Introduction
Ischemia-reperfusion injury (IRI) caused by
hypoxia/reoxygenation (H/R) in the liver is a common cause of liver
damage during liver resection, liver transplantation and hepatic
failure (1–4). It has been widely demonstrated that
interleukin-6 (IL-6) is a vital mediator, and IL-6/signal
transducer and activator of transcription 3 (STAT3) signaling is of
major importance in restoring normal hepatic function following
liver IRI (5–7). Studies have found that IL-6 is
transported to the liver in an endocrine manner from immunocytes at
the wound site, or in a paracrine manner from hepatic immunocytes
within the liver (8,9). However, the biological mechanisms of
IL-6 synthesis in parenchymal hepatic cells, not hepatic
immunocytes, has previously been overlooked.
Additionally, the prevalence of rs1800795 (-174G/C)
and rs1800797 locus polymorphisms in the IL-6 gene promoter
is extremely rare, whereas the allele C at the IL-6
rs1800796 locus is frequently observed in East Asian populations
(10-13). Although studies have reported that
IL-6 rs1800796 (also designated as -572C/G or -634G>C)
single nucleotide polymorphisms (SNPs) affect IL-6
transcriptional activity, their conclusions were not always
consistent among similar populations (13,14). The rs1800796 locus SNPs may be
present in a regulatory DNA binding site, leading to altered
affinity with the regulatory protein, and may subsequently
influence IL-6 transcription efficiency (15). Furthermore, only donor IL-6
rs1800796 SNPs affect the metabolism of tacrolimus, an
immunosuppressive drug, mainly by altering liver function and
regeneration in response to hepatic H/R following liver
transplantation (16). Therefore,
we hypothesized that IL-6 and its rs1800796 SNPs in
parenchymal hepatic cells may protect liver function during H/R,
and that the sequence variants of rs1800796 SNPs (G→C) may increase
the transcription efficiency of IL-6. Thus, in order to
confirm our hypothesis, in the present study, we investigated the
response of IL-6 and its rs1800796 locus SNPs to hepatic H/R
stimulus in parenchymal liver cells.
Materials and methods
Collection of liver tissue samples
Disease-free liver tissues were collected from 84
male Chinese volunteers at the Shanghai Jiao Tong University
Affiliated First People's Hospital (Shanghai, China) after
obtaining written informed consent. The mean age of the subjects
was 35.8±8.3 years. The subjects had no hepatic disease,
cardiovascular or cerebrovascular disease that affected the serum
IL-6 levels. Each sample was immediately cryopreserved in liquid
nitrogen, and then transferred to refrigerator at -80°C. All the
surgical protocols were conducted similarly. This study was
conducted according to the Declaration of Helsinki and its
amendments, and was approved by the Ethics Committee of the Medical
Faculties of Shanghai Jiao Tong University.
Materials
Rosewell Park Memorial Institute (RPMI)-1640 medium
and fetal bovine serum (FBS) were purchased from Gibco (Grand
Island, NY, USA). Dulbecco's modified Eagle's medium (DMEM), TRIzol
reagemt and the SuperScript VILO cDNA Synthesis kit were purchased
from Invitrogen (Carlsbad, CA, USA). Wild-type overexpressed
IL-6 vectors named p-IL-6(WT), negative control IL-6
overexpression vectors, HG transgene reagent, all lentivectors, and
the Lentiviral Packaging Plasmid Mix (Lenti-HG Mix) were obtained
from GenomeDitech Co., Ltd. (Shanghai, China). Restriction
endonucleases (BamHI, ClaI and EcoRI) were
purchased from MBI Fermentas (MBI Fermentas, Burlington, ON
Canada). The QuickChange™ Site-Directed Mutagenesis kit was
obtained from Stratagene (La Jolla, CA, USA). The RNeasy mini kits
and the AllPrep DNA/RNA Mini kits were obtained from Qiagen
(Venlos, The Netherlands). The SuperScript VILO cDNA synthesis kits
were obtained from Invitrogen. The cell counting kit-8 (CCK-8) and
alanine aminotransferase (AST) enzyme-linked immunosorbent assay
(ELISA) kits were purchased from R&D Systems (Minneapolis, MN,
USA). The antibodies used were as follows: anti-IL-6 and
anti-STAT3 polyclonal antibodies (ab6672 and ab68153; Abcam,
Cambridge, UK). Ampicillin, puromycin, or blasticidin were obtained
from MP Biomedicals (Irvine, CA, USA). The secondary antibody [goat
anti-rabbit immunoglobulin G (ZB-2301)] was obtained from Golden
Bridge Biotechnology Co., Ltd. (Beijing, China). The human normal
liver cell line LO2 (17) and
293T cells were purchased from the Shanghai Institute of Cell
Biology (Shanghai, China) and the cells were cultured in DMEM
supplemented with 10% FBS. All normal cultures were maintained at
37°C in saturated humidified incubator with 5% CO2.
Stable cell transfection with IL-6 short
hairpin RNA
Four IL-6-targeted short hairpin RNAs (IL-6-shRNAs),
as well as one negative control mismatch sequence (IL-6-NC-shRNA),
were synthesized by Sangon Biotech Co., Ltd. (Shanghai, China). The
targeting shRNA sequences containing recognition sites for
BamHI in the forward primer and EcoRI in the reverse
primers are shown in Table I
(Primers 1–5). Subsequently, the pGMLV-SC7-IL-6-shRNA lentiviral
vectors were constructed using double restriction enzyme digestion
and gene recombination. The sequence of these lentivectors was
confirmed using direct sequencing. Subsequently, 293T cells were
transiently transfected with the recombinant lentivectors, Lenti-HG
Mix and HG transgene reagent as described in the manufacturer's
manual. The packaging virus was collected from the cell supernatant
through lysis after 3 consecutive freeze-thaw cycles. The viral
titer was approximately 1×109 TU/ml, which was tested
using end point dilution assay. The LO2 cells were then steadily
transfected with lentivirus particles at a multiplicity of
infection (MOI) of 20, and the green fluorescence protein in the
cells was observed using a laser fluorescence microscope (serial
no. N-STORM; Nikon, Tokyo, Japan). Following transfection, the
cells were subjected to drug screening with the corresponding
antibiotics.
 | Table IPrimer sequences used in this
study. |
Table I
Primer sequences used in this
study.
| No. | Name | Primer
sequences |
|---|
| Primer 1 | IL-6-shRNA1 | F:
5′-gatccGCCCTGAGAAAGGAGACATGTTTCAAGAGAACATGTCTCCTTTCTCAGGGCTTTTTTg-3′ |
| | R:
5′-aattcAAAAAAGCCCTGAGAAAGGAGACATGTTCTCTTGAAACATGTCTCCTTTCTCAGGGCg-3′ |
| Primer 2 | IL-6-shRNA2 | F:
5′-gatccGGATGCTTCCAATCTGGATTCTTCAAGAGAGAATCCAGATTGGAAGCATCCTTTTTTg-3′ |
| | R:
5′-aattcAAAAAAGGATGCTTCCAATCTGGATTCTCTCTTGAAGAATCCAGATTGGAAGCATCCg-3′ |
| Primer 3 | IL-6-shRNA3 | F:
5′-gatccGGAGTTTGAGGTATACCTAGATTCAAGAGATCTAGGTATACCTCAAACTCCTTTTTTg-3′ |
| | R:
5′-aattcAAAAAAGGAGTTTGAGGTATACCTAGATCTCTTGAATCTAGGTATACCTCAAACTCCg-3′ |
| Primer 4 | IL-6-shRNA4 | F:
5′-gatccGCAGGACATGACAACTCATCTTTCAAGAGAAGATGAGTTGTCATGTCCTGCTTTTTTg-3′ |
| | R:
5′-aattcAAAAAAGCAGGACATGACAACTCATCTTCTCTTGAAAGATGAGTTGTCATGTCCTGCg-3′ |
| Primer 5 | IL-6-NC-shRNA | F:
5′-gatccGTTCTCCGAACGTGTCACGTTTCAAGAGAACGTGACACGTTCGGAGAACTTTTTTACGCGTg-3′ |
| | R:
5′-aattcACGCGTAAAAAAGTTCTCCGAACGTGTCACGTTCTCTTGAAACGTGACACGTTCGGAGAACg-3′ |
| Primer 6 | Cell IL-6 | F:
5′-CAGACAGCCACTCACCTCTTCAG-3′ |
| | R:
5′-CTGCCAGTGCCTCTTTGCTG-3′ |
| Primer 7 | Cell STAT3 | F:
5′-CTTTGAGACCGAGGTGTATCACC-3′ |
| | R:
5′-GGTCAGCATGTTGTACCACAGG-3′ |
| Primer 8 | Cell GAPDH | F:
5′-GTCTCCTCTGACTTCAACAGCG-3′ |
| | R:
5′-ACCACCCTGTTGCTGTAGCCAA-3′ |
| Primer 9 | p-ProG | F:
5′-GGCCatcgatTCTAAGTGGGCTGAAGCAGGTGAAG-3′ |
| | R:
5′-GCCGgaattcTGGGCTCCTGGAGGGGA-3′ |
| Primer 10 | p-ProC | F:
5′-TCTACAACAGCCCCTCACAGGGAGAGC-3′ |
| | R:
5′-GCTCTCCCTGTGAGGGGCTGTTGTAGA-3′ |
| Primer 11 | IL-6(WT) | F:
5′-CCGgaatccGCCACC ATG
AACTCCTTCTCCACAAGCGC-3′ |
| | R:
5′-CCGggatccCTACATTTGCCGAAGAGCCCTCA-3′ |
| Primer 12 | IL-6(MT) | F:
5′-CCAAAGATGGCTGAAAAAGATGGCTGTTTTCAGAGCGGCTTTAATGAGGAGA-3′ |
| | R:
5′-CACCAGGCAAGTCTCCTCATTAAAGCCGCTCTGAAAACAGCCATCTTTTTCA-3′ |
| Primer 13 | IL-6 in liver
tissue | F:
5′-GACCCAACCACAAATGCCAG-3′ |
| | R:
5′-ACATTTGCCGAAGAGCCCTC-3′ |
| Primer 14 | β-actin in liver
tissue | F:
5′-CTCCATCCTGGCGCTGT-3′ |
| | R:
5′-GCTGTCACCTTCACCGTTCC-3′ |
Subsequently, IL-6 expression was measured by
RT-qPCR as described in the section entitled 'RNA isolation and
quantitative RT-PCR'. The primer sequences of the target gene
(IL-6) and housekeeping gene (GAPDH) are listed in
Table I (Primers 6 and 8).
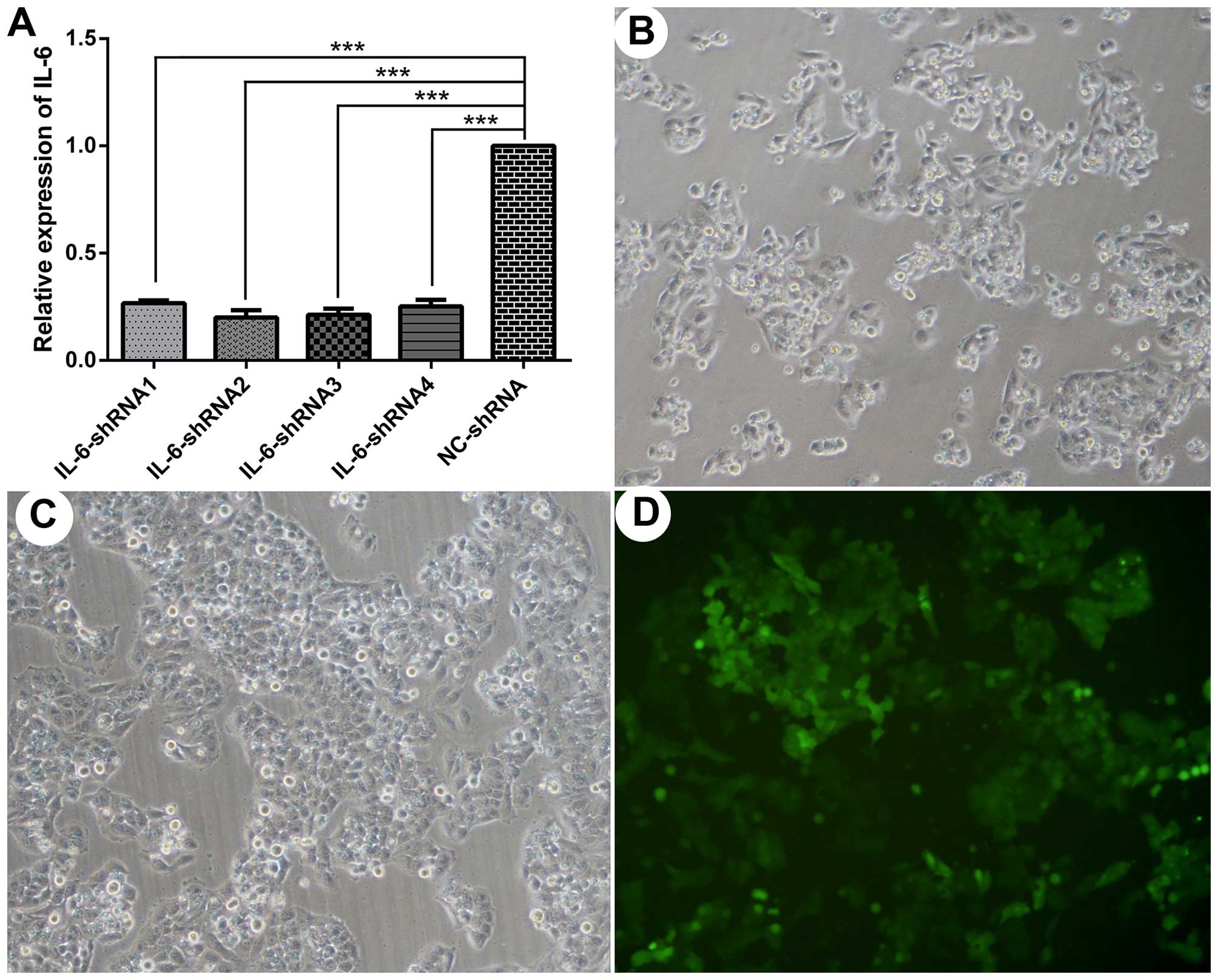
Compared with the control group, the mRNA expression of IL-6
was reduced by 73, 80, 78 and 74% in the cells transfected with
IL-6-shRNA 1, 2, 3 and 4, respectively, when measured at 48
h post-transfection (Fig. 3A).
Therefore, the lentiviral particles harboring IL-6-shRNA2
were selected for use in the subsequent experiments.
Stable cell transfection and clone
selection
First, total DNA was extracted from the LO2 cells
using an AllPrep DNA/RNA mini kit according to the manufacturer's
instructions. A 751-bp sequence (−635 to +116) of the IL-6
promoter region was amplified from the genomic DNA of LO2
homozygous for the -572G allele at the rs1800796 locus (18). Specific primer sequences for the
751-bp amplification contained recognition sites for ClaI in
the forward primer and EcoRI in the reverse primer. The
751-bp fragments containing the -572G allele represented the
wild-type promoters derived from the LO2 cells. The fragments
carrying the -572G allele and the pGMLV-PA2 lentiviral vectors were
doubly digested with restriction endonuclease EcoRI and
ClaI. The cytomegalovirus promoter was removed during this
process in the linear lentiviral vector. Subsequently, the linear
lentivectors and 751-bp fragments were ligated with T4 DNA ligase.
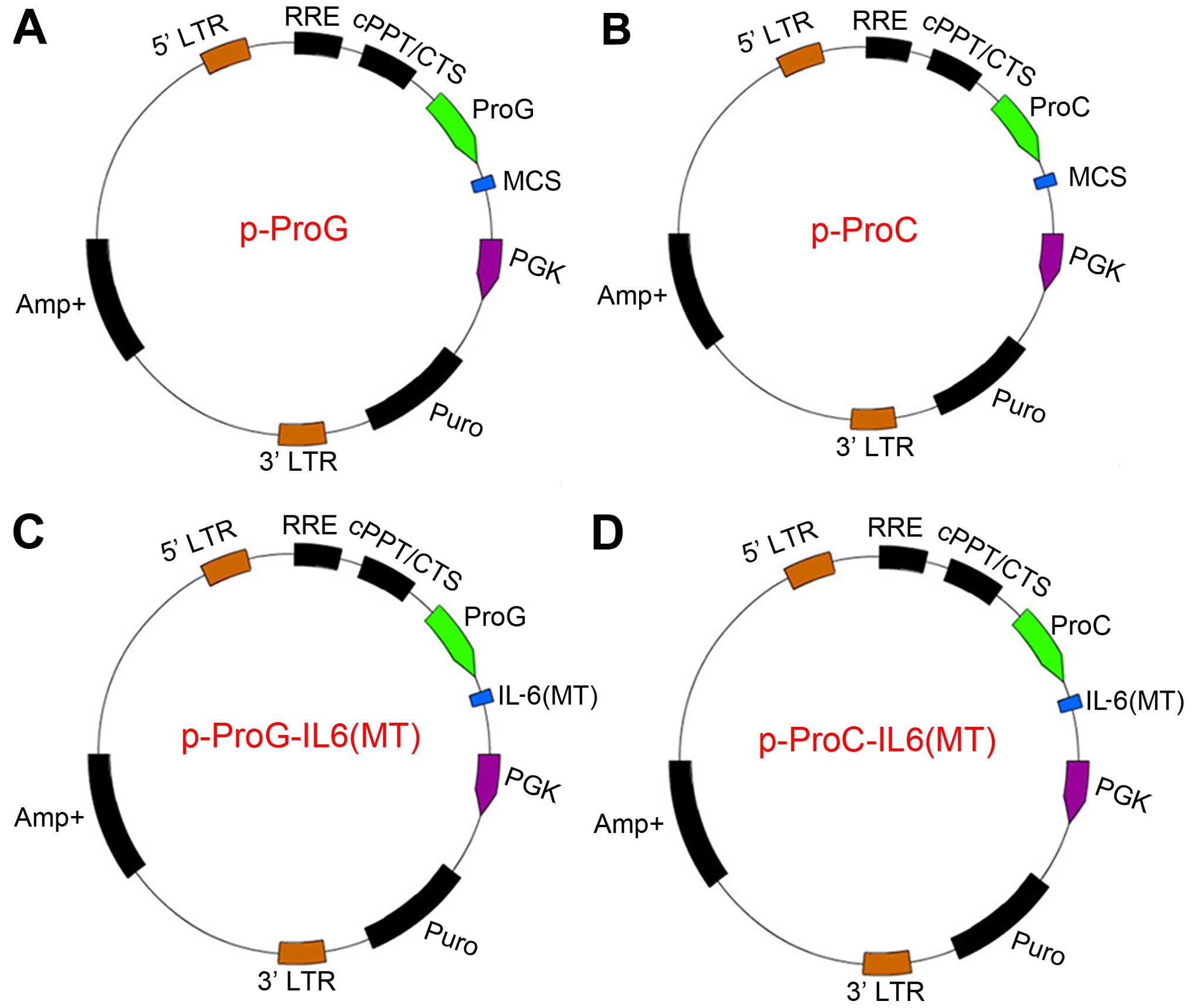
The resulting lentivectors were named p-ProG, which carried the
-572G allele. The G allele at the IL-6 rs1800796 locus was
site-specifically mutated to C with the template of wild-type
promoter and the QuickChange™ Site-Directed Mutagenesis kit, as
previously described (18). The
p-ProC plasmids were synthesized as the p-ProG ones. Specific
primer sequences and the plasmid profile for constructing p-ProG
and p-ProC are listed in Table I
(Primers 9 and 10) and Fig. 1A and
B. The sequence of p-ProG and p-ProC was confirmed using direct
DNA sequencing.
Second, nonsense mutations of
IL-6-overexpressing fragments were obtained based on the
IL-6-shRNA2 sequence and the template of p-IL-6(WT) using
the QuickChange™ Site-Directed Mutagenesis kit, as previously
described (18). The resulting
fragments were named p-NM-IL-6(WT), and guaranteed that IL-6
overexpression was not reduced by IL-6-shRNA2 in the LO2
cells. In particular, NM of p-NM-IL-6(WT) refers to nonsense
mutations. Specific primers used in this process contained
recognition sites for ClaI in the forward primer and
EcoRI in the reverse primer (Table I; Primers 11 and 12). Later, the
p-ProG lentivectors and p-NM-IL-6(WT) were doubly digested with
restriction endonucleases EcoRI and BamHI.
Subsequently, the linear p-ProG vectors and
IL-6-overexpressing fragments were ligated with T4 DNA
ligase. The resulting lentiviral vector was named p-ProG-IL-6(MT)
(Fig. 1C). Subsequently, the
p-ProC-IL-6(MT) lentiviral vector (Fig. 1D) was constructed as was the
p-ProG-IL-6(MT). The altered sequences were verified using direct
sequencing before stable cell transfection.
Third, the negative control
IL-6-overexpressing lentivirus particles (no expression of
IL-6) and IL-6-overexpressing lentivirus particles [carrying
ProG-IL-6(MT) or ProC-IL-6(MT)] were packaged as
pGMLV-SC7-shRNA-IL-6 described in the previous section. Finally,
the LO2 cells with an 80% suppression of basic IL-6 were
cultured in a 6-well culture plate at a density of approximately
2×105 cells/well for 24 h, and then stably transfected
with IL-6-overexpressing lentivirus particles at an
efficiency of approximately 70%. These stably transfected cells
were named LO2C and LO2G, respectively. Similarly, the LO2 cells
stably transfected with IL-6-NC-shRNA were again transfected
with the negative control IL-6-overexpressing lentivirus
particles. The resulting cells were designated as NC-LO2 (negative
control LO2 cells). Following transfection, the cells were
subjected to drug screening with the corresponding antibiotics
(ampicillin, puromycin, or blasticidin).
