Introduction
Alzheimer's disease (AD) is a progressive
neurodegenerative disease characterized by the accumulation of
senile plaques, the formation of neurofibrillary tangles, neuronal
loss and progressive cognitive impairment. Amyloid β (Aβ)
aggregation and accumulation in the brain is considered a causative
factor in the pathogenesis of AD, while the progression of AD is
considered to be multifactorial, and involves an increase in
oxidative stress and inflammation (1–3).
The increased accumulation of cellular reactive oxygen species
(ROS) and oxidative damage to lipids, nucleic acids and proteins,
as well as the activation of inflammatory responses are considered
to play major roles in the process of age-related neurodegeneration
and cognitive decline (3–7).
Many inflammatory pathways have been implicated in
AD. Reactive glial cells, microglia and astrocytes produce multiple
inflammatory factors, including cytokines, chemokines, ROS and
cyclooxygenase-2 (COX-2). Elevated levels of these factors have
been discovered in AD and are among the major mechanisms of AD
neuropathology (3,8). Anti-inflammatory drugs, such as
non-steroidal anti-inflammatory drugs, peroxisome
proliferator-activated receptor-γ agonists, minocycline, and tumor
necrosis factor-α (TNF-α) inhibitors attenuate microglial
activation, amyloid deposition and tau hyperphosphorylation in
animal models of AD (9–14). Moreover, it has been demonstrated
that several natural antioxidants and anti-inflammatory compounds,
such as curcumin and grape-derived polyphenols can inhibit Aβ
aggregation and attenuate cognitive deterioration (15–18).
Humulus japonicus Siebold & Zucc. (HJ),
also known as 'Japanese hop', is a perennial herb that belongs to
the Cannabaceae family and is commonly distributed in Asian
countries including Korea, Japan and China. In western countries,
HJ was known to be originally imported for use as an ornamental
vine, but it is now considered as an ecologically threatening plant
due to the rapid growth and spreading capacity of the HJ vine,
resulting in the blocking of light to plants beneath (19). According to previous studies, HJ
has demonstrated scavenging effects on active oxygen molecules,
such as superoxide radicals, hydroxyl radicals and hydrogen
peroxide (20,21). It has been reported that HJ exerts
potent anti-inflammatory effects by blocking the generation of
lipopolysaccharide (LPS)-induced TNF-α, interleukin (IL)-1β, IL-6,
inducible nitric oxide synthase (iNOS) and COX-2 expression in the
macrophage cell line, RAW264.7. Furthermore, HJ was found to
inhibit the LPS-induced phosphorylation of the inhibitor of κB-α
(IκBα) (22). However, the
effects of HJ on AD have not yet been elucidated, at least to the
best of our knowledge. In the present study, we investigated
whether HJ confers a beneficial effect on glial-mediated
inflammation in the central nervous system (CNS) and in APP/PS1
transgenic (Tg-APP/PS1) mice, a murine model of AD, and
demonstrated that HJ has a therapeutic potential to attenuate
neuroinflammation, AD-like pathology and cognitive impairment.
Materials and methods
Animals
Tg-APPswe/PS1dE9 transgenic mice (Tg-APP/PS1 mice)
were obtained from the Jackson Laboratory (Bar Harbor, ME, USA) and
maintained in a hemizygote state by crossing transgenic C57BL6 x
C3H F1 hybrid mice and non-transgenic C57BL6 x C3H F1 mice
(23,24). Mice not expressing the transgene
were used as wild-type controls. The mice were housed in regular
polycarbonate plastic cages in an environment with controlled
temperature (21–22°C) and humidity (50–60%) with a 12-h light/dark
cycle (lights on at 7 a.m.). Furthermore, the animals were
maintained on an ad libitum diet of laboratory chow (2018S;
Harlan Teklad, Madison, WI, USA) and had free access to water. The
cages were filled to an approximate depth of 1.5 cm with bedding
made of chopped wood particles (JSBio, Daejeon, Korea). All
materials used were autoclaved and gamma-irradiated. The animal
room was maintained under specific-pathogen-free (SPF) conditions.
The Tg-APP/PS1 mice at 5 months of age were randomized into the
control (n=10) and HJ (n=10) groups. The methanolic extract of HJ
at 500 mg/kg/day or the vehicle were administered to the mice by
oral gavage for 2.5 months from 5 months of age. All animal
experiments were approved by the Institutional Animal Use and Care
Committee of the Korea Research Institute of Bioscience and
Biotechnology and were performed in accordance with the Guide for
the Care and Use of Laboratory Animals published by the US National
Institutes of Health (NIH publication, 8th edition, 2011).
Preparation and administration of HJ
HJ was purchased from Gangwon Herbs, Gangwon,
Republic of Korea, on July, 2014. The voucher specimen was
identified by Professor W.K. Oh, and a voucher specimen
(SNU-2014-0004) was deposited at the College of Pharmacy, Seoul
National University, Korea. The HJ extract was then prepared and
supplied by the Korea Bioactive Natural Material Bank (Seoul,
Korea). Briefly, the dried aerial parts of HJ were soaked in 100%
methanol in an extraction container for 2 days at room temperature.
The methanol-soluble extract was filtered through a cheesecloth,
concentrated exhaustively and dried to produce a methanolic extract
under reduced pressure. This HJ extract was suspended in 0.5%
carboxymethylcellulose (CMC) at a concentration of 50 mg/ml as a
stock solution. The 5-month-old male and female mice were
administered orally 500 mg/kg/day of HJ-methanol extract or 0.5%
CMC (vehicle) for 10 weeks.
Cell culture
BV-2 microglial cells (a gift from Dr B.S. Han,
KRIBB, Daejeon, Korea) were maintained in Dulbecco's modified
Eagle's medium (DMEM, Cat. no. SH30243.01; HyClone, Logan, UT, USA)
supplemented with 10% heat-inactivated fetal bovine serum (FBS) and
1% penicillin/streptomycin (both from Gibco, Grand Island, NY,
USA). The cells were incubated at 37°C under 5% CO2 and
95% relative humidity. They were seeded in 12-well plates
(1×105 cells/well), incubated overnight, and thereafter
co-treated with differently diluted HJ extracts (100 and 500
µg/ml) for 24 h. In this preliminary experiment, HJ
concentrations of 50, 100, 200 and 500 µg/ml were not shown
to have cytotoxic effects on the BV-2 cells (data not shown).
Subsequently, two different concentrations of HJ (100 and 500
µg/ml) were selected for use in further experiments, and the
BV-2 cells were cultured under the same conditions with additional
LPS (1 µg/ml, Sigma-Aldrich, St. Louis, MO, USA) for 24
h.
Measurement of nitric oxide (NO)
production
Nitrite in the culture supernatants was measured by
the Griess reaction as an indicator of NO production. The BV2 cells
were seeded in 12-well plates (1×105 cells/well),
incubated overnight, and thereafter co-treated with various
concentrations of HJ (100 and 500 µg/ml) and LPS (1
µg/ml) for 24 h. Following LPS stimulation, a 100 µl
volume of culture supernatants from each sample were mixed with the
same volume of Griess reagent (G4410; Sigma-Aldrich) and then
incubated at room temperature for 15 min. The NO concentration was
determined by measuring the absorbance at 540 nm on a microplate
reader (Thermo Multiskan Spectrum; Thermo Fisher Scientific,
Waltham, MA, USA). The nitrite concentration was calculated with
reference to a sodium nitrite standard curve generated with known
concentrations.
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
RNA preparation and qPCR were performed as
previously described (25).
