Introduction
Gamma-aminobutyric acid (GABA) acts mainly as an
inhibitory neurotransmitter at GABA type A (GABA A) receptors to
mediate the synaptic inhibition of neuronal activity in the
mammalian brain (1). The GABA A
receptor is a heteropentamer, consisting of five subunits that form
a chloride ion channel. In addition to being located on central
neurons, functional GABA A receptors, particularly the GABA A
receptor π subunit (GABRP), have also been found in peripheral
tissues, such as the uterus, prostate, ovaries, placenta, gall
bladder, lung and small intestine (2,3).
It has been reported that the assembly of GABRP into a recombinant
GABA A receptor may change the sensitivity of the receptor to
modulatory agents such as GABA and allopregnanolone (4). In addition, due to its upregulation
in the human endometrium during the window of implantation, GABRP
may play an important role in the generation of a receptive
endometrium (5). In our previous
study, GABRP was detected in both human and mouse placenta;
however, the precise function of GABRP in trophoblastic cells
during placentation remains poorly defined (7).
In the present study, we report that GABRP
overexpression may result in the decreased invasion of HTR-8/SVneo
cells, possibly induced by elevated apoptosis. It has been
suggested that the elevated apoptosis of cytotrophoblasts (CTBs) in
preeclampsia (PE) contributes to the placental dysfunction, which
is characteristic of this disorder (6). Given the role of GABRP in enhancing
the apoptosis of HTR-8/SVneo trophoblastic cells, we hypothesized
that there may be a correlation between GABRP expression and PE. As
expected, the results of reverse transcription-quantitative
polymerase chain reaction (RT-qPCR) and immunohistochemical
analysis revealed that the expression of GABRP was increased in
placental tissues from patients with PE compared with that in the
normal controls, which may contribute to the shallow trophoblastic
invasion and hence, result in the onset of PE. In our laboratory,
we have previously demonstrated that the GABRP protein level was
decreased in cultured mouse stromal cells during the
decidualization process, and that GABA suppressed this procress
through GABA A receptors, which suggests that GABRP impaired the
remodeling of the decidua (7).
Based on these results, we concluded that GABRP may adversely
affect placental function complicated with PE through intrinsic
(trophoblastic) factors in combination with extrinsic (maternal
uterine) factors. Further studies are warranted to determine
whether GABRP has the potential to be used as a biomarker or a
therapeutic molecular target for PE.
Materials and methods
Human placental tissue samples
Six placentas from patients with PE and six
placentas from normal pregnancies were obtained from patients at
the First Affiliated Hospital of Chongqing Medical University
(Chongqing, China). The criteria for the diagnosis of severe PE
were strictly based on those of the American Congress of
Obstetricians and Gynecologists (2002). The study was approved by
the Ethics Committee of Chongqing Medical University (Chongqing,
China) and informed consent was obtained from all patients. The
tissue samples were fixed in buffered formalin (4%, m/v) and
embedded in paraffin according to standard procedures.
Immunohistochemistry (IHC) and
immumocytochemistry (ICC)
IHC was performed with a
biotin-streptavidin-peroxidase (SP; Zhongshan Goldenbridge,
Beijing, China) and 3,3′-diaminobenzidine (DAB; Zhongshan
Goldenbridge). The sections (5 mm) were deparaffinized and
rehydrated in xylene and ethanol gradients. The slides were boiled
in citrate buffer (10 mM citrate sodium, 10 mM citric acid, pH 6.0)
in a microwave oven at 92–98°C for 15 min for antigen retrieval.
The sections were then sequentially incubated with 3%
H2O2 in methanol for 10 min to quench
endogenous peroxidase, and in normal goat serum as the blocking
solution for 20 min. The sections were incubated with primary
antibodies against GABRP (ab26055; Abcam, Cambridge, UK) or rabbit
IgG isotypic control at 4°C overnight and biotinylated secondary
antibody. The sections were then incubated in HRP-streptavidin
(Zhongshan Goldenbridge) for 30 min at room temperature and then
detected using the chromogenic peroxidase substrate, DAB, and
counterstained with hematoxylin.
For ICC staining, the HTR-8/SVneo trophoblastic
cells (obtained from Professor H. Wang, Institute of Zoology,
Chinese Academy of Sciences, Beijing, China) were cultured on
coverslips for 48 h. The coverslips were fixed in 4%
paraformaldehyde for 20 min at room temperature, washed in
phosphate-buffered saline (PBS) and permeabilized for 10 min with
0.25% Triton-100 in PBS. The cells were then incubated with 1%
bovine serum albumin (BSA) in PBS/Tween 20 (PBST) for 30 min to
block the non-specific binding of the antibodies. The cells were
incubated with primary antibody or rabbit IgG isotype control
overnight at 4°C, and then incubated with a peroxidase-conjugated
secondary antibody for 60 min at 37°C. The slides were stained with
DAB and counterstained with hematoxylin.
Cell culture and treatment
An immortalized, first trimester extravillous
trophoblast (EVT) cell line, HTR-8/SVneo, which was derived from a
short-lived, primary EVT cell line, was used in the present study.
The HTR-8/SVneo cells were maintained in RPMI-1640 medium,
supplemented with 10% heat-inactivated fetal bovine serum (FBS;
Gibco-BRL, Carlsbad, CA, USA), 100 U/ml penicillin (Beyotime
Biotech, Jiangsu, China) and 100 µg/ml streptomycin
(Beyotime Biotech) under standard culture conditions (37°C in a 5%
humidified CO2 incubator). The cells were transiently
transfected with full-length GABRP after being seeded onto 35-mm
dishes overnight. In order to exogenously express GABRP, the GABRP
sequence was sub-cloned into the pIRES2-EGFP plasmid vector. After
the GABRP sequence was inserted into the vector, sequencing
analysis was conducted to ensure that this vector was capable of
specifically expressing GABRP (constructed by Invitrogen, Carlsbad,
CA, USA).
Matrigel invasion assay
An invasion assay was performed in Matrigel-coated
(BD Biosciences, Bedford, MA, USA) Transwell inserts (6.5 mm;
Costar, Cambridge, UK) containing polycarbonate filters with a pore
size of 8 µm. Briefly, the inserts were pre-coated with 100
µl Matrigel matrix (1 mg/ml) at 37°C for 4 h for gelling.
