometer (Thermo Fisher Scientific, Inc.). LKB1 and Vec
cells were seeded in 6-well plates (1,000 cells/well) in
triplicate, and then incubated for 10–14 days until visible
colonies had formed. The colonies were fixed with methanol, washed
with water, stained with crystal violet, and then counted.
Cell cycle assessment
GC cells were seeded in 6-cm cell culture dishes,
transfected with GV341-LKB1 or GV341-vector, and harvested by
trypsinization 24–72 h post-transfection. The cells were then fixed
with 70% ethanol overnight at 4°C, and incubated with RNaseA
(R4875; Sigma-Aldrich) (50 µg/ml) for 30 min and
subsequently stained with propidium iodide (PI). Cell cycle
distribution was analyzed by flow cytometric analysis using a BD
FACSCalibur.
In vivo cell proliferation assay
Equal numbers (0.2 ml×1×106) of AGS cells
stably expressing GV341-LKB1 or the empty vector were injected
subcutaneously into the left armpit of 4-week-old immunodeficient
BALB/c nude mice (Charles River, Tokyo, Japan). Twenty-four BALB/c
nude mice were purchased from Vitalriver Company (Beijing, China),
and were sacrificed by high concentrations of CO2. Tumor
formation was measured once every 4 days, and tumor volume was
calculated by 1/2 × length × width2 for 3 weeks. All
mouse experiments were approved by the Ethics Committee of the
Chinese PLA General Hospital. All experiments involving mice were
performed in accordance with internationally recognized guidelines
and the Helsinki Declaration.
Statistical analysis
All experiments were performed at least three times.
The results of experiments comparing two groups were analyzed using
a two-tailed Student's t-test. Pearson's χ2 test was
used to assess the various clinicopathological characteristics as a
function of LKB1 expression determined by IHC analysis. The
correlation between LKB1 and Yap or β-catenin was determined using
Spearman's correlation coefficient test. Cumulative survival curves
were drawn using the Kaplan-Meier method. The difference between
the curves was analyzed using the log-rank test. A P-value <0.05
was considered to indicate a statistically significant difference.
The Statistical Package for the Social Sciences version 19.0 (SPSS
Inc., Chicago, IL, USA) was used for all statistical analyses.
Results
LKB1 expression and the protein ratio of
p-Yap/Yap are low in GAC tissues
To determine the expression of LKB1 and its
association with Yap and β-catenin, we first measured the mRNA and
protein levels in GAC tissues and adjacent NC tissues. The relative
mRNA levels were measured using RT-qPCR in the 54 pairs of GAC and
NC tissues. It was clear that LKB1 mRNA expression was lower
(0.33±0.04 vs. 1.0±0.10, P=0.001) and Yap (1.84±0.13 vs. 1.0±0.11,
P=0.019) and β-catenin (1.63±0.11 vs. 1.0±0.09, P=0.027) mRNA
expression levels were higher in GAC tissues compared with NC
tissues (Fig. 1A). The LKB1,
p-Yap, Yap and β-catenin protein levels were measured using western
blotting, and it was revealed that LKB1 and p-Yap were lower and
that Yap and β-catenin were higher in GAC tissues compared with NC
tissues (Fig. 1B). The relative
LKB1 protein level in GAC tissues was significantly lower in GAC
tissues than in NC tissues (0.37±0.061 vs. 0.59±0.108, P=0.006)
(Fig. 1C). Further analysis
showed that the protein expression ratio of p-Yap/Yap was also
lower in GAC tissues compared with NC tissues (0.179±0.085 vs.
0.8±0.126, P=0.001) (Fig. 1D).
Taken together, these results demonstrate that the LKB1 expression
level and the ratio of p-Yap/Yap were lower in GAC tissues compared
with NC tissues.
LKB1 expression in GAC tissues correlates
negatively with the nuclear accumulation of Yap and β-catenin
In order to determine whether aberrant decreased
LKB1 in GAC tissues was associated with the expression and location
of Yap and β-catenin, we evaluated them using IHC in 109 GAC
tissues and 54 adjacent NC tissues. LKB1 staining was weak in GAC
tissues (43%) but intensely positive in NC tissues (61%)
(χ2=4.678, P=0.0306) (Fig.
2A, panels a and b, and Fig.
2B). Nuclear Yap was negative in NC tissues but intensely
positive in GAC tissues (Fig. 2A,
panels c and d). β-catenin was also negative in NC tissues, whereas
moderately positive staining was noted in GAC tissues (Fig. 2A, panels e and f). Spearman's
correlation coefficient analysis indicated a negative correlation
between LKB1 expression and the nuclear staining of both Yap
(r=−0.6997) and β-catenin (r=−0.3510) (Table I). Therefore, LKB1 expression was
negatively correlated with the nuclear accumulation of Yap and
β-catenin in GAC tissues.
 | Table ICorrelation between LKB1 expression
and clinicopathological features of GC patients. |
Table I
Correlation between LKB1 expression
and clinicopathological features of GC patients.
| Clinicopathological
variables | na | LKB1
| χ2 | P-value |
|---|
| Positive | Negative |
|---|
| All patients | 109 | 47 | 62 | | |
| Age (years) | | | | 3.1320 | 0.0768 |
| <60 | 50 | 17 | 33 | | |
| ≥60 | 59 | 30 | 29 | | |
| Gender | | | | 0.0245 | 0.8757 |
| Male | 71 | 31 | 40 | | |
| Female | 38 | 16 | 22 | | |
| Tumor size
(cm) | | | | 6.9373 | 0.0084c |
| <4 | 32 | 20 | 12 | | |
| ≥4 | 77 | 27 | 50 | | |
| Histopathological
grading | | | | 0.1069 | 0.7437 |
|
Differentiated | 63 | 28 | 35 | | |
|
Undifferentiated | 46 | 19 | 27 | | |
| Lymphatic
invasion | | | | 0.1470 | 0.7015 |
| 0 | 44 | 18 | 26 | | |
| 1–3 | 65 | 29 | 36 | | |
| TNM
classification | | | | | |
| T categories | | | | 13.1648 | 0.0043c |
| T1 | 18 | 12 | 6 | | |
| T2 | 29 | 17 | 12 | | |
| T3 | 45 | 15 | 30 | | |
| T4 | 17 | 3 | 14 | | |
| N categories | | | | 6.3017 | 0.0428c |
| N0 | 29 | 18 | 11 | | |
| N1/2 | 37 | 15 | 22 | | |
| N3 | 43 | 14 | 29 | | |
| TNM stage
(AJCC)b | | | | 8.2511 | 0.0411c |
| I | 25 | 14 | 11 | | |
| II | 47 | 24 | 23 | | |
| III | 28 | 7 | 21 | | |
| IV | 9 | 2 | 7 | | |
| Yap nuclear
location | | | | 53.3644 | 0.0000c |
| Positive | 64 | 9 | 55 | r=−0.6997 | |
| Negative | 45 | 38 | 7 | | |
| β-catenin nuclear
location | | | | 13.4260 | 0.0002c |
| Positive | 59 | 16 | 43 | r=−0.3510 | |
| Negative | 50 | 31 | 19 | | |
LKB1 expression in GAC is positively
correlated with better prognosis
Since the clinicopathological associations between
LKB1 expression and the GAC patients were unclear, we assessed the
clinicopathological characteristics and the prognosis of these GAC
patients. The clinicopathological characteristics of patients with
GC were analyzed using Pearson's χ2 test, which showed a
negative association between LKB1 expression level and tumor size
(P=0.0084), tumor infiltration (P=0.0043), lymph node metastasis
(P=0.0428), and TNM stage (P= 0.0411) (Table I). In cumulative survival curves
obtained using Kaplan-Meier analysis, postoperative overall
survival was longer in patients with LKB1-positive tumors (median,
55 months) than in those with LKB1-negative tumors (median, 34
months; χ2=11.778, P=0.001) (Fig. 2C). In a univariate analysis using
the Cox proportional hazard regression model, LKB1 expression
(P=0.0053), tumor size (P=0.0002), tumor infiltration (P=0.0011),
and lymph node metastasis (P=0.0028) were significantly associated
with cause-specific survival (Table
II). In a multivariate analysis using forward and backward
stepwise Cox regression, LKB1 expression was an independent
prognostic factor (P=0.0077; hazard ratio, 3.527; 95% confidence
interval, 1.491–10.630).
| Table IIUnivariate and multivariate analyses
of overall survival of GC patients. |
Table II
Univariate and multivariate analyses
of overall survival of GC patients.
| Variables | Univariatea
|
Multivariateb
|
|---|
| P-value | HR | 95% CI | P-value |
|---|
| Gender | | | | |
| Male vs.
female | 0.7750 | 1.230 | 0.857–2.035 | 0.0740 |
| Age (years) ≥60 vs.
