Introduction
Despite a recent decline in incidence, gastric
cancer (GC) remains one of the most common health issues with a
significant mortality rate (1,2).
There are approximately 800,000 GC-related deaths worldwide each
year (1,3). However, the pathogenetic mechanisms
and the origin of GC are not yet clear. Previous research has
postulated that, by contrast to conventional stochastic theory,
cancers may initiate from a rare subpopulation of cells within the
tumor, known as cancer stem cells (CSCs), which have been
considered the 'heartbeat' of cancer (4,5).
This theory can be traced back to 1997, when Bonnet and Dick first
isolated stem cells from acute myeloid leukemia (AML) (6). Since then, CSCs have been discovered
in various types of tumors, including breast (7), prostate (8), liver (9), colon (10) and pancreatic (11) cancer. Gastric CSCs (GCSCs) have
already been isolated, identified and proven to drive the growth
and metastasis of GC cells (12,13). Moreover, GCSCs possess
chemo-resistant and radio-resistant abilities and differ from their
parental cancer cells (14,15). The emergence of GCSCs may lead to
a paradigm shift in GC research, and may assist in the
establishment of a novel therapeutic tool for the treatment of
GC.
Quercetin (3,3′,4′,5,7-pentahydroxyavone), which is
widely found in edible fruits and vegetables, exerts
anti-inflammatory and antiviral effects (16). Increasing attention has been paid
to its anti-tumor effects on many cancer cell models, including GC
(17,33). Quercetin inhibits the development
of GC possibly by regulating cell cycle arrest or by inducing
protective autophagy (18–20).
Studies on the synergetic, inhibitory effects of quercetin on the
self-renewal ability of pancreatic CSCs have noted the potential
selective toxicity of quercetin to pancreatic CSCs (21–23). Moreover, this synergetic
phenomenon was also observed in prostate CSCs by Tang et al
(24). Moreover, Chang et
al showed that quercetin significantly reduced self-renewal,
stemness signature expression and the migratory ability of head and
neck cancer-derived sphere cells (25). However, to the best of our
knowledge, no studies regarding the effects of quercetin on GCSCs
were completed prior to the present study. Thus, in this study, we
explored the role of quercetin in controlling the proliferation and
apoptosis of GCSCs, and aimed to elucidate the potential molecular
mechanisms involved.
Materials and methods
Culture of parental and spheroid
body-forming cells
The human GC cell line, MGC803, was obtained from
the Cell Bank of the Chinese Academy of Sciences (Shanghai, China).
The cells were cultured in RPMI-1640 medium (Gibco-BRL, Grand
Island, NY, USA) containing 10% fetal bovine serum (FBS) (Gibco,
Rockville, MD, USA), 100 kU/l penicillin, 0.1 g/l streptomycin, 0.3
g/l L-glutamine (Invitrogen, Carlsbad, CA, USA) and 0.85 g/l
NaHCO3 at 37°C in a humidified atmosphere containing 5%
CO2. Spheroid bodies were derived from parental cells.
Briefly, the parental cells were first cultured in serum-free
RPMI-1640 medium with 20 ng/ml human fibroblast growth factor 2
(FGF-2), 100 ng/ml epidermal growth factor (EGF) (Chemicon, Japan),
1% N-2 supplement, 2% B-27 supplement (Invitrogen) and 1%
antibiotic mixture (Gibco). First, parental GC cells were plated
into 96-well ultra-low attachment plates at 1,000 cells/well.
Subsequently, the formation of spheroid bodies was observed under
an inverted microscope once in a while. Primary spheroid bodies
were noted within 2 weeks. The spheroid bodies were then collected
and dissociated by trypsinization for passage and amplification in
serum-free medium.
Colony formation assay
The subspheroid body formation assay was performed
using crystal violet staining. The collected spheroid bodies were
dissociated and centrifuged into single suspended cells. The cells
were seeded into 6-well plates at a density of 1×102
cells/well, as well as the digested GC cells. The serum-free
culture medium was changed every 3–4 days. Two weeks later, the
culture medium was removed and the cells were washed in PBS 3
times. Subsequently, 4% paraformaldehyde was added for 30 min to
fix the cells, which were then stained with crystal violet (Sigma,
St. Louis, MO, USA) for 10 min. Obvious cell colonies were observed
under an inverted microscope following 10 washes with PBS. Colonies
containing <50 cells were excluded.
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
A Qiagen RNeasy mini kit (Qiagen, Valencia, CA, USA)
was used for total RNA extraction from the parental cells and
spheroid body-forming cells. Agarose gel electrophoresis was used
to detect the integrity and purification of RNA samples. Absorption
under 260 nm was used to determine the RNA concentration. cDNA
synthesis was conducted using a High-Capacity cDNA Reverse
Transcription kit (Applied Biosystems, Foster City, CA, USA); qPCR
was performed with SYBR-Green (Takara, Dalian, China) as a
fluorescent dye to determine the changes in the levels of
stem-specific genes [octamer-binding transcription factor 4 (Oct4),
SRY (sex determining region Y)-box 2 (Sox2), Nanog and CD44]
following the manufacturer's instructions. β-actin was used as a
normalizing control. The primer sequences were as follows: Oct4,
5′-AGCCCTCATTTCACCAGGCC-3′ (forward) and 5′-CCCCCACAGAACTCATACGG-3′
(reverse); Sox2, 5′-GAAAAACGAGGGAAATGGG-3′ (forward) and
5′-GCTGTCATTTGCTGTGGGT-3′ (reverse); Nanog,
5′-CTCCTCCCATCCCTCATA-3′ (forward) and 5′-AGGCTCCAACCATACTCC-3′
(reverse); CD44, 5′-CATCCCAGACGAAGACAGTCC-3′ (forward) and
5′-TGATCAGCCATTCTGGAATTTG-3′ (reverse); and β-actin,
5′-CTGAGAGGGAAATCGTGCGT-3′ (forward) and
5′-CCACAGGATTCCATACCCAAGA-3′ (reverse).
MTT assay
An in vitro analysis of the cytotoxic effects
of quercetin on GCSCs was carried out by MTT assay. In brief, 200
µl cells were seeded into a 96-well plate and incubated for
12 h, and the medium was then removed. Following treatment with
various concentrations of quercetin (0, 20, 40, 60, 80 and 100
µM) for the indicated perdios of time at 37°C, 20 µl
5 mg/ml MTT (Sigma) solution were added to each well. Following a 4
h of incubation, DMSO (100 µl) was added to dissolve the
formazan crystals for 15 min. The optical density at 570 nm was
measured to calculate the inhibitory function of quercetin from the
cytotoxicity curves. Cells without treatment were used as controls.
Quercetin (99% pure) was purchased from Sigma, dissolved in DMSO,
and stored at −20°C. Its structure is illustrated in Fig. 1A. The half maximal inhibitory
concentration (IC50) was defined as the drug
concentration which inhibited cell growth by 50%, and was
calculated using a linear regression based on the results of MTT
assay.
Cell sorting by flow cytometry
The cells were counted and transferred to a 5-ml
tube, washed with HBSS containing 2% heat-inactivated FBS, and we
reconstituted cells at a concentration of 106/100
µl with Sandoglobulin solution (Sandoz Pharmaceuticals
Corp., East Hanover, NJ, USA). Subsequently, the cells were
incubated on ice for 20 min. After washing twice with HBSS, the
cells were resuspended in HBSS containing 1 mM HEPES and 2% FBS.
After filtering with a sterile mesh filter (40 µm), the
cells were incubated with anti-CD44-FITC at a dilution of 1:400
(no. 560977; BD Biosciences, Mountain View, CA, USA) for 30 min.
The cells were then rinsed with HBSS and subsequently resuspended
in HBSS solution with 2% FBS, 1 mM HEPES and 1%
penicillin-streptomycin sulfate. Subsequently, the samples were
examined using FACSAria (serial no. 342975; BD Immunocytometry
Systems, Franklin Lakes, NJ, USA) for fluorescence-activated cell
sorting. The cells were routinely sorted twice, as previously
described (26).
Determination of apoptosis by flow
cytometry
The quantification of the apoptosis of the GCSCs
treated with quercetin was examined by flow cytometry. The cells
were treated with 53.0 µM quercetin for 24 h, and then
centrifuged at 1,000 × g for 5 min, and washed twice with PBS. An
Annexin V-fluorescein isothiocyanate (FITC) apoptosis detection kit
(Invitrogen) was used for the detection of apoptotic cells. After
staining with 5 µl Annexin V-FITC and 5 µl propidium
iodide for 15 min in the dark at room temperature, the cells were
immediately subjected to flow cytometric observation and analysis
within 1 h.
Determination of mitochondrial membrane
potential
The resultant fluorescence intensity was used as an
arbitrary unit representing mitochondrial membrane potential. The
GCSCs were treated with the AKT activator, fumonisin B1 (FB1, 25
µM) (Sigma) for 2 h, in prior to exposure to quercetin for
24 h and rinsed with PBS. The cells were resuspended in 1 ml
rhodamine 123 (1 µg/ml; Sigma) for 30 min in the dark at
37°C. After rinsing with PBS 3 times, the fluorescence intensity of
the cells was examined by fluorescence-activated cell sorting at
507 nm (excitation) and 529 nm (emission).