The HTR-8/SVneo cells (1×105 cells) in 200 µl
serum-free medium were plated in the upper chamber, whereas medium
with 10% FBS was added to the lower well. After incubating for 24
h, the cells on the Matrigel side of the insert were scraped by a
cotton swab. The inserts were then fixed in methanol for 10 min at
room temperature and stained with hematoxylin and eosin (H&E;
Zhongshan Goldenbridge). The cells which had invaded to the other
side of the insert were counted under a light microscope (IX51;
Olympus, Tokyo, Japan) in ten random fields at magnification of
x200. The assay was repeated three times, and the results are
represented as the percentage means of invasion ± standard
deviation (SD) compared with the control.
Western blot analysis
The HTR-8/SVneo cells transfected with
overexpressing plasmid groups were lysed for protein extraction
using RIPA supplemented with phenylmethylsulfonyl fluoride (PMSF)
(both from Beyotime Biotech). The Bradford assay (Beyotime Biotech)
was used to determine the protein concentration of each sample.
Samples containing 50–100 µg of extracted protein were
separated by 10% sodium dodecyl sulfate-polyacrylamide gel
electrophoresis (SDS-PAGE). The separated proteins were transferred
to 0.22 µm nitrocellulose membranes and were incubated with
primary antibodies against GABRP (ab26055; Abcam); caspase-3
(19677-1-AP; ProteinTech Group, Wuhan, China); cleaved caspase-3
(Asp175; Cell Signaling Technology, Inc., Danvers, MA, USA); Bcl-2
(AP13823c), Bad (AP1314c) and Bax (AP18517a) (all from Abgent,
Inc., San Diego, CA, USA); and GAPDH (AP0063; Bioworld Technology,
Inc., St. Louis Park, MN, USA) at 1:1,000 dilution. The secondary
antibody was goat anti-rabbit IgG (1:1,000; Zhongshan
Goldenbridge). All experiments were repeated at least three
times.
Flow cytometry (FCM)
The HTR-8/SVneo cells were transiently transfected
with overexpressing (pIRES2-EGFP and pIRES2-GABRP-EGFP) plasmid
groups and after 72 h, they were harvested using trypsin without
EDTA, washed with PBS, resuspended in 1 ml binding buffer, and
stained for 15 min with fluorescein isothiocyanate (FITC)-Annexin V
and propidium iodide (PI) in the dark at room temperature,
according to the manufacturer's instructions. Cell analysis was
performed by a flow cytometer (FACScan) equipped with CellQuest
software (both from BD Biosciences). The cells were sorted into
living, necrotic, early apoptotic and late apoptotic cells. The
relative ratio of early and late apoptotic cells was calculated for
further comparison. This assay was repeated at least three
times.
RT-qPCR
Total RNA extraction from the placental tissues and
the HTR-8/SVneo cells was performed using TRIzol reagent
(Invitrogen) according to the manufacturer's instructions, and
quantities of RNA were determined using a spectrophotometer (Thermo
Fisher Scientific, Inc., Waltham, MA, USA) at 260 nm. Two
micrograms of total RNA was reverse transcribed using SuperScript
III Reverse Transcriptase (Invitrogen).
qPCR was performed using SYBR Premix Ex Taq II
(RR820A; Takara Bio, Inc., Kusatsu, Japan) in a CFX96™ Real-Time
PCR detection system (Bio-Rad, Berkeley, CA, USA) and analyzed
using the ΔΔCt method according to the following equation: ΔCt
(test) = Ct (target, test) − Ct (ref, test); ΔCt (calibrator) = Ct
(target, calibrator) − Ct (ref, calibrator); ΔΔCt = ΔCt (test) −
ΔCt (calibrator); 2−ΔΔCt = relative quantitative value.
The following PCR primers were used: GABRP (NM_014211.2) sense,
5′-CGACCGTGTTATCAATGACC-3′ and antisense,
5′-CCCCAAACACAAAGCTAAAGCA-3′; Bcl-2 (NM_000657.2) sense,
5′-TTGTTCAAACGGGATTCACA-3′ and antisense,
5′-GAGCAAGTGCAGCCACAATA-3′; Bax (NM_001291428.1) sense,
5′-GCTGGACATTGGACTTCCTC-3′ and antisense,
5′-CTCAGCCCATCTTCTTCCAG-3′; Bad (NM_004322.3) sense,
5′-CCTCAGGCCTATGCAAAA-3′ and antisense, 5′-AAACCCAAAACTTCCGATGG-3′;
and GAPDH (NM_002046.5) sense 5′-AGCCACATCGCTCAGACAC-3′ and
antisense, 5′-TGGACTCCACGACGTACTC-3′. This assay was repeated at
least three times.
Cell viability assay
A Cell Counting Kit-8 (CCK-8; Beyotime Biotech) was
employed in this experiment to quantitatively evaluate cell
viability. Following 24, 48 and 72 h of plasmid transfection, a
CCK-8 assay was performed using the HTR-8/SVneo cells. Briefly, the
HTR-8/SVneo cells were seeded at 0.5×104/well in 96-well
plates. RPMI-1640 culture medium (100 µl/well) was added to
10 µl CCK-8 reagent after 20, 44 and 68 h and incubated at
37°C for a further 4 h, to form water-soluble formazan. The
absorbances at 450 and 630 nm (calibrated wave) were determined
using a microplate reader (Multiskan MK33; Thermo Fisher
Scientific, Inc.). RPMI-1640 medium containing 10% CCK-8 was used
as a control. The untreated control was set at 100%, and the
treated samples were normalized to this value according to the
following equation: survival rate (%)=optical density (OD) of the
treated cells - OD of blank control/OD of negative control - OD of
blank control x 100.
Statistical analysis
The bands from western blot analysis were quantified
using ImageJ2x software (http://imagej.net/ImageJ2). All data are presented as
the means ± SD and statistical analysis was performed using SPSS
software (SPSS Inc., Chicago, IL, USA). Furthermore, P<0.05 was
considered to indicate a statistically significant difference.
Results
GABRP expression in HTR-8/SVneo
cells
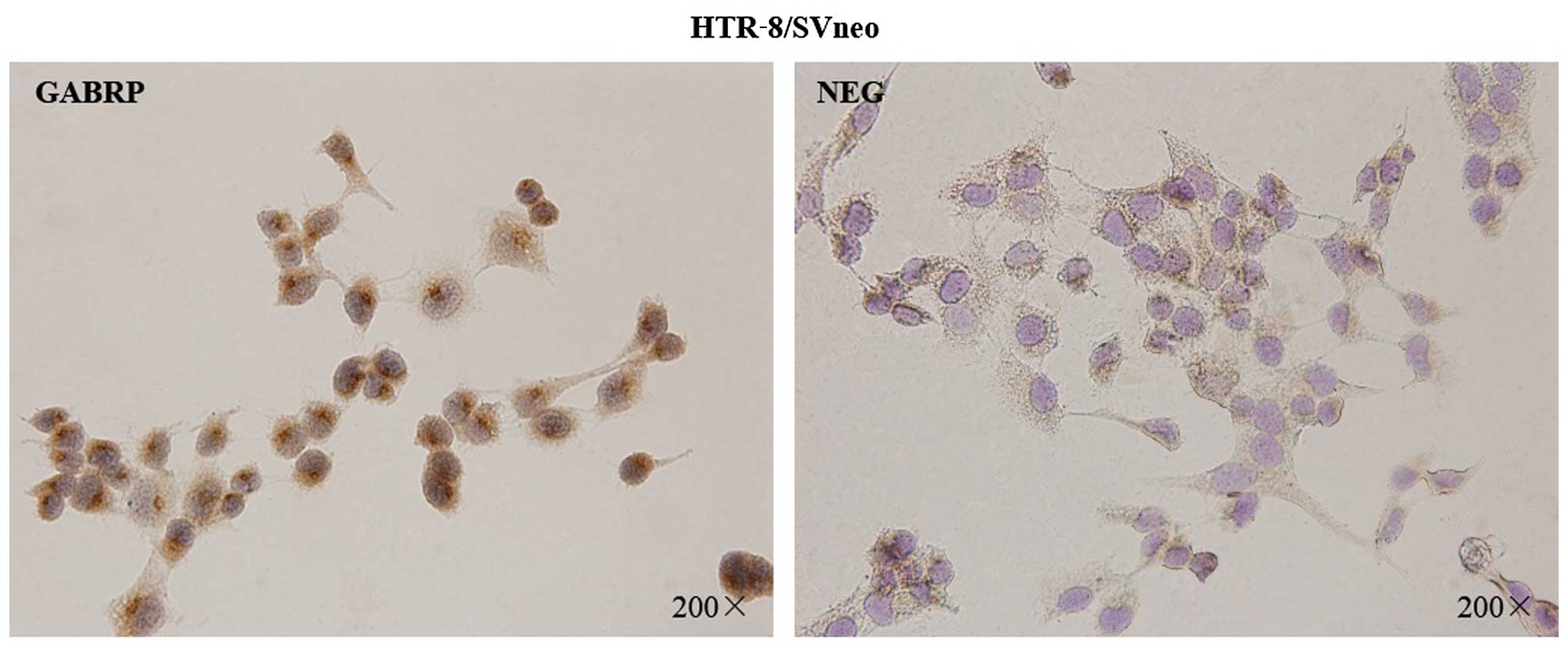
To assess the localization pattern of GABRP within
the cells, immunocytochemical analysis was performed on the
HTR-8/SVneo trophoblastic cells (Fig.
1). The presence of GABRP was identified by the specific
brown-colored staining of the cytoplasm and the nuclear membrane of
the HTR-8/SVneo cells. Notably, GABRP expression was mainly located
on the apical side of the nuclear membrane which showed particular
polarization to a certain extent.
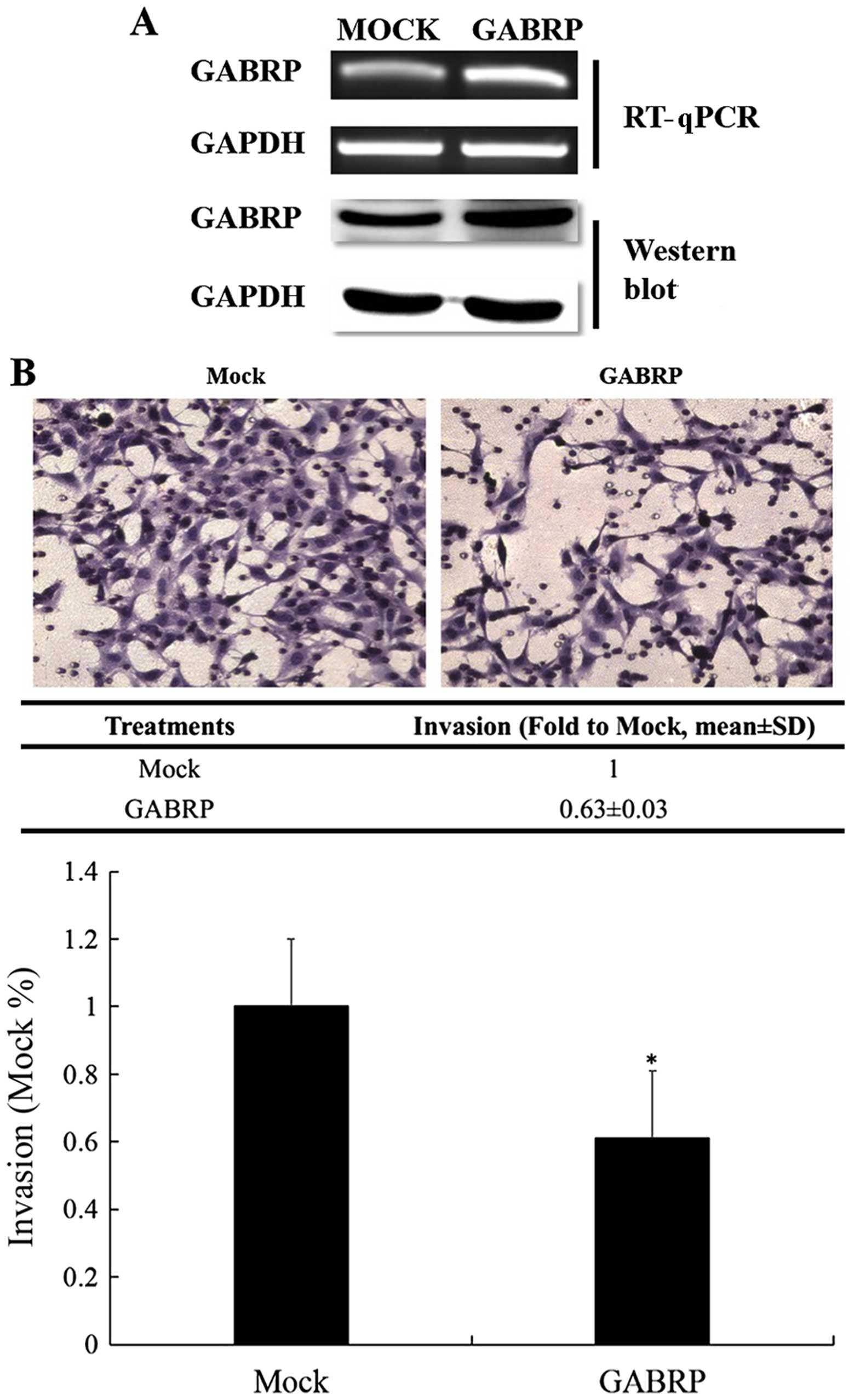
GABRP overexpression significantly
inhibits the invasion of HTR-8/SVneo cells
GABRP expression in the trophoblastic cell line may
play a role in controlling trophoblastic cell invasion. Thus, we
constructed a pIRES2-GABRP-EGFP plasmid and used Matrigel cell
invasion models in the HTR-8/SVneo cells to verify this assumption.
As shown in Fig. 2A, the
exogenous expression of GABRP was evaluated by RT-qPCR and western
blot analysis. The results demonstrated that the number of invading
HTR-8/SVneo cells overexpressing GABRP was decreased compared with
that in the control groups (P<0.05) (Fig. 2B).
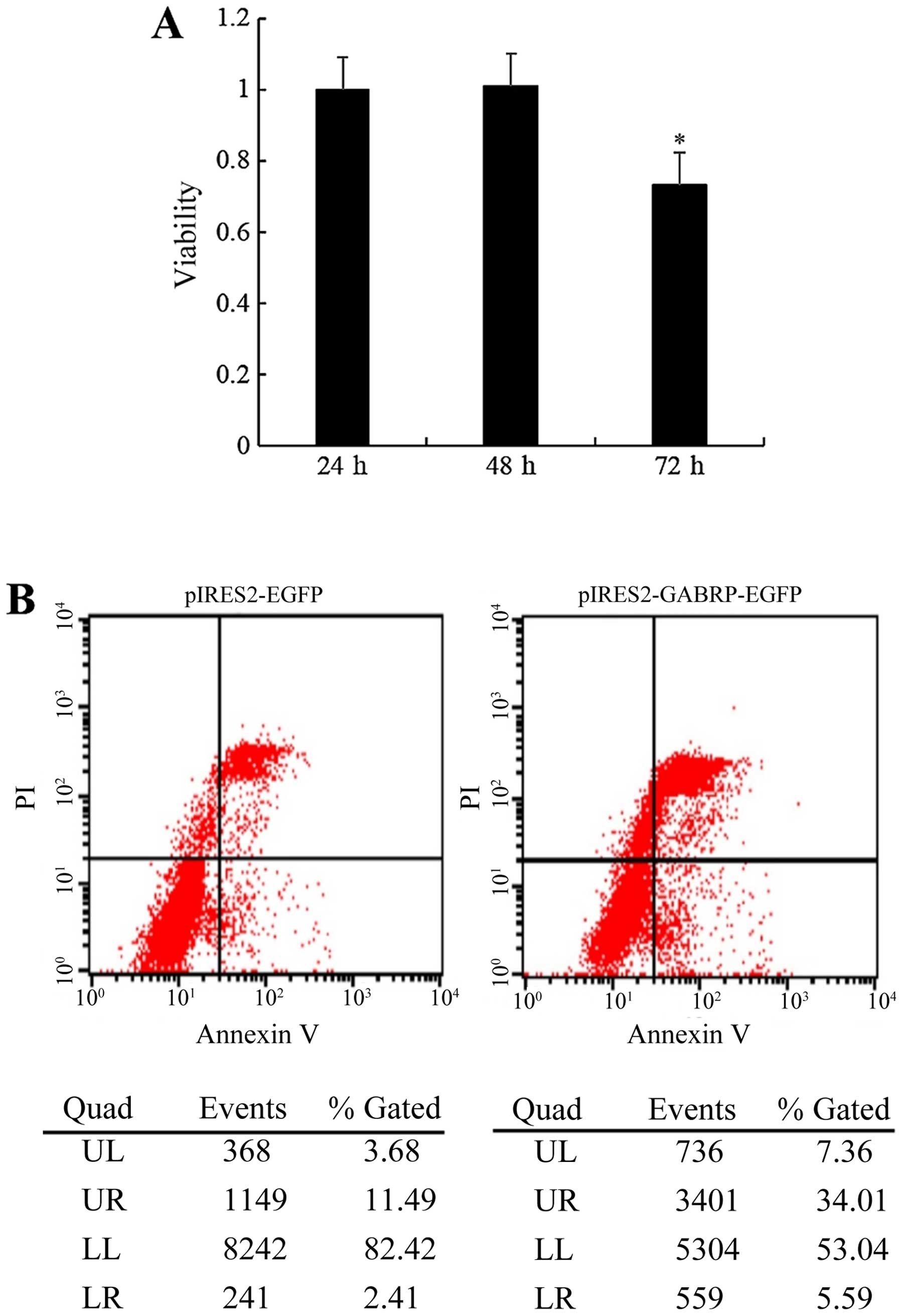
GABRP overexpression decreases cell
viability and promotes the apoptosis of HTR-8/SVneo cells
To determine whether the observed changes in
invasiveness were due to the effects of GABRP on cell apoptosis
and/or viability, we examined the effects of GABRP on the viability
and apoptosis of trophoblastic cells. The HTR-8/SVneo cells
transfected with the pIRES2-GABRP-EGFP plasmid were subjected to
the CCK-8 assay and FCM, respectively. Compared with the control
groups, the viability of the HTR-8/SVneo cells was decreased
(P<0.05) (Fig. 3A) and the
proportion of apoptotic HTR-8/SVneo cells was increased (Fig. 3B) by GABRP 72 h after
transfection.
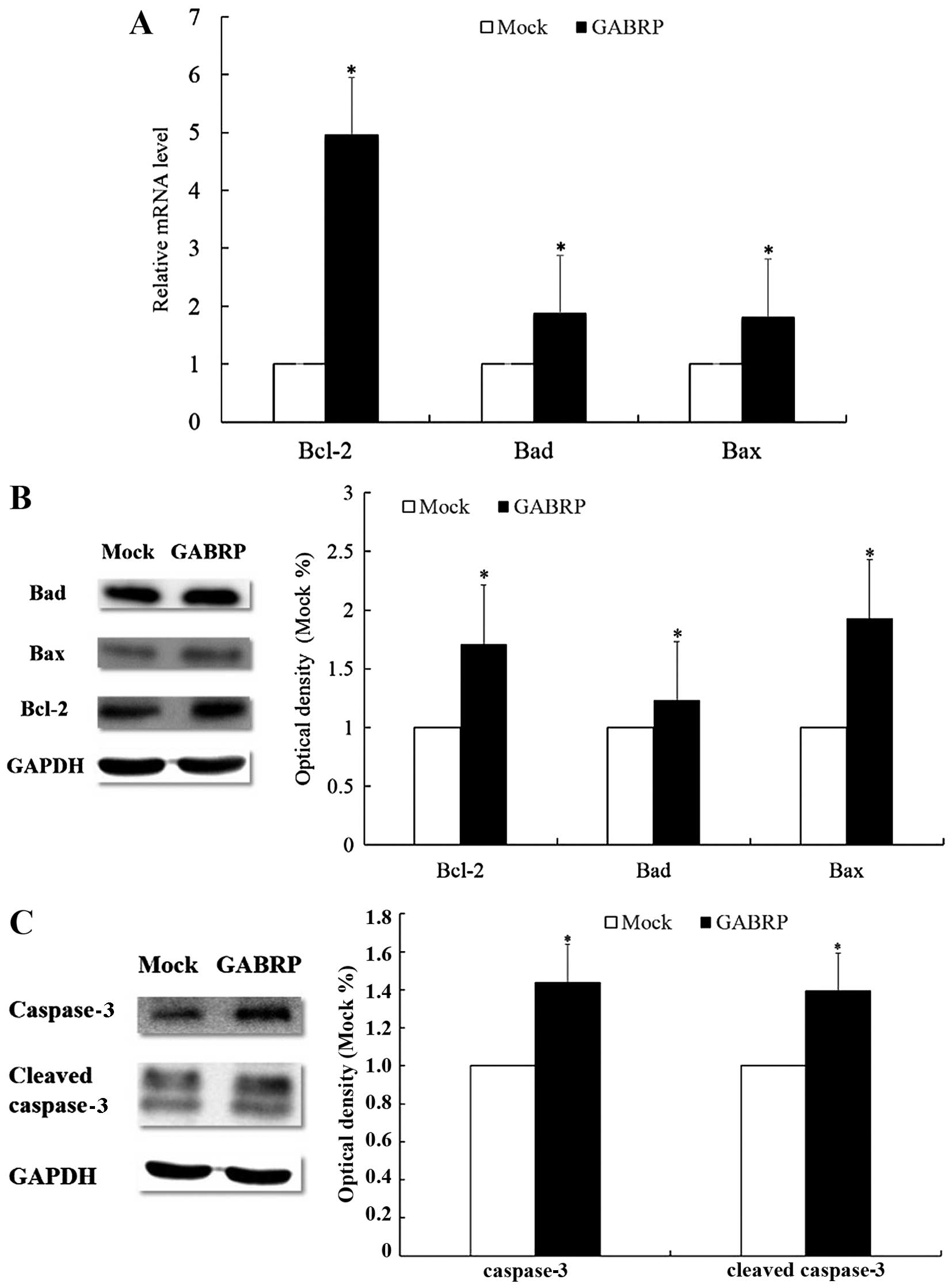
Members of the Bcl-2 family mediate
increased apoptosis of HTR-8/SVneo cells through GABRP
overexpression
As members of the Bcl-2 family are the key
regulators of apoptosis, we evaluated the expression of three
important members of the family, namely Bcl-2, Bad and Bax. The
mRNA levels of pro-apoptotic Bad and Bax were upregulated after
GABRP transfection (P<0.05) (Fig.
4A), which was consistent with the protein levels (P<0.05)
(Fig. 4B). Unexpectedly, the mRNA
and protein levels of anti-apoptotic Bcl-2 were also upregulated
(P<0.05) (Fig. 4A and B).
Further evaluation by western blot analysis revealed that GABRP
overexpression increased the protein expression of cleaved
caspase-3 (active form) and caspase-3 (inactive form) (P<0.05)
(Fig. 4C).
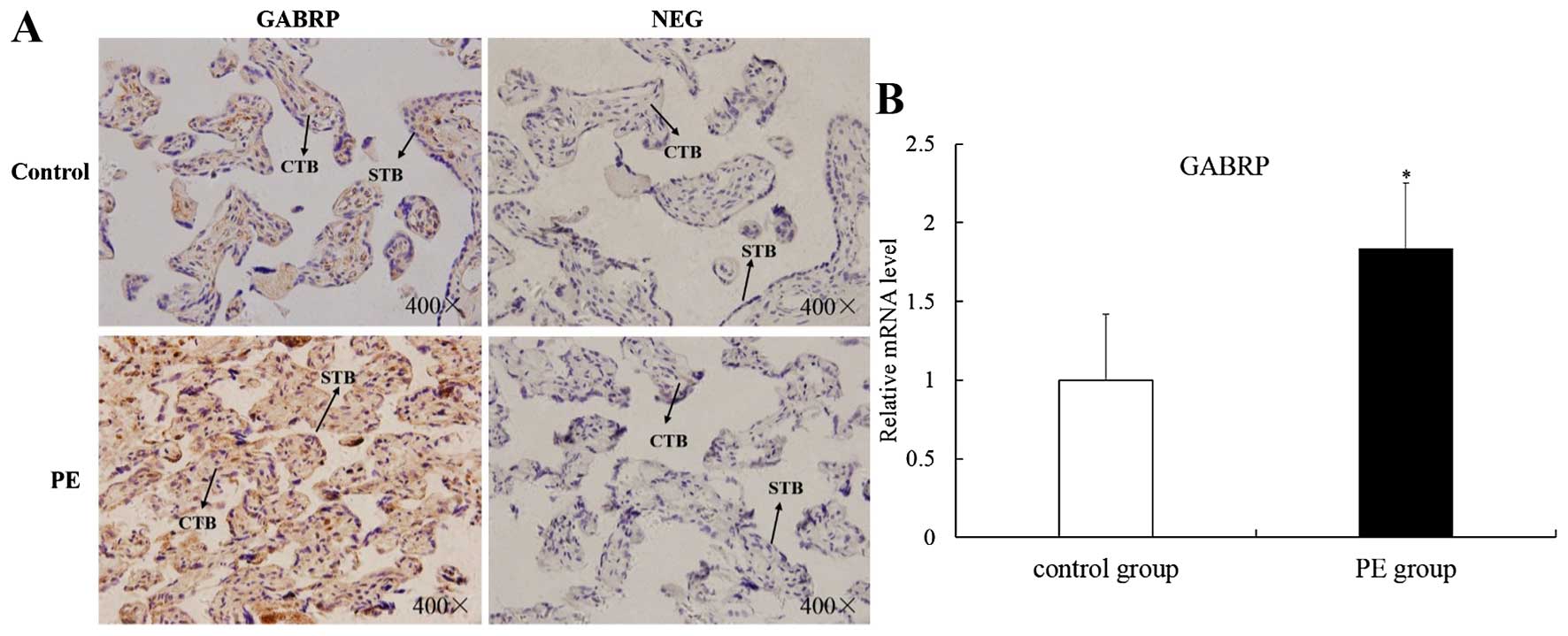
GABRP is upregulated in placental villi
from patients with PE
The apoptosis of villous trophoblasts is increased
in PE and it is generally believed that PE is associated with
impaired trophoblast invasion (8,9).
Thus, we hypothesized that GABRP may be involved in the onset of PE
and we collected placental villi from patients with PE and from
normal pregnancy controls of similar gestational stages for IHC
analysis and RT-qPCR. Immunoreactivity for GABRP was consistently
found in the villous syncytiotrophoblasts (STB) and CTBs of both
groups. However, GABRP was strongly expressed in the PE group
compared with the normal control group (Fig. 5A). Compared with the normal
control group, the mRNA expression of GABRP was upregulated in the
PE group (P<0.05) (Fig.
5B).
Discussion
Apoptosis is an essential feature of normal
placental development; however, it may also be involved in the
pathophysiology of pregnancy-related diseases, such as PE (10). Although the apoptosis of
trophoblastic cells increases in normal placentas with advancing
gestation, increased apoptosis of trophoblastic cells has been
observed in pregnancies complicated by PE (11,12). The precise etiology of PE remains
unclear; however, it may be associated with shallow trophoblastic
invasion or impaired decidualization. In our previous study, we
suggested that GABRP impaired remodeling of the decidua (7), and in the present study, we have
demonstrated that GABRP promotes apoptosis and hence, inhibits the
invasion of trophoblastic cells. Based on these findings, we
hypothesized that GABRP may be a novel candidate which explains the
association between the pathologies of the fetal placenta and
maternal decidua in PE, and GABRP may adversely affect placental
function through intrinsic (trophoblastic) factors in combination
with extrinsic (maternal uterine) factors.
As important regulators of apoptosis, members of the
Bcl-2 family may be divided into two functionally antagonistic
groups: i) apoptosis suppressors such as Bcl-2, Bcl-xL
and Bcl-w; and ii) apoptosis promoters such as Bax,
Bcl-xS, Bak and Bad (13–15). To identify the members of this
protein family that may regulate trophoblast apoptosis, we assessed
the expression of three important members of the Bcl-2 gene family
in GABRP-overexpressing HTR-8/SVneo cells. As expected, the mRNA
and protein expression of pro-apoptotic Bad and Bax was upregulated
following GABRP overexpression. Unexpectedly, the anti-apoptotic
Bcl-2 expression was also increased. It appears contradictory that
GABRP activates both suppressors and promoters of apoptosis during
the apoptosis cascade of HTR-8/SVneo cells. Even though it has been
demonstrated that Bcl-2 may inhibit further propagation of the
death signal at the mitochondrial level through mediating the
activity of initiator caspases and the activation of execution
caspases (16,17), the increase in Bcl-2 following
GABRP overexpression in the HTR-8/SVneo cells appears inadequate to
prevent the whole apoptosis cascade when the execution caspases,
such as caspase-3 and its active form, cleaved caspase-3, were also
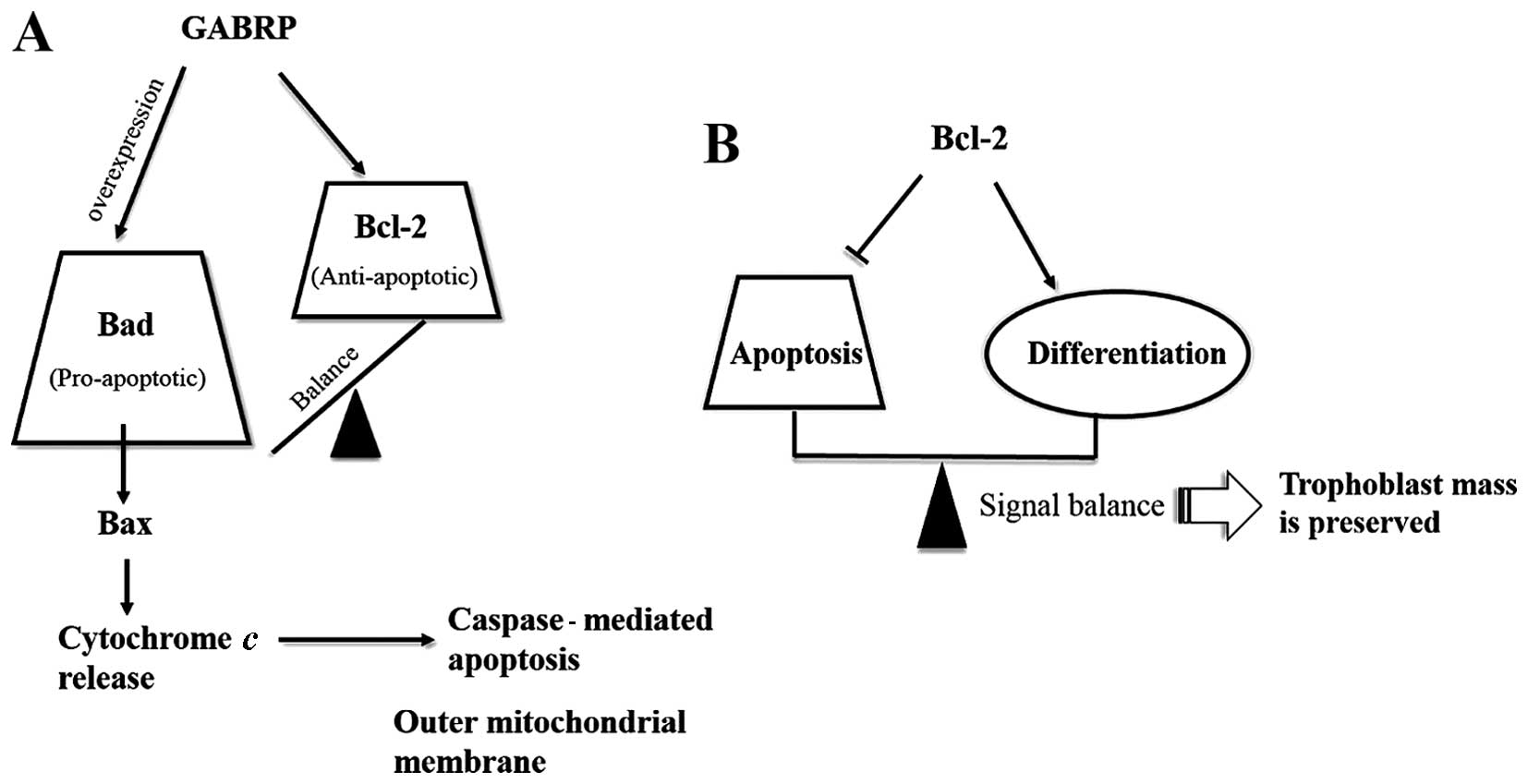
activated. Thus, we hypothesized that GABRP maintains a state of
equilibrium between anti-apoptotic Bcl-2 and pro-apoptotic Bad.
However, excessive GABRP may damage this balance and induce
apoptosis along the Bad pathway (Fig.
6A). In addition, as Bcl-2 expression is STB-specific during
pregnancy (18–22), it is tempting to speculate that
Bcl-2 is associated with the differentiation of trophoblasts. In
addition, the treatment of JEG-3 cells with 8-Br-cAMP, which
induces genes characteristic of STBs, was shown to raise Bcl-2
protein levels approximately 2-fold (23). Thus, we speculate that: i) there
is a differentiation-dependent pattern of Bcl-2 expression in the
placenta, with the protein being expressed in the STBs throughout
gestation and increased in JEG-3 choriocarcinoma cells following
treatment with the differentiation inducing reagent, 8-Br-cAMP; and
ii) Bcl-2 expression establishes a delicate balance between the
differentiation and apoptosis of trophoblasts and this may be one
mechanism through which trophoblast mass is preserved during
pregnancy (Fig. 6B). In fact,
there is evidence to suggest that syncytial fusion is directly or
indirectly associated with apoptotic events (24), and Bcl-2 is involved in the
retardation of the apoptosis cascade in the execution stages. In
the initiation stages of the apoptosis cascade, caspase-8 is
believed to be required for syncytial fusion (25,26). Nevertheless, the presumption that
GABRP may be involved in the differentiation of trophoblasts
through the upregulation of Bcl-2 merits further investigation.
In mammalian brains, the synaptic inhibition of
neuronal activity is principally mediated by GABA at GABA A
receptors, and functional GABA A receptors have also been detected
in non-neuronal cells (1). Among
16 known GABA A receptor subunits, GABRP mRNA was confirmed to be
highly expressed in the human uterus as well as in normal breast
tissue by RT-qPCR (2). In fact,
there have been many studies regarding the role of GABRP in uterine
and breast tissues. For example, GABRP mRNA expression and membrane
translocation were suggested to be regulated by HOXA10, which is
likely to be involved in initiating and maintaining a receptive
endometrium (5). Furthermore, the
dynamic expression of GABRP mRNA throughout pregnancy is suggested
to correlate with parturition (4). Despite the abundance of GABRP in
breast tissue, the role of GABRP in breast tissue remains
controversial according to the findings of different groups. For
example, Zafrakas et al postulated that GABRP may exert a
tumor-suppressing effects in breast tissue (2), whereas Sizemore et al
suggested that GABRP stimulates basal-like breast cancer cell
migration through the activation of extracellular-regulated kinase
1/2 (27) and the distinct
results may be due to the heterogeneity of breast cancer. In
addition to uterine and breast tissues, it has been demonstrated
that GABRP is overexpressed in pancreatic tumors (28) and promotes pancreatic cancer
growth through GABA stimulation (29).
Emerging evidence indicates that GABA acts as a
tumor signaling molecule in peripheral non-neuronal cells (30–32). Given the role of GABRP in the
uterus and some tumor tissues, it is important to consider the
potential function of GABRP in the trophoblastic cells of the
reproductive system, which share a similar capacity to migrate and
invade surrounding tissues with malignant cells (33–35). Herein, we have demonstrated that
GABRP promotes apoptosis and hence, inhibits the invasion of
trophoblastic cells, which may be associated with the onset of PE.
In addition, the upregulation of GABRP was detected in placental
tissues obtained from patients with PE compared with that in normal
control placental tissues which is consistent with our hypothesis.
This suggests that GABRP may possibly be used as a tissue marker
with diagnostic significance in PE. On the other hand, GABRP
overexpression may contribute to the progression of PE and GABRP
may be a promising molecular target for the development of new
therapeutic strategies for PE.
Acknowledgments
The present study was supported by grants from the
National Natural Science Foundation of China (no. 31171436).
References
|
1
|
Hedblom E and Kirkness EF: A novel class
of GABAA receptor subunit in tissues of the reproductive system. J
Biol Chem. 272:15346–15350. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Zafrakas M, Chorovicer M, Klaman I,
Kristiansen G, Wild PJ, Heindrichs U, Knüchel R and Dahl E:
Systematic characterisation of GABRP expression in sporadic breast
cancer and normal breast tissue. Int J Cancer. 118:1453–1459. 2006.
View Article : Google Scholar
|
|
3
|
Gladkevich A, Korf J, Hakobyan VP and
Melkonyan KV: The peripheral GABAergic system as a target in
endocrine disorders. 124:1–8. 2006.
|
|
4
|
Fujii E and Mellon SH: Regulation of
uterine gamma-aminobutyric acid(A) receptor subunit expression
throughout pregnancy. Endocrinology. 142:1770–1777. 2001.PubMed/NCBI
|
|
5
|
Sadeghi H and Taylor HS: HOXA10 regulates
endometrial GABAA (pi) receptor expression and membrane
translocation. Am J Physiol Endocrinol Metab. 298:E889–E893. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Longtine MS, Chen B, Odibo AO, Zhong Y and
Nelson DM: Villous trophoblast apoptosis is elevated and restricted
to cytotrophoblasts in pregnancies complicated by preeclampsia,
IUGR, or preeclampsia with IUGR. Placenta. 33:352–359. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Luo W, Liu Z, Tan D, Zhang Q, Peng H, Wang
Y and Tan Y: Gamma-amino butyric acid and the A-type receptor
suppress decidualization of mouse uterine stromal cells by
down-regulating cyclin D3. Mol Reprod Dev. 80:59–69. 2013.
View Article : Google Scholar
|
|
8
|
Allaire AD, Ballenger KA, Wells SR,
McMahon MJ and Lessey BA: Placental apoptosis in preeclampsia.
Obstet Gynecol. 96:271–276. 2000.PubMed/NCBI
|
|
9
|
Zhang Q, Chen Q, Lu X, Zhou Z, Zhang H,
Lin HY, Duan E, Zhu C, Tan Y and Wang H: CUL1 promotes trophoblast
cell invasion at the maternal-fetal interface. Cell Death Dis.
4:e5022013. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Sharp AN, Heazell AE, Crocker IP and Mor
G: Placental apoptosis in health and disease. Am J Reprod Immunol.
64:159–169. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Straszewski-Chavez SL, Abrahams VM and Mor
G: The role of apoptosis in the regulation of trophoblast survival
and differentiation during pregnancy. Endocr Rev. 26:877–897. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Crocker IP, Cooper S, Ong SC and Baker PN:
Differences in apoptotic susceptibility of cytotrophoblasts and
syncytiotrophoblasts in normal pregnancy to those complicated with
preeclampsia and intrauterine growth restriction. Am J Pathol.
162:637–643. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Marzioni D, Mühlhauser J, Crescimanno C,
Banita M, Pierleoni C and Castellucci M: BCL-2 expression in the
human placenta and its correlation with fibrin deposits. Hum
Reprod. 13:1717–1722. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Krajewski S, Krajewska M and Reed JC:
Immunohistochemical analysis of in vivo patterns of Bak expression,
a proapoptotic member of the Bcl-2 protein family. Cancer Res.
56:2849–2855. 1996.PubMed/NCBI
|
|
15
|
Cory S, Huang DC and Adams JM: The Bcl-2
family: roles in cell survival and oncogenesis. Oncogene.
22:8590–8607. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Huppertz B and Kingdom JC: Apoptosis in
the trophoblast - role of apoptosis in placental morphogenesis. J
Soc Gynecol Investig. 11:353–362. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Huppertz B, Kadyrov M and Kingdom JC:
Apoptosis and its role in the trophoblast. Am J Obstet Gynecol.
195:29–39. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Ratts VS, Tao XJ, Webster CB, Swanson PE,
Smith SD, Brownbill P, Krajewski S, Reed JC, Tilly JL and Nelson
DM: Expression of BCL-2, BAX and BAK in the trophoblast layer of
the term human placenta: a unique model of apoptosis within a
syncytium. Placenta. 21:361–366. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Murakoshi H, Matsuo H, Laoag-Fernandez JB,
Samoto T and Maruo T: Expression of Fas/Fas-ligand, Bcl-2 protein
and apoptosis in extravillous trophoblast along invasion to the
decidua in human term placenta. Endocr J. 50:199–207. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Ho S, Winkler-Lowen B, Morrish DW, Dakour
J, Li H and Guilbert LJ: The role of Bcl-2 expression in EGF
inhibition of TNF-alpha/IFN-gamma-induced villous trophoblast
apoptosis. Placenta. 20:423–430. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Candelier JJ, Frappart L, Yadaden T, Poaty
H, Picard JY, Prévot S and Coullin P: Altered p16 and Bcl-2
expression reflects pathologic development in hydatidiform moles
and choriocarcinoma. Pathol Oncol Res. 19:217–227. 2013. View Article : Google Scholar
|
|
22
|
Lea RG, al-Sharekh N, Tulppala M and
Critchley HO: The immunolocalization of bcl-2 at the maternal-fetal
interface in healthy and failing pregnancies. Hum Reprod.
12:153–158. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Sakuragi N, Matsuo H, Coukos G, Furth EE,
Bronner MP, VanArsdale CM, Krajewsky S, Reed JC and Strauss JF III:
Differentiation-dependent expression of the BCL-2 proto-oncogene in
the human trophoblast lineage. J Soc Gynecol Investig. 1:164–172.
1994.PubMed/NCBI
|
|
24
|
Huppertz B, Tews DS and Kaufmann P:
Apoptosis and syncytial fusion in human placental trophoblast and
skeletal muscle. Int Rev Cytol. 205:215–253. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Black S, Kadyrov M, Kaufmann P, Ugele B,
Emans N and Huppertz B: Syncytial fusion of human trophoblast
depends on caspase 8. Cell Death Differ. 11:90–98. 2004. View Article : Google Scholar
|
|
26
|
Gauster M, Siwetz M, Orendi K, Moser G,
Desoye G and Huppertz B: Caspases rather than calpains mediate
remodelling of the fodrin skeleton during human placental
trophoblast fusion. Cell Death Differ. 17:336–345. 2010. View Article : Google Scholar
|
|
27
|
Sizemore GM, Sizemore ST, Seachrist DD and
Keri RA: GABA(A) receptor pi (GABRP) stimulates basal-like breast
cancer cell migration through activation of extracellular-regulated
kinase 1/2 (ERK1/2). J Biol Chem. 289:24102–24113. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Johnson SK and Haun RS: The
gamma-aminobutyric acid A receptor pi subunit is overexpressed in
pancreatic adenocarcinomas. JOP. 6:136–142. 2005.PubMed/NCBI
|
|
29
|
Takehara A, Hosokawa M, Eguchi H, Ohigashi
H, Ishikawa O, Nakamura Y and Nakagawa H: Gamma-aminobutyric acid
(GABA) stimulates pancreatic cancer growth through overexpressing
GABAA receptor pi subunit. Cancer Res. 67:9704–9712. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Young SZ and Bordey A: GABA's control of
stem and cancer cell proliferation in adult neural and peripheral
niches. Physiology (Bethesda). 24:171–185. 2009. View Article : Google Scholar
|
|
31
|
Watanabe M, Maemura K, Oki K, Shiraishi N,
Shibayama Y and Katsu K: Gamma-aminobutyric acid (GABA) and cell
proliferation: focus on cancer cells. Histol Histopathol.
21:1135–1141. 2006.PubMed/NCBI
|
|
32
|
Ortega A: A new role for GABA: inhibition
of tumor cell migration. Trends Pharmacol Sci. 24:151–154. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Perry JK, Lins RJ, Lobie PE and Mitchell
MD: Regulation of invasive growth: similar epigenetic mechanisms
underpin tumour progression and implantation in human pregnancy.
Clin Sci (Lond). 118:451–457. 2009. View Article : Google Scholar
|
|
34
|
Murray MJ and Lessey BA: Embryo
implantation and tumor metastasis: common pathways of invasion and
angiogenesis. Semin Reprod Endocrinol. 17:275–290. 1999. View Article : Google Scholar
|
|
35
|
Ferretti C, Bruni L, Dangles-Marie V,
Pecking AP and Bellet D: Molecular circuits shared by placental and
cancer cells, and their implications in the proliferative, invasive
and migratory capacities of trophoblasts. Hum Reprod Update.
13:121–141. 2007. View Article : Google Scholar
|