Introduction
Zinc (Zn) is an essential nutrient and one of the
most abundant biological trace metals which is intimately involved
in growth and reproduction in males and females (1,2).
It is well known that Zn content is high in the adult testis, and
the prostate has a higher concentration of Zn than any other organ
of the body. Zn plays pivotal roles in testicular development, the
maintenance of testicular function and spermatogenesis (3,4).
In humans, for example, Zn is a major constituent of seminal plasma
and it is necessary for the formation and maturation of spermatozoa
as well as for fertilization (5,6).
Epidemiological investigations in humans have demonstrated that the
testes have a strict Zn requirement and a severe Zn deficiency may
lead to low seminal plasma Zn levels, and the subsequent depletion
of testosterone as well as the inhibition of spermatogenesis. This
has effects on the fertility of a population, and in one study Zn
deficiency was found to be the main cause of a decline in fertility
rates (5,7,8).
In addition, Zn deficiency has been demonstrated to cause Leydig
cells to appear smaller and show endoplasmic reticulum
abnormalities, when examined under an electron microscope (9). Notably, Zn-deficient (ZnD) Leydig
cells are capable of taking up cholesterol and neutral lipids,
which are the precursors of sex steroids; however, they are
incapable of converting them into sex steroids. This has been shown
to lead to the arrest of spermatogenesis and the impairment of
fertilization (10). Indeed, Zn
deficiency has been shown to result in oligospermia, which
coincides with poor Leydig cell function and lower testosterone
concentrations, and these effects may be reversed with Zn
supplementation (11,12). Thus, Zn, which is known to be an
important cation necessary for the normal functioning of the testes
and the regulation of fertility, is now attracting increasing
attention from researchers.
Previous studies have confirmed that changes in
intracellular Zn concentrations exerted marked effects on
testicular steroido-genesis processes induced by various physical,
chemical and immunological stimuli (11,13,14). Zn homeostasis is regulated by two
different Zn-transporter families: the Zrt- and Irt-like proteins
(ZIPs; SLC39a) and the Zn transporter (ZnT) proteins (SLC30a). ZIPs
facilitate the influx of Zn into the cytosol from the outside of
cells or from the lumen of intracellular compartments whereas ZnTs
ensure Zn efflux from the cytosol to the outside of cells or to
intracellular organelles (15–17). ZnT1 is ubiquitously expressed and
the only ZnT known to be involved in Zn efflux across the plasma
membrane in many different cells, thus conferring resistance to Zn
(15). ZnT3 is mainly located in
the presynaptic vesicles of neurons and is responsible for the
transportation of Zn into synaptic vesicles involved in nerve
transmission (18,19). ZnT5 is thought to transport Zn
into the insulin-containing secretory granules of pancreatic
β-cells (20). ZnT7 is located in
the Golgi apparatus and it plays an important role in regulating
the efflux of Zn from the cytosol into the extracellular space or
increasing the influx of Zn into the Golgi network (14,15,21). ZnT8 is predominantly expressed in
the insulin-containing granules in pancreatic β-cells and
transports cytoplasmic Zn ions into the secretory granules for
insulin maturation and crystallization prior to secretion (22,23). Previous findings have revealed
that coordinated Zn mobilization by ZnTs is essential for the
maintenance of Zn homeostasis. Thus, Zn plays roles in the
regulation of critical physiological functions and profoundly
affects health positively or negatively, as it is involved in a
wide variety of diseases (17,24). The study of ZnTs is therefore
receiving great clinical interest and attention at present
(17), although little is known
about the functional characteristics of ZnTs, particularly ZnT7, in
endocrine tissues such as the mammalian testis.
In the present study, we aimed to examine the role
of ZnT7 in testosterone synthesis and to elucidate the underlying
mechanisms. Several important transport proteins and enzymes
involved in testosterone synthesis were selected for evaluation,
including steroidogenic acute regulatory protein (StAR),
cholesterol side-chain cleavage enzyme (P450scc) and
3β-hydroxysteroid dehydrogenase/D5-D4 isomerase (3β-HSD). Our
results revealed that double immunofluorescence staining between
ZnT7 and StAR was mainly observed in the interstitial compartment
cells in the mouse testis, and the intensity of staining was weaker
in the ZnD group. Silencing the ZnT7 gene in the mouse Leydig tumor
cell line (MLTC-1) cells decreased the levels of StAR, P450scc and
3β-HSD, and subsequently decreased progesterone synthesis in the
MLTC-1 cells. These data suggest that ZnT7 may be involved in
steroidogenesis in the mouse testis.
Materials and methods
Animals
Four-week-old male CD-1 mice, weighing approximately
13±2 g, were purchased from the Experimental Animal Center of China
Medical University (Shenyang, China). The animals were divided into
two groups: 20 animals were fed a ZnD diet (0.85 ppm; egg
white-based AIN-76A; Research Diets Inc., New Brunswick, NJ, USA)
and provided with deionized water; the other 20 animals were
treated with a Zn-adequate (30 ppm) diet and received normal
drinking water ad libitum. The mice were housed in cages in
a controlled environment (22–25°C; 50% humidity; and 12-h/12-h
light/dark cycle). The mice were fed either ZnD or Zn-adequate
diets for 5 weeks. All animal experimental procedures were approved
by the Experimental Animal Ethics Committee of China Medical
University (Shenyang, China), in accordance with the criteria
described in the National Institutes of Health (NIH) Guide for the
Care and Use of Laboratory Animals.
Analysis of Zn levels
Following decapitation, the testes of five mice from
each group were carefully removed, and the samples were digested
with concentrated nitric acid; the digests were diluted to a
constant volume with deionized water. The Zn concentration was then
determined from the resultant solution by performing atomic
absorption spectrophotometry (AAS) at the Experimental Center of
China Medical University (Shenyang, China). The results are
expressed as mg/g of wet weight (wet wt) and as averages of at
least two determinations/sample.
Double immunofluorescence staining
Four animals from the ZnD group and four from the
control group were deeply anesthetized with sodium pentobarbital
and perfused with isotonic saline, followed by 4% paraformaldehyde
in 0.1 M phosphate-buffered saline (PBS; pH 7.4). The testes were
removed and post-fixed with 4% paraformaldehyde overnight at 4°C.
Serial paraffin sections (5 μm) were prepared, dewaxed in
xylene and rehydrated using gradient alcohol solutions. The
cryostat sections were then pre-incubated with normal donkey serum
(1:20) for 1 h and incubated overnight at room temperature (RT)
with anti-ZnT7 (polyclonal antibody; HPA018034; Sigma, St. Louis,
MO, USA) at 1:100 and anti-StAR (polyclonal antibody; sc-25806;
Santa Cruz Biotechnology, Inc., Santa Cruz, CA, USA) at 1:100.
Secondary antibodies including fluorescein isothiocyanate
(FITC)-conjugated donkey anti-rabbit (FITC-DAR; 711-542-152) IgG
and Texas Red-conjugated donkey anti-goat (Texas Red-DAG;
705-007-003) IgG were purchased from Jackson ImmunoResearch
Laboratories, Inc. (West Grove, PA, USA). After rinsing with PBS,
the sections were incubated for 2 h with FITC-DAR (1:50) and Texas
Red-DAG (1:50) at RT. The sections were mounted and examined under
a confocal laser scanning microscope (CLSM; SP2; Leica Microsystems
GmbH, Wetzlar, Germany).
Cell culture and treatment with a Zn ion
chelating agent
The MLTC-1 cell line was purchased from the Cell
Bank, Shanghai Institutes for Biological Sciences, Chinese Academy
of Sciences (Shanghai, China) and the cells were maintained in an
RPMI-1640 medium (RPMI-1640; Gibco BRL, Grand Island, NY, USA)
supplemented with 10% fetal bovine serum (FBS), 100 IU/ml of
penicillin and 100 μg/ml of streptomycin, at 37°C in a 5%
CO2 incubator with humidified 95% air. To deplete the
intracellular Zn stores, the Zn chelator
N,N,N′,N′-tetrakis(2-pyridylmethyl)ethylenediamine (TPEN; 1
μM) was added and incubated with the cells for 12 h.
Assessment of cell viability
Cell viability was measured in 96-well plates by a
quantitative colorimetric assay using
3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT),
which is an indicator of mitochondrial activity in living cells.
Briefly, at the indicated time after treatment, 10 μl MTT
(final concentration, 500 μg/ml) was added to the medium and
the cells were incubated at 37°C for 3 h. The MTT solution was
removed and 100 μl dimethyl sulfoxide (DMSO) was added for
15 min in order to dissolve the resulting colored formazan
crystals. The absorbance at 540 nm for each sample aliquot was
measured using a Sunrise RC microplate reader (Tecan Group, Ltd.,
Männedorf, Switzerland). Cell viability was expressed as the ratio
of the signal obtained from the treated cultures to the
control.
Transient transfection with mZnT7
(NM_023214) small interfering (si)RNA (Stealth RNAi)
The cells were plated in 6-well plates at a density
of 2×105 cells/well in 2 ml RPMI-1640. ZnT7 siRNA
(Stealth RNAi) was prepared by Invitrogen Life Technologies
(Carlsbad, CA, USA). The siRNA sequence targeting a specific
sequence in ZnT7 mRNA used was 5′-GCCAUAGUCACGAAGCCAATT-3′. The
scrambled siRNA sequence used was 5′-UUGGCUUCGUGACUAUGGCTT-3′. The
target sequences of ZnT7 siRNA and control ZnT7 siRNA were aligned
against the GenBank database using the Basic Local Alignment Search
Tool (http://blast.ncbi.nlm.nih.gov/Blast.cgi). Transient
transfections were performed according to the manufacturer's
instructions using Lipofectamine® 2000 (Invitrogen Life
Technologies) in order to silence the expression of ZnT7. All
experiments were performed in 6-well plates, with the cells plated
to reach 50–60% confluence on the day of transfection. The cells
were incubated in growth medium with 10% FBS for 24 h after
transfection. The knockdown of ZnT7 was evaluated using western
blot analysis.
Hormone assays
Blood was collected by cardiac puncture with a
heparinized syringe and serum was immediately collected by
centrifugation (4,000 rpm at 4°C) and stored at −20°C for further
analysis. The serum sample concentrations of luteinizing hormone
(LH) and testosterone were measured using an enzyme-linked
immunosorbent assay as described in the instructions provided with
the assay kits (DRG Instruments GmbH, Marburg, Germany). For the
cell experiments, following transfection or treatment with TPEN
(Sigma), the supernatants were collected 2 h after human chorionic
gonadotrophin (hCG) stimulation. The total progesterone levels were
detected in the culture medium using the above-mentioned
method.
Western blot analysis
The decapsulated testes were placed in a 15-ml
conical tube containing 2–10 ml DMEM/Ham's F12 (DMEM/F12) and
subjected to rocking horizontal incubation at room temperature at
250 rpm for 40 min. Shortly after resting for 1 min, suspended
cells in the upper fraction were separated from the bottom
fraction. Upper fraction cells primarily consisting of Leydig cells
were centrifuged at 200 × g for 5 min and subjected to western blot
analysis. The Leydig cells isolated from the testes of 11 mice were
minced into small pieces and homogenized in a chilled lysis buffer
(150 mM of NaCl, 50 mM of Tris-HCl, 1% Nonidet P-40, 0.25% sodium
deoxycholate, 0.1% SDS, 1 mM of PMSF, 10 mg/ml of leupeptin, 1 mM
of Na3VO4, and 1 Mm of NaF) overnight at 4°C.
The MLTC-1 cell pellets were lysed in a chilled lysis buffer for an
hour at 4°C. The lysates were collected, centrifuged at 12,000 rpm
for 30 min, and quantified for total protein using a UV 1700
PharmaSpec ultraviolet spectrophotometer (Shimadzu, Kyoto, Japan).
The supernatant was removed, portioned into aliquots, and then
stored at −80°C.
Fifty micrograms of total protein from each sample
were subjected to SDS-PAGE using 10% gradient Tris/glycine gels.
The proteins were then transferred to polyvinylidene difluoride
(PVDF) membranes (Millipore Corp., Temecula, CA, USA). After
blocking with 5% fat-free milk for 1 h, the blots were incubated
with the following primary antibodies: anti-ZnT7 (1:1,000;
HPA018034; Sigma), anti-StAR (1:1,000; sc-25806; Santa Cruz
Biotechnology, Inc.), anti-P450SCC (1:1,000; ABS235; Millipore
Corp.), anti-3β-HSD (1:1,000; sc-28206; Santa Cruz Biotechnology,
Inc.), and anti-GAPDH (1:10,000; G8795; KangChen Biotech, Shanghai,
China) at 4°C overnight. The membranes were incubated with an
horseradish peroxide (HRP)-conjugated goat anti-rabbit IgG (H + L;
170-6515; Biocompare, San Francisco, CA, USA), HRP-conjugated goat
anti-mouse IgG (H + L; 170-6515; Biocompare), and HRP-conjugated
rabbit anti-goat IgG (H + L; ab6721; Abcam, Cambridge, UK)
(1:5,000) for 2 h at room temperature. Immunoreactive bands were
visualized using a SuperSignal West Pico Chemiluminescent substrate
(Pierce Biotechnology, Rockford, IL, USA) with a ChemDoc XRS
imaging system and Quantity One software (Bio-Rad, Hercules, CA,
USA).
Statistical analysis
Continuous variables are expressed as the means ±
standard error of the mean (SEM). One-way analysis of variance
(ANOVA) was used to analyze the data. Tukey's multiple comparison
test was used. P<0.05 was considered to indicate a statistically
significant difference.
Results
Colocalization of ZnT7 and StAR in the
Leydig cells of mouse testes
To examine the association between ZnT7 and
testosterone synthesis as well as testosterone-related factors in
Leydig cells, we first examined whether ZnT7 proteins are expressed
in the Leydig cells from the mouse testis. Testosterone is
essential for spermatogenesis, fertility and the maintenance of the
male phenotype (25). StAR
protein and testosterone biosynthetic enzymes play critical roles
in the synthesis of testosterone in Leydig cells (26–28). Double immunofluorescence staining
for ZnT7 and StAR was performed to analyze the distribution and
localization of both proteins in the mouse testis. Using a CLSM,
ZnT7-positive cells were found to be distributed in the
interstitial compartment and the seminiferous tubules of the mouse
testis. The intensity of ZnT7 staining in the interstitial
compartment cells was stronger than that in the seminiferous
tubule; StAR staining was exclusively located in the interstitial
compartment cells (Fig. 1). The
merged images illustrated that ZnT7 was colocalized with StAR in
the interstitial compartment cells in the mouse testis (Fig. 1). Furthermore, we found that the
staining for both ZnT7 and StAR was weaker in the Leydig cells from
the mice in the ZnD group compared with the control group. These
results demonstrated that ZnT7 was expressed in the interstitial
compartment cells of the mouse testes and may be involved in
steroidogenesis.
Effects of Zn deficiency on Zn
concentrations in mouse testes and serum levels of LH and
testosterone
In an in vivo experiment, AAS was used to
determine the concentration of Zn ions in the mouse testis. As
compared with the control group, both the Zn concentrations in the
testis (total) and the isolated Leydig cells were significantly
decreased in the ZnD group (Fig.
2a). In addition, we also evaluated the circulating LH content
and testosterone levels in the serum. As expected, the mice fed a
ZnD diet had significantly lower serum concentrations of
testosterone and higher serum concentrations of LH than in the
control group (Fig. 2b and
c).
Effects of Zn deficiency on the protein
expression of ZnT7 and testosterone-related factors in the mouse
Leydig cells
To assess the effect of Zn deficiency on the
expression of ZnT7 and testosterone-related factors in mouse Leydig
cells, several important transport proteins and enzymes involved in
testosterone synthesis were selected for evaluation, namely StAR,
P450scc and 3β-HSD (29–31). P450scc is a hormonally-regulated
mitochondrial enzyme which converts insoluble cholesterol to
soluble pregnenolone (30).
Pregnenolone is then metabolized to androgens by 3β-HSD (25,30). Notably, Zn deficiency which
downregulated ZnT7 levels also decreased the protein expression of
StAR, P450scc and 3β-HSD compared with that in the control groups
(Fig. 3). Overall, these results
suggest that ZnT7 plays an essential role in the reduced
testosterone synthesis that occurs in response to Zn
deficiency.
RNAi-mediated knockdown of ZnT7 and Zn
deficiency may inhibit hCG-induced testosterone synthesis and
downregulate testosterone-related factors in MLTC-1 cells
In the present study, MLTC-1 cells were used as a
model cell line and hCG was used to induce steroidogenesis in order
to elucidate the mechanism underlying the involvement of ZnT7 in
testosterone synthesis. The established MLTC-1 cell line has the
capability to synthesize progesterone as the key steroid in
response to hCG and to keep to a consistent gonadotropin response
before progesterone synthesis compared with the primary Leydig cell
line (32). The cells were
treated with increasing amounts of hCG for 30 min, and the
progesterone levels in the media were measured using a
chemiluminescent enzyme immunoassay. The saturating amount of hCG
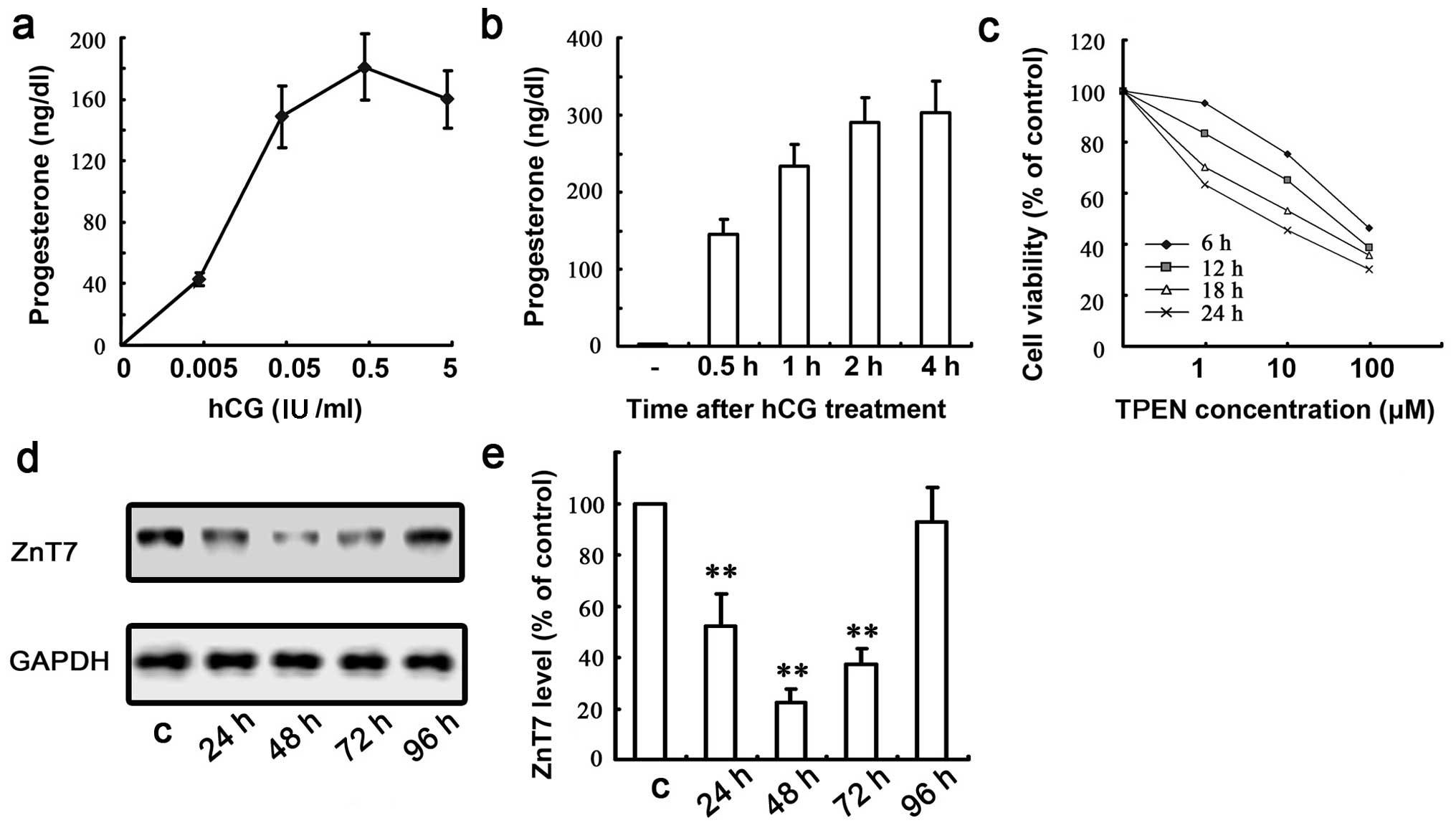
was approximately 0.5 IU/ml (Fig.
4a). This amount of hCG (0.5 IU/ml) was used in subsequent
experiments. The hCG-induced steroid production was consistently
detectable as early as 0.5 h post-treatment and reached a steady
peak at approximately 2–4 h post-treatment (Fig. 4b). We selected 2 h as the optimum
incubation time for subsequent experiments. To avoid potential
toxic effects resulting in cell death in the following experiments,
we performed a cell viability assay in order to determine the
optimal TPEN concentration and incubation time (Fig. 4c). Accordingly, a concentration of
1 μm TPEN and a 12 h incubation time were selected.
To examine the effects of ZnT7 on testosterone
synthesis, we determined whether ZnT7 knockdown was capable of
preventing the events of steroidogenesis. Firstly, western blot
analysis was performed in order to detect ZnT7 protein expression
following the transfection of MLTC-1 cells with mZnT7 siRNA, which
revealed that the optimal effect was achieved at 48 h
post-transfection and the inhibition rate of ZnT7 RNAi reached
>80% (Fig. 4d and e).
Subsequently, hCG-stimulated cells were then used to determine
whether Zn homeostasis and ZnT7 expression are involved in
steroidogenesis in MLTC-1 cells (33–35). The MLTC-1 cells were treated with
0.5 IU/ml of hCG for 2 h with 1 μM TPEN or ZnT7 siRNA and
progesterone production in the media was measured at 2 h after hCG
treatment. ZnT7 knockdown decreased progesterone production and
downregulated the expression of StAR, P450scc and 3β-HSD following
hCG stimulation of the MLTC-1 cells. In addition, exposure to TPEN
decreased progesterone levels and the factors mentioned above
(Fig. 5a–e). Taken together,
these findings suggest that ZnT7 appears to play a role in
steroidogenesis induced by hCG, by modulating the expression of
several steroidogenic enzymes.
Discussion
Previous studies have reported that Zn plays
important roles in the production, storage and secretion of a
number of hormones including testosterone (26,36). Studies in animal models as well as
clinical studies have demonstrated that hypogonadism is a prominent
feature of Zn deficiency in humans and animals (37,38); however, the underlying mechanisms
responsible for these endocrine effects observed in Zn deficiency
remain unclear. The present study has reported a significant
decrease in serum testosterone levels and Zn concentrations in the
testis and Leydig cells of mice, which is in line with the findings
of previous studies on a number of male mammal species (37–40). In addition, ZnT7 was prinicipally
expressed and colocalized with StAR in the Leydig cells of male
CD-1 mice. ZnT7 expression was downregulated in the mice fed a ZnD
(0.85 ppm) diet as were the enzymes involved in testosterone
synthesis, including P450scc and 3β-HSD, as well as testosterone
levels. Furthermore, ZnT7 gene silencing downregulated the levels
of StAR, P450scc, 3β-HSD and progesterone in the hCG-stimulated
MLTC-1 cells. These results are the first to demonstrate that Znt7
is involved in testosterone synthesis in the mouse testis and
MLTC-1 cells, to the best of our knowledge. The mechanism
underlying this process may involve the modulation of the
expression levels of testosterone-related factors in MLTC-1
cells.
Zn is critical for the production, storage and
secretion of various enzymes, including a number of Zn-dependent
enzymes, that are involved in cellular signaling pathways as well
as for the activity of certain transcription factors (41,42). Studies in humans and animal models
have determined that severe Zn deficiency negatively affects male
reproduction by inducing testicular atrophy, reducing the
functioning of LH receptor and steroid synthesis, and causing
Leydig cell damage as a result of oxidative stress (9,43).
In addition, Zn deficiency may cause the degeneration of Leydig
cells involved in the processing of spermatozoa after meiotic
division (40). More
specifically, ZnD testicular Leydig cells are capable of-taking up
cholesterol and neutral lipids which are precursors of sex
steroids; however, they are unable to convert them into sex
steroids. This leads to the arrest of spermatogenesis and the
impairment of fertilization (10,38). The present study demonstrated that
in male mice, dietary Zn deficiency significantly reduced the level
of testosterone and increased the circulating level of LH, thus
indicating that the release and synthesis of LH occurred under
conditions of Zn depletion. These data are in agreement with
previous findings obtained by other researchers, who demonstrated
that rats exhibited significant decreases in the serum levels of
testosterone and progesterone together with increases in serum
levels of follicle-stimulating hormone (FSH) and LH (3,5,44).
Furthermore, in vitro experiments have demonstrated that
decreased levels of progesterone synthesis by Leydig cells as well
as the decreased expression of StAR, P450scc and 3β-HSD in the
Leydig cells occurred under conditions of Zn depletion. The results
of in vitro and in vivo experiments in the present
study have therefore indicated that Zn deficiency may exert a
direct inhibitory effect on testosterone synthesis in Leydig cells.