Cell model of H/R
To induce H/R after ischemia reperfusion in the
liver during liver transplantation and resection, the cell model of
H/R was constructed as follows: cellular hypoxia was maintained by
flushing the incubator set with a continuous gas flow comprising a
mixture of 95% N2 and 5% CO2, as previously
described (19,20). Re-oxygenation was ensured by
continuously flushing with humidified a 95% air and 5%
CO2 gas mixture. This method resulted in an oxygen
content of 0.1% in the chamber. Subsequently, during the
logarithmic growth phase of the LO2C cells, LO2G cells, normal LO2
cells (control LO2) and NC-LO2 cells were inoculated into 96-well
plates or 3.5 cm2 culture dishes, and incubated with
normal medium (DMEM supplemented with 10% FBS) under normoxic
conditions (at 37°C in a saturated humidified incubator with 5%
CO2 and 95% air). Each of the cell types was prepared in
4 groups. Following overnight culture, the medium of these 4 cell
types was changed to serum-free
Krebs-Henseleit-4-(2-hydroxyethyl)-1-piperazineethanesul fonic acid
buffer, as previously described (19), and subjected to hypoxia for 6 h.
Subsequently, all the cells were transferred to fresh, warm,
oxygenated medium and returned to reoxygenation conditions for a
further 1, 6, 12 and 24 h, represented as H/R1h, H/R6h, H/R12h and
H/R24h, respectively. Additionally, another 4 normoxic groups of
cells, as the controls, were incubated for 12 h along with these
H/R-exposed cells. Each group mentioned above was treated in
triplicate.
Other methods
Genomic DNA isolation and genotype
determination
Genomic DNA was extracted from the 84 liver
specimens with an AllPrep DNA/RNA Mini kit according to the
manufacturer's instructions. These samples were previously stored
at −80°C until genotyping. Genotyping of the SNPs was performed
using a Sequenom MassARRAY SNP genotyping platform, as previously
described (21). The protocols
mainly included DNA and primer preparation, PCR amplification,
primer extension, spotting of primer extension products on the
SpectroCHIP, and detection of the primer extension products by mass
spectrometry.
Construction of tissue microarray and
immunohistochemistry assay
Tissue microarray (TMA) was constructed from
disease-free liver tissues (n=84) in collaboration with Shanghai
Biochip (Shanghai, China) as previously described (22). Immunohistochemistry assay was then
performed using the primary anti-IL-6 polyclonal antibody
(1:600). The secondary antibody was goat anti-rabbit immunoglobulin
G. As the negative controls for all the experiments, the primary
antibody was omitted.
Immunohistochemical scoring was performed
independently in a blinded manner by two pathologists. The scoring
standard has been previously described (23).
RNA isolation and quantitative RT-PCR
(RT-qPCR)
Total RNA from the 84 disease-free liver tissues and
all types of LO2 cells was extracted using TRIzol reagent and the
mini RNeasy® kit. First, single-stranded cDNAs were
synthesized using a SuperScript VILO cDNA Synthesis kit.
Subsequently, RT-qPCR was performed using 2× SYBR-Green qPCR mix
and a Mastercycler® Ep Realplex. The fold change of
relative mRNA copies was expressed as the relative quantification
calculated using the 2−ΔΔCt method for each group and
calculated as previously described (23). Namely, the data were normalized to
a housekeeping gene (β-actin or GAPDH). The primer
sequences of the target gene and housekeeping gene are shown in
Table I (Primers 6 to 8, and 13
and 14).
Cell counting kit-8 assay
A total of 2×103 LO2C cells, LO2G cells,
normal LO2 cells (control LO2 cells) and NC-LO2 cells in the
logarithmic growth phase were seeded into 96-well plates, and each
group was allocated at least 3 wells. Subsequently, the cells were
incubated with normal culture medium and subjected to H/R, and the
effects of different H/R conditions on the cell growth or cell
viability were evaluated using the CCK-8 assay according to the
instructions of the manufacturer. The cells were incubated with
CCK-8 (10 μl/well) at 37°C for 3 h. The absorbance (value A)
of individual wells was determined at a 450 nm wavelength. The
formula for calculating cell viability was (value A of treatment
group - value A of blank group)/(value A of control group - value A
of blank group) ×100.
ELISA for determining the AST
level
After being subjected to different H/R conditions or
normal culture, the degree of H/R injury in the LO2 cells was
directly quantified by measuring the concentrations of AST in the
cell supernatants using ELISA kits according to the manufacturer's
instructions. Inter- and intra-detection coefficients of variation
were <10% and <5%, respectively.
Western blot analysis
The expression of STAT3 was determined by
western blot analysis as previously described (4). In brief, total cellular protein was
extracted and separated by electrophoresis on 12% sodium dodecyl
sulfate polyacrylamide gels and transferred onto polyvinylidene
difluoride membranes. The membranes were then washed, blocked and
incubated with the primary antibodies (1:1,000 dilution for
STAT3; 1:1,000 dilution for β-actin) overnight at
4°C, and then incubated with an appropriate horseradish
peroxidase-conjugated secondary antibodies (1:5,000). Signals were
detected by chemiluminescence using ECL luminous fluid and scanned
by Quantity One software. The protein expression of STAT3 was
determined by normalization of the gray value of the STAT3
gene to that of the control housekeeping gene (β-actin).
Then, the resting relative level of STAT3 protein was calculated
using the blank control group as a reference sample.
Statistical analysis
All data were analyzed using SPSS version 19.0
statistical software (SPSS Inc., Chicago, IL, USA) and GraphPad
Prism version 6.02 (GraphPad Software, San Diego, CA, USA).
Continuous variables were reported as the means ± standard
deviation (SD) and examined using the Student's t-test or ANOVA.
Categorical data were analyzed using the Chi-square test. Two-sided
probability values were used for all statistical tests.
Significance was established at the P<0.05 level.
Results
Distribution of genotypes and
alleles
The frequencies of genotypes CC, CG and GG were
46.4, 47.6 and 6.0%, respectively. The allele C and G frequencies
were 70.2 and 29.8% (data not shown). The genotype CG/GG carriers
were pooled into one group, as previously described (16) (Table II). There were no significant
differences between the CC group and CG/GG group with regard to
race, gender, age distribution and body mass index (P>0.05; data
not shown).
| Table IIIL-6 expression in human disease-free
liver tissues in the TMA assay. |
Table II
IL-6 expression in human disease-free
liver tissues in the TMA assay.
