Introduction
The skin serves as the physical and chemical barrier
between the interior body and the exterior environment, which
consists of an underlying dermis of mesodermal origin and an
overlaying epidermis of ectodermal origin. Although these two
layers perform different functions, they communicate in various
ways and at different levels. The vital barrier function of skin is
primarily provided by its upper stratified epidermis, composed of
proliferating basal and differentiated suprabasal keratinocytes
(1). To complete these functions,
stem cells in the basal layer are capable of self-renewal
throughout life and they produce daughter cells [transit-amplifying
cells (TAs)] that undergo differentiation (2). Thus, the balance between the
proliferation and differentiation of the basal keratinocytes is
essential for epidermal integrity, and it ensures the occurrence of
tissue renewal which is necessary to complete normal physiological
functions. In addition to maintaining tissue homeostasis, stem
cells residing in the epidermis and hair follicles also participate
in the repair of the epidermis following injury (3,4).
In the skin, different phases of wound healing evolve in dynamic
interactions between epidermal cells, dermal cells and bone
marrow-derived cells. Keratinocytes represent the 'first line of
defense' of the body against the outside environment. Therefore,
they are the first responders to injury. With the release of
inflammatory cytokines and the contraction of collagenous fibers,
the basal epidermal keratinocytes are activated and migrate into
the wound area, where they proliferate and differentiate until the
cellular content of the tissue has been renewed (5). A number of observations have
suggested that the epidermis has an effect on the pathology of
cutaneous scars (6,7). In burn patients, hypertrophic scars
occur more frequently when wounds heal by secondary intention,
whereas early skin grafting appears to suppress scar formation
(8). Funayama et al
demonstrated that keloid-derived fibroblasts co-cultured with
keloid-derived keratinocytes were significantly more proliferative
and resistant to apoptosis than those co-cultured with normal
skin-derived keratinocytes (9).
In a similar co-culture system, it was found that keloid-derived
keratinocytes promoted increased collagen production, as well as
the increased proliferation of normal fibroblasts (10). In particular, keloid-derived
keratinocytes have been shown to exhibit the increased expression
of a set of histone genes essential for DNA replication in
comparison with the expression levels in normal cells, and
keratinocytes in the epidermis of hypertrophic scars have entered
an alternative differentiation pathway and express a proliferative
phenotype (11,12). These results suggest that the
balance between the proliferation and differentiation of
keratinocytes is dysregulated in scar tissues, and that fundamental
abnormalities in keratinocytes may play a more important role in
skin regeneration and its pathogenesis than was previously
appreciated.
In the present study, normal, wound edge and
hypertrophic scar tissues were obtained and analysed using
histological and immunofluorescence methods to investigate the
differences in morphology, as well as keratin expression profiles
during epidermal regeneration. Our results indicated that there was
a variation in the keratin expression profiles in the epidermal
keratinocytes from normal, wound edge and hypertrophic scar
tissues, which corresponded with abnormalities in the structure of
the basement membrane (BM). By using a panel of antibodies to BM
components, we validated our hypothesis and determined that the BM
was a positive regulator of the recruitment of precursor-like cells
to the basal layer of the epidermis. Our data indicate that
alterations in the structure of the BM promotes basal keratinocytes
to adopt a proliferative phenotype both in vivo and in
vitro.
Materials and methods
Tissues samples
Hypertrophic scar tissue (4 females and 3 males; age
range 18–40 years) and wound edge tissue (3 females and 5 males;
age range, 18–40 years) samples were obtained from patients who had
undergone prior reconstructive burn surgery. Normal skin tissue (4
females and 3 males; age range, 18–40 years) adjacent to the scar
excision during surgery was used as a control. Foreskins were also
obtained from males undergoing circumcision (10 males; age range,
18–20 years). The present study was approved by the Ethics
Committee of the Chinese PLA General Hospital (Beijing, China), and
written informed consent was obtained from all individuals prior to
obtaining the samples.
Immunofluorescence staining
Normal skin, wound edge and hypertrophic scar tissue
samples were fixed in 10% buffered formalin, dehydrated through
ethanol solutions with increased concentration successively, and
finally embedded in paraffin, respectively. Paraffin-embedded
tissues were cut into 4-μm-thick sections for staining with
hematoxylin and eosin (H&E), Masson's trichrome, and
methenamine silver (all from Zhongshan Golden Bridge, Beijing,
China). Images were captured using an optical microscope (BX53;
Olympus, Tokyo, Japan). To perform immunofluorescence staining, the
sections were fixed in 4% paraformaldehyde for 30 min, and then
permeabilized in 0.2% Triton X-100 (T8787; Sigma-Aldrich, St.
Louis, MO, USA) in phosphate-buffered saline (PBS) for 10 min. The
sections were incubated with primary anti bodies at 4°C overnight
and then with secondary antibodies for 2 h at room temperature. The
following primary anti bodies were used: mouse anti-human
cytokeratin (CK)10 (1:200, ab111447) and rabbit anti-human CK14
(1:200, ab7800) (both from Abcam, Cambridge, MA, USA), rabbit
anti-human CK5 (1:200, ZA-0518; Zhongshan Goldenbridge, Beijing,
China), mouse anti-human CK19 (1:200, ab53119; Abcam), rabbit
anti-human integrin-β1 (1:200, PB0063; Boster, Wuhan, China); mouse
anti-human integrin-β4 (1:200, ab128068; Abcam), mouse anti-human
laminin (1:200, ZM-0181; Zhongshan Goldenbridge), rabbit anti-human
laminin-5 (1:200, ab14509; Abcam) and mouse anti-human collagen IV
(1:200, ZM-0081; Zhongshan Goldenbridge). The following secondary
antibodies were used: Alexa-Fluor 488-conjugated anti-mouse (1:200,
ab150117) and Alexa-Fluor 594-conjugated anti-rabbit (1:200,
ab150080) (both from Abcam). The nuclei were stained with DAPI
(H-1200; Vector Laboratories, Burlingame, CA, USA).
Immunofluorescence images were captured using a confocal laser
scanning microscope (SP8; Leica, Solms, Germany).
Cell culture and treatment
Primary human epidermal keratinocytes (HEKs) were
isolated from the male foreskins as previously described (13), with minor modifications. The human
immortalized keratinocyte (HaCaT) cells were purchased from the
China Infrastruture of Cell Line Resources (3111C0001CCC000373;
Beijing, China). Both the HEKs and the HaCaT cells were incubated
in EpiLife medium (M-EPI-500-CA; Invitrogen Life Technologies,
Carlsbad, CA, USA) supplemented with 0.06 mM Ca2+, 1%
EpiLife defined growth supplement (S-001-5; Invitrogen Life
Technologies), and 1% penicillin/streptomycin (P1400; Solarbio,
Beijing, China). To investigate the roles of the BM in regulating
keratinocyte behavior in vitro, the HEKs and HaCaT cells
were plated in 60 mm-dishes coated with MaxGel ECM (E0282) and
collagen type IV (C7521) (both from Sigma-Aldrich), respectively.
For Ca2+ treatment, the cells were incubated in EpiLife
medium supplemented with 1.5 mM Ca2+ for 0, 12 and 24 h.
The MaxGel ECM contains human extracellular matrix (ECM) components
including collagens, laminin, fibronectin, tenascin and elastin, as
well as a number of proteoglycans and glycosaminoglycans. The cells
not treated with Ca2+ were used as controls.