This explains the reduction in testosterone synthesis, which was
confirmed by evidence that progesterone production was decreased
and the expression of the key steroidogenic enzymes was
downregulated in the mice with dietary Zn deficiency.
It is believed that ZnTs are involved in a number of
physiological pathways and processes, such as insulin synthesis and
secretion in the pancreas, cognitive function, bone development and
body adiposity (37,45). The roles of ZnTs in the
pathogenesis of disease have received great attention and
monitoring cellular Zn status may have an important impact on
disease prevention and treatment (45,46). Several studies have demonstrated
that ZnT7 is located in the Golgi apparatus and may participate in
the transport of cytoplasmic Zn into the Golgi apparatus (21,47). ZnT7-deficient rats had lower
levels of Zn in their cells and tissues, a reduced food intake, and
poor growth; all of which are clinical manifestations of Zn
deficiency (39). In the present
study, double immunofluorescence staining revealed the
colocalization of ZnT7 and StAR in the mouse Leydig cells. The
silencing of ZnT7 gene expression demonstrated the impact of
changes in ZnT7 expression on testosterone secretion as well as the
expression of the enzymes involved in testosterone synthesis in
MLTC-1 cells which suggested that ZnT7 may play a role in the the
regulation of steroidogenesis. It is well known that ZnT7 may play
a direct role in maintaining Zn concentrations within the Golgi
apparatus, which is a vital organelle for protein synthesis and
post-translational modification (48–50). Previous research has demonstrated
that the overexpression of ZnT7 in Chinese hamster ovarian cells
led to an accumulation of Zn in the Golgi apparatus, indicating
that ZnT7 may play an important role in delivering Zn to newly
synthesized Zn-requiring enzymes, proteins, or in the storage of Zn
in the Golgi apparatus (49,51). In the present study, we found that
the protein expression of StAR, P450scc and 3β-HSD were
significantly decreased in the ZnT7-silenced MLTC-1 cells, compared
with that in the control groups. The changes we observed suggest
that ZnT7 may affect testosterone synthesis in MLTC-1 cells by
regulating the levels of testosterone-related factors through the
metal-responsive element in testosterone synthesis promoters and
Zn-requiring proteins or processes, or through the export of Zn
from the Golgi apparatus through ZnTs.
In conclusion, in the present study we present novel
evidence regarding the association between ZnT7 and testosterone
synthesis through in vivo experiments on the tests of male
CD-1 mice and in vitro experiments on MLTC-1 cells. Our
results have demonstrated that the expression of ZnT7 as well as
StAR, P450scc and 3β-HSD was reduced, respectively, in the ZnD
group together with a decreased serum testosterone level. In
particular, the silencing of the ZnT7 gene resulted in the reduced
expression of StAR, P450scc and 3β-HSD as well as in decreased
progesterone levels. Given the important role that testosterone
synthesis plays in the development and progression of gonadal
differentiation, formation and maturation, the identification of
ZnT7 as a key regulator of testosterone synthesis represents an
important finding. Future studies may confirm ZnT7 as a potentially
important target for therapeutic intervention, in an attempt to
regulate steroidogenesis in order to limit the decline in fertility
observed in patients with Zn deficiency.
Abbreviations:
|
MLTC-1 cells
|
mouse Leydig tumor cell line
|
|
FITC
|
fluorescein isothiocyanate
|
|
TPEN
|
N,N,N′,N′-tetrakis(2-pyridylmethyl)
ethylenediamine
|
|
PBS
|
phosphate-buffered saline
|
|
ZnT
|
zinc transporter
|
|
LH
|
luteinizing hormone
|
|
FSH
|
follicle-stimulating hormone
|
|
3β-HSD
|
3-β-hydroxysteroid dehydrogenase
|
|
P450scc
|
cholesterol side-chain cleavage
enzyme
|
|
StAR
|
steroidogenic acute regulatory
protein
|
|
CLSM
|
confocal laser scanning microscope
|
|
AAS
|
atomic absorption
spectrophotometry
|
Acknowledgments
The present study was supported by the National
Grand Fundamental Research 973 Program of China (no. 2012CB722405),
the Natural Science Foundation of China (no. 81170561), and the
Liaoning Province Science and Technology Plan Project (nos.
2012408002 and 2013225090).
References
|
1
|
Hidiroglou M and Knipfel JE: Zinc in
mammalian sperm: a review. J Dairy Sci. 67:1147–1156. 1984.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Coleman JE: Zinc proteins: enzymes,
storage proteins, transcription factors, and replication proteins.
Annu Rev Biochem. 61:897–946. 1992. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Hartoma TR, Nahoul K and Netter A: Zinc,
plasma androgens and male sterility. Lancet. 2:1125–1126. 1977.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Aihara K, Nishi Y, Hatano S, Kihara M,
Ohta M, Sakoda K, Uozumi T and Usui T: Zinc, copper, manganese, and
selenium metabolism in patients with human growth hormone
deficiency or acromegaly. J Pediatr Gastroenterol Nutr. 4:610–615.
1985. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Bedwal RS and Bahuguna A: Zinc, copper and
selenium in reproduction. Experientia. 50:626–640. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
El Hendy HA, Yousef MI and Abo El-Naga NI:
Effect of dietary zinc deficiency on hematological and biochemical
parameters and concentrations of zinc, copper, and iron in growing
rats. Toxicology. 167:163–170. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Matossian MK: Fertility decline in Europe
1875–1913. Was zinc deficiency the cause? Perspect Biol Med.
34:604–616. 1991. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Brown KH and Wuehler SE: Zinc and human
health: results of recent trials and implications for program
interventions and research. Micronutrient Initiative; Ottawa:
2000
|
|
9
|
Hesketh JE: Effects of dietary zinc
deficiency on Leydig cell ultrastructure in the boar. J Comp
Pathol. 92:239–247. 1982. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Lei KY, Abbasi A and Prasad AS: Function
of pituitary-gonadal axis in zinc-deficient rats. Am J Physiol.
230:1730–1732. 1976.PubMed/NCBI
|
|
11
|
Hadwan MH, Almashhedy LA and Alsalman AR:
Study of the effects of oral zinc supplementation on peroxynitrite
levels, arginase activity and NO synthase activity in seminal
plasma of Iraqi asthenospermic patients. Reprod Biol Endocrinol.
12:12014. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Abbasi AA, Prasad AS and Rabbani PR:
Experimental zinc deficiency in man: effect on spermatogenesis.
Trans Assoc Am Physicians. 92:292–302. 1979.PubMed/NCBI
|
|
13
|
Khan MS, Zaman S, Sajjad M, Shoaib M and
Gilani G: Assessment of the level of trace element zinc in seminal
plasma of males and evaluation of its role in male infertility. Int
J Appl Basic Med Res. 1:93–96. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Chasapis CT, Loutsidou AC, Spiliopoulou CA
and Stefanidou ME: Zinc and human health: an update. Arch Toxicol.
86:521–534. 2012. View Article : Google Scholar
|
|
15
|
Kambe T, Yamaguchi-Iwai Y, Sasaki R and
Nagao M: Overview of mammalian zinc transporters. Cell Mol Life
Sci. 61:49–68. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Palmiter RD and Huang L: Efflux and
compartmentalization of zinc by members of the SLC30 family of
solute carriers. Pflugers Arch. 447:744–751. 2004. View Article : Google Scholar
|
|
17
|
Kambe T, Hashimoto A and Fujimoto S:
Current understanding of ZIP and ZnT zinc transporters in human
health and diseases. Cell Mol Life Sci. 71:3281–3295. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Palmiter RD, Cole TB, Quaife CJ and
Findley SD: ZnT-3, a putative transporter of zinc into synaptic
vesicles. Proc Natl Acad Sci USA. 93:14934–14939. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Cole TB, Wenzel HJ, Kafer KE,
Schwartzkroin PA and Palmiter RD: Elimination of zinc from synaptic
vesicles in the intact mouse brain by disruption of the ZnT3 gene.
Proc Natl Acad Sci USA. 96:1716–1721. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Kambe T, Narita H, Yamaguchi-Iwai Y,
Hirose J, Amano T, Sugiura N, Sasaki R, Mori K, Iwanaga T and Nagao
M: Cloning and characterization of a novel mammalian zinc
transporter, zinc transporter 5, abundantly expressed in pancreatic
beta cells. J Biol Chem. 277:19049–19055. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Kirschke CP and Huang L: ZnT7, a novel
mammalian zinc transporter, accumulates zinc in the Golgi
apparatus. J Biol Chem. 278:4096–4102. 2003. View Article : Google Scholar
|
|
22
|
Lemaire K, Chimienti F and Schuit F: Zinc
transporters and their role in the pancreatic β-cell. J Diabetes
Investig. 3:202–211. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Chimienti F, Devergnas S, Favier A and
Seve M: Identification and cloning of a beta-cell-specific zinc
transporter, ZnT-8, localized into insulin secretory granules.
Diabetes. 53:2330–2337. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Kambe T, Weaver BP and Andrews GK: The
genetics of essential metal homeostasis during development.
Genesis. 46:214–228. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Jiang MH, Cai B, Tuo Y, Wang J, Zang ZJ,
Tu X, Gao Y, Su Z, Li W, Li G, et al: Characterization of
Nestin-positive stem Leydig cells as a potential source for the
treatment of testicular Leydig cell dysfunction. Cell Res.
24:1466–1485. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Wang H, Wang Q, Zhao XF, Liu P, Meng XH,
Yu T, Ji YL, Zhang H, Zhang C, Zhang Y and Xu DX: Cypermethrin
exposure during puberty disrupts testosterone synthesis via
downregu-lating StAR in mouse testes. Arch Toxicol. 84:53–61. 2010.
View Article : Google Scholar
|
|
27
|
Caron KM, Soo SC, Wetsel WC, Stocco DM,
Clark BJ and Parker KL: Targeted disruption of the mouse gene
encoding steroidogenic acute regulatory protein provides insights
into congenital lipoid adrenal hyperplasia. Proc Natl Acad Sci USA.
94:11540–11545. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
King SR, Manna PR, Ishii T, Syapin PJ,
Ginsberg SD, Wilson K, Walsh LP, Parker KL, Stocco DM, Smith RG and
Lamb DJ: An essential component in steroid synthesis, the
steroidogenic acute regulatory protein, is expressed in discrete
regions of the brain. J Neurosci. 22:10613–10620. 2002.PubMed/NCBI
|
|
29
|
Stocco DM and Clark BJ: The role of the
steroidogenic acute regulatory protein in steroidogenesis.
Steroids. 62:29–36. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Raucci F, D'Aniello A and Di Fiore MM:
Stimulation of androgen production by D-aspartate through the
enhancement of StAR, P450scc and 3β-HSD mRNA levels in vivo rat
testis and in culture of immature rat Leydig cells. Steroids.
84:103–110. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Stocco DM: StAR protein and the regulation
of steroid hormone biosynthesis. Annu Rev Physiol. 63:193–213.
2001. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Hu Y, Dong C, Chen M, Lu J, Han X, Qiu L,
Chen Y, Qin J, Li X, Gu A, et al: Low-dose monobutyl phthalate
stimulates steroidogenesis through steroidogenic acute regulatory
protein regulated by SF-1, GATA-4 and C/EBP-beta in mouse Leydig
tumor cells. Reprod Biol Endocrinol. 11:722013. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Pakarinen P, Vihko KK, Voutilainen R and
Huhtaniemi I: Differential response of luteinizing hormone receptor
and steroidogenic enzyme gene expression to human chorionic
gonadotropin stimulation in the neonatal and adult rat testis.
Endocrinology. 127:2469–2474. 1990. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Baba Y: Effect of HCG administration in
the gossypol-treated male rats. Hinyokika Kiyo. 34:643–647. 1988.In
Japanese. PubMed/NCBI
|
|
35
|
Scott IS, Charlton HM, Cox BS, Grocock CA,
Sheffield JW and O'Shaughnessy PJ: Effect of LH injections on
testicular steroidogenesis, cholesterol side-chain cleavage P450
mRNA content and Leydig cell morphology in hypogonadal mice. J
Endocrinol. 125:131–138. 1990. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Xu Z, Kawai M, Bandiera SM and Chang TK:
Influence of dietary zinc deficiency during development on hepatic
CYP2C11, CYP2C12, CYP3A2, CYP3A9, and CYP3A18 expression in
postpubertal male rats. Biochem Pharmacol. 62:1283–1291. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Croxford TP, McCormick NH and Kelleher SL:
Moderate zinc deficiency reduces testicular Zip6 and Zip10
abundance and impairs spermatogenesis in mice. J Nutr. 141:359–365.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Favier AE: The role of zinc in
reproduction. Hormonal mechanisms. Biol Trace Elem Res. 32:363–382.
1992. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Om AS and Chung KW: Dietary zinc
deficiency alters 5 alpha-reduction and aromatization of
testosterone and androgen and estrogen receptors in rat liver. J
Nutr. 126:842–848. 1996.PubMed/NCBI
|
|
40
|
Qureshi IZ and Abbas Q: Modulation of
testicular and whole blood trace element concentrations in
conjunction with testosterone release following kisspeptin
administration in male rabbits (Oryctolagus cuniculus). Biol Trace
Elem Res. 154:210–216. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Sunderman FW Jr: The influence of zinc on
apoptosis. Ann Clin Lab Sci. 25:134–142. 1995.PubMed/NCBI
|
|
42
|
Vallee BL and Falchuk KH: The biochemical
basis of zinc physiology. Physiol Rev. 73:79–118. 1993.PubMed/NCBI
|
|
43
|
Prasad AS: Clinical and biochemical
manifestation zinc deficiency in human subjects. J Pharmacol.
16:344–352. 1985.PubMed/NCBI
|
|
44
|
Root AW, Duckett G, Sweetland M and Reiter
EO: Effects of zinc deficiency upon pituitary function in sexually
mature and immature male rats. J Nutr. 109:958–964. 1979.PubMed/NCBI
|
|
45
|
Huang L and Tepaamorndech S: The SLC30
family of zinc transporters - a review of current understanding of
their biological and pathophysiological roles. Mol Aspects Med.
34:548–560. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Kelleher SL, Seo YA and Lopez V: Mammary
gland zinc metabolism: regulation and dysregulation. Genes Nutr.
4:83–94. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Chi ZH, Feng WY, Gao HL, Zheng W, Huang L
and Wang ZY: ZNT7 and Zn2+ are present in different cell
populations in the mouse testis. Histol Histopathol. 24:25–30.
2009.
|
|
48
|
Chi ZH, Ren H, Wang X, Rong M, Huang L and
Wang ZY: The cellular and subcellular localization of zinc
transporter 7 in the mouse spinal cord. Histol Histopathol.
23:781–787. 2008.PubMed/NCBI
|
|
49
|
Huang L, Yan M and Kirschke CP:
Over-expression of ZnT7 increases insulin synthesis and secretion
in pancreatic beta-cells by promoting insulin gene transcription.
Exp Cell Res. 316:2630–2643. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Mukherjee S, Chiu R, Leung SM and Shields
D: Fragmentation of the Golgi apparatus: an early apoptotic event
independent of the cytoskeleton. Traffic. 8:369–378. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Chi ZH, Wang X, Wang ZY, Gao HL, Dahlstrom
A and Huang L: Zinc transporter 7 is located in the cis-Golgi
apparatus of mouse choroid epithelial cells. Neuroreport.
17:1807–1811. 2006. View Article : Google Scholar : PubMed/NCBI
|