| Groups | No. of case | IL-6 expression
| P-value |
|---|
| Weak (%) | Intermediate
(%) |
|---|
| CC | 39 | 11 (28.2) | 28 (71.8) | 0.006 |
| CG/GG | 45 | 26 (57.8) | 19 (42.2) | |
Expression of IL-6 in human disease-free
liver tissues
The expression of IL-6 in the 84 human
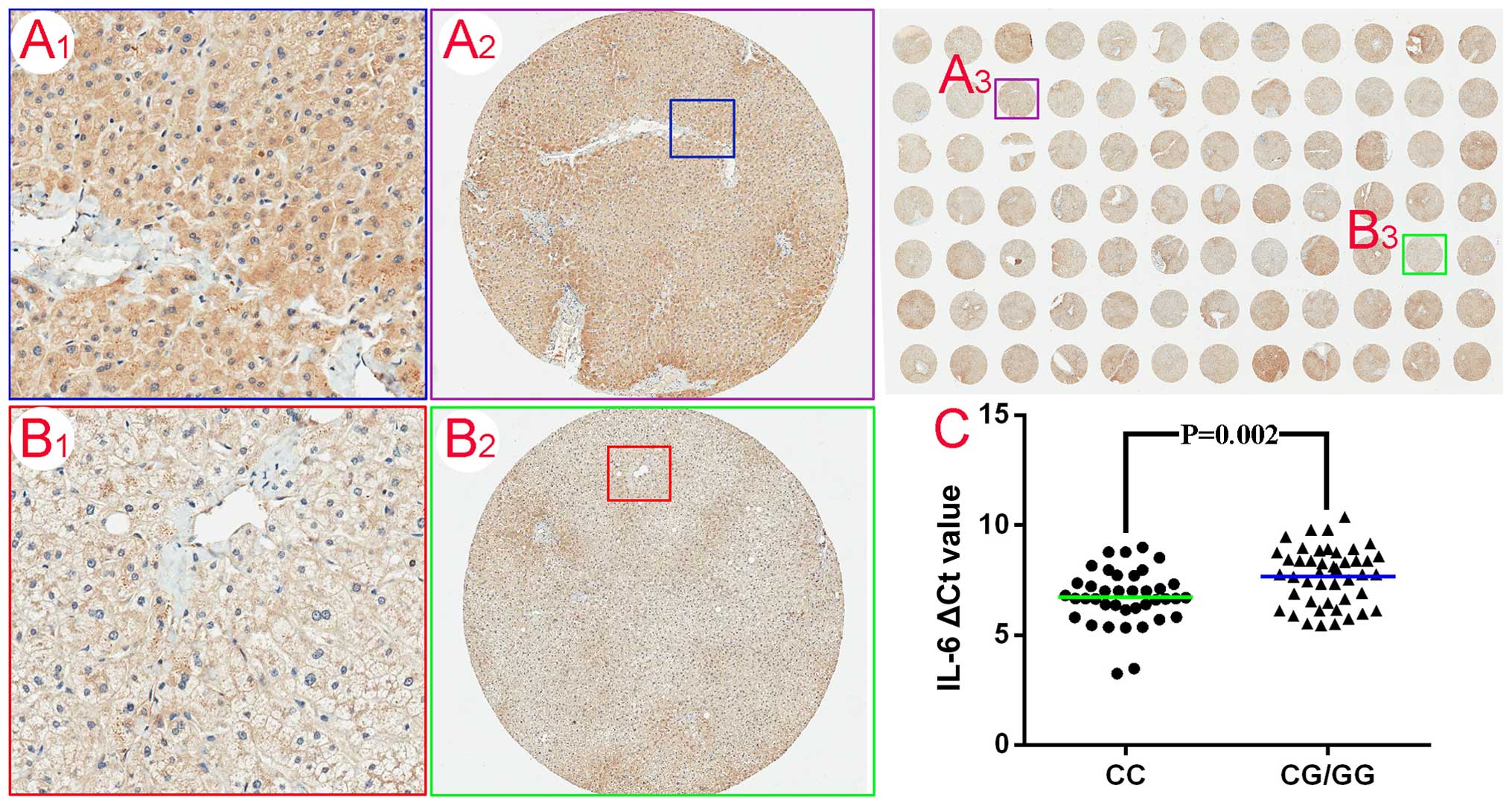
disease-free liver tissues was evaluated using the TMA. The panels
in Fig. 2A1–A3 and B1–B3
represent the intermediate and weak immunostaining profiles of IL-6
protein in hepatocytes. Among the 45 genotype (CG/GG) carriers,
57.8% (26/45) of the specimens exhibited weak staining and 42.2%
(19/45) were intermediately immunoreactive for IL-6 (Table II). However, of the 39 genotype
CC carriers, there was differential IL-6 expression, with
intermediate staining in 71.8% (28/39) of the samples and weak
staining in 28.2% (11/39) of the samples (P=0.006) (Table II). Subsequently, RT-qPCR
analysis was undertaken to further determine the IL-6 mRNA
levels in the 84 liver tissue samples. The ∆Ct value for the
genotype CC carriers was less than that for the CG/GG carriers
(6.7±1.2 vs. 7.6±1.3, P=0.002) (Fig.
2C), confirming the results of immunostaining analysis.
Further, when comparing the results of RT-qPCR between all the
intermediate samples and all weak staining samples, the difference
was more obvious (P<0.001). The protein (staining intensity) and
IL-6 mRNA expression were well correlated.
Construction of LO2 cell lines
The LO2C cells and LO2G cells, stably transfected
with IL-6-shRNA2 lentivirus particles and IL-6
overexpression lentivirus particles, were successfully constructed.
Therefore, the LO2C and LO2G cells produced ectopic IL-6
overexpression and approximately 20% of basic IL-6.
Similarly, the NC-LO2 cells were constructed by stable transfection
with IL-6-NC-shRNA and negative control IL-6-overexpressing
lentiviral particles. The foregoing stable transfection protocol
was described in the Materials and methods. Notably, the NC-LO2
cells served as the negative controls. Normal LO2 cells were used
as blank controls. The LO2G cells contained the wild-type promoter
derived from the LO2 cells, and the WT promoter for the control
compared with the LO2C cells. The double transfection of lentiviral
particles did not alter the cell morphology of the LO2 cells
significantly compared with the normal LO2 cells (Fig. 3B and C) at 48 h. Additionally,
fluorescence analysis of the LO2 cells revealed an infection
lentiviral transduction efficiency of approximately 70% in
accordance with the manufacturer's instructions. There were no
cells in the blank zone, and the fluorescence was 100% after 0.5
μg/ml puromycin selection (Fig. 3D).
H/R injury level of LO2 cells
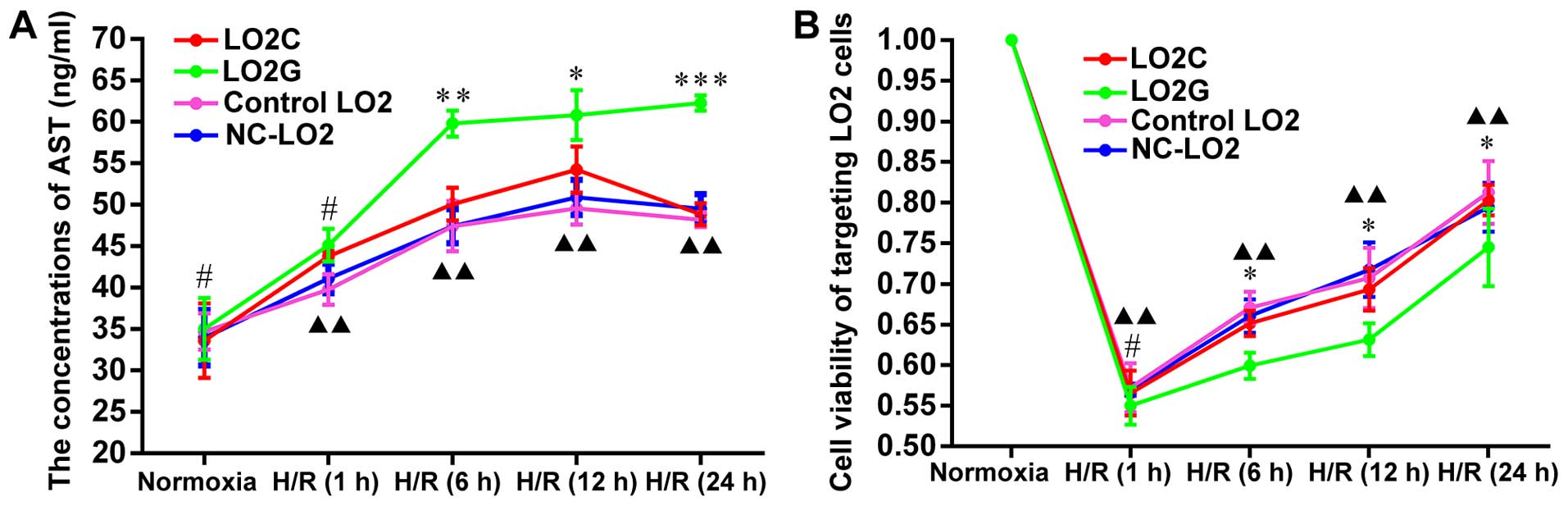
Compared with the normoxic group cells, the
concentrations of AST (ng/ml) in the LO2 cells increased
significantly in all the H/R-exposed groups ranging from 30.18 to
78.98%, suggesting varying degrees of damage in the LO2 cells
(Fig. 4A). In addition, the
concentrations of AST in the LO2G cells were much higher than those
in the LO2C cells at H/R6h, H/R12h and H/R24h (P=0.003, P=0.041 and
P<0.001, respectively) (Fig.
4A). The differences between the experimental groups (LO2C and
LO2G) and the control groups (control LO2 and NC-LO2) were more
significant at all H/R exposure time points (P<0.01). However,
there was no obvious difference between the control LO2 and NC-LO2
cells, which suggested that the lentiviral particles had a
decreased effect on H/R injury in LO2 cells. This difference
between the LO2C and LO2G cells was mainly caused by the sequence
variants of the IL-6 rs1800796 locus SNPs (G→C).
Viability of LO2 cells
Following H/R injury, the rate of survival in the 4
groups of cells was significantly decreased by approximately 45%
compared with the normoxic group of cells. In addition, the
viability of the LO2C cells was signficantly increased compared to
that of the LO2G cells from H/R6h to H/R24h (P=0.034, P=0.016 and
P=0.031) (Fig. 4B). In addition,
there were significant differences between the experimental group
and the control group at all H/R exposure time points (P<0.01)
(Fig. 4B). There were no obvious
differences in cell viability between the control LO2 and NC-LO2
cells, which suggested that the lentiviral particles had less
prominent effects on the biological activity of the LO2 cells. The
results were consistent with the those obtained for the AST
levels.
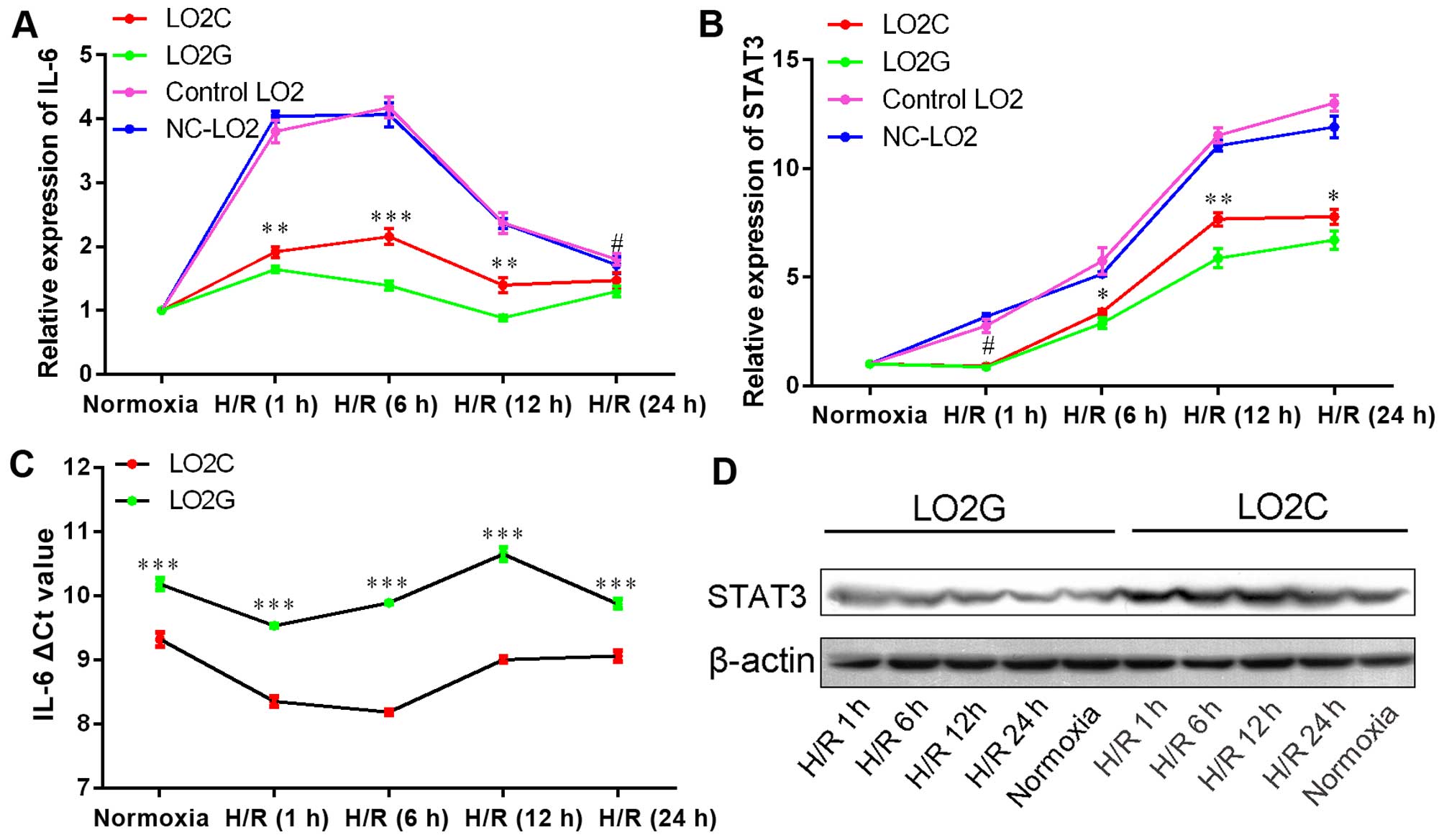
Expression of IL-6 and STAT3 in LO2
cells
Compared with IL-6 expression in the normoxic
groups, the increase in the mRNA expression of IL-6 in the
LO2C cells was greater than that in the LO2G cells from H/R1h to
H/R24h (P=0.009, P<0.001, P=0.002 and P<0.05, respectively)
(Fig. 5A). Similarly,
STAT3 expression also increased from H/R1h to H/R24h
(P<0.05, P=0.034, P=0.004 and P=0.027) (Fig. 5B). Additionally, relative to the
housekeeping gene (GAPDH), IL-6 expression was lower
in the LO2G cells than in the LO2C cells under normoxic conditions
(P<0.001) (Fig. 5C). This
result was consistent with that of IL-6 expression in human
disease-free liver tissues. We also investigated whether STAT3
protein expression was increased under H/R conditions by conducting
subsequent western blot analysis, which confirmed that the STAT3
level was also higher at all time points. Additionally, STAT3
expression was higher in the LO2C cells compared with the LO2G
cells (Fig. 5D). The result was
confirmed by densitometry (integral optical density, IOD) to
measure the STAT3 levels from H/R1 to H/R24 (P<0.05).
Discussion
Hepatic IRI caused by H/R is a common pathological
phenomenon during liver surgery, liver transplantation and severe
liver trauma (1–4). The simultaneous activation and
interaction of multiple cytokines, particularly multifunctional
IL-6, regulate cell injury and death during liver regeneration.
IL-6 plays an essential role in triggering liver
regeneration during the priming phase (24). Furthermore, protective
IL-6/STAT3 signaling is initiated within minutes to
hours after partial liver hepatectomy (4,25).
Recently, the IL-6 rs1800796 locus SNPs were thoroughly
investigated in a healthy population (13), and under several clinical
conditions (26–28). It has been reported that only
donor IL-6 rs1800796 SNPs affect the metabolism of
tacrolimus, mainly by influencing liver function and regeneration
following liver transplantation (16). Therefore, in this study, we
examined IL-6 expression in 84 disease-free liver tissues using
TMA. The IL-6 level was higher in genotype CC carriers than in
genotype CG/GG carriers, due to a higher intermediate expression
rate (P=0.006) (Fig. 2A and B and
Table II). Subsequently, it was
verified at the mRNA level (Fig.
2C). Therefore, allele C promotes IL-6 transcription
efficiency, which is consistent with a study involving basic
promoter experiments using a reporter gene system (18). As shown in Fig. 2A and B, the IL-6 was mainly
expressed in the cytoplasm of hepatocytes. It has been reported
that approximately 70% of liver cells are hepatocytes (29). Furthermore, studies have suggested
that primary rodent and LO2 hepatocytes synthesize and secrete IL-6
(9,30,31). Therefore, the biological
mechanisms of IL-6 and its rs1800796 locus SNPs, in human
hepatocytes, warrant further investigation.
The immortalized human normal liver cell line, LO2,
was derived from primary normal human hepatocytes, and still
retains the biological characteristics of normal adult hepatocytes
(17). Additionally, the current
third-generation lentivirus used in this study, proved that human
immunodeficiency virus-based vectors are valid manufacturing tools
for stable genetic modification (32,33). The LO2G cells and LO2C cells
successfully generated via lentivirus-mediated method stably
produced ectopic IL-6 overexpression and approximately 20% of
endogenic IL-6. The LO2G and LO2C cells, respectively, carried
allele G or C at the IL-6 rs1800796 locus. The NC-LO2 cells,
as the negative control group, were also successfully constructed.
Furthermore, cell morphology was not altered significantly
following double lentivirus transfection (Fig. 3B and C). Subsequently, based on
the LO2C cells, the LO2G cells, normal LO2 cells (blank control
group) and NC-LO2 cells, a cell model of H/R was successfully
established in order to examine H/R injury (19,20).
In this study, the concentrations of AST were higher
in the LO2C cells than in the LO2G cells from H/R6h to H/R24h
(P<0.05) (Fig. 4A). Similarly,
there were also marked differences between the LO2C and LO2G cells
as regards cell viability from H/R6h to H/R24h (P<0.05)
(Fig. 4B). The differences in the
AST data or cell viability between the experimental and the control
groups were significant at all H/R time-points (P<0.01)
(Fig. 4). Interestingly, these
data did not differ between the control LO2 and NC-LO2 cells, which
suggested that the lentiviral particles exerted less prominent
effects on the biological properties of LO2 cells. Therefore, the
differences between LO2C and LO2G cells were mainly attributed to
allele G mutating into C at the rs1800796 locus.
The relative increase in IL-6 expression in the LO2C
cells was higher than that in the LO2G cells from H/R1h to H/R12h
(Fig. 5A). LO2C cells expressed
greater IL-6 than LO2G cells in the normoxia group (P<0.001)
(Fig. 5C). The expression of
STAT3, downstream signal molecules of IL-6, in LO2C cells increased
higher than in LO2G cells, from H/R6h to H/R24h (P<0.05)
(Fig. 5B). Similarly, it was also
found that the STAT3 level was higher at all time-points under H/R
conditions. This phenomenon was more obvious in LO2C cells compared
with LO2G cells (Fig. 5D) and
confirmed by grey level of STAT3 from H/R1 to H/R24
(P<0.05).
The results suggest that LO2 cells synthesized IL-6,
and the subsequent IL-6/STAT3 signaling pathway was triggered
within minutes to hours following H/R injury. IL-6 exerted
hepatoprotective effects and decreased cell activity through the
upregulation of IL-6/STAT3 signaling. Although IL-6 production by
non-parenchymal cells, such as Kupffer cells may play a role
(8,9), these findings suggested that IL-6
produced in hepatocytes promoted the activation of the IL-6/STAT3
signaling pathway and also exerted hepatoprotective effects in an
autocrine manner.
It was observed that IL-6 rs1800796 SNPs
affected IL-6 transcription efficiency under normoxic or H/R
conditions. Furthermore, the allele G→C substitution accelerated
the production of IL-6 and attenuated H/R injury in hepatocytes.
These SNPs were present in the IL-6 promoter region, and
altered the affinity with the regulatory protein, and subsequently
influenced IL-6 transcription (15). For example, SNPs in the
osteopontin promoter region have been shown to alter DNA binding
affinity to transcription factors SP1/SP3 (34).
The present study has several limitations. First,
although the use of TMAs enables the analysis of 84 samples in a
relatively short period of time, the results may be unreliable due
to the small sample size. Thus, these results need to be confirmed
with a larger population. Second, in vitro studies using the
LO2 cell line, but not adult hepatocytes, may be confounded by the
cell type and culture conditions. Therefore, further in vivo
studies are warranted in order to elucidate the function of
IL-6 gene and SNPs. Third, this study mainly investigated
the changes in STAT3 and IL-6 RNA or protein levels. Although many
studies have fully established the association between IL-6 protein
and mRNA expression, its functional significance requires further
analysis. Nevertheless, our results provide insight into the
potential role of IL-6 and its SNPs in human
hepatocytes.
In conclusion, this study, to the best of our
knowledge, was the first of its kind to demonstrate that sequence
variants of IL-6 rs1800796 locus SNPs (G→C) increase the
transcription efficiency of the IL-6 gene and protect
hepatocytes following H/R. H/R injury triggered the synthesis of
IL-6 and STAT3 in the liver parenchymal cells. Our results
facilitate efficient liver regeneration and restoration following
liver surgery. Additionally, these data establish the framework for
exploring the role of the IL-6 gene and its polymorphisms in
other H/R conditions, such as those affecting the brain and
kidneys.
Acknowledgments
The authors of this study would like to thank Lijuan
Feng (GenomeDitech Co., Ltd., Shanghai, China) for her experimental
and technical advice. This study was supported by the National
Natural Science Foundation of China (grant no. 81370579) and the
Leading Project of Science, Technology Commission of Shanghai
Municipality (grant no. 134119a6300).
References
|
1
|
Serracino-Inglott F, Habib NA and Mathie
RT: Hepatic ischemia-reperfusion injury. Am J Surg. 181:160–166.
2001. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Klune JR and Tsung A: Molecular biology of
liver ischemia/reperfusion injury: Established mechanisms and
recent advancements. Surg Clin North Am. 90:665–677. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Hiranuma S, Ito K, Noda Y, Ozasa H, Koike
Y and Horikawa S: Amelioration of hepatic ischemia/reperfusion
injury in the remnant liver after partial hepatectomy in rats. J
Gastroenterol Hepatol. 22:2167–2172. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Taki-Eldin A, Zhou L, Xie HY and Zheng SS:
Liver regeneration after liver transplantation. Eur Surg Res.
48:139–153. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Streetz KL, Luedde T, Manns MP and
Trautwein C: Interleukin 6 and liver regeneration. Gut. 47:309–312.
2000. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Tiberio L, Tiberio GA, Bardella L, Cervi
E, Cerea K, Dreano M, Garotta G, Fra A, Montani N, Ferrari-Bravo A,
et al: Mechanisms of interleukin-6 protection against
ischemia-reperfusion injury in rat liver. Cytokine. 34:131–142.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
da Silva CG, Studer P, Skroch M, Mahiou J,
Minussi DC, Peterson CR, Wilson SW, Patel VI, Ma A, Csizmadia E and
Ferran: A20 promotes liver regeneration by decreasing SOCS3
expression to enhance IL-6/STAT3 proliferative signals. Hepatology.
57:2014–2025. 2013. View Article : Google Scholar :
|
|
8
|
Selzner N, Selzner M, Odermatt B, Tian Y,
Van Rooijen N and Clavien PA: ICAM-1 triggers liver regeneration
through leukocyte recruitment and Kupffer cell-dependent release of
TNF-alpha/IL-6 in mice. Gastroenterology. 124:692–700. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Yoshiya S, Shirabe K, Imai D, Toshima T,
Yamashita Y, Ikegami T, Okano S, Yoshizumi T, Kawanaka H and
Maehara Y: Blockade of the apelin-APJ system promotes mouse liver
regeneration by activating Kupffer cells after partial hepatectomy.
J Gastroenterol. 50:573–582. 2015. View Article : Google Scholar
|
|
10
|
Kämäräinen OP, Solovieva S, Vehmas T,
Luoma K, Riihimäki H, Ala-Kokko L, Männikkö M and Leino-Arjas P:
Common interleukin-6 promoter variants associate with the more
severe forms of distal interphalangeal osteoarthritis. Arthritis
Res Ther. 10:R212008. View
Article : Google Scholar : PubMed/NCBI
|
|
11
|
Li J, Song J, Jiang MH, Zheng JG, Gao SP,
Zhu JH and Pan M: Interleukin-6 promoter polymorphisms and
susceptibility to atrial fibrillation in elderly Han Chinese
patients with essential hypertension. J Interferon Cytokine Res.
32:542–547. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Gao SP, Liang S, Pan M, Sun RL, Chen C,
Luan H and Jiang MH: Interleukin-6 genotypes and serum levels in
Chinese Hui population. Int J Clin Exp Med. 7:2851–2857.
2014.PubMed/NCBI
|
|
13
|
Pan M, Gao SP, Jiang MH, Guo J, Zheng JG
and Zhu JH: Interleukin 6 promoter polymorphisms in normal Han
Chinese population: Frequencies and effects on inflammatory
markers. J Investig Med. 59:272–276. 2011. View Article : Google Scholar
|
|
14
|
Cheung BM, Ong KL, Tso AW, Leung RY,
Cherny SS, Sham PC, Thomas GN, Lam TH and Lam KS; Investigators of
the Hong Kong Cardiovascular Risk Factor Prevalence Study:
Relationship of plasma interleukin-6 and its genetic variants with
hypertension in Hong Kong Chinese. Am J Hypertens. 24:1331–1337.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Liao PY and Lee KH: From SNPs to
functional polymorphism: The insight into biotechnology
applications. Biochem Eng J. 49:149–158. 2010. View Article : Google Scholar
|
|
16
|
Chen D, Fan J, Guo F, Qin S, Wang Z and
Peng Z: Novel single nucleotide polymorphisms in interleukin 6
affect tacrolimus metabolism in liver transplant patients. PLoS
One. 8:e734052013. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Cheng B, Zheng Y, Guo X, Wang Y and Liu C:
Hepatitis B viral X protein alters the biological features and
expressions of DNA repair enzymes in LO2 cells. Liver Int.
30:319–326. 2010. View Article : Google Scholar
|
|
18
|
Chen J, Liu RY, Yang L, Zhao J, Zhao X, Lu
D, Yi N, Han B, Chen XF, Zhang K, et al: A two-SNP IL-6 promoter
haplotype is associated with increased lung cancer risk. J Cancer
Res Clin Oncol. 139:231–242. 2013. View Article : Google Scholar
|
|
19
|
Carini R, De Cesaris MG, Splendore R,
Bagnati M and Albano E: Ischemic preconditioning reduces Na(+)
accumulation and cell killing in isolated rat hepatocytes exposed
to hypoxia. Hepatology. 31:166–172. 2000. View Article : Google Scholar
|
|
20
|
Bhogal RH, Weston CJ, Curbishley SM, Bhatt
AN, Adams DH and Afford SC: Variable responses of small and large
human hepatocytes to hypoxia and hypoxia/reoxygenation (H–R). FEBS
Lett. 585:935–941. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Gabriel S, Ziaugra L and Tabbaa D: SNP
genotyping using the Sequenom MassARRAY iPLEX platform. Curr Protoc
Hum Genet. Chapter 2: Unit 2.12. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Li D, Yan D, Tang H, Zhou C, Fan J, Li S,
Wang X, Xia J, Huang F, Qiu G and Peng Z: IMP3 is a novel
prognostic marker that correlates with colon cancer progression and
pathogenesis. Ann Surg Oncol. 16:3499–3506. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Yan DW, Li DW, Yang YX, Xia J, Wang XL,
Zhou CZ, Fan JW, Wen YG, Sun HC, Wang Q, et al: Ubiquitin D is
correlated with colon cancer progression and predicts recurrence
for stage II–III disease after curative surgery. Br J Cancer.
103:961–969. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Cressman DE, Greenbaum LE, DeAngelis RA,
Ciliberto G, Furth EE, Poli V and Taub R: Liver failure and
defective hepatocyte regeneration in interleukin-6-deficient mice.
Science. 274:1379–1383. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Jin LM, Jin SF, Liu YX, Zhou L, Xie HY,
Yan S, Xu X and Zheng SS: Ischemic preconditioning enhances
hepatocyte proliferation in the early phase after ischemia under
hemi-hepatectomy in rats. Hepatobiliary Pancreat Dis Int.
11:521–526. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Motoyama S, Nakatsu T, Miura M, Hinai Y,
Minamiya Y and Ogawa J: Interleukin-6 -634G>C genetic
polymorphism is associated with prognosis following surgery for
advanced thoracic esophageal squamous cell carcinoma. Dig Surg.
29:194–201. 2012. View Article : Google Scholar
|
|
27
|
Oishi Y, Watanabe Y, Shinoda S, Naka M,
Ozawa Y, Matsuyama T, Morozumi K and Fuke Y: The IL-6 gene
polymorphism -634C>G and IL17F gene polymorphism 7488T>C
influence bone mineral density in young and elderly Japanese women.
Gene. 504:75–83. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Zhang G, Zhou B, Wang W, Zhang M, Zhao Y,
Wang Z, Yang L, Zhai J, Feng CG, Wang J and Chen X: A functional
single-nucleotide polymorphism in the promoter of the gene encoding
interleukin 6 is associated with susceptibility to tuberculosis. J
Infect Dis. 205:1697–1704. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Weibel ER, Stäubli W, Gnägi HR and Hess
FA: Correlated morphometric and biochemical studies on the liver
cell. I. Morphometric model, stereologic methods, and normal
morphometric data for rat liver. J Cell Biol. 42:68–91. 1969.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Xiang WQ, Feng WF, Ke W, Sun Z, Chen Z and
Liu W: Hepatitis B virus X protein stimulates IL-6 expression in
hepatocytes via a MyD88-dependent pathway. J Hepatol. 54:26–33.
2011. View Article : Google Scholar
|
|
31
|
Norris CA, He M, Kang LI, Ding MQ, Radder
JE, Haynes MM, Yang Y, Paranjpe S, Bowen WC, Orr A, et al:
Synthesis of IL-6 by hepatocytes is a normal response to common
hepatic stimuli. PLoS One. 9:e960532014. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Lee CL, Chou M, Dai B, Xiao L and Wang P:
Construction of stable producer cells to make high-titer lentiviral
vectors for dendritic cell-based vaccination. Biotechnol Bioeng.
109:1551–1560. 2012. View Article : Google Scholar :
|
|
33
|
Li M, Husic N, Lin Y and Snider BJ:
Production of lentiviral vectors for transducing cells from the
central nervous system. J Vis Exp. 24:e40312012.
|
|
34
|
Giacopelli F, Marciano R, Pistorio A,
Catarsi P, Canini S, Karsenty G and Ravazzolo R: Polymorphisms in
the osteopontin promoter affect its transcriptional activity.
Physiol Genomics. 20:87–96. 2004. View Article : Google Scholar : PubMed/NCBI
|