Reverse transcription-quantitative
(real-time) PCR (RT-qPCR)
Total RNA was isolated from the cells after they
were subjected to the various treatments using TRIzol reagent
(15596-026; Invitrogen, Carlsbad, CA, USA) and then cDNA was
synthesized using GoScript reverse transcriptase (A5001; Promega,
Madison, WI, USA) according to the manufacturer's instructions. The
primer sequences used for gene amplification are listed in Table I. qPCR reactions were performed
with GoTaq qPCR Master Mix (A6001; Promega) using an ABI 7500
Real-Time PCR system and software (Applied Biosystems, Foster City,
CA, USA). The reaction protocol consisted of the following cycles:
95°C for 15 sec, 55°C for 30 sec, and 72°C for 30 sec for 40 cycles
of PCR amplification.
 | Table IPrimers used for RT-qPCR in this
study. |
Table I
Primers used for RT-qPCR in this
study.
| Transcript | | Primer sequences
(5′→3′) |
|---|
| CK10 | Sense: |
GGCAAAATCAAGGAGTGGTATG |
| Antisense: |
GAAGCAGGATGTTGGCATTATC |
| CK14 | Sense: |
GAAGTGAAGATCCGTGACTGGT |
| Antisense: |
GTGGCTGTGAGAATCTTGTTCC |
| CK19 | Sense: |
GCCACTACTACACGACCATCCA |
| Antisense: |
AGAGCCTGTTCCGTCTCAAACT |
| GAPDH | Sense: |
TGAAGGTCGGAGTCAACGGAT |
| Antisense: |
CTGGAAGATGGTGATGGGATT |
Western blot analysis
Total protein was isolated from the cells and 20
μg of protein was dissolved in substrate-soluble buffer. The
proteins were then separated by SDS-PAGE and transferred onto
polyvinylidene difluoride (PVDF) membranes (IPFL00010; Millipore,
Billerica, MA, USA). After blocking, the blots were probed with the
primary antibodies overnight at 4°C. The antibodies used for
western blot analysis included CK10 (1:1000, ab111447), CK14
(1:500, ab7800), CK19 (1:500, ab53119), integrin-β4 (1:500,
ab128068), and mouse anti-human β-actin antibody (1:5000, ab6276)
(all from Abcam). After washing, the membranes were incubated with
secondary antibodies, which were goat anti-rabbit and goat
anti-mouse IgG conjugated to horseradish peroxidase (sc-2004,
1:1000; sc-2005, 1:1000; Santa Cruz Biotechnology, Santa Cruz, CA,
USA). Immunoreactive bands were detected by enhanced
chemiluminescence (ECL) kit (PE-0010-100; Solarbio, Beijing, China)
and imaged using the ImageQuant LAS 4000 system (GE Healthcare
Bio-Sciences, Pittsburgh, USA).
Statistical analysis
All experiments were repeated at least 3 times,
unless otherwise indicated. Data are presented as the means ±
standard deviation (SD). Statistical analysis was performed using
SPSS 20.0 software. Data comparison was performed using the
independent Student's t-test and one-way ANOVA followed by the LSD
test. A P-value <0.05 was considered to indicate a statistically
significant difference.
Results
Morphological differences in normal,
wound edge and hypertrophic scar tissues
Skin wound repair is a precise remodeling process,
which is divided classically into four overlapping phases and
involves the interaction of many different tissues and cell
lineages (5). In chronic wounds,
however, the normal healing process is interrupted, which results
in a pathological cycle of inflammation and protease release
leading to the development of a pathological scar. In this study,
to evaluate the morphological changes occurring in the regenerating
epidermis during skin wound healing, specimens of normal, wound
edge and hypertrophic scar tissues were obtained in order to
perform routine H&E and Masson's trichrome staining. In the
normal skin samples, the histological structure of the skin was
visible, showing layers of the epidermis and dermis. The epidermis
contained 4 to 5 layers, in which keratinocytes differentiated and
gradually matured into spinous cells, granular cells and cornified
cells during their outward passage (Fig. 1A). Moreover, in the underlying
dermis, fibroblasts made up loose connective tissue, which was rich
in parallel collagenous fibers (Fig.
1D). In the tissues around the wound edge, however, the
epidermis proliferated; it was comprised of multiple layers of
keratinocytes of distinct morphology and was of visibly increased
thickness (Fig. 1B and E). The
epidermis and dermis interlocked through finger-like projections
(called rete ridges), which increased the surface area of contact
between the epidermis and the dermis. In both the normal and wound
edge tissues, the basal keratinocytes were arranged as a regimented
single layer of cuboidal or low columnar cells, which connected to
the spinous cell layers above (Fig.
1A and B). The histological structure of the hypertrophic scar
tissue differed from that of normal skin tissue by having a rich
blood supply, and a thick epidermal layer (Fig. 1C and F). The dermis was composed
largely of dense, disorganized connective tissue that includes
tough collagenous fibers of an irregular shape and a larger
diameter (Fig. 1F). To further
clarify the differences in histological structure among the normal,
wound edge and hypertrophic scar tissues, methenamine silver
staining was performed to assess the formation of the BM, which
indicated that the BM structure was detectable in the sections of
the normal and wound edge tissues (Fig. 1G and H). In the scar tissues,
however, BM staining was absent (Fig.
1I).
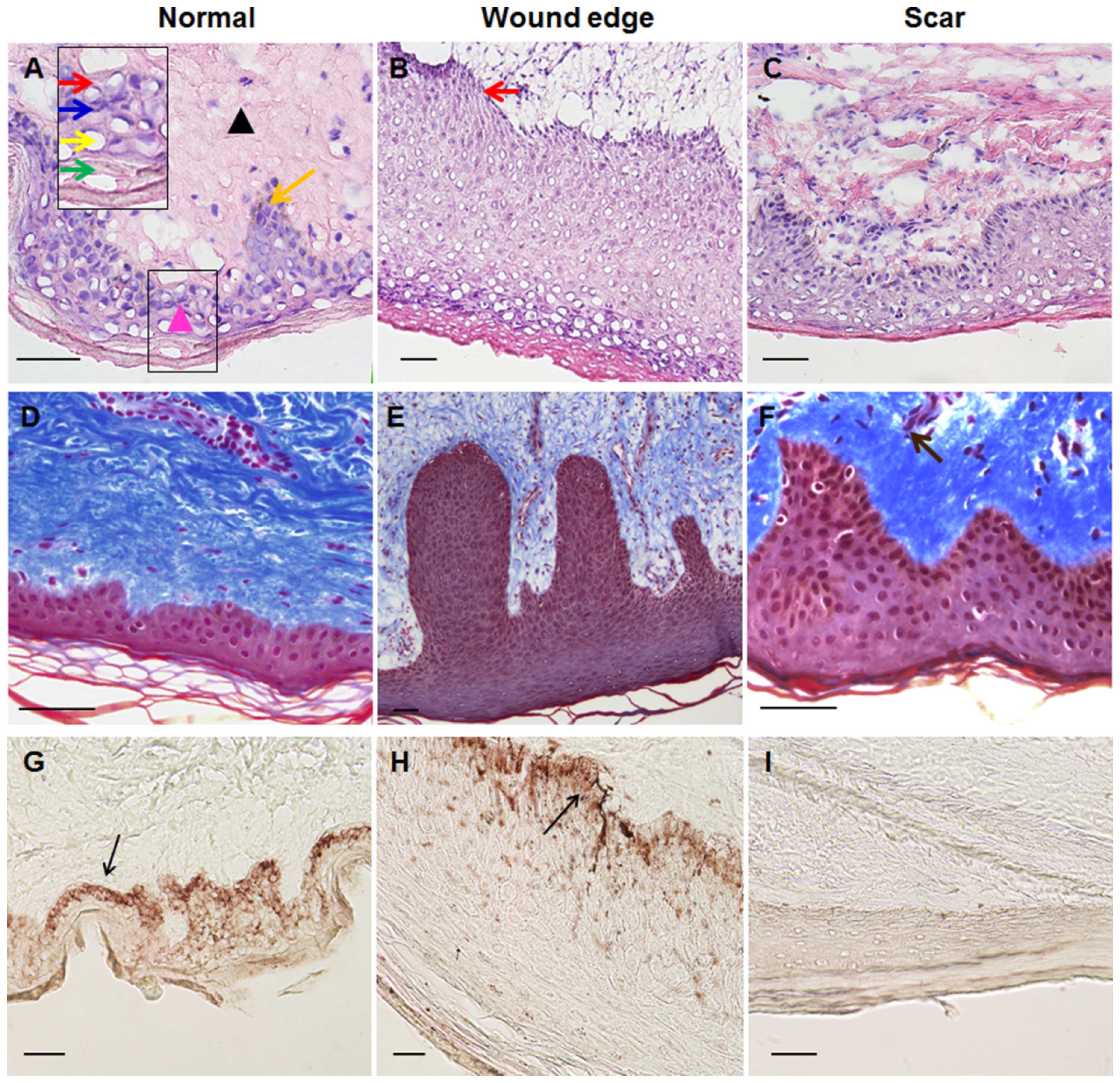 | Figure 1Morphological differences between
normal, wound edge and hypertrophic scar tissues. (A–C) Hematoxylin
and eosin (H&E) staining was performed to visualize the
histological structures in normal, wound edge and scar tissues. (A)
In the normal skin sample, the histological structure of the skin
was visible, showing layers of the epidermis and dermis. The
epidermis contained 4 to 5 layers of keratinocytes. In the
underlying dermis, fibroblasts made up loose connective tissue,
which was rich in parallel collagenous fibers (D). Inset shows a
close-up view of area within the black markings. Black triangle
denotes dermis, while pink triangle denotes epidermis. Orange arrow
denotes rete ridge. Red arrow denotes basal keratinocytes. Blue
arrow denotes spinous cells. Yellow arrow denotes granular cells.
Green arrows denote cornified cells. (B) In the wound edge tissue
sample, the epidermis proliferated and was of increased thickness
and was comprised of multiple layers of keratinocytes of distinct
morphology. Red arrow denotes basal keratinocytes, which arranged
as a regimented single layer of cuboidal or low columnar cells. (C)
The histological structure of hypertrophic scar tissue differed
from normal skin by having a rich blood supply, high mesenchymal
density, and a thick epidermal layer. The dermis was composed
largely of dense, disorganized connective tissue that includes
tough collagenous fibers of an irregular shape and a larger
diameter. (D–F) Masson's staining was performed to visualize the
distribution of collagen fibers in (D) normal, (E) wound edge and
(F) scar tissues. Brown arrow denotes tough collagens with rich
blood supply. (G–I) Methenamine silver staining was performed to
detect the basement membrane (BM) structure in (G) normal, (H)
wound edge and (I) scar tissues. BM was present in normal and wound
edge tissue, but absent in scar tissue. (G and H) Black arrows
denote the BM. All scale bars represent 50 μm. |
Differences in the expression of cell
markers in epidermal keratinocytes in normal, wound edge and
hypertrophic scar tissues
Given the morphological differences which were
observed in the normal, wound edge and hypertrophic scar tissues,
we examined whether the basal keratinocytes exhibited a different
cellular behavior during scar formation. Initially, we performed
immunofluorescence staining to detect the expression of CK10, CK14,
CK5, CK19 and integrin-β1 in the sections of normal, wound edge and
scar tissues. Previous studies have verified that CK10 is a marker
of differentiated keratinocytes, that CK14 and CK5 are markers of
proliferating basal keratinocytes or TAs, and that CK19 is a
putative marker of epidermal progenitor cells or stem cells in the
skin (14–17). Integrin-β1 is also thought to be a
marker of precursor cells in the skin (18,19). Our results indicated that CK10 was
expressed in the outer layer of the epidermis in the normal and
wound edge tissues (Fig. 2A and
B), and in the suprabasal, terminally differentiating cells in
the epidermis of the hypertrophic scar tissue (Fig. 2C). CK14 was expressed in both
basal and suprabasal layers in the stratified epidermis of all
three tissues (Fig. 2D–F), and
there was an extensive distribution of CK14 in the multilayered
epidermis of the scar tissue (Fig.
2F). The distribution pattern of CK5 was similar to that of
CK14 (Fig. 2G–I), which indicated
an increased number of proliferating cells in the
hyperproliferative epidermis. Integrin-β1 and CK19 were expressed
in the basal layer of the normal epidermis (Fig. 2J and M) and in the basal and
supra-basal layers of the wound edge epidermis (Fig. 2K and N), but were not detectable
in the epidermis of the hypertrophic scar tissues (Fig. 2L and O). Collectively, these
observations indicate that the keratin expression profile of basal
keratinocytes differed between the normal, wound edge and
hypertrophic scar tissues. Although fewer CK19-expressing cells
were detected in the basal and suprabasal layers of the
hypertrophic scar tissue (Fig.
2P), a proliferative phenotype was revealed due to the
increased expression of CK14 and CK5 in the multilayered
epidermis.
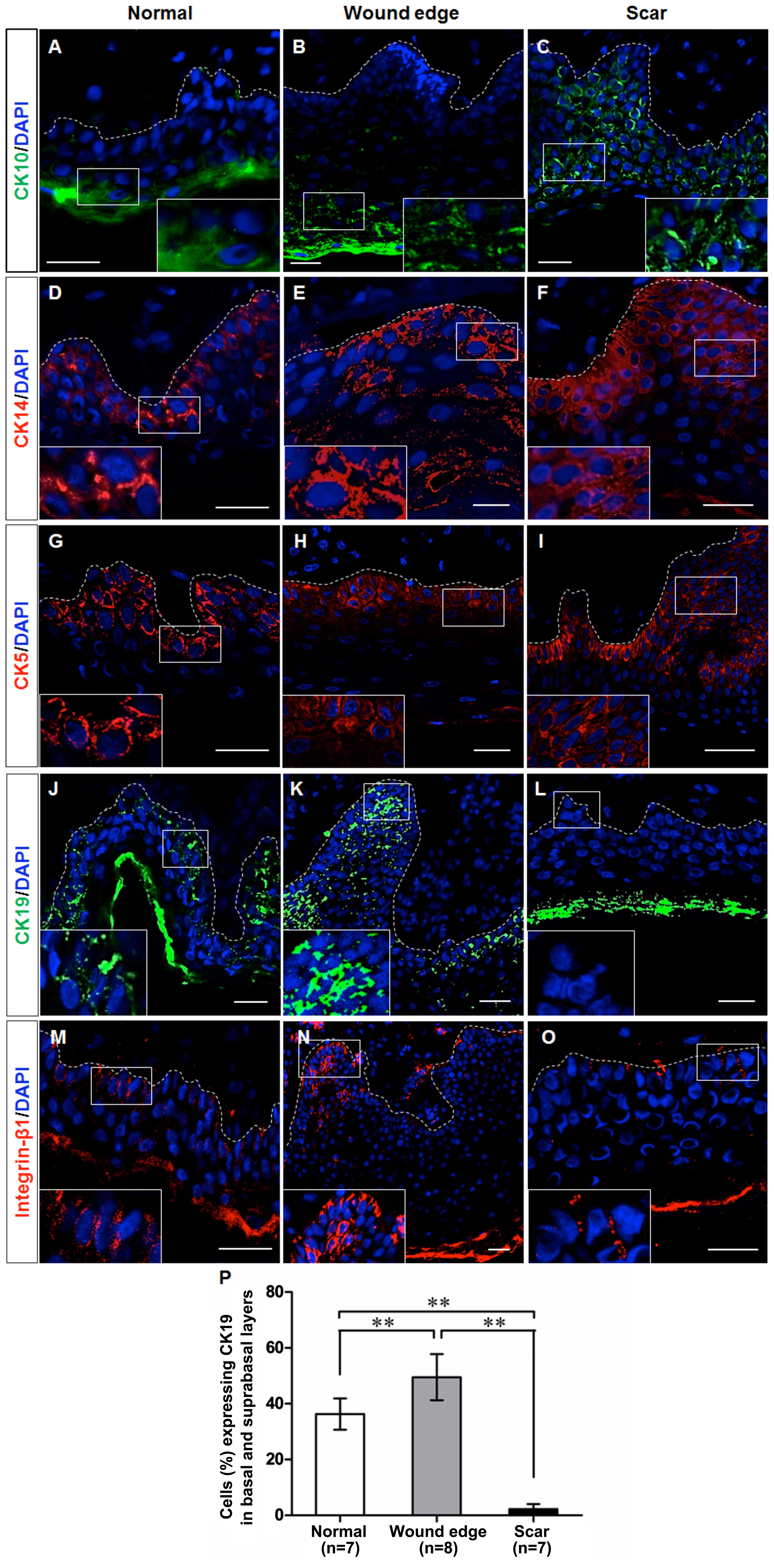 | Figure 2Cell marker expression in epidermal
keratinocytes in normal, wound edge and hypertrophic scar tissue.
Representative fluorescence images showing the localization of
CK10, CK14, CK5, CK19 and integrin-β1 in the sections of normal,
wound edge and scar tissues. (A–C) CK10, a marker of differentiated
keratinocytes, was expressed in the outer layer of the epidermis of
(A) normal and (B) wound edge tissues, and in (C) the suprabasal,
terminally differentiating cells in the epidermis of hypertrophic
scar tissues. (D–F) CK14 was expressed in both basal and suprabasal
layers in the stratified epidermis of all three tissues. In
particular, extensive distribution of CK14 was detectable in (F)
the multilayered epidermis of the scar tissue. (G–I) The
distribution patterns of CK5 were similar to those of CK14, which
indicated an increased number of proliferating cells in the
hyperproliferative epidermis. (J–O) CK19 and integrin-β1 were
expressed in the basal layer of (J and M) normal epidermis, and in
the basal and suprabasal layers of (K and N) wound edge epidermis,
but were not detected in (L and O) the epidermis of hypertrophic
scar tissues. White dotted lines denote the basement membrane,
which separated the epidermis and dermis. Insets show a close-up
view of area within the white markings. All scale bars represent 25
μm. (P) Statistical analysis of CK19-expressing cells in the
basal and suprabasal layers in normal, wound edge and hypertrophic
scar tissues. Data are the means ± SD. **P<0.01. |
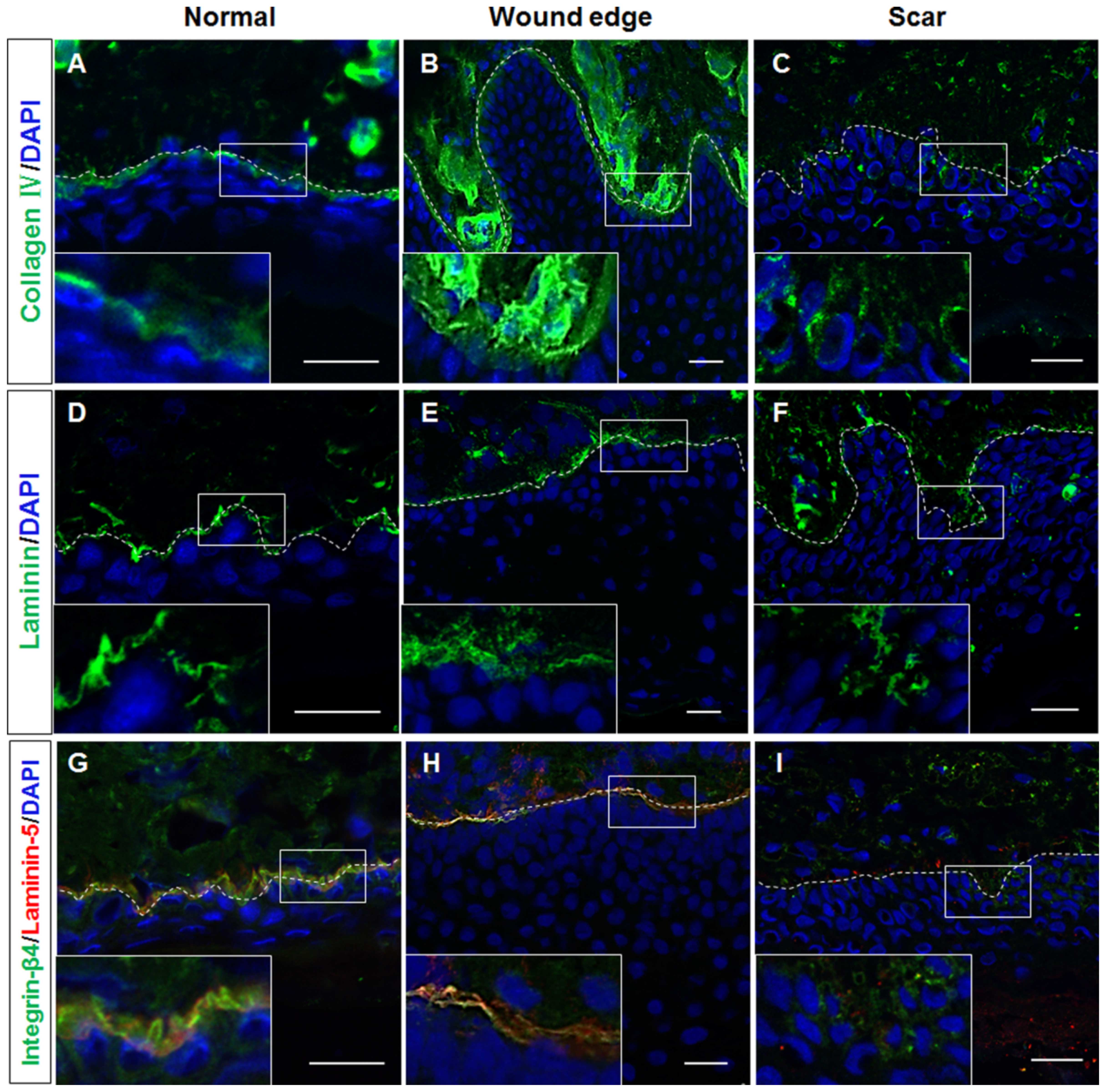
Altered BM structure contributes to the
differential keratin expression profile in epidermal keratinocytes
in vivo
As a key component of the stem cell niche, the ECM
not only anchors stem cells but also directs their fate (20). As mentioned above, the
histological structure of the BM appeared to be absent in the
hypertrophic scar tissue samples. Given the differences in the
keratin expression profiles in the basal layer of normal, wound
edge and hypertrophic scar epidermis, we then speculated that the
structural abnormalities of the BM play a role in regulating the
cell fate decision of basal keratinocytes during wound healing. To
examine this hypothesis, immunofluorescence staining was performed
to visualize the components of the BM in the normal, wound edge and
hypertrophic scar tissue samples. Our results revealed that
collagen IV expression was detected in the BM area in both normal
(Fig. 3A) and wound edge
(Fig. 3B) tissues. However, the
formation of BM-like structures with an absence of collagen IV
expression was observed in the epidermis of hypertrophic scar
tissues (Fig. 3C). Although the
expression of laminin was detected in all three tissue types
(Fig. 3D–F), the double-labelling
of laminin-5 and its receptor integrin-β4 further verified our
observation that the structure of the BM was altered in the
epidermis of the hypertrophic scar tissues; negative staining of
both laminin-5 and integrin-β4 (Figs.
3I and 4G–I) was observed as
compared with that in the normal (Figs. 3G and 4A–C) and wound edge tissues (Figs. 3H and 4D–F). Given these findings, it is likely
that the normal BM contributed to the recruitment of integrin-β1-
and CK19-expressing cells in the basal layer of the normal and
wound edge epidermis. Abnormalities in the BM structure, however,
induced the basal keratinocytes to differentiate and adopt a
proliferative phenotype during scar pathogenesis.
Altered BM structure promotes epidermal
keratinocytes to adopt a proliferative phenotype in vitro
The mechanical support provided by the BM is
determined primarily by its type IV collagen scaffold (21), into which the laminin networks and
perlecan oligomers are integrated by the cross-linking nidogens to
finally assemble the sheet-like BM complex (22). To further clarify the roles of the
BM in the regulation of epidermal keratinocyte behavior, we used
the human BM extract-derived ECM to coat the culture dishes in
order to control the pattern of cell-matrix adhesion. As shown in
Fig. 5, ECM administration
resulted in the upregulation of integrin-β4 expression in the HEKs
(Fig. 5A) and the HaCaT (Fig. 5B) cells, which was present on the
plasma membrane and at the sites of cell-matrix attachment. We also
performed western blot analysis to examine the protein levels of
integrin-β4 in the epidermal cell lines with or without ECM
coating. Our data demonstratd that higher levels of integrin-β4
were detectable in the ECM-treated groups than in the controls,
which further confirmed the fluorescent immunostaining findings
(Fig. 5C). Moreover, the HEKs and
HaCaT cells were switched to medium containing 1.5 mM
Ca2+ for the indicated periods of time to generate
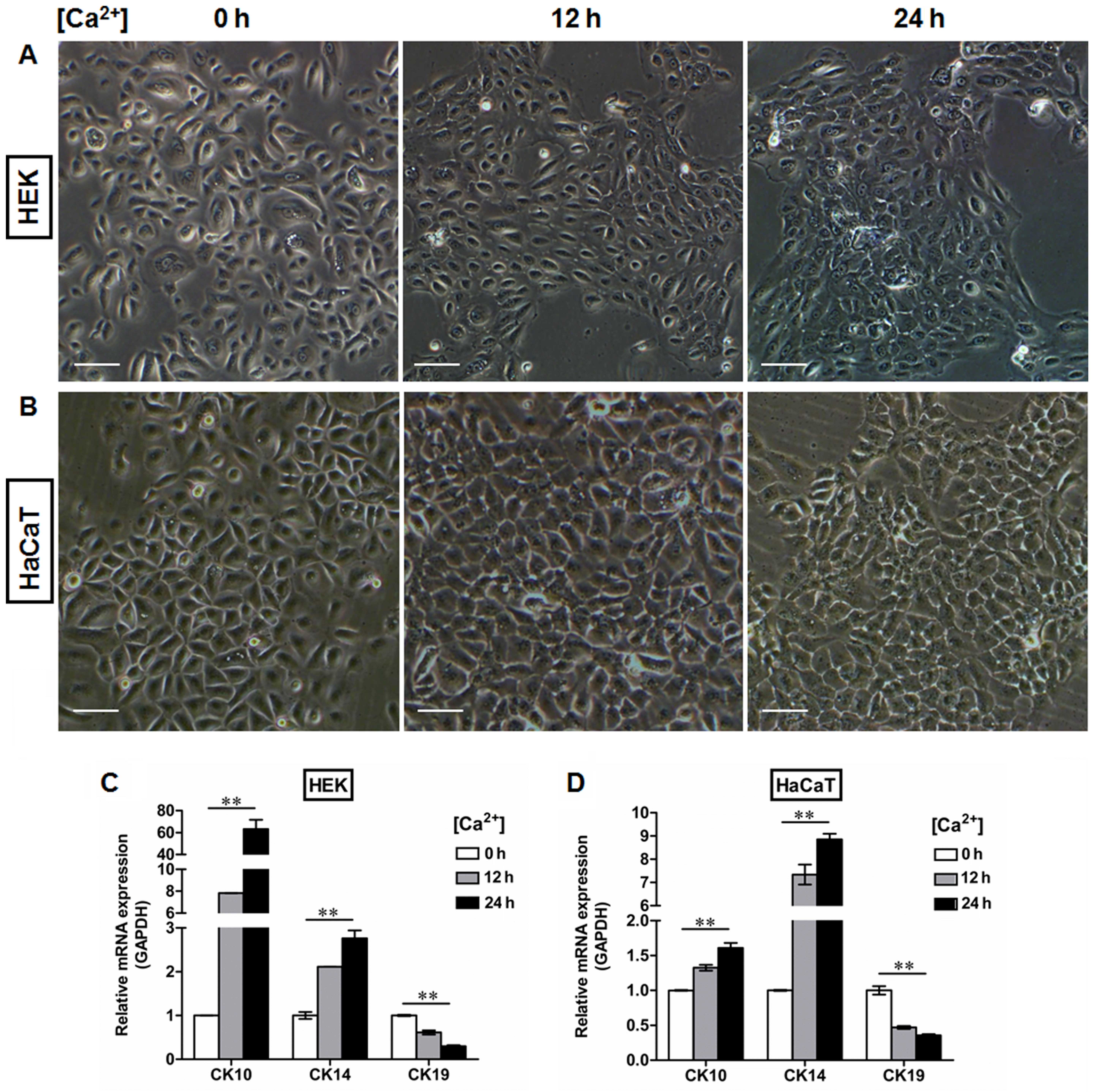
differentiating epidermal cells as previously described (23,24). Following Ca2+
treatment, the morphological alterations in both the HEKs (Fig. 6A) and HaCaT cells (Fig. 6B) were observed under a light
microscope, which included cell enlargement and flattening.
Ca2+ induced the expression of CK10 (Figs. 5D, G and J, and 6C and D) and CK14 (Figs. 5E, H and J, and 6C and D) in a time-dependent manner in
epidermal cell lines at both the mRNA (Figs. 5D, E, G and H, and 6C and D) and protein (Fig. 5J) levels, with peak levels of CK10
and CK14 expression being observed 24 h after initiating 1.5 mM
Ca2+ treatment in the HEKs (Figs. 5D, E and J and 6C) and the HaCaT cells (Figs. 5G, H and J and 6D). Following Ca2+ treatment,
the expression of CK19 was downregulated in the HEKs, reaching its
lowest level at 24 h after the start of Ca2+ treatment
(Figs. 5F and J and 6C). Similar results were observed in the
HaCaT cells (Figs. 5I and J and
6D). ECM treatment, however,
reduced the differentiating responses of keratinocytes associated
with Ca2+ administration in the HEKs and HaCaT cells,
and enhanced the expression of CK19 at 12 h and 24 h after
Ca2+ treatment (Fig. 5F, I
and J). To further determine the potential role of the BM in
the regulation of epidermal keratinocyte behavior, type IV collagen
was used to form a BM-like structure in vitro. Collagen IV
treatment contributed to CK19 expression and reduced the expression
of CK10 and CK14 at the mRNA and protein levels in both the HEKs
(Fig. 7A–C and G) and the HaCaT
cells (Fig. 7D–F and G). These
results further validated our in vivo findings that the BM
appears to be a positive regulator of the recruitment of
precursor-like cells in the basal layer of the epidermis.
Alterations in the BM structure promoted the development of a
proliferative phenotype in basal keratinocytes as indicated by the
enhanced expression of the markers associated with cell
proliferation, e.g., CK14 and CK5.
 | Figure 5Extracellular matrix (ECM) coating
regulates the expression of cytokeratin (CK)10, CK14 and CK19 in
epidermal keratinocytes in vitro. Human basement membrane
(BM) extract-derived ECM was used to minic the BM structure in
vitro. (A and B) Representative fluorescence images showing the
localization of integrin-β4 (green) in (A) human epidermal
keratinocytes (HEKs) and (B) human immortalized keratinocyte
(HaCaT) cells with or without ECM coating. Nuclei were stained with
DAPI (blue). All scale bars represent 25 μm. (C) Western
blot analysis of integrin-β4 expression in HEKs and HaCaT cells
with or without ECM coating. (D–I) After primary cultured HEKs and
HaCaT cells were switched to the medium containing 1.5 mM
Ca2+ for the indicated time periods, the relative levels
of CK10, CK14, and CK19 expression were assessed by real-time qPCR.
Ca2+ induced the expression of (D and G) CK10 and (E and
H) CK14 in a time-dependent manner in the epidermal cell lines,
with a peak expression level 24 h after 1.5 mM Ca2+
treatment in (D and E) HEKs and (G and H) HaCaT cells. ECM
treatment, however, reduced the differentiating responses of
keratinocytes associated with Ca2+ administration in (F)
HEKs and (I) HaCaT cells, and enhanced expression of CK19 at 12 h
and 24 h after Ca2+ addition. Data are the means ± SD.
*P<0.05, **P<0.01, ns= not significant.
(J) Western blot analysis of CK10, CK14, and CK19 expression in
HEKs and HaCaT cells with or without ECM coating. |
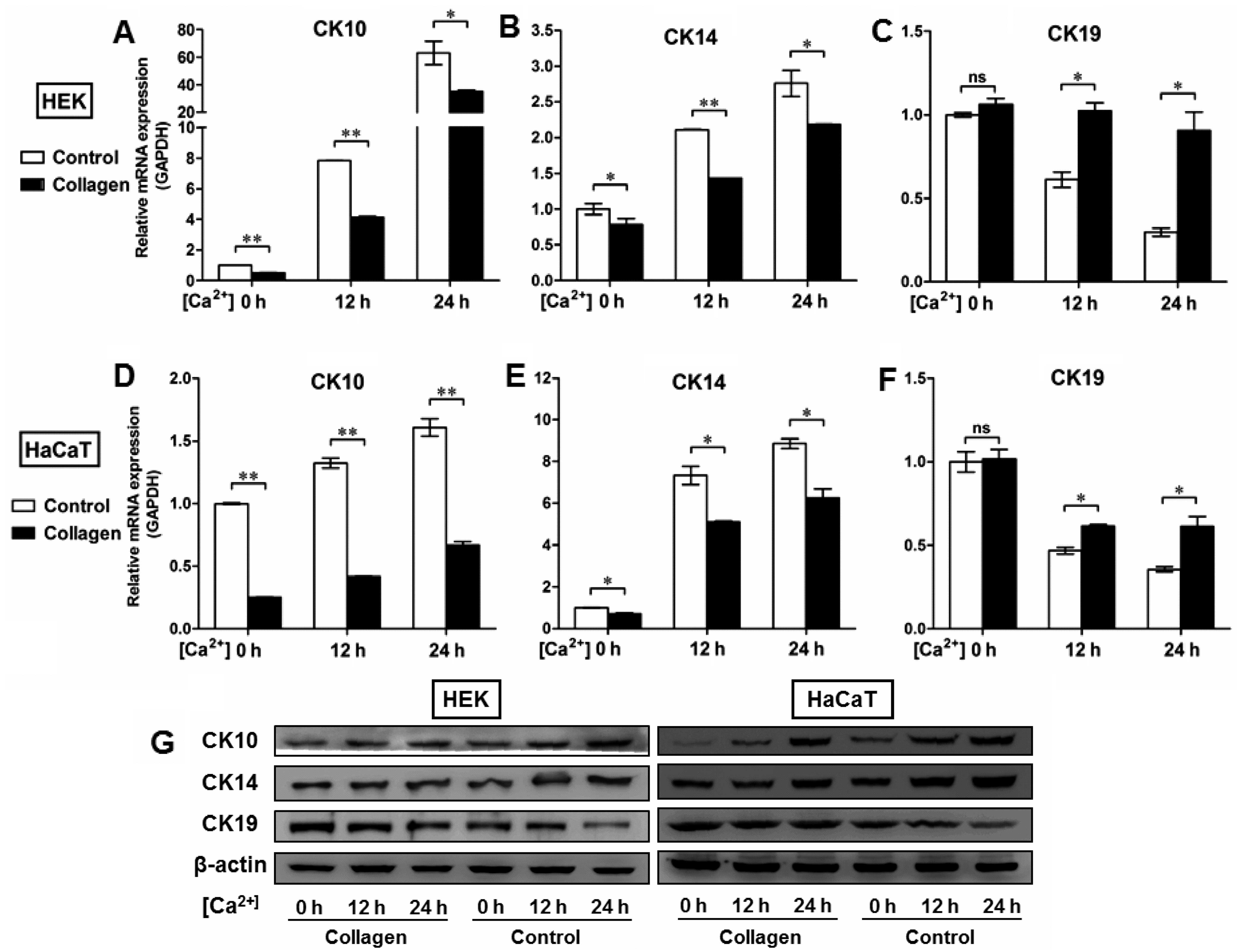 | Figure 7Type IV collagen coating regulates
the expression of CK10, CK14 and CK19 in epidermal keratinocytes
in vitro. (A–F) To further determine the potential role of
basement membrane (BM) in the regulation of epidermal keratinocyte
behavior, type IV collagen was used to form a BM-like structure
in vitro. After human epidermal keratinocytes (HEKs) and
human immortalized keratinocyte (HaCaT) cells were switched to the
medium containing 1.5 mM Ca2+, the relative levels of
CK10, CK14, and CK19 expression were assessed by RT-qPCR. Although
Ca2+ contributed to the expression of (A and D) CK10 and
(B and E) CK14 in a time-dependent manner in epidermal cell lines,
type IV collagen treatment reduced the mRNA expression of CK10 and
CK14 in both (A and B) HEKs and (D and E) HaCaT cells. Type IV
collagen treatment also increased the mRNA levels of CK19 in (C)
HEKs and (F) HaCaT cells in vitro. Data are the means ± SD.
*P<0.05, **P<0.01, ns, not significant.
(G) Western blot analysis of CK10, CK14, and CK19 expression in
HEKs and HaCaT cells with or without collagen IV treatment. |
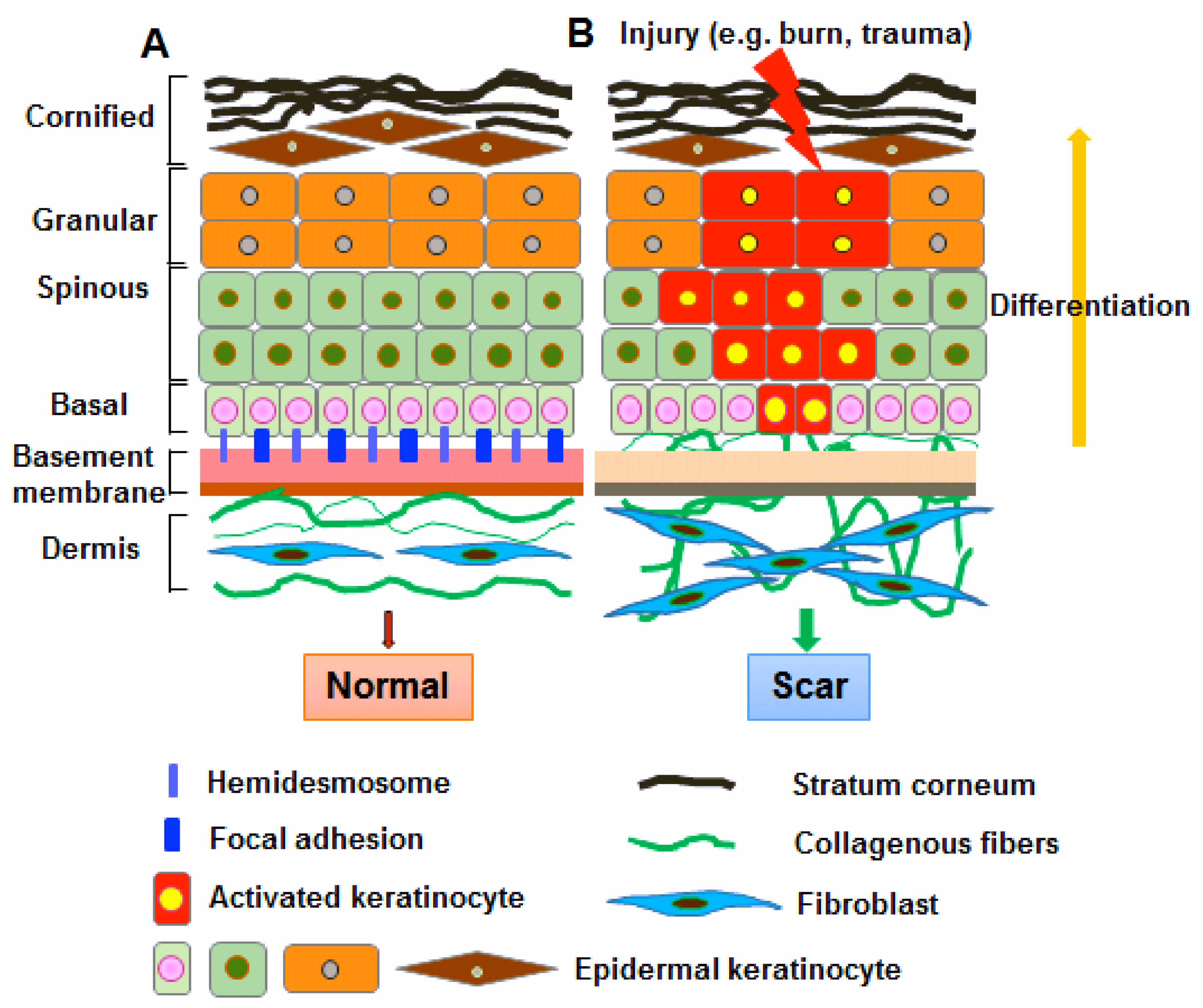
Discussion
In the present study, we first investigated the
morphological changes which occur during epidermal regeneration as
part of the wound healing process. Our data demonstrated that the
histological structure of the hypertrophic scar tissues differed
from that of normal skin tissue, with a significant increase in
epidermal thickness between the basal layer and stratum corneum
being observed. Notably, staining of the BM appeared to be absent
in the scar tissue. Moreover, immunofluorescence staining for CK10,
CK14, CK5, CK19 and integrin-β1 indicated that the differential
expression of basal keratinocyte markers differed among the samples
of normal, wound edge and hypertrophic scar tissues, and
furthermore the epidermis of the hypertrophic scar tissue exhibited
a proliferative phenotype by switching the expression of
integrin-β1 and CK19 to CK14 and CK5, markers of cell
proliferation, in the multilayered epidermis. Given these findings,
we thus hypothesized that the BM plays a role in regulating the
cell fate decision of epidermal keratinocytes during wound healing.
By using a panel of antibodies associated with BM components, we
validated our hypothesis that the structure of the BM was altered
in the hypertrophic scar tissues. Our results indicated that the BM
contributed to the recruitment of CK19-expressing cells in the
basal layer of normal and wound edge epidermis, whereas
abnormalities in the structure of the BM induced the basal
keratinocytes to differentiate and adopt a proliferative phenotype
during scar pathogenesis. By using ECM and collagen IV to mimic the
BM structure in vitro, we further confirmed our in
vivo observations and showed that the BM reduced the
differentiating responses of keratinocytes induced by
Ca2+ administration in the epidermal cell lines and
enhanced the expression of CK19, a putative marker of epidermal
progenitor cells.
The BM plays a fundamental role in the
differentiation, proliferation, survival and migration of cells
during embryonic development. In the skin, the BM physically
separates the epidermis from the underlying dermis. The basal
keratinocytes in the epidermis attach to the underlying BM through
integrins, which bind to some BM components, such as collagen,
laminin and fibronectin (25).
Thus, the BM not only serves as a selective barrier and structural
scaffold, but also provides a communication interface between
dermal fibroblasts and epidermal keratinocytes (2,26).
The BM also binds to a number of cytokines and growth factors,
serving as a reservoir for their controlled release, which
regulates keratinocyte-fibroblast interactions to promote the
development of skin, as well as wound healing (27,28). Previous studies on scar formation
have mainly focused on fibroblasts and little is known about the
role of the overlying epidermal keratinocytes (29,30). There is growing evidence that
keratinocytes may play an important role in the development of
pathological fibrosis through paracrine regulation of fibroblast
function (6,9,10).
It has also been shown that keratinocytes derived from scar tissues
differed from normal keratinocytes by exhibiting differential gene
expression profiles (12). The
mechanisms responsible for the fundamental abnormalities in
keratinocytes during keloid and hypertrophic scarring remain
elusive. Our data indicated a potential link between BM remodeling
and the maintenance of keratinocyte function during wound repair
and regeneration. It is well known that adhesion molecules, such
integrins and E- and N-cadherin, anchor stem cells to the ECM
(22). In light of our
observations, it appears that the BM may represent part of the
niche components for the recruitment of epidermal progenitor cells
in the basal layer of the skin epidermis. Our results indicate that
the abnormal functioning of the BM reduces the attachment of basal
progenitor cells to their surrounding microenvironment, and induces
them to adopt a proliferative phenotype, which may account for the
pathogenesis of scarring (Fig.
8).
Acknowledgments
This study was supported in part by grants from the
Novel Program of Beijing (nos. 2008B53 and 2009A38) and the
National Natural Science Foundation of China (nos. 30901564,
81101883, 81372067, 81121004 and 81230041) and the National Basic
Science and Development Program (973 Program, 2012CB518105).
References
|
1
|
Fuchs E and Raghavan S: Getting under the
skin of epidermal morphogenesis. Nat Rev Genet. 3:199–209. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Breitkreutz D, Mirancea N and Nischt R:
Basement membranes in skin: unique matrix structures with diverse
functions? Histochem Cell Biol. 132:1–10. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Blanpain C and Fuchs E: Epidermal stem
cells of the skin. Annu Rev Cell Dev Biol. 22:339–373. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Snippert HJ, Haegebarth A, Kasper M, Jaks
V, van Es JH, Barker N, van de Wetering M, van den Born M, Begthel
H, Vries RG, et al: Lgr6 marks stem cells in the hair follicle that
generate all cell lineages of the skin. Science. 327:1385–1389.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Broughton G II, Janis JE and Attinger CE:
The basic science of wound healing. Plast Reconstr Surg. 117(Suppl
7): 12S–34S. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Bellemare J, Roberge CJ, Bergeron D,
Lopez-Vallé CA, Roy M and Moulin VJ: Epidermis promotes dermal
fibrosis: role in the pathogenesis of hypertrophic scars. J Pathol.
206:1–8. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Andriessen MP, Niessen FB, Van de Kerkhof
PC and Schalkwijk J: Hypertrophic scarring is associated with
epidermal abnormalities: an immunohistochemical study. J Pathol.
186:192–200. 1998. View Article : Google Scholar
|
|
8
|
Teepe RG, Kreis RW, Koebrugge EJ,
Kempenaar JA, Vloemans AF, Hermans RP, Boxma H, Dokter J, Hermans
J, Ponec M, et al: The use of cultured autologous epidermis in the
treatment of extensive burn wounds. J Trauma. 30:269–275. 1990.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Funayama E, Chodon T, Oyama A and Sugihara
T: Keratinocytes promote proliferation and inhibit apoptosis of the
underlying fibroblasts: an important role in the pathogenesis of
keloid. J Invest Dermatol. 121:1326–1331. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Lim IJ, Phan TT, Bay BH, Qi R, Huynh H,
Tan WT, Lee ST and Longaker MT: Fibroblasts cocultured with keloid
keratinocytes: normal fibroblasts secrete collagen in a keloidlike
manner. Am J Physiol Cell Physiol. 283:C212–C222. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Hahn JM, Glaser K, McFarland KL, Aronow
BJ, Boyce ST and Supp DM: Keloid-derived keratinocytes exhibit an
abnormal gene expression profile consistent with a distinct causal
role in keloid pathology. Wound Repair Regen. 21:530–544. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Machesney M, Tidman N, Waseem A, Kirby L
and Leigh I: Activated keratinocytes in the epidermis of
hypertrophic scars. Am J Pathol. 152:1133–1141. 1998.PubMed/NCBI
|
|
13
|
Orazizadeh M, Hashemitabar M, Bahramzadeh
S, Dehbashi FN and Saremy S: Comparison of the enzymatic and
explant methods for the culture of keratinocytes isolated from
human foreskin. Biomed Rep. 3:304–308. 2015.PubMed/NCBI
|
|
14
|
Su L, Morgan PR and Lane EB: Keratin 14
and 19 expression in normal, dysplastic and malignant oral
epithelia. A study using in situ hybridization and
immunohistochemistry. J Oral Pathol Med. 25:293–301. 1996.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Michel M, Török N, Godbout MJ, Lussier M,
Gaudreau P, Royal A and Germain L: Keratin 19 as a biochemical
marker of skin stem cells in vivo and in vitro: keratin 19
expressing cells are differentially localized in function of
anatomic sites, and their number varies with donor age and culture
stage. J Cell Sci. 109:1017–1028. 1996.PubMed/NCBI
|
|
16
|
Khanom R, Sakamoto K, Pal SK, Shimada Y,
Morita K, Omura K, Miki Y and Yamaguchi A: Expression of basal cell
keratin 15 and keratin 19 in oral squamous neoplasms represents
diverse pathophysiologies. Histol Histopathol. 27:949–959.
2012.PubMed/NCBI
|
|
17
|
Fuchs E: Skin stem cells: rising to the
surface. J Cell Biol. 180:273–284. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Yeh YC, Lin HH and Tang MJ: A tale of two
collagen receptors, integrin β1 and discoidin domain receptor 1, in
epithelial cell differentiation. Am J Physiol Cell Physiol.
303:C1207–C1217. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Borowska K, Jedrych B, Czerny K and
Zabielski S: The role of integrins in the physiologic and
pathogenic processes. Pol Merkur Lekarski. 21:362–366. 2006.Article
in Polish.
|
|
20
|
Lane SW, Williams DA and Watt FM:
Modulating the stem cell niche for tissue regeneration. Nat
Biotechnol. 32:795–803. 2014. View
Article : Google Scholar : PubMed/NCBI
|
|
21
|
Breitkreutz D, Koxholt I, Thiemann K and
Nischt R: Skin basement membrane: the foundation of epidermal
integrity - BM functions and diverse roles of bridging molecules
nidogen and perlecan. Biomed Res Int. 2013(179784)2013. View Article : Google Scholar
|
|
22
|
Timpl R and Brown JC: Supramolecular
assembly of basement membranes. BioEssays. 18:123–132. 1996.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Hennings H, Michael D, Cheng C, Steinert
P, Holbrook K and Yuspa SH: Calcium regulation of growth and
differentiation of mouse epidermal cells in culture. Cell.
19:245–254. 1980. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Hennings H and Holbrook KA: Calcium
regulation of cell-cell contact and differentiation of epidermal
cells in culture. An ultrastructural study. Exp Cell Res.
143:127–142. 1983. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Burgeson RE and Christiano AM: The
dermal-epidermal junction. Curr Opin Cell Biol. 9:651–658. 1997.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
LeBleu VS, Macdonald B and Kalluri R:
Structure and function of basement membranes. Exp Biol Med
(Maywood). 232:1121–1129. 2007. View Article : Google Scholar
|
|
27
|
Iozzo RV: Basement membrane proteoglycans:
from cellar to ceiling. Nat Rev Mol Cell Biol. 6:646–656. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Werner S, Krieg T and Smola H:
Keratinocyte-fibroblast interactions in wound healing. J Invest
Dermatol. 127:998–1008. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Moulin V, Castilloux G, Auger FA, Garrel
D, O'Connor-McCourt M and Germain L: Modulated response to
cytokines of human wound healing myofibroblasts compared to dermal
fibroblasts. Exp Cell Res. 238:283–293. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Gabbiani G, Ryan GB and Majne G: Presence
of modified fibroblasts in granulation tissue and their possible
role in wound contraction. Experientia. 27:549–550. 1971.
View Article : Google Scholar : PubMed/NCBI
|