<60 | 0.1330 | 1.453 | 0.793–2.274 | 0.0628 |
| Tumor size
(cm) | | | | |
| ≥4 vs. <4 | 0.0002c | 5.490 | 2.105–14.143 | 0.0152c |
| Histopathological
grading | | | | |
| Differentiated vs.
undifferentiated | 0.8103 | 1.144 | 0.9102–1.572 | 0.0751 |
| Tumor
infiltration | | | | |
| T3-4 vs. T1-2 | 0.0011c | 1.930 | 1.550–2.584 | 0.0018c |
| Lymph node
metastasis | | | | |
| N1-3 vs. N0 | 0.0028c | 1.562 | 1.460–4.150 | 0.0320 |
| LKB1
expression | | | | |
| Positive vs.
negative | 0.0053c | 3.527 | 1.491–10.630 | 0.0077c |
LKB1 causes the increase in p-Yap
expression and the suppression of GC cell proliferation in
vitro
Although LKB1 has been reported to suppress the
proliferation and survival of many types of cancer cells (11,14,23), its effects on GC cells required
further research. Therefore, AGS GC cells were transfected with a
full-length LKB1 vector (LKB1 cells) or an empty vector (Vec
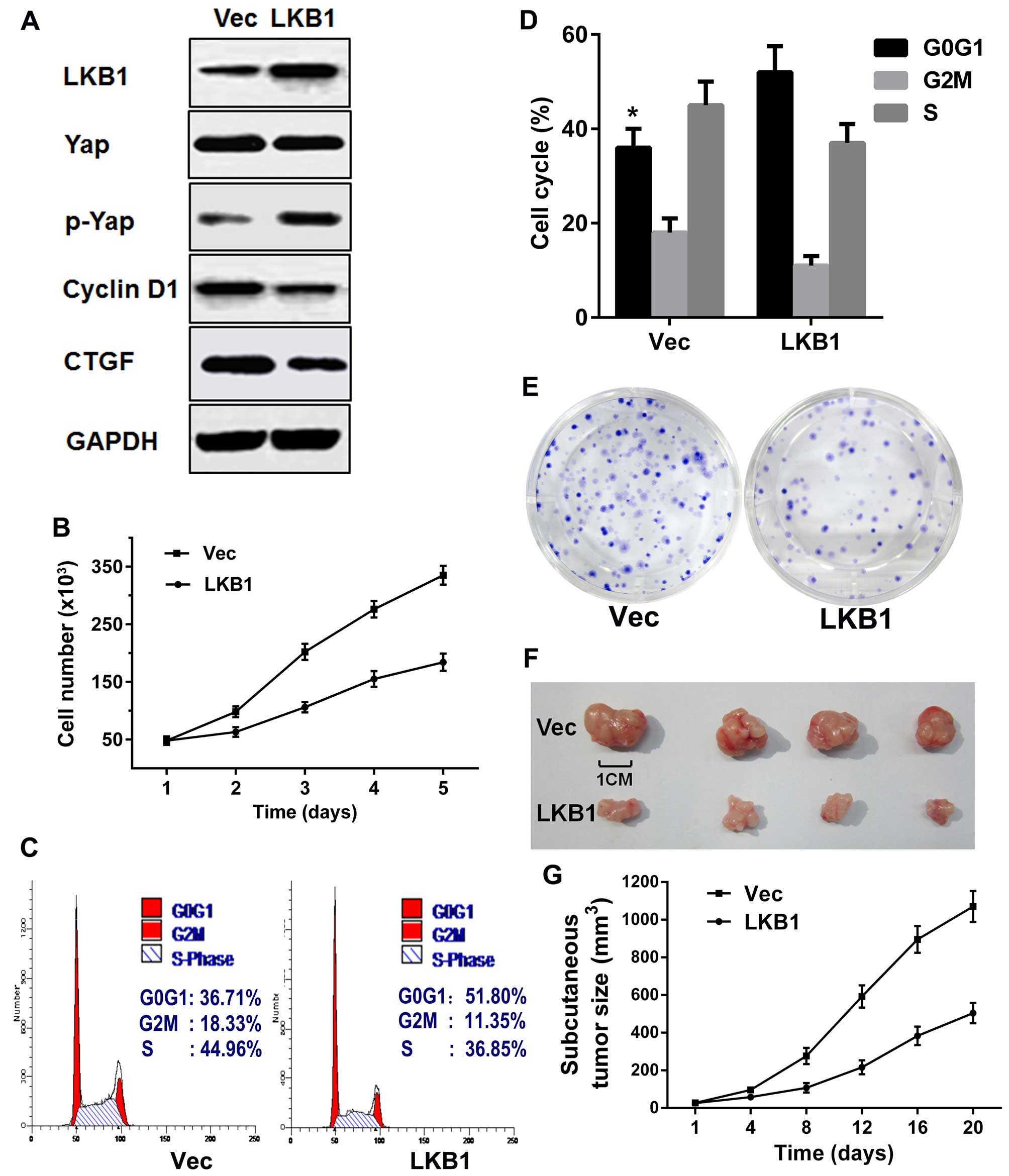
cells). Western blotting revealed that LKB1 and p-Yap expression
levels were notably increased in the LKB1 cells compared with Vec
cells, but Yap expression barely changed (Fig. 3A). Moreover, the target protein of
Yap, CTGF, and the target protein of β-catenin, cyclin D1, which
promote cell proliferation and cell cycle progression, were
decreased in LKB1 cells compared with Vec cells (Fig. 3A). Flow cytometric analysis
carried out at 48 h post-transfection indicated a decrease in the
number of cells in LKB1 cells compared with Vec cells (Fig. 3B). A cell cycle analysis showed an
increase in the number of cells in the G0G1 phase and a decrease in
the numbers of cells in S and G2/M phases in LKB1 vs. Vec cells
(53.01±4.7 vs. 36.41±3.2%, P=0.0022) (Fig. 3C and D). The clonogenic assays
aimed at evaluating the impact of LKB1 on cell growth in
vitro were measured using a Multiskan spectrum
spectrophotometer at 570 nm, and revealed fewer LKB1 compared with
Vec cell colonies (1.98±0.64 vs. 0.85±0.20, P=0.03) (Fig. 3E). These results suggest that LKB1
induced the increase of p-Yap and suppressed proliferation of GC
cells in vitro.
LKB1 suppresses the proliferation of GC
cells in vivo
To evaluate the impact of LKB1 on cell growth in
vivo, equal numbers of LKB1 or Vec cells were injected into the
subcutaneous tissues of BALB/c nude mice, and the volume of the
resulting subcutaneous tumors was measured every 4 days for 3
weeks. Tumors derived from LKB1 cells were found to grow more
slowly and grew to a smaller size compared with those derived from
Vec cells (Fig. 3F and G),
suggesting that LKB1 inhibits the proliferation of GC cells in
vivo.
LKB1 inhibits the nuclear translocation
of Yap in GC cells
To determine whether the location of Yap was related
to the regulation of LKB1 in GC cells, the subcellular expression
levels of p-Yap and Yap were assessed using western blotting in
LKB1 cells and Vec cells. The cytoplasmic p-Yap and Yap expression
levels were increased in LKB1 cells, but nuclear Yap expression was
decreased compared with the Vec cells (Fig. 4A). In addition no p-Yap was
detected in the nucleus of GC cells (Fig. 4A). Further analysis revealed that
the cytoplasmic Yap ratio of LKB1 cells to Vec cells was higher
than the nuclear Yap ratio (1.27±0.26 vs. 0.43±0.18, P=0.017)
(Fig. 4B). However, the relative
mRNA level of Yap was barely altered by LKB1 overexpression
(Fig. 4D). The results suggest
that LKB1 phosphorylates Yap and inhibits its nuclear
translocation, which is consistent with the results of previous
studies (10,16).
LKB1 induces the cytoplasmic accumulation
of β-catenin via Yap in GC cells
To better understand the role of LKB1 in the
regulation of Yap and β-catenin, LKB1 and Yap were individually
silenced using shRNAs in LKB1-overexpressing cells, after which the
levels of Yap and β-catenin in the cytoplasm and nucleus were
determined. The levels of Yap and β-catenin in the cytoplasm were
higher in LKB1-overexpressing GC cells than in Vec cells, whereas
transfection with LKB1-shRNA or Yap-shRNA made these levels
decrease (Fig. 4C). In the
nucleus, however, Yap and β-catenin levels were lower in LKB1 than
in Vec cells, an effect that was reversed by LKB1-shRNA
transfection. In LKB1-overexpressing GC cells, the transfection of
Yap-shRNA led to a further decrease of nuclear Yap and an increase
in nuclear β-catenin (Fig. 4C).
Taken together, these results suggest that LKB1 induces the
accumulation of cytoplasmic β-catenin in GC cells, which, at least
partially, is due to the cytoplasmic accumulation of Yap.
The role of LKB1 and Yap in the regulation of
β-catenin was further examined by measuring the relative mRNA
levels of LKB1, Yap and β-catenin in LKB1 cells and Vec cells.
Silencing of both LKB1 or Yap had almost no effect on β-catenin
mRNA levels (Fig. 4D). Based on
these observations, we concluded that LKB1 expression promotes the
accumulation of cytoplasmic β-catenin in GC cells by inhibiting the
nuclear translocation of Yap, rather than by suppressing the
transcription of β-catenin.
Discussion
LKB1 acts as a critical regulator in the AMPK/mTOR,
Wnt/β-catenin, and Hippo signaling pathways in various types of
cancers (10,11,14). However, the underlying mechanisms
linking LKB1 to its diverse functions in GC remain unclear, and
they are further complicated by the cross-talk between different
signaling pathways. Previous studies have reported that sporadic
LKB1 mutations are rare in metastatic (3.4%) (6) and advanced (2.1%) (7) GC, and thus wild-type LKB1 plays an
important function in the regulation of associated signaling
pathways.
In the present study, we found that the LKB1
expression level and protein ratio of p-Yap/Yap was lower in GAC
tissues compared with adjacent NC tissues. Further IHC analysis
revealed significantly lower LKB1 staining in GAC tissues than in
NC tissues and a negative correlation between LKB1 and the nuclear
staining of Yap and β-catenin. The elevated expression and nuclear
accumulation of Yap has previously been shown to be associated with
promoted GC progression (27).
The nuclear translocation of β-catenin has been implicated in
gastric carcinogenesis in murine adult gastric epithelium (28), and in the invasiveness of GC
(29). In the present study, we
found that LKB1 expression was negatively associated with tumor
size, tumor infiltration, lymph node metastasis, and TNM stage, and
positively associated with longer overall survival of GC patients.
Taken together, these results suggest that LKB1 expression is
decreased in GAC tissues and is negatively associated with the
nuclear accumulation of Yap and β-catenin, which is related to poor
prognosis of GC patients.
Mohseni et al (10) reported that LKB1 inhibits Yap
activation via the canonical Hippo-LATSYap signaling axis, whereas
Nguyen et al (11) found
that LKB1 phosphorylates Yap independently of LATS phosphorylation.
In order to determine whether LKB1 expression affects the
expression and location of Yap, in the present study, we
transfected a full-length LKB1 into the AGS GC cells and observed
an increase in p-Yap expression whereas Yap expression remained
almost unchanged. The potential regulation of Yap by LKB1 in GC
cells was assessed by examining the expression and subcellular
location of Yap. We found that LKB1 overexpression induced an
increase in cytoplasmic p-Yap and the protein ratio of p-Yap/Yap,
leading to a decrease in nuclear unphosphorylated Yap. A decrease
in the expression of CTGF (27),
the target gene of Yap has been noted, but the Yap mRNA level was
not markedly altered between LKB1 and vector-transfected cells.
Therefore, we believe that LKB1 induces the nuclear translocation
of Yap.
Moreover, LKB1 overexpression led to a cell cycle
arrest in G1, a decrease in the cell cycle protein, cyclin D1,
which is an important target gene of β-catenin (30). Previous studies reported that the
aberrant nuclear location of Yap (31) and β-catenin (30), effectors of Hippo and
Wnt/β-catenin signaling pathways, are related to proliferation and
cell cycle progression of cancer cells. Consistent with previous
studies, LKB1-overexpressing GC cells demonstrated a decreased
proliferation in vitro and in vivo. Taken together,
LKB1 inhibits the proliferation of GC cells via suppressing the
nuclear translocation of Yap and associated with the nuclear
accumulation of β-catenin.
LKB1 has been shown to induce the degradation of
β-catenin (32,33) by binding to and activating the
APC/GSK3β destruction complex, resulting in Wnt/β-catenin signaling
inhibition and tumor suppression. Yap has been shown to preserve
cytoplasmic β-catenin by recruiting β-TrCP ubiquitin ligase away
from the Axin/GSK3β destruction complex, leading to the
inactivation of the latter in cancer cells (18). Yap also binds β-catenin directly
in the cytoplasm, which prevents its nuclear translocation and
thereby inhibits the expression of Wnt/β-catenin target genes
(19). In the present study, we
assessed the expression and subcellular location of Yap and
β-catenin in LKB1 cells and in Yap shRNA-transfected LKB1 cells,
and found that LKB1 overexpression induced the accumulation of Yap
and β-catenin in the cytoplasm, thus preventing their nuclear
localization. Interestingly, Yap silencing in LKB1-transfected
cells led to a decrease of β-catenin in the cytoplasm but an
increase of the protein in the nucleus, which suggested that LKB1
and cytoplasmic Yap inhibit the nuclear accumulation of β-catenin
in GC cells, as previously reported (18). By measuring the relative mRNA
levels, we were able to demonstrate that the transcription of
β-catenin was almost unchanged in LKB1 cells and in Yap-silenced
LKB1 cells. Taken together, our results suggest that LKB1
phosphorylates Yap and inhibits its nuclear accumulation via the
Hippo signaling pathway and thereby induces nuclear translocation
of β-catenin in GC cells.
Cross-talk between Yap and β-catenin in cancer cells
has previously been demonstrated in several studies. Rosenbluh
et al reported that the binding of Yap to β-catenin results
in a transcriptional complex that promotes tumor progression in
β-catenin-driven cancer cells, including AGS cells (34). Konsavage et al have
reported that β-catenin drives Yap transcription in colorectal
cancer cells (35). Yap is also a
target of SOX2 and binds β-catenin, thus indirectly dampening Wnt
signals that regulate osteoblastic and adipocytic stem cell
lineages (36). Given the
complexity of the cross-talk between Yap and β-catenin, we did not
further examine the mechanism underlying the regulation of
β-catenin by Yap in GC cells.
In conclusion, the present study revealed that LKB1
expression is decreased in GAC tissues and is correlated negatively
with the nuclear accumulation of Yap and β-catenin. IHC analysis
demonstrated a negative association between LKB1 expression and
tumor size, infiltration, lymph node metastasis and TNM stage.
Higher levels of LKB1 were also associated with the longer overall
survival of GC patients. Further investigation revealed that LKB1
inhibited the nuclear translocation of Yap and β-catenin,
suggesting a mechanism which accounts for its suppression of the
proliferation of GC cells observed in vitro and in
vivo.
Acknowledgments
The study was supported by a grant from the Project
supported by the National High Technology Research and Development
Program of China (no. 2012AA02A504), a grant from the Special
Scientific Research Foundation of Health Sector from the National
Health and Family Planning Commission of China (no. 201302016), a
grant from the PLA Medical Technology Key Project of Scientific
Research in the 12th research projects in 12th Five-Year-Plan (no.
BWS12J049), and a grant from the People's Liberation Army Medical
Technology Project for Fostering Youth (no. 13QNP185),
References
|
1
|
Hemminki A, Markie D, Tomlinson I,
Avizienyte E, Roth S, Loukola A, Bignell G, Warren W, Aminoff M,
Höglund P, et al: A serine/threonine kinase gene defective in
Peutz-Jeghers syndrome. Nature. 391:184–187. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Gill RK, Yang SH, Meerzaman D, Mechanic
LE, Bowman ED, Jeon HS, Roy Chowdhuri S, Shakoori A, Dracheva T,
Hong KM, et al: Frequent homozygous deletion of the LKB1/STK11 gene
in non-small cell lung cancer. Oncogene. 30:3784–3791. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Wingo SN, Gallardo TD, Akbay EA, Liang MC,
Contreras CM, Boren T, Shimamura T, Miller DS, Sharpless NE,
Bardeesy N, et al: Somatic LKB1 mutations promote cervical cancer
progression. PLoS One. 4:e51372009. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Kim CJ, Cho YG, Park JY, Kim TY, Lee JH,
Kim HS, Lee JW, Song YH, Nam SW, Lee SH, et al: Genetic analysis of
the LKB1/STK11 gene in hepatocellular carcinomas. Eur J Cancer.
40:136–141. 2004. View Article : Google Scholar
|
|
5
|
Birnbaum DJ, Adélaïde J, Mamessier E,
Finetti P, Lagarde A, Monges G, Viret F, Gonçalvès A, Turrini O,
Delpero JR, et al: Genome profiling of pancreatic adenocarcinoma.
Genes Chromosomes Cancer. 50:456–465. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Kim S, Lee J, Hong ME, Do IG, Kang SY, Ha
SY, Kim ST, Park SH, Kang WK, Choi MG, et al: High-throughput
sequencing and copy number variation detection using formalin fixed
embedded tissue in metastatic gastric cancer. PLoS One.
9:e1116932014. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Lee J, van Hummelen P, Go C, Palescandolo
E, Jang J, Park HY, Kang SY, Park JO, Kang WK, MacConaill L and Kim
KM: High-throughput mutation profling identifies frequent somatic
mutations in advanced gastric adenocarcinoma. PLoS One.
7:e388922012. View Article : Google Scholar
|
|
8
|
Fassan M, Simbolo M, Bria E, Mafficini A,
Pilotto S, Capelli P, Bencivenga M, Pecori S, Luchini C, Neves D,
et al: High-throughput mutation profiling identifies novel
molecular dysregulation in high-grade intraepithelial neoplasia and
early gastric cancers. Gastric cancer. 17:442–449. 2014. View Article : Google Scholar
|
|
9
|
Martin-Belmonte F and Perez-Moreno M:
Epithelial cell polarity, stem cells and cancer. Nat Rev Cancer.
12:23–38. 2012.
|
|
10
|
Mohseni M, Sun J, Lau A, Curtis S,
Goldsmith J, Fox VL, Wei C, Frazier M, Samson O, Wong KK, et al: A
genetic screen identifies an LKB1-MARK signalling axis controlling
the Hippo-YAP pathway. Nat Cell Biol. 16:108–117. 2014. View Article : Google Scholar :
|
|
11
|
Nguyen HB, Babcock JT, Wells CD and
Quilliam LA: LKB1 tumor suppressor regulates AMP
kinase/mTOR-independent cell growth and proliferation via the
phosphorylation of Yap. Oncogene. 32:4100–4109. 2013. View Article : Google Scholar
|
|
12
|
Patel K, Foretz M, Marion A, Campbell DG,
Gourlay R, Boudaba N, Tournier E, Titchenell P, Peggie M, Deak M,
et al: The LKB1-salt-inducible kinase pathway functions as a key
gluconeogenic suppressor in the liver. Nat Commun. 5:45352014.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Lin YY, Kiihl S, Suhail Y, Liu SY, Chou
YH, Kuang Z, Lu JY, Khor CN, Lin CL, Bader JS, et al: Functional
dissection of lysine deacetylases reveals that HDAC1 and p300
regulate AMPK. Nature. 482:251–255. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Jian SF, Hsiao CC, Chen SY, Weng CC, Kuo
TL, Wu DC, Hung WC and Cheng KH: Utilization of liquid
chromatography mass spectrometry analyses to identify LKB1-APC
interaction in modulating Wnt/β-catenin pathway of lung cancer
cells. Mol Cancer Res. 12:622–635. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Jacob LS, Wu X, Dodge ME, Fan CW, Kulak O,
Chen B, Tang W, Wang B, Amatruda JF and Lum L: Genome-wide RNAi
screen reveals disease-associated genes that are common to Hedgehog
and Wnt signaling. Sci Signal. 4:ra42011. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Gao Y, Zhang W, Han X, Li F, Wang X, Wang
R, Fang Z, Tong X, Yao S, Li F, et al: YAP inhibits squamous
transdifferentiation of Lkb1-deficient lung adenocarcinoma through
ZEB2-dependent DNp63 repression. Nat Commun. 5:46292014. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Hsu YL, Hung JY, Chou SH, Huang MS, Tsai
MJ, Lin YS, Chiang SY, Ho YW, Wu CY and Kuo PL: Angiomotin
decreases lung cancer progression by sequestering oncogenic YAP/TAZ
and decreasing Cyr61 expression. Oncogene. 34:4056–4068. 2015.
View Article : Google Scholar
|
|
18
|
Azzolin L, Panciera T, Soligo S, Enzo E,
Bicciato S, Dupont S, Bresolin S, Frasson C, Basso G, Guzzardo V,
et al: YAP/TAZ incorporation in the β-catenin destruction complex
orchestrates the Wnt response. Cell. 158:157–170. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Imajo M, Miyatake K, Iimura A, Miyamoto A
and Nishida E: A molecular mechanism that links Hippo signalling to
the inhibition of Wnt/β-catenin signalling. EMBO J. 31:1109–1122.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Lee SJ, Kang BW, Chae YS, Kim HJ, Park SY,
Park JS, Choi GS, Jeon HS, Lee WK and Kim JG: Genetic variations in
STK11, PRKAA1, and TSC1 associated with prognosis for patients with
colorectal cancer. Ann Surg Oncol. 21(Suppl 4): S634–S639. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Frau M, Feo F and Pascale RM: Pleiotropic
effects of methionine adenosyltransferases deregulation as
determinants of liver cancer progression and prognosis. J Hepatol.
59:830–841. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Tsai LH, Chen PM, Cheng YW, Chen CY, Sheu
GT, Wu TC and Lee H: LKB1 loss by alteration of the NKX2-1/p53
pathway promotes tumor malignancy and predicts poor survival and
relapse in lung adenocarcinomas. Oncogene. 33:3851–3860. 2014.
View Article : Google Scholar
|
|
23
|
Morton JP, Jamieson NB, Karim SA, Athineos
D, Ridgway RA, Nixon C, McKay CJ, Carter R, Brunton VG, Frame MC,
et al: LKB1 haploinsufficiency cooperates with Kras to promote
pancreatic cancer through suppression of p21-dependent growth
arrest. Gastroenterology. 139:586–597. 597.e581–586. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Bouchekioua-Bouzaghou K, Poulard C,
Rambaud J, Lavergne E, Hussein N, Billaud M, Bachelot T, Chabaud S,
Mader S, Dayan G, et al: LKB1 when associated with methylatedERα is
a marker of bad prognosis in breast cancer. Int J Cancer.
135:1307–1318. 2014.PubMed/NCBI
|
|
25
|
Xi HQ, Wu XS, Wei B and Chen L: Aberrant
expression of EphA3 in gastric carcinoma: correlation with tumor
angiogenesis and survival. J Gastroenterol. 47:785–794. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Wood LD, Calhoun ES, Silliman N, Ptak J,
Szabo S, Powell SM, Riggins GJ, Wang TL, Yan H, Gazdar A, et al:
Somatic mutations of GUCY2F, EPHA3, and NTRK3 in human cancers. Hum
Mutat. 27:1060–1061. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Jiao S, Wang H, Shi Z, Dong A, Zhang W,
Song X, He F, Wang Y, Zhang Z, Wang W, et al: A peptide mimicking
VGLL4 function acts as a YAP antagonist therapy against gastric
cancer. Cancer Cell. 25:166–180. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Radulescu S, Ridgway RA, Cordero J,
Athineos D, Salgueiro P, Poulsom R, Neumann J, Jung A, Patel S,
Woodgett J, et al: Acute WNT signalling activation perturbs
differentiation within the adult stomach and rapidly leads to
tumour formation. Oncogene. 32:2048–2057. 2013. View Article : Google Scholar :
|
|
29
|
Aihara R, Mochiki E, Nakabayashi T,
Akazawa K, Asao T and Kuwano H: Clinical significance of mucin
phenotype, beta-catenin and matrix metalloproteinase 7 in early
undifferentiated gastric carcinoma. Br J Surg. 92:454–462. 2005.
View Article : Google Scholar
|
|
30
|
Zhang N, Wei P, Gong A, Chiu WT, Lee HT,
Colman H, Huang H, Xue J, Liu M, Wang Y, et al: FoxM1 promotes
β-catenin nuclear localization and controls Wnt target-gene
expression and glioma tumorigenesis. Cancer Cell. 20:427–442. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Ehsanian R, Brown M, Lu H, Yang XP,
Pattatheyil A, Yan B, Duggal P, Chuang R, Doondeea J, Feller S, et
al: YAP dysregulation by phosphorylation or ΔNp63-mediated gene
repression promotes proliferation, survival and migration in head
and neck cancer subsets. Oncogene. 29:6160–6171. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Ossipova O, Bardeesy N, DePinho RA and
Green JB: LKB1 (XEEK1) regulates Wnt signalling in vertebrate
development. Nat Cell Biol. 5:889–894. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Yan D, Avtanski D, Saxena NK and Sharma D:
Leptin-induced epithelial-mesenchymal transition in breast cancer
cells requires β-catenin activation via Akt/GSK3- and MTA1/Wnt1
protein-dependent pathways. J Biol Chem. 287:8598–8612. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Rosenbluh J, Nijhawan D, Cox AG, Li X,
Neal JT, Schafer EJ, Zack TI, Wang X, Tsherniak A, Schinzel AC, et
al: β-Catenin-driven cancers require a YAP1 transcriptional complex
for survival and tumorigenesis. Cell. 151:1457–1473. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Konsavage WM Jr, Kyler SL, Rennoll SA, Jin
G and Yochum GS: Wnt/β-catenin signaling regulates Yes-associated
protein (YAP) gene expression in colorectal carcinoma cells. J Biol
Chem. 287:11730–11739. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Seo E, Basu-Roy U, Gunaratne PH, Coarfa C,
Lim DS, Basilico C and Mansukhani A: SOX2 regulates YAP1 to
maintain stemness and determine cell fate in the osteo-adipo
lineage. Cell Rep. 3:2075–2087. 2013. View Article : Google Scholar : PubMed/NCBI
|