Caspase activity assay
Caspase activity was measured using a caspase
colorimetric assay kit from Sigma. Briefly, the cells were treated
with quercetin for 24 h and then lysed. The centrifugal supernatant
was placed into a 96-well microplate with reaction buffer. The
microplate was incubated at 37°C overnight and substrate was added.
The activity was measured at 405 nm using a spectrophotometer
(U-5100; Techcomp Ltd., Shanghai, China).
Western blot analysis
Before western blot analysis was performed, the
cells were lysed with RIPA lysis buffer [100 mM NaCl, 50 mM
Tris-HCl pH 7.5, 1% Triton X-100, 1 mM ethylenediaminetetraacetic
acid (EDTA), 10 mM β-glycerophosphate and 2 mM sodium vanadate and
protease inhibitor] to collect whole-cell extracts. To prepare the
cytosolic extracts, the cells were lysed with RIPA lysis buffer
containing protease inhibitor cocktail for 30 min on ice, and were
then homogenized by 6 passages through a 25-gauge needle. Following
centrifugation (14,000 × g) at 4°C for 10 min, the resultant
supernatant (cytosol) was subjected to western blot analysis to
evaluate the release of cytochrome c (Cyt-c). The protein
samples were then separated using SDS-PAGE on a 12% gel and
transferred onto polyvinylidene difluoride membranes (Amersham
Pharmacia, Freiburg, Germany). Non-specific binding was blocked by
incubation with 5% non-fat milk in Tris-buffered saline with
Tween-20 (TBST) buffer at room temperature for 1 h. Incubation with
the primary antibody was carried out overnight in a cold room, and
the membranes were subsequently washed in TBST 3 times. Primary
antibodies were as follows: anti-Bax (no. 2774, 1:400), anti-Bcl-2
(no. 2872, 1:200), anti-Cyt-c (no. 4272, 1:300) and anti-β-actin
(no. 4967S) were purchased from Cell Signaling Technology (Danvers,
MA, USA). A secondary antibody conjugated to horseradish peroxidase
(1:5,000; Sigma) was added to the membranes followed by incubation
for 1 h at room temperature, followed by enhanced chemiluminescence
using an ECL kit (Amersham Pharmacia, Piscataway, NJ, USA).
Finally, protein bands were visualized using ECL and quantified
using Quantity One software (Bio-Rad, Hercules, CA, USA).
Statistical analysis
All experimental data were derived from at least 3
independent experiments, and all values are expressed as the means
± SD. Statistical analyses were performed using the SPSS 16.0
statistical software package (SPSS Inc., Chicago, IL, USA). Any
differences were assessed using the Student's t-test or one-way
ANOVA. A value of p<0.05 was considered to indicate a
statistically significant difference.
Results
Isolation of GCSCs from MGC803 cells
The MGC803 parental cells cultured in serum-free
medium grew as non-adherent, three-dimensional spheroid clusters,
termed spheroid bodies. As time progressed, the cell spheroid
bodies gradually increased not only in volume, but also in number.
The self-renewal capacity of these spheroid body-forming cells was
preliminarily assessed by counting the subspheroid bodies that they
generated. As shown in Fig. 1B,
compared with the parental cells, the rate of spheroid body
formation was 25.4±3.51% in the spheroid body cells, whereas for
the parental cells it was only 5.0±1.04%, reflecting the high
capacity for self-renewal of these isolated spheroid body cells.
Furthermore, the results of RT-qPCR revealed that the mRNA levels
of stem-specific Oct4, Sox2, Nanog and CD44 in the spheroid
body-forming cells were significantly higher than those in the
parental cells (Fig. 1C). In
addition, the results of western blot analysis revealed the high
expression of the stem-cell markers, Oct4, Sox2, Nanog and CD44, in
the spheroid body cells, as compared to the parental cells
(Fig. 1D). The relative protein
levels of CD44, Nanog, Sox2 and Oct4 in the spheroid body-forming
cells were approximately double those of the parental cells, at
least for CD44, Nanog and Sox2 (Fig.
1E). Taken together, these results indicate the successful
isolation of GCSCs from the MGC803 GC cells.
Quercetin inhibits the viability of
GCSCs
To determine the cytotoxity of quercetin on the
GCSCs, we used an MTT assay to examine cell viability. As shown in
Fig. 2A, treatment with quercetin
significantly increased the inhibition rate of viable cells in a
time- and dose-dependent manner. Moreover, after treating the GCSC
cells with 20 and 100 µM quercetin for 24 h, cell viability
decreased to 82±6.73% (20 µM) and 20.5±2.10% (100 µM)
(Fig. 2B), and quercetin strongly
restrained GCSC cell viability with an IC50 of
approximately 53 µM. Additionally, we noted that the MGC803
cells were more sensitive than the GCSCs to quercetin, reflecting
the relative stronger chemoresistance of GCSCs in comparison to the
parental cells. Moreover, a decrease of >50% in the number of
Oct4+CD44+ cells was observed following
treatment with quercetin (Fig.
2C), indicating the diminished proliferative ability of GCSCs
upon treatment with quercetin.
 | Figure 2Effects of quercetin on the growth of
gastric cancer stem cells (GCSCs). (A) Increasing concentrations of
quercetin (20, 40, 60, 80 and 100 µM) were added and the
cells were incubated for different periods of time (12, 24, 36 and
48 h), and the absorption at 570 nm was measured. (B) After 24 h of
treatment, the dose-dependent effects of quercetin (20, 40, 60, 80
and 100 µM) on cell viability were determined by MTT assay.
(C) Effects of quercetin on GCSCs. Cells were treated with 53
µM (IC50 value) quercetin for 24 h. Flow
cytometry was applied to detect the percentage of cells bearing the
stemness markers, Oct4 and CD44 (Oct4+CD44+
cells). (D) Flow cytometry was applied for the detection of
quercetin-induced cell apoptosis after 24 h of treatment with
quercetin at 53 µM. (E) The quantification of cell apoptosis
is also demonstratedted by the values in the quadrants Q4 (early
apoptotic cells) and Q2 (late apoptotic cells) shown in (E).
*p<0.05 and **p<0.01 compared to
control groups. |
Quercetin induces the apoptosis of
GCSCs
In order to further analyze the effects of quercetin
on GCSC survival, flow cytometry was used to assess
quercetin-induced apoptosis. As shown in Fig. 2D, in comparison with the control
(untreated) group, quercetin significantly induced apoptosis. When
the cells were treated with 53.0 µM quercetin for 24 h, GCSC
apoptosis was significantly enhanced (Fig. 2E). However, the sensitivity of the
GCSCs to quercetin was less than that of the MGC803 cells. These
data reflect the inhibitory effects which quercetin exerts on the
growth of GCSCs; however, the mechanisms involved require further
investigation.
Quercetin and the mitochondrial apoptotic
pathway
Mitochondrial apoptosis plays an important role in
GC growth inhibition. To ascertain the underlying mechanisms
involved in the quercetin-triggered inhibition of cell growth, the
mitochondrial apoptotic pathway was analyzed. As shown in Fig. 3A, quercetin decreased
mitochondrial membrane potential in a time-dependent manner. The
fluorescence intensity at 12 h was higher than after 24 h of
treatment with quercetin. Concomitantly, as central molecules of
the apoptotic cascade, caspase-3 and caspase-9 were also
investigated, and we noted that quercetin upregulated the
expression levels of both caspase-3 and caspase-9 (Fig. 3B). Our findings demonstrated that
quercetin-induced apoptotic cell death was associated with the
activation of caspase-3 and caspase-9. In order to further explore
the involvement of the mitochondrial apoptotic pathway in the
quercetin-induced inhibition of GCSC growth, the expression of Bax,
Bcl-2 and Cyt-c was examined by western blot anlaysis. As shown in
Fig. 3C, an increase in the
levels of Bax and Cyt-c occurred, together with a significant
decrease in the expression of the anti-apoptotic protein, Bcl-2,
upon treatment with quercetin. The variations in the levels of
these proteins can be directly noted in Fig. 3D. These results suggest that
quercetin inhibits the growth of GCSCs by regulating the
mitochondrial apoptotic pathway.
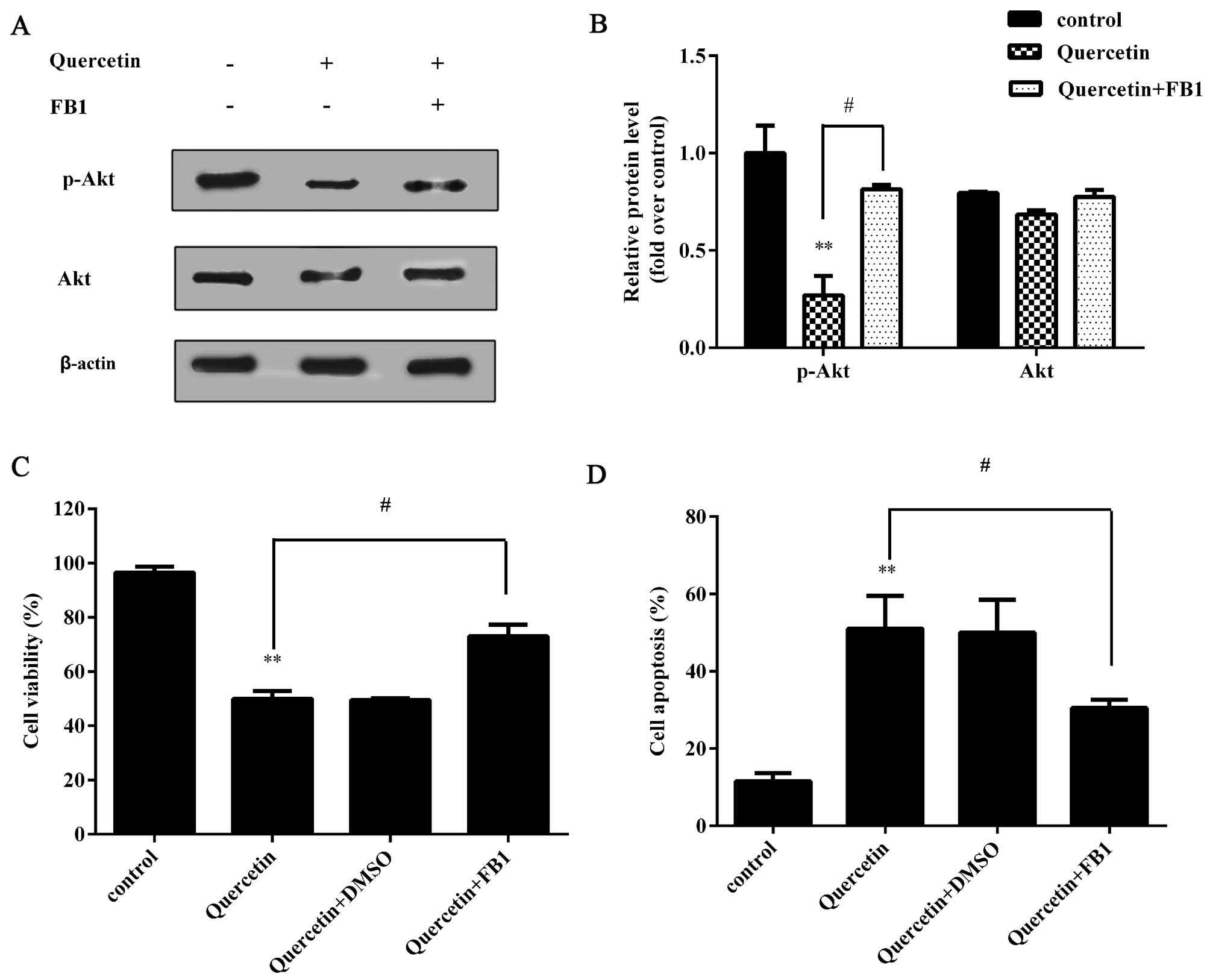
Akt is involved in quercetin-induced
growth inhibition
Previous studies have demonstrated that quercetin
induces cell apoptosis in many types of cancer by blocking the
phosphoinositide 3-kinase (PI3K)-Akt pathway, which plays a
negative role in the intrinsic mitochondrial pathway (27–29). Therefore, in the present study,
the phosphorylation of Akt was investigated. As was expected, an
attenuated level of p-Akt was observed following treatment with
quercetin (Fig. 4A). Treatment
with quercetin induced only a slight decrease in the expression of
Akt (Fig. 4B). On the other hand,
the addition of fumonisin B1 (FB1, Akt activator) significantly
attenuated the inhibitory effects of quercetin on Akt
phosphorylation (Fig. 4A).
Moreover, the quercetin-induced inhibition of GCSC viability was
almost completely abrogated once Akt was activated by treatment
with FB1 (Fig. 4C). At the same
time, the upregulation of cell apoptosis induced by quercetin was
also markedly decreased when the cells were pre-treated with FB1
(Fig. 4D). These results suggest
that the PI3K-Akt signaling pathway is involved in the
quercetin-induced inhibition of the growth of GCSCs.
Association between PI3K-Akt and the
mitochondrial pathway
In order to gain further insight into the
involvement of Akt in the mitochondrial apoptotic pathway, we
examined the influence of FB1 on the quercetin-induced
mitochondrial pathway. As shown in Fig. 5A, treatment with quercetin
significantly reduced mitochondrial membrane potential, while the
addition of FB1 reversed this effect. FB1 markedly abated the
increase in the level of caspase-3 (Fig. 5B) and caspase-9 (Fig. 5C) induced by quercetin.
Furthermore, the effects of quercetin on the expression of Cyt-c,
Bax and Bcl-2 were attenuated by the Akt activator, FB1 (Fig. 5D). Their relative levels were
calculated and are depicted in Fig.
5E. These results clearly illustrate the involvement of Akt in
the quercetin-induced activation of the mitochondrial apoptotic
pathway.
Discussion
The incidence of GC, which is increasingly perceived
as a leading cause of mortality particularly in developing
countries, has increased due to the increased aging populations.
Conventional curative treatments, such as surgery, chemotherapy and
radiation therapy, are not completely effective, and thus GCSC
theory provides new insight into the study of tumor initiation, as
well as the development of novel strategies for GC treatment
(30,31). Recently, modern tools have been
used to investigate the characteristics of GCSCs, including
self-renewal pathways, chemoresistance mechanisms and even microRNA
expression profiles (31,32). In spite of the significance of
GCSCs for GC genesis and development, limited research has been
undertaken on the control of GCSCs by effective compounds which
serve to block the origination of GCs.
Quercetin (3,3′,4′,5,7-pentahydroxyavone) is a
plant-based chemical that is commonly found in various types of
fruits and vegetables (33).
Although quercetin has been evaluated in relation to several types
of cancer (17,22,24,33), very little is known about its
effects on GCSCs. In the present study, we provide evidence that
quercetin exerts a desirable toxic effect on GCSCs, and inhibits
their survival. Although the exact mechanisms responsible for the
quercetin-induced apoptosis of GCSCs remain unclear, some
similarities should exist due to the homology between GCSCs and GCs
(13). In 2012, it was
hypothesized that quercetin-induced GC cell apoptosis was mediated
via the mitochondrial intrinsic apoptotic pathway, which was an
important theory that involved the release of apoptotic signals
(35). The mitochondrial
apoptotic pathway in mammals is regulated by members of the Bcl-2
family of proteins (34).
Similarly, in the present study, the collapse of the membrane
potential, and the activation of caspase-3 and caspase-9, as well
as the upregulation of Cyt-c following treatment with quercetin,
suggested the involvement of the mitochondrial apoptotic pathway.
Moreover, the downregulation of the anti-apoptotic protein, Bcl-2,
and the upregulation of the pro-apoptotic protein, Bax, further
reinforces the hypothesis that quercetin-induced growth inhibition
occurrs through mitochondrial-dependent signaling in GCSCs.
PI3K-Akt is a central regulatory protein which is
involved in cell survival and plays a significant role in the
regulation of many cellular functions, such as proliferation, cell
viability and cell apoptosis (27,28,36). In this study, we noted that Akt
phosphorylation was decreased following treatment with quercetin.
When the cells were pre-treated with the Akt activator, FB1, the
quercetin-induced inhibition of the phosphorylation of Akt was
markedly attenuated. Furthermore, the quecetin-induced inhibition
of cell growth and the enhancement of cell apoptosis were also
attenuated by treatment with FB1. These data strongly suggest the
involvement of the PI3K-Akt pathway in quercetin-induced growth
inhibition, which is consistent with the findings of previous
studies on the inhibitory effects of quercetin on Akt activation
(27,28,36). Based on previous studies which
suggested that the PI3K-Akt pathway plays a negative role in the
mitochondrial intrinsic pathway (37,38), we confirmed the involvement of the
PI3K-Akt pathway in the mitochondrial pathway. The effects of
quercetin on the expression of Cyt-c, Bcl-2 and Bax, the activation
of caspase-3 and caspase-9, as well as on mitochondrial membrane
potential, were found to be significantly abated by FB1. These
findings suggest that the inhibitory effecfts exerted by quercetin
against GCSCs are regulated by the mitochondrial apoptotic pathway
via the PI3K-Akt pathway.
In conclusion, in the present study, we provide
evidence that quercetin has the potential to effectively intervene
and prevent GCSC growth. Moreover, we noted that quercetin-induced
apoptosis occurred via the mitochondrial-dependent pathway, which
was mediated via the PI3K-Akt pathway. However, quercetin was less
effective in the GCSCs than the MGC803 cells. Hence, promoting the
specificity of quercetin to GCSCs is an aim for futuer research. We
hypothesize that understanding the modulatory mechanisms of
quercetin and promoting its specificity to GCSCs may aid in the
development of quercetin as a potential chemotherapeutic agent for
GC.
References
|
1
|
Boku N: HER2-positive gastric cancer.
Gastric Cancer. 17:1–12. 2014. View Article : Google Scholar :
|
|
2
|
Pang X, Wei W, Leng W, Chen Q, Xia H, Chen
L and Li R: Radiotherapy for gastric cancer: a systematic review
and meta-analysis. Tumour Biol. 35:387–396. 2014. View Article : Google Scholar
|
|
3
|
Jemal A, Center MM, DeSantis C and Ward
EM: Global patterns of cancer incidence and mortality rates and
trends. Cancer Epidemiol Biomarkers Prev. 19:1893–1907. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Haraguchi N, Ishii H, Sakai D, Satoh T,
Doki Y and Mori M: The concept and significance of cancer stem
cells. Nippon Geka Gakkai Zasshi. 113:204–209. 2012.In
Japanese.
|
|
5
|
Xu G, Shen J, Ou Yang X, Sasahara M and Su
X: Cancer stem cells: the 'heartbeat' of gastric cancer. J
Gastroenterol. 48:781–797. 2013. View Article : Google Scholar
|
|
6
|
Bonnet D and Dick JE: Human acute myeloid
leukemia is organized as a hierarchy that originates from a
primitive hematopoietic cell. Nat Med. 3:730–737. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Marsden CG, Wright MJ, Pochampally R and
Rowan BG: Breast tumor-initiating cells isolated from patient core
biopsies for study of hormone action. Methods Mol Biol.
590:363–375. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Collins AT, Berry PA, Hyde C, Stower MJ
and Maitland NJ: Prospective identification of tumorigenic prostate
cancer stem cells. Cancer Res. 65:10946–10951. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Yang ZF, Ngai P, Ho DW, Yu WC, Ng MN, Lau
CK, Li ML, Tam KH, Lam CT, Poon RT and Fan ST: Identification of
local and circulating cancer stem cells in human liver cancer.
Hepatology. 47:919–928. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
O'Brien CA, Pollett A, Gallinger S and
Dick JE: A human colon cancer cell capable of initiating tumour
growth in immunodeficient mice. Nature. 445:106–110. 2007.
View Article : Google Scholar
|
|
11
|
Li C, Lee CJ and Simeone DM:
Identification of human pancreatic cancer stem cells. Methods Mol
Biol. 568:161–173. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Dewi DL, Ishii H, Kano Y, Nishikawa S,
Haraguchi N, Sakai D, Satoh T, Doki Y and Mori M: Cancer stem cell
theory in gastrointestinal malignancies: recent progress and
upcoming challenges. J Gastroenterol. 46:1145–1157. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Li K, Dan Z and Nie YQ: Gastric cancer
stem cells in gastric carcinogenesis, progression, prevention and
treatment. World J Gastroenterol. 20:5420–5426. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Zhou ZH, Zhang JD, Xu GF and Zhao HB:
Isolation of gastric cancer stem cells based on colony morphology
and detection of their sensitivity to 5-fluorouracil. Zhonghua Wei
Chang Wai Ke Za Zhi. 16:376–380. 2013.In Chinese. PubMed/NCBI
|
|
15
|
Liu J, Ma L, Xu J, Liu C, Zhang J, Liu J,
Chen R and Zhou Y: Spheroid body-forming cells in the human gastric
cancer cell line MKN-45 possess cancer stem cell properties. Int J
Oncol. 42:453–459. 2013.
|
|
16
|
Boots AW, Haenen GR and Bast A: Health
effects of quercetin: from antioxidant to nutraceutical. Eur J
Pharmacol. 585:325–337. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Russo GL, Russo M, Spagnuolo C, Tedesco I,
Bilotto S, Iannitti R and Palumbo R: Quercetin: a pleiotropic
kinase inhibitor against cancer. Cancer Treat Res. 159:185–205.
2014. View Article : Google Scholar
|
|
18
|
Wang K, Liu R, Li J, Mao J, Lei Y, Wu J,
Zeng J, Zhang T, Wu H, Chen L, et al: Quercetin induces protective
autophagy in gastric cancer cells: Involvement of Akt-mTOR- and
hypoxia-induced factor 1α-mediated signaling. Autophagy. 7:966–978.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Wang HY, Guo LM, Chen Y, Zhao XH, Cheng
CL, Wu MY and He LY: Quercetin inhibits growth and induces
apoptosis of human gastric carcinoma cells. Xi Bao Yu Fen Zi Mian
Yi Xue Za Zhi. 22:585–587. 2006.In Chinese. PubMed/NCBI
|
|
20
|
Yoshida M, Sakai T, Hosokawa N, Marui N,
Matsumoto K, Fujioka A, Nishino H and Aoike A: The effect of
quercetin on cell cycle progression and growth of human gastric
cancer cells. FEBS Lett. 260:10–13. 1990. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Adikrisna R, Tanaka S, Muramatsu S, Aihara
A, Ban D, Ochiai T, Irie T, Kudo A, Nakamura N, Yamaoka S and Arii
S: Identification of pancreatic cancer stem cells and selective
toxicity of chemotherapeutic agents. Gastroenterology. 143:234–245
e237. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Srivastava RK, Tang SN, Zhu W, Meeker D
and Shankar S: Sulforaphane synergizes with quercetin to inhibit
self-renewal capacity of pancreatic cancer stem cells. Front Biosci
(Elite Ed). 3:515–528. 2011. View
Article : Google Scholar
|
|
23
|
Zhou W, Kallifatidis G, Baumann B, Rausch
V, Mattern J, Gladkich J, Giese N, Moldenhauer G, Wirth T, Büchler
MW, et al: Dietary polyphenol quercetin targets pancreatic cancer
stem cells. Int J Oncol. 37:551–561. 2010.PubMed/NCBI
|
|
24
|
Tang SN, Singh C, Nall D, Meeker D,
Shankar S and Srivastava RK: The dietary bioflavonoid quercetin
synergizes with epigallocathechin gallate (EGCG) to inhibit
prostate cancer stem cell characteristics, invasion, migration and
epithelial-mesenchymal transition. J Mol Signal. 5:142010.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Chang WW, Hu FW, Yu CC, Wang HH, Feng HP,
Lan C, Tsai LL and Chang YC: Quercetin in elimination of tumor
initiating stem-like and mesenchymal transformation property in
head and neck cancer. Head Neck. 35:413–419. 2013. View Article : Google Scholar
|
|
26
|
Li C, Heidt DG, Dalerba P, Burant CF,
Zhang L, Adsay V, Wicha M, Clarke MF and Simeone DM: Identification
of pancreatic cancer stem cells. Cancer Res. 67:1030–1037. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Granado-Serrano AB, Martín MA, Bravo L,
Goya L and Ramos S: Quercetin induces apoptosis via caspase
activation, regulation of Bcl-2, and inhibition of PI-3-kinase/Akt
and ERK pathways in a human hepatoma cell line (HepG2). J Nutr.
136:2715–2721. 2006.PubMed/NCBI
|
|
28
|
Xiang T, Fang Y and Wang SX: Quercetin
suppresses HeLa cells by blocking PI3K/Akt pathway. J Huazhong Univ
Sci Technolog Med Sci. 34:740–744. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Pan HC, Jiang Q, Yu Y, Mei JP, Cui YK and
Zhao WJ: Quercetin promotes cell apoptosis and inhibits the
expression of MMP-9 and fibronectin via the AKT and ERK signalling
pathways in human glioma cells. Neurochem Int. 80:60–71. 2015.
View Article : Google Scholar
|
|
30
|
Takaishi S, Okumura T and Wang TC: Gastric
cancer stem cells. J Clin Oncol. 26:2876–2882. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Stojnev S, Krstic M, Ristic-Petrovic A,
Stefanovic V and Hattori T: Gastric cancer stem cells: therapeutic
targets. Gastric Cancer. 17:13–25. 2014. View Article : Google Scholar
|
|
32
|
Liu J, Ma L, Wang Z, Wang L, Liu C, Chen R
and Zhang J: MicroRNA expression profile of gastric cancer stem
cells in the MKN-45 cancer cell line. Acta Biochim Biophys Sin
(Shanghai). 46:92–99. 2014. View Article : Google Scholar
|
|
33
|
Borska S, Chmielewska M, Wysocka T,
Drag-Zalesinska M, Zabel M and Dziegiel P: In vitro effect of
quercetin on human gastric carcinoma: targeting cancer cells death
and MDR. Food Chem Toxicol. 50:3375–3383. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Brunelle JK and Letai A: Control of
mitochondrial apoptosis by the Bcl-2 family. J Cell Sci.
122:437–441. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Wang P, Zhang K, Zhang Q, Mei J, Chen CJ,
Feng ZZ and Yu DH: Effects of quercetin on the apoptosis of the
human gastric carcinoma cells. Toxicol In Vitro. 26:221–228. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Spencer JP, Rice-Evans C and Williams RJ:
Modulation of pro-survival Akt/protein kinase B and ERK1/2
signaling cascades by quercetin and its in vivo metabolites
underlie their action on neuronal viability. J Biol Chem.
278:34783–34793. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Zhang Y, Sun S, Chen J, Ren P, Hu Y, Cao
Z, Sun H and Ding Y: Oxymatrine induces mitochondria dependent
apoptosis in human osteosarcoma MNNG/HOS cells through inhibition
of PI3K/Akt pathway. Tumour Biol. 35:1619–1625. 2014. View Article : Google Scholar
|
|
38
|
Cheong JW, Min YH, Eom JI, Kim SJ, Jeung
HK and Kim JS: Inhibition of CK2α and PI3K/Akt synergistically
induces apoptosis of CD34+CD38− leukaemia
cells while sparing haematopoietic stem cells. Anticancer Res.
30:4625–4634. 2010.PubMed/NCBI
|