Following 24 h of stimulation with LPS or treatment with LPS + HJ,
the mRNA expression levels of TNF-α, IL-1β, IL-6, iNOS and β-actin
in BV2 microglial cells were evaluated by RT-qPCR. In addition,
following the administration of HJ for 2.5 months in vivo,
mice were anesthetized with Avertin (0.3 g/kg, i.p.) and the mouse
brain was extracted and dissected using Rodent Brain Matrix (ASI
Instruments, Warren, MI, USA). The cortex was removed and the
expression levels of TNF-α, IL-1β and IL-6 were evaluated by
RT-qPCR. Total RNA was purified from the cells and tissues using
TRI reagent (Sigma-Aldrich). Reverse transcription was carried out
using a Promega RT-PCR kit (Promega, Madison, WI, USA). PCR
reaction was prepared with a mix of 5 µl of 2X SYBR-Green
mix and performed using the StepOne Real-Time PCR system (both from
Applied Biosystems, Foster City, CA, USA). The cycle number at
which the fluorescence emission exceeds the fixed threshold was
defined as threshold cycle (CT). The cycling conditions were as
follows: 10 min at 95°C, followed by 41 cycles of 20 sec at 95°C,
30 sec at 60°C, and 20 sec at 72°C. The difference in amplification
fold was calculated based on qPCR amplification of the target gene
vs. β-actin as a reference using built-in software of the Gene
Expression Analysis for StepOne Software version 2.1 (Applied
Biosystems). qPCR was repeated 4 times. The following primer sets
were used: TNF-α forward, 5′-ATCCGCGACGTGGAACTG-3′ and reverse,
5′-ACCGCCTGGAGTTCTGGAA-3′; IL-1β forward,
5′-CTACAGGCTCCGAGATGAACAAC-3′ and reverse,
5′-TCCATTGAGGTGGAGAGCTTTC-3′; IL-6 forward,
5′-TTCCATCCAGTTGCCTTCTTG-3′ and reverse,
5′-GGGAGTGGTATCCTCTGTGAAGTC-3′; iNOS forward,
5′-GTTCTCAGCCCAACAATACAAGA-3′ and reverse,
5′-GTGGACGGGTCGATGTCAC-3′; and β-actin forward,
5′-TATTGGCAACGAGCGGTTCC-3′ and reverse,
5′-GGCATAGAGGTCTTTACGGATGT-3′.
Cytokine assays
The levels of TNF-α, IL-1β, and IL-6 produced were
measured using ELISA kits (BD Biosciences, San Diego, CA, USA)
according to the manufacturer's instructions. Briefly, the BV2
microglial cells were seeded in 12-well plates (1×105
cells/well), incubated overnight and thereafter co-treated with
various concentrations of HJ (100 and 500 µg/ml) and LPS (1
µg/ml) for 24 h. A total of 100 µl of culture
supernatants was collected to determine the TNF-α, IL-1β, and IL-6
concentrations by ELISA.
Novel object recognition test
Novel object recognition is a validated and widely
used test for assessing recognition memory (26,27). The mice were placed individually
in a 40×20×20 cm3 testing chamber for 10 min with two
identical objects (familiar, acquisition session). The mice were
then returned to home cages and one day they later placed back in
the testing chamber in the presence of one of the original objects
and one novel object (novel, recognition session) for 10 min. The
original objects were cylindrical wooden blocks 10 cm high × 2 cm
in diameter. The novel object was a 10×2.5×2 cm rectangular wooden
block. The acquisition and recognition sessions were video-recorded
and an observer who was blinded to the drug treatment scored the
time spent exploring the objects. The chambers and objects were
cleaned with ethanol between trials. Exploration was defined as
sniffing and touching the object with the nose and/or forepaws.
Sitting on the object was not considered exploratory behavior. In
preliminary experiments, wild-type mice exhibited no significant
preference for the cylindrical or rectangular block. The time spent
exploring both objects was calculated. A discrimination index was
calculated for each animal and expressed by the ratio of time spent
exploring the novel object − time spent exploring the familiar
object/(time spent exploring the novel object + time spent
exploring the familiar object) on day 2, as previously described
(28).
Spontaneous Y-maze alternation test
The Y-maze test was conducted as previously
described (29). A Y-maze with
three identical arms of plexiglass (51.5×11.5×12 cm) 120° apart was
placed in the center of a room. The walls of each arm had a
distinct design that provided visual cues. Each mouse was placed at
the end of one arm facing the center and allowed to explore the
maze for a period of 10 min. The sessions were video-recorded and
scored for entries into arms. The percentage of spontaneous
alternation was calculated as the ratio of the actual to possible
alternations (defined as the total number of arm entries minus 2)
multiplied by 100: alternation (%) = [(number of
alternation)/(total arm entries − 2)] ×100.
Immunohistochemistry
Immunohistochemistry was conducted as previously
described (30). The mice were
transcardially perfused with phosphate-buffered saline (PBS)
followed by 4% paraformaldehyde in PBS. The perfused brains were
dissected, post-fixed overnight and then cut into
40-µm-thick coronal sections on a vibratome (Vibratome
VT1000A; Leica, Mannheim, Germany). Free-floating sections were
incubated in PBS containing 3% H2O2 (v/v),
rinsed 3 times in PBS, and blocked with serum for 1 h at room
temperature. The sections were incubated overnight at 4°C with the
primary antibodies. The primary antibodies were mouse monoclonal
antibodies against Bam10 (4% BSA, Cat. no. A5213; Sigma-Aldrich),
rabbit polyclonal antibodies against ionized calcium-binding
adapter molecule 1 (Iba-1, 5% HS, Cat. no. 019-19741; Wako,
Richmond, VA, USA), glial fibrillary acidic protein (GFAP, 5% GS,
Cat. no. Z0334; Dako) and mouse polyclonal antibody against AT8
(phospho-tau, 5% GS, Cat. no. MN1020; Thermo Fisher Scientific).
Immunohistochemistry was then performed using biotinylated
secondary anti-rabbit IgG, and avidin-biotinylated peroxidase
complex (ABC kit) (both from Vector Laboratories, Burlingame, CA,
USA), and 3,3′-diaminobenzidine (Sigma-Aldrich). Sections
containing the parietal cortex were selected and the number of
plaques in the cortex was counted under a microscope (Olympus
Corp., Tokyo, Japan). The assessments of Bam10-stained areas in the
cortex were performed using the MetaMorph image analyzer (Molecular
Devices Inc., Sunnyvale, CA, USA).
Western blot analysis
Western blot analysis was performed as described in
a previous study (25). The mice
were sacrificed and the brain tissues were rapidly removed and
homogenized in homogenization buffer (50 mM Tris-HCl, pH 8.0, 150
mM NaCl, 1% Nonidet P-40, 0.1% sodium dodecyl sulfate, and 0.1%
sodium deoxycholate) containing a cocktail of protease inhibitors
(Roche, Mannheim, Germany). Protein samples were resolved by
performing sodium dodecyl sulfate-polyacrylamide gel
electrophoresis, and were then transferred onto polyvinylidene
fluoride membranes (Bio-Rad Laboratories, Inc., Hercules, CA, USA).
The blots were incubated with primary antibodies followed by
secondary antibodies, and specific signals were visualized using an
enhanced chemiluminescence kit (Intron Biotechnology, Seoul,
Korea). The western blot images were quantified using Quantity One
1-D analysis software version 4.6.1 (Bio-Rad Laboratories, Inc.).
The primary antibodies were C-terminal fragments (CTFs, Cat.no.
A8717; Sigma-Aldrich), phospho-amyloid precursor protein (p-Thr668
APP, Cat.no. 6986; Cell Signaling Technology, Inc., Danvers, MA,
USA), a disintegrin and metalloproteinase 10 (ADAM10, Cat. no.
AB1997) and beta-site APP-cleaving enzyme 1 (BACE1, Cat. no.
AB2077) (both from Abcam, Cambridge, UK), neprilysin (NEP, Cat. no.
SC-9149; Santa Cruz Biotechnology, Inc., Dallas, TX, USA), insulin
degrading enzyme (IDE, Cat. no. 97-273; Calbiochem, Darmstadt,
Germany), vesicular glutamate transporter 1 (vGlut1, Cat. no. 135
302; Synaptic Systems, Göttingen, Germany), N-methyl-D-aspartate
receptor subtype 2B (NR2B, Cat. no. 06-600; Merck Millipore,
Darmstadt, Germany), synaptophysin (SYP, Cat. no. S5768;
Sigma-Aldrich), β-actin (Cat. no. MAB1501; Merck Millipore) and
post-synaptic density protein 95 (PSD-95, a gift from Dr J.-R Lee;
KRIBB). The secondary antibodies were goat anti-rabbit IgG (Cat.
no. sc-2004; Santa Cruz Biotechnology, Inc.) and goat anti-mouse
IgG (Cat. no. sc-2005; Santa Cruz Biotechnology, Inc.).
Statistical analysis
Two-sample comparisons were carried out using the
Student's t-test, while multiple comparisons were made using
one-way ANOVA followed by Tukey-Kramer's post hoc test. All data
are presented as the means ± SEM and statistical differences were
accepted at the 5% level (p<0.05) unless otherwise
indicated.
Results
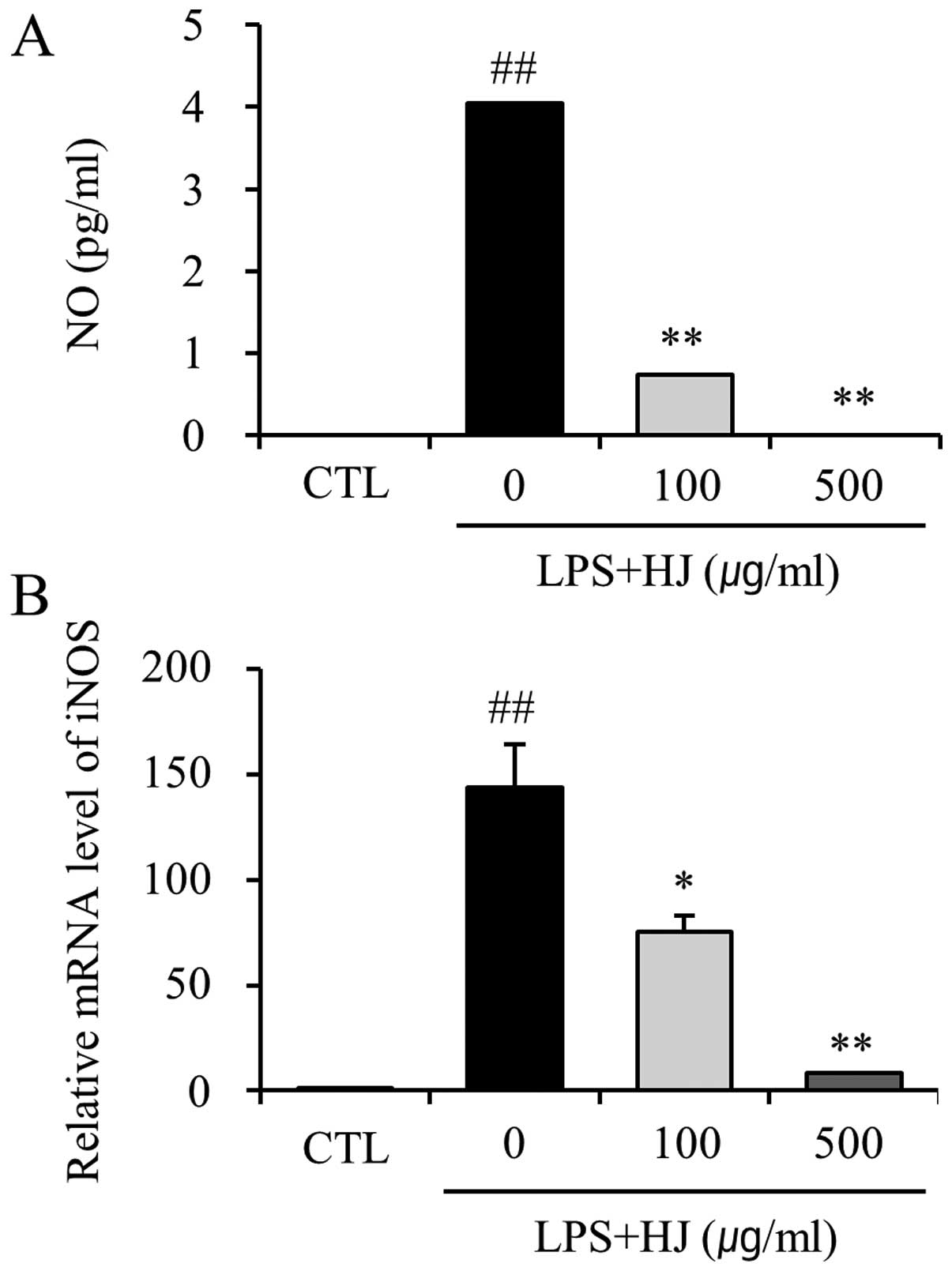
HJ suppresses LPS-induced NO production
by microglial cells
To examine the inhibitory effects of HJ on
LPS-induced NO production by BV2 cells, the NO levels in the
culture medium were measured by Griess reaction. Stimulation of the
BV2 cells with LPS caused a marked induction of NO production.
However, treatment with HJ resulted in the significant inhibition
of LPS-induced NO production (Fig.
1A). We also performed RT-qPCR to determine whether the
inhibition of NO production by HJ in the LPS-stimulated BV2 cells
is associated with the decreased levels of iNOS, which produces NO
as a key mediator of inflammation. Treatment with HJ resulted in a
significant decrease in the mRNA expression of iNOS in the
LPS-stimulated BV2 cells (Fig.
1B). These results indicated that the HJ-induced decrease in
the expression of iNOS is responsible for the inhibition of NO
production.
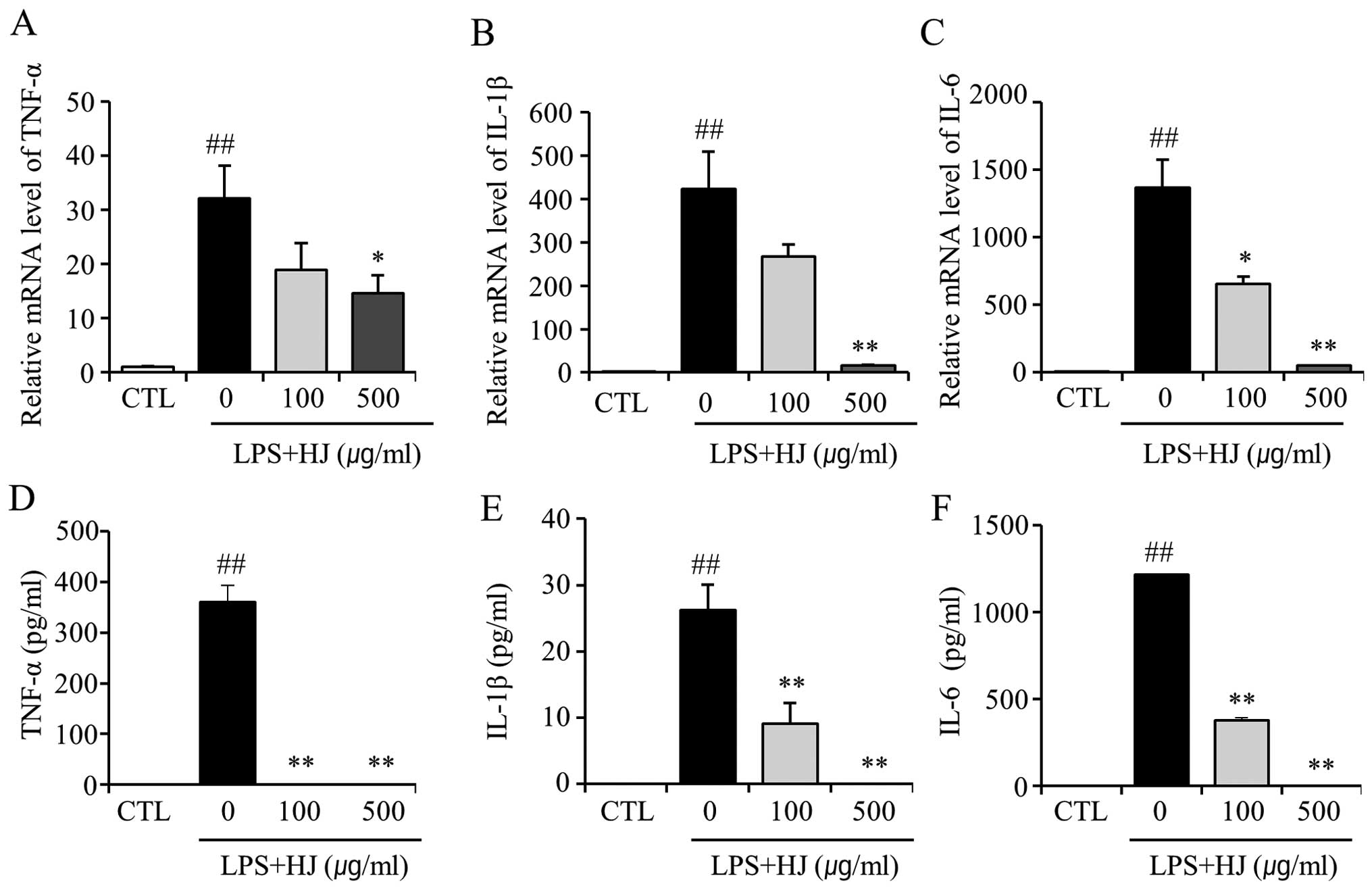
HJ decreases the mRNA expression and the
release of LPS-induced pro-inflammatory cytokines from microglial
cells
In order to investigate whether HJ inhibits the
expression of LPS-induced pro-inflammatory cytokines in the murine
microglial cell line, BV2, the cells were co-treated with LPS and
HJ for 24 h and the mRNA levels of TNF-α, IL-1β and IL-6 were then
analyzed by RT-qPCR. As shown in Fig.
2A–C, the LPS-induced increase in the expression of TNF-α,
IL-1β and IL-6 was significantly decreased by treatment with HJ.
The production of TNF-α, IL-1β and IL-6 induced by LPS in the
culture supernatants was evaluated by ELISA. The levels of TNF-α,
IL-1β and IL-6 were significantly increased in the culture medium
of LPS-stimulated BV2 cells; however, co-treatment with HJ resulted
in a marked decrease in the release of these pro-inflammatory
cytokines (Fig. 2D–F). These
results suggest that HJ is effective in the suppression of
pro-inflammatory cytokine production through the alteration of the
transcript levels of TNF-α, IL-1β and IL-6 in activated BV2
microglial cells.
HJ improves cognitive function in
Tg-APP/PS1 mice
Based on the in vitro anti-inflammatory
effects of HJ, we examined whether HJ has a therapeutic effect on
an animal model of AD. Tg-APP/PS1 mice were administered HJ at a
dose of 500 mg/kg/day for 2 months from 5 months of age. The body
weight of the HJ group during the treatment period was similar to
that of the controls (data not shown). To investigate whether the
treatment of Tg-APP/PS1 mice with HJ functionally attenuates
AD-type cognitive deterioration, we evaluated object recognition
memory (27) and spatial working
learning and memory (31) using
the object recognition test and spontaneous Y-maze alteration test
in HJ- or vehicle-treated mice. The HJ-treated Tg-APP/PS1 mice
exhibited increased exploration preference of the novel object
during the recognition session of the object recognition test
compared with vehicle-treated Tg-control mice (Fig. 3A). In the spontaneous Y-maze
alteration test, the HJ-treated Tg-APP/PS1 mice exhibited increased
alteration percentage compared with the vehicle-treated Tg-control
mice (Fig. 3B). This increase in
the percentage of spontaneous alterations was statistically
significant (p<0.05). These results indicated that treatment
with HJ for 2 months improved the cognitive decline of Tg-APP/PS1
mice.
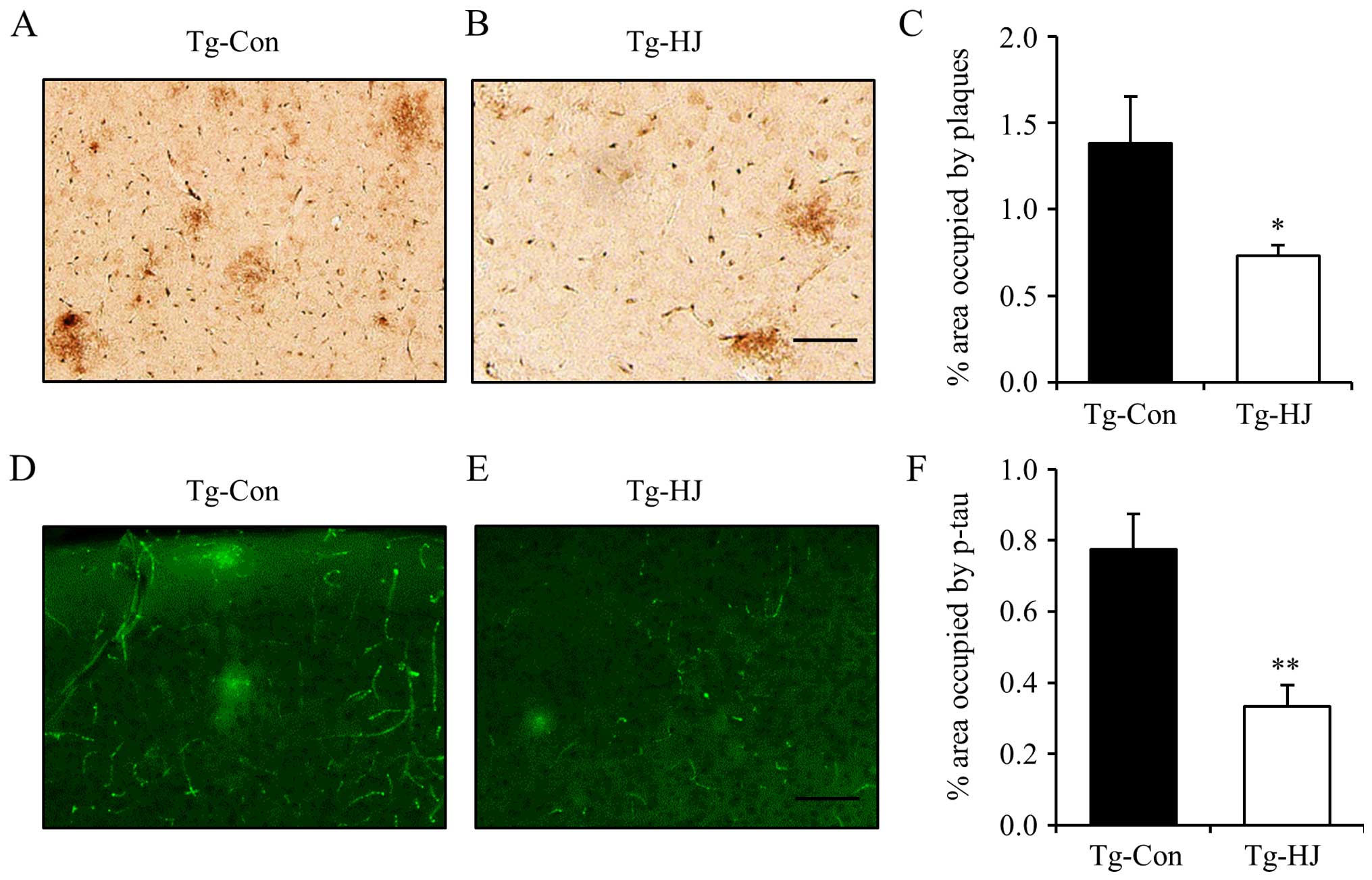
HJ reduces the Aβ level and tau
phosphorylation in the brains of Tg-APP/PS1 mice
Subsequently, to investigate whether HJ can
attenuate the levels of Aβ and tau hyperphosphorylation in
Tg-APP/PS1 mice, we measured the percentage area occupied by
plaques and phospho-tau in the cortex by immunohistochemical assay.
The Tg-APP/PS1 mice were administered HJ at a dose of 500 mg/kg/day
for 2.5 months from 5 months of age. The administration of HJ
effectively decreased the percentage area occupied by amyloid
plaques in the cortex (Fig. 4A–C;
p<0.05). Tau hyperphosphorylation was also significantly
decreased in the HJ-treated Tg-APP/PS1 mice compared with the
vehicle-treated Tg-control mice (Fig.
4D–F; p<0.01).
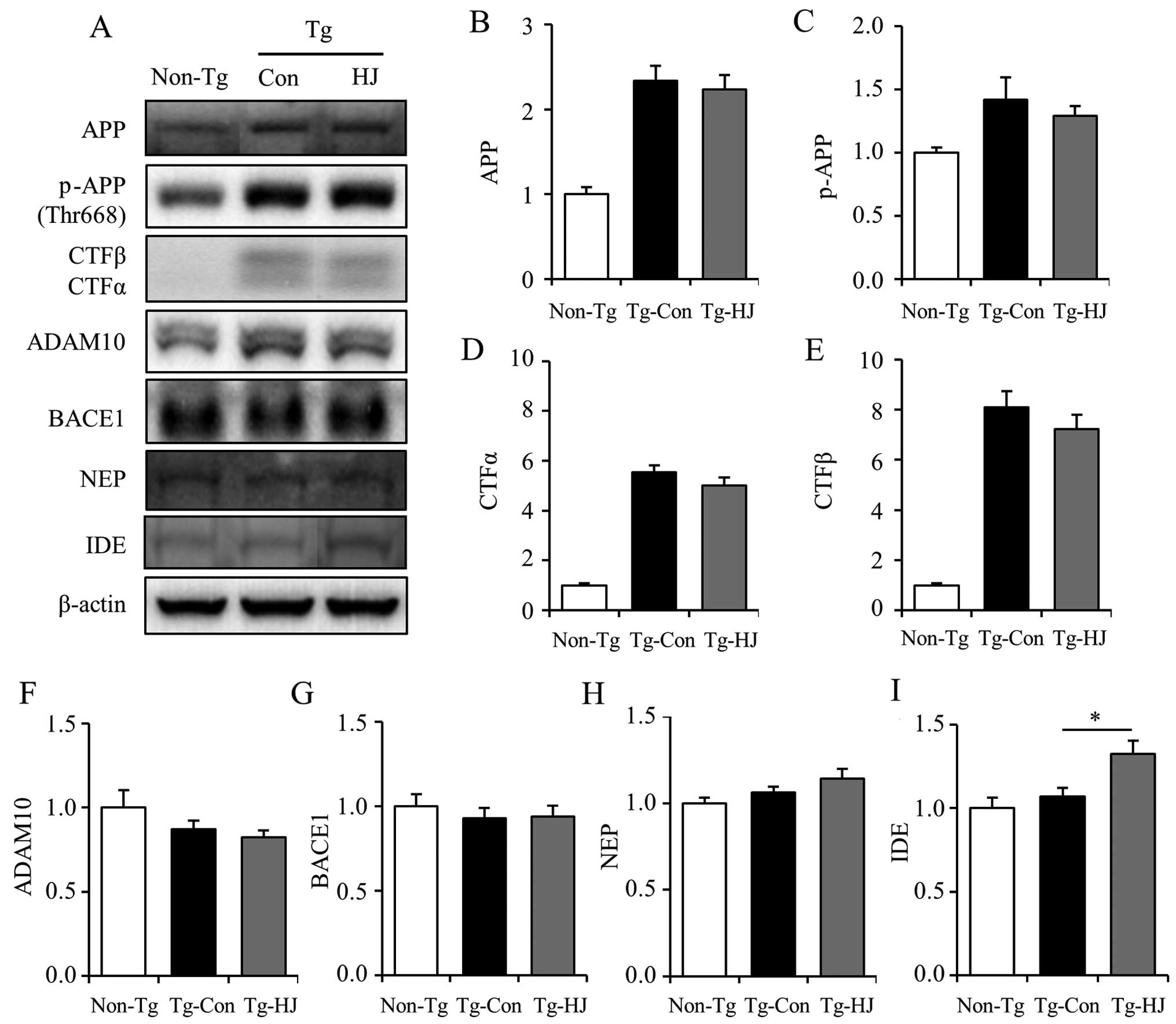
HJ increases IDE protein expression for
Aβ clearance in the brains of Tg-APP/PS1 mice
In order to further investigate which factors
contributed to the reduction of Aβ in HJ-treated Tg-APP/PS1 mice,
the changes in APP processing and Aβ clearance-related factors were
analyzed using brain tissues from both groups of mice. As shown by
the results of western blot analysis, HJ had no effect on the
protein expression of total APP (Fig.
5A and B), phospho-APP (Fig. 5A
and C), and the β-cleavage product (CTFβ) and α-cleavage
product (CTF α) (Fig. 5A, D and
E). In addition, treatment with HJ did not alter the levels of
ADAM10 (α-secretase) and BACE1 (β-secretase) (Fig. 5A, F and G). Moreover, since the
proteolytic degradation of Aβ is known to be a major route for the
clearance of plaques and neprilysin, and insulin degrading enzyme
and major Aβ-degrading metalloproteases are involved in the
downregulation of the levels of Aβ secreted extracellularly
(32,33), the protein levels of NEP and IDE
in brain tissues were determined in both groups. Immunoreactivity
assessed by western blot analysis of IDE expression was
significantly increased in the HJ-treated Tg-APP/PS1 mice (Fig. 5A and I), but the level of NEP was
not altered (Fig. 5A and H).
These results suggest that HJ has an effect on Aβ clearance through
the increased level of IDE, but not by APP processing.
HJ attenuates neuroinflammation in the
brains of Tg-APP/PS1 mice
Previously, it has been reported that Aβ is able to
initiate inflammation, which implicates the activation of microglia
and the recruitment of astrocytes, and the release of cytokines,
such as TNF-α, interferon-γ, IL-1β, IL-1α and COX-2, and ROS that
are involved in neuronal and synaptic damage (4,34).
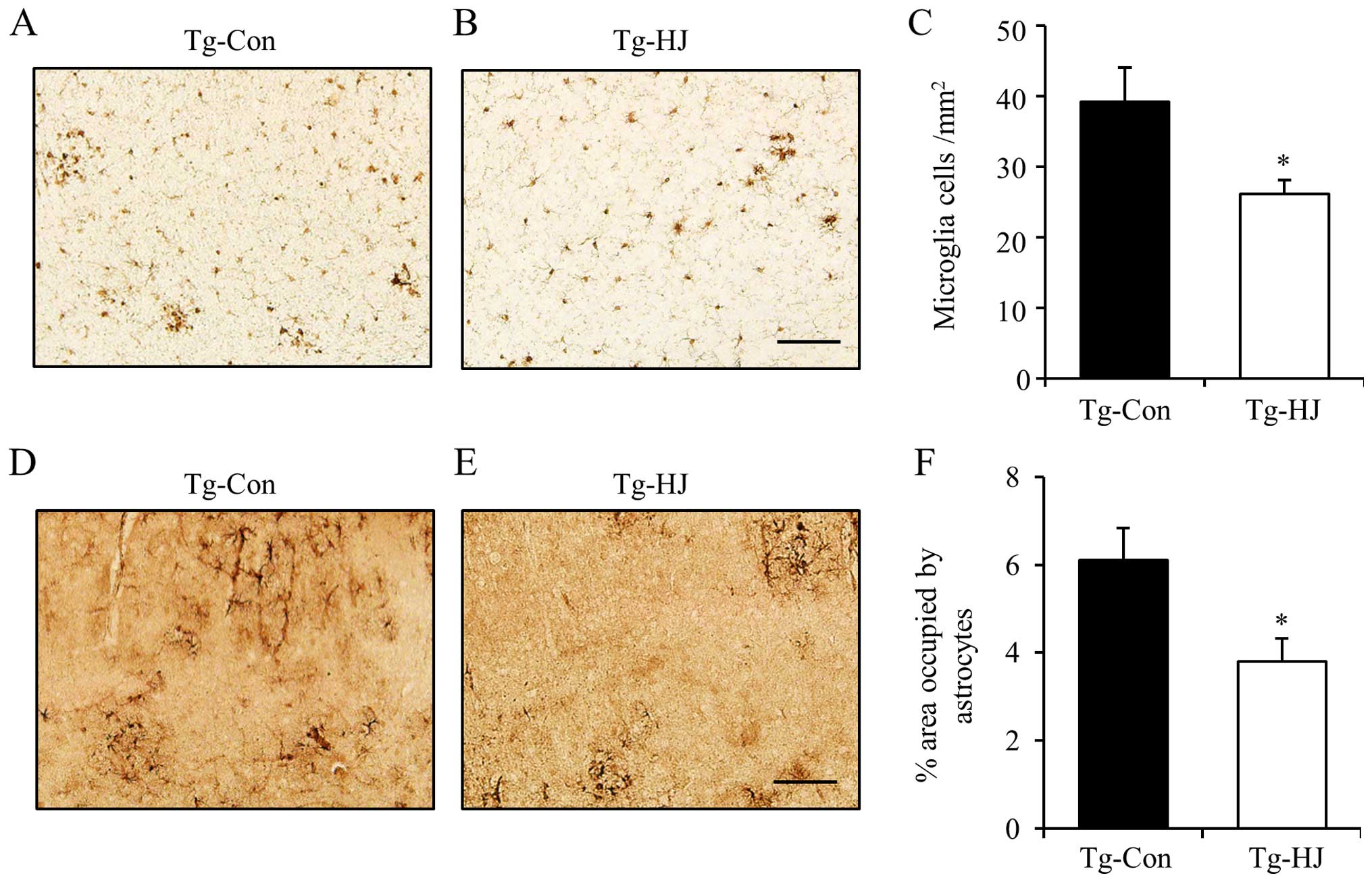
Thus, to investigate whether HJ can reduce the activation of
microglia and astrocytes in Tg-APP/PS1 mice, we measured the number
of Iba-1 immunoreactive cells and the areas of GFAP
immunoreactivity in the cortex (Fig.
6). The number of Iba-1-positive cells was significantly
decreased in the cortex tissues of the HJ-treated Tg-APP/PS1 mice
(Fig. 6A–C; p<0.05). Moreover,
the percentage area occupied by active astrocytes was also
significantly decreased in the cortex tissues of the HJ-treated
Tg-APP/PS1 mice (Fig. 6D–F;
p<0.05). Additionally, the mRNA levels of TNF-α and IL-6 in the
cortex homogenates in the HJ-treated Tg-APP/PS1 group were
significantly lower than those in the Tg-APP/P1 control group
(Fig. 7A and B; p<0.05 and
p<0.01). However, the mRNA expression levels of IL-1β in the
HJ-treated Tg-APP/PS1 group were not significantly altered
(Fig. 7C). These findings
indicated that the activation of microglia and astrocytes, and the
expression levels of the pro-inflammatory cytokines, TNF-α and
IL-6, were markedly reduced by treatment with HJ for 2.5 months in
the animal model of AD. These results suggest that treatment with
HJ effectively attenuates neuroinflammation in the brains of
Tg-APP/PS1 mice.
Discussion
In the present study, we demonstrated that HJ
inhibits inflammation in BV2 microglial cells. HJ significantly
decreased the LPS-stimulated production of pro-inflammatory
cytokines, such as TNF-α, IL-1β and IL-6, as well as the production
of the pro-inflammatory mediator, NO. HJ improved object
recognition and spatial working memory in a mouse model of AD, and
reduced Aβ deposition and tau hyperphosphorylation in the cortex
tissues of Tg-APP/PS1 mice. We found that treatment with HJ
increased the protein expression of IDE, which is one of the
degrading enzymes of Aβ, in the cortex tissues of Tg-APP/PS1 mice.
Further, we observed that HJ has anti-inflammatory activities, such
as the attenuation of the activation of microglia and astrocytes,
and the decreased mRNA levels of TNF-α and IL-6 in the cortex
tissues of HJ-treated Tg-APP/PS1 mice.
In the present study, to investigate the
anti-inflammatory effects of HJ on LPS-activated BV2 microglia
cells, the effects of HJ on the expression and production of
pro-inflammatory cytokines, and the expression of iNOS and the
production of NO were determined. The results revealed that HJ
inhibited the LPS-induced increase in the expression of IL-1β,
IL-6, TNF-α, and iNOS in the BV2 cells. The production of IL-1β,
IL-6, TNF-α and NO in the supernatant of the BV2 cells was also
markedly inhibited by treatment with HJ. Furthermore, the
anti-inflammatory activities of HJ were revealed in HJ-treated
Tg-APP/PS1 mice; the activation of microglia and astrocytes, and
the mRNA expression of TNF-α and IL-6, the major cytokines in the
process of AD pathogenesis, was markedly decreased in the cortex
tissues of HJ-treated Tg-APP/PS1 mice. Chronic neuroinflammation
induced by pro-inflammatory cytokines released from activated
microglia and astrocytes has been recognized as one of the major
mechanisms of AD pathophysiology (3). Aβ plaques and tangles in the brains
of patients with AD stimulate a chronic inflammatory reaction to
clear this debris (35–37). However, chronically activated glia
can contribute to neuronal dysfunction and cell death by releasing
highly toxic products, such as ROS, NO and excitatory amino acids
(38,39). In addition, γ-aminobutyric acid
released from reactive and activated astrocytes in AD can impair
synaptic plasticity, and learning and memory in AD (40). A number of cytokines, including
IL-1α, IL-1β, IL-6, TNF-α and IL-8 are increased in AD-affected d
brain tissue (41). Many
compounds that alleviate AD-type pathology and cognitive deficits
inhibit the expression and release of these cytokines (15–18,36,42). Based on these studies, the
anti-inflammatory effects of HJ may attenuate Aβ deposition, tau
hyperphosphorylation, and cognitive deterioration in an animal
model of AD.
Aβ is known to be generated by sequential
proteolysis of APP, a large integral membrane protein that is
cleaved by BACE1 at first, followed by CTFβ in the membrane and by
γ-secretase in the transmembrane domain (43,44). The production of Aβ is avoided by
an alternate APP cleavage pathway mediated by the α-secretase
followed by γ-secretase (43,44). HJ had no effect on the expression
of total APP, CTFα, and APP cleavage form mediated by the
α-secretase and CTFβ. The expression of BACE1 and ADAM10 was not
altered by treatment with HJ. These results suggest that HJ has no
effect on APP processing in Tg-APP/PS1 mice.
The degradation and clearance of amyloid plaques, as
well as alternative APP processing is an interesting therapeutic
approach, and the proteases, NEP and IDE, have shown to cleave Aβ
(45–47). In our study, IDE protein levels in
the cortex tissues of Tg-APP/PS1 mice were significantly increased
by treatment with HJ; however, the NEP protein levels were not
altered in the HJ-treated Tg-APP/PS1 mice. IDE is the main
extracellular protease degrading Aβ (33). The increased expression of IDE
reduces the Aβ level and inhibits amyloid plaque formation
(46,47), and the genetic inactivation of IDE
in mice increases brain Aβ levels (48). IDE is expressed and released by
microglia and neurons (33,47). A decrease in IDE levels with the
concomitant upregulation of TNF-α and IL-1β was observed in
8-month-old Tg-APP/PS1 mice, and the inhibition of microglial
activation by donepezil has been shown to lead to alterations in
IDE levels (42). Unlike previous
studies (42,49), we observed that IDE expression in
the cortex tissues of Tg-APP/PS1 mice was not decreased (Fig. 5), but treatment with HJ increased
IDE protein levels. Therefore, it is conceivable that this
increased IDE expression may enhance Aβ clearance.
Our in vitro and in vivo analysis with
microglial cells and an animal model of AD demonstrated that HJ has
strong anti-inflammatory properties, and attenuates AD
pathophysiology in the CNS. The anti-inflammatory capability of HJ
may contribute to its anti-microbial, anti-mycobacterial and
anticancer effects (20,21,50,51). Although it was not verified in
this study, previous studies have shown that HJ has an antioxidant
activity in vitro (20,21,51). The reduction in cellular
expression and activity of antioxidant proteins and the resulting
increased oxidative stress can contribute to the progression of
brain aging and the development of neurodegenerative diseases,
including AD (52). These data
suggest that this natural plant, HJ, may prove to be useful in the
treatment of inflammatory diseases and several neurodegenerative
diseases that are associated with microglial cell activation and
oxidative stress.
Abbreviations:
|
Aβ
|
amyloid β
|
|
AD
|
alzheimer's disease
|
|
ADAM10
|
a disintegrin and metalloproteinase
10
|
|
BACE1
|
beta-site APP-cleaving enzyme 1
|
|
IDE
|
insulin degrading enzyme
|
|
IL-1β
|
interleukin-1β
|
|
IL-6
|
interleukin-6
|
|
iNOS
|
inducible nitric oxide synthase
|
|
NEP
|
neprilysin
|
|
TNF-α
|
tumor necrosis factor α
|
Acknowledgments
This study was supported by KRIBB Research
Initiative Program of the Republic of Korea. The authors would like
to thank Mr. In-Bok Lee, Ms. Jung-Hyun Choi and Ms. Yun-Jeong Seo
for their technical assistance.
References
|
1
|
Ali FEA, Barnham KJ, Barrow CJ and
Separovic F: Metal-catalyzed oxidative damage and oligomerization
of the amyloid-peptide of Alzheimer's disease. Aust J Chem.
57:511–518. 2004. View
Article : Google Scholar
|
|
2
|
Baldeiras I, Santana I, Proença MT,
Garrucho MH, Pascoal R, Rodrigues A, Duro D and Oliveira CR:
Oxidative damage and progression to Alzheimer's disease in patients
with mild cognitive impairment. J Alzheimers Dis. 21:1165–1177.
2010.PubMed/NCBI
|
|
3
|
Zhang F and Jiang L: Neuroinflammation in
Alzheimer's disease. Neuropsychiatr Dis Treat. 11:243–256. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Birch AM, Katsouri L and Sastre M:
Modulation of inflammation in transgenic models of Alzheimer's
disease. J Neuroinflammation. 11:252014. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Finch CE and Morgan TE: Systemic
inflammation, infection, ApoE alleles, and Alzheimer disease: a
position paper. Curr Alzheimer Res. 4:185–189. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Frank B and Gupta S: A review of
antioxidants and Alzheimer's disease. Ann Clin Psychiatry.
17:269–286. 2005. View Article : Google Scholar
|
|
7
|
Zhu X, Su B, Wang X, Smith MA and Perry G:
Causes of oxidative stress in Alzheimer disease. Cell Mol Life Sci.
64:2202–2210. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Heneka MT and O'Banion MK: Inflammatory
processes in Alzheimer's disease. J Neuroimmunol. 184:69–91. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
El Khoury J and Luster AD: Mechanisms of
microglia accumulation in Alzheimer's disease: therapeutic
implications. Trends Pharmacol Sci. 29:626–632. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Mandrekar-Colucci S, Karlo JC and Landreth
GE: Mechanisms underlying the rapid peroxisome
proliferator-activated receptor-γ-mediated amyloid clearance and
reversal of cognitive deficits in a murine model of Alzheimer's
disease. J Neurosci. 32:10117–10128. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Noble W, Garwood C, Stephenson J, Kinsey
AM, Hanger DP and Anderton BH: Minocycline reduces the development
of abnormal tau species in models of Alzheimer's disease. FASEB J.
23:739–750. 2009. View Article : Google Scholar
|
|
12
|
Sastre M and Gentleman SM: NSAIDs: how
they work and their prospects as therapeutics in Alzheimer's
disease. Front Aging Neurosci. 2:202010.PubMed/NCBI
|
|
13
|
Seabrook TJ, Jiang L, Maier M and Lemere
CA: Minocycline affects microglia activation, Abeta deposition, and
behavior in APP-tg mice. Glia. 53:776–782. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Tweedie D, Ferguson RA, Fishman K,
Frankola KA, Van Praag H, Holloway HW, Luo W, Li Y, Caracciolo L,
Russo I, et al: Tumor necrosis factor-α synthesis inhibitor
3,6′-dithiothalidomide attenuates markers of inflammation,
Alzheimer pathology and behavioral deficits in animal models of
neuroinflammation and Alzheimer's disease. J Neuroinflammation.
9:1062012. View Article : Google Scholar
|
|
15
|
Chan MM, Huang HI, Fenton MR and Fong D:
In vivo inhibition of nitric oxide synthase gene expression by
curcumin, a cancer preventive natural product with
anti-inflammatory properties. Biochem Pharmacol. 55:1955–1962.
1998. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Wang J, Ho L, Zhao W, Ono K, Rosensweig C,
Chen L, Humala N, Teplow DB and Pasinetti GM: Grape-derived
polyphenolics prevent Abeta oligomerization and attenuate cognitive
deterioration in a mouse model of Alzheimer's disease. J Neurosci.
28:6388–6392. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Xia EQ, Deng GF, Guo YJ and Li HB:
Biological activities of polyphenols from grapes. Int J Mol Sci.
11:622–646. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Yang F, Lim GP, Begum AN, Ubeda OJ,
Simmons MR, Ambegaokar SS, Chen PP, Kayed R, Glabe CG, Frautschy SA
and Cole GM: Curcumin inhibits formation of amyloid beta oligomers
and fibrils, binds plaques, and reduces amyloid in vivo. J Biol
Chem. 280:5892–5901. 2005. View Article : Google Scholar
|
|
19
|
Jeong KY, Han IS, Choi SY, Lee JH, Lee JS,
Hong CS and Park JW: Allergenicity of recombinant profilins from
Japanese hop, Humulus japonicus. J Investig Allergol Clin Immunol.
23:345–350. 2013.PubMed/NCBI
|
|
20
|
Lee YR, Kim KY, Lee SH, Kim MY, Park HJ
and Jeong HS: Antioxidant and antitumor activities of methanolic
extracts from Humulus japonicus. Korean J Food Nutr. 25:357–361.
2012. View Article : Google Scholar
|
|
21
|
Park SW, Woo CJ, Chung SK and Chung KT:
Antimicrobial and antioxidative activities of solvent fraction from
Humulus japonicus. Korean J Food Sci Technol. 26:464–470. 1994.
|
|
22
|
Hwang SY, Jo MJ, Kim SC and Jee SY:
Anti-inflammaory effects of the MeOH extract of Humulus japonicus
in vivo. J Korean Orient Med Ophthalmol Otolaryngol Derm.
22:92–103. 2009.
|
|
23
|
Jankowsky JL, Fadale DJ, Anderson J, Xu
GM, Gonzales V, Jenkins NA, Copeland NG, Lee MK, Younkin LH, Wagner
SL, et al: Mutant presenilins specifically elevate the levels of
the 42 residue beta-amyloid peptide in vivo: evidence for
augmentation of a 42-specific gamma secretase. Hum Mol Genet.
13:159–170. 2004. View Article : Google Scholar
|
|
24
|
Jankowsky JL, Slunt HH, Ratovitski T,
Jenkins NA, Copeland NG and Borchelt DR: Co-expression of multiple
transgenes in mouse CNS: a comparison of strategies. Biomol Eng.
17:157–165. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Park HY, Kang YM, Kang Y, Park TS, Ryu YK,
Hwang JH, Kim YH, Chung BH, Nam KH, Kim MR, et al: Inhibition of
adenylyl cyclase type 5 prevents L-DOPA-induced dyskinesia in an
animal model of Parkinson's disease. J Neurosci. 34:11744–11753.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Akkerman S, Blokland A, Reneerkens O, van
Goethem NP, Bollen E, Gijselaers HJ, Lieben CK, Steinbusch HW and
Prickaerts J: Object recognition testing: methodological
considerations on exploration and discrimination measures. Behav
Brain Res. 232:335–347. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Antunes M and Biala G: The novel object
recognition memory: neurobiology, test procedure, and its
modifications. Cogn Process. 13:93–110. 2012. View Article : Google Scholar :
|
|
28
|
Wasilewska-Sampaio AP, Santos TG, Lopes
MH, Cammarota M and Martins VR: The growth of glioblastoma
orthotopic xenografts in nude mice is directly correlated with
impaired object recognition memory. Physiol Behav. 123:55–61. 2014.
View Article : Google Scholar
|
|
29
|
Belforte JE, Zsiros V, Sklar ER, Jiang Z,
Yu G, Li Y, Quinlan EM and Nakazawa K: Postnatal NMDA receptor
ablation in cortico-limbic interneurons confers schizophrenia-like
phenotypes. Nat Neurosci. 13:76–83. 2010. View Article : Google Scholar
|
|
30
|
Kim KS, Kang YM, Kang Y, Park TS, Park HY,
Kim YJ, Han BS, Kim CH, Lee CH, Ardayfio PA, et al: Pitx3 deficient
mice as a genetic animal model of co-morbid depressive disorder and
parkinsonism. Brain Res. 1552:72–81. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Sierksma AS, Prickaerts J, Chouliaras L,
Rostamian S, Delbroek L, Rutten BP, Steinbusch HW and van den Hove
DL: Behavioral and neurobiological effects of prenatal stress
exposure in male and female APPswe/PS1dE9 mice. Neurobiol Aging.
34:319–337. 2013. View Article : Google Scholar
|
|
32
|
Hafez D, Huang JY, Huynh AM, Valtierra S,
Rockenstein E, Bruno AM, Lu B, DesGroseillers L, Masliah E and Marr
RA: Neprilysin-2 is an important β-amyloid degrading enzyme. Am J
Pathol. 178:306–312. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Qiu WQ, Walsh DM, Ye Z, Vekrellis K, Zhang
J, Podlisny MB, Rosner MR, Safavi A, Hersh LB and Selkoe DJ:
Insulin-degrading enzyme regulates extracellular levels of amyloid
beta-protein by degradation. J Biol Chem. 273:32730–32738. 1998.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Sastre M, Klockgether T and Heneka MT:
Contribution of inflammatory processes to Alzheimer's disease:
molecular mechanisms. Int J Dev Neurosci. 24:167–176. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
McGeer PL and McGeer EG: The inflammatory
response system of brain: implications for therapy of Alzheimer and
other neurodegenerative diseases. Brain Res Brain Res Rev.
21:195–218. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Rubio-Perez JM: A review: inflammatory
process in Alzheimer's disease, role of cytokines. ScientificWord
Journal. 2012:7563572012.
|
|
37
|
Town T, Nikolic V and Tan J: The
microglial 'activation' continuum: from innate to adaptive
responses. J Neuroinflammation. 2:242005. View Article : Google Scholar
|
|
38
|
Brown GC and Bal-Price A: Inflammatory
neurodegeneration mediated by nitric oxide, glutamate, and
mitochondria. Mol Neurobiol. 27:325–355. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Halliday G, Robinson SR, Shepherd C and
Kril J: Alzheimer's disease and inflammation: a review of cellular
and therapeutic mechanisms. Clin Exp Pharmacol Physiol. 27:1–8.
2000. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Jo S, Yarishkin O, Hwang YJ, Chun YE, Park
M, Woo DH, Bae JY, Kim T, Lee J, Chun H, et al: GABA from reactive
astrocytes impairs memory in mouse models of Alzheimer's disease.
Nat Med. 20:886–896. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Akiyama H, Barger S, Barnum S, Bradt B,
Bauer J, Cole GM, Cooper NR, Eikelenboom P, Emmerling M, Fiebich
BL, et al: Inflammation and Alzheimer's disease. Neurobiol Aging.
21:383–421. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Guo HB, Cheng YF, Wu JG, Wang CM, Wang HT,
Zhang C, Qiu ZK and Xu JP: Donepezil improves learning and memory
deficits in APP/PS1 mice by inhibition of microglial activation.
Neuroscience. 290:530–542. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Edbauer D, Winkler E, Regula JT, Pesold B,
Steiner H and Haass C: Reconstitution of gamma-secretase activity.
Nat Cell Biol. 5:486–488. 2003. View
Article : Google Scholar : PubMed/NCBI
|
|
44
|
Vassar R, Bennett BD, Babu-Khan S, Kahn S,
Mendiaz EA, Denis P, Teplow DB, Ross S, Amarante P, Loeloff R, et
al: Beta-secretase cleavage of Alzheimer's amyloid precursor
protein by the transmembrane aspartic protease BACE. Science.
286:735–741. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Iwata N, Tsubuki S, Takaki Y, Watanabe K,
Sekiguchi M, Hosoki E, Kawashima-Morishima M, Lee HJ, Hama E,
Sekine-Aizawa Y and Saido TC: Identification of the major
Abeta1-42-degrading catabolic pathway in brain parenchyma:
suppression leads to biochemical and pathological deposition. Nat
Med. 6:143–150. 2000. View
Article : Google Scholar : PubMed/NCBI
|
|
46
|
Leissring MA, Farris W, Chang AY, Walsh
DM, Wu X, Sun X, Frosch MP and Selkoe DJ: Enhanced proteolysis of
beta-amyloid in APP transgenic mice prevents plaque formation,
secondary pathology, and premature death. Neuron. 40:1087–1093.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Vekrellis K, Ye Z, Qiu WQ, Walsh D,
Hartley D, Chesneau V, Rosner MR and Selkoe DJ: Neurons regulate
extracellular levels of amyloid beta-protein via proteolysis by
insulin-degrading enzyme. J Neurosci. 20:1657–1665. 2000.PubMed/NCBI
|
|
48
|
Leal MC, Dorfman VB, Gamba AF, Frangione
B, Wisniewski T, Castaño EM, Sigurdsson EM and Morelli L:
Plaque-associated overexpression of insulin-degrading enzyme in the
cerebral cortex of aged transgenic tg2576 mice with Alzheimer
pathology. J Neuropathol Exp Neurol. 65:976–987. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Cook DG, Leverenz JB, McMillan PJ, Kulstad
JJ, Ericksen S, Roth RA, Schellenberg GD, Jin LW, Kovacina KS and
Craft S: Reduced hippocampal insulin-degrading enzyme in late-onset
Alzheimer's disease is associated with the apolipoprotein
E-epsilon4 allele. Am J Pathol. 162:313–319. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Hong MS, Son ES, Lee SJ, Lee SK, Lee YJ,
Song SD, Cho SN, Barry CE III and Eum SY: Anti-mycobacterial
Effects of the Extract of Humulus japonicas. Korean J Food Sci
Technol. 46:94–99. 2014. View Article : Google Scholar
|
|
51
|
Park SW, Kim SH and Chung SK:
Antimutagenic effects and isolation of flavonoids from HJ extract.
Korean J Food Sci Technol. 27:897–901. 1995.
|
|
52
|
Di Bona D, Scapagnini G, Candore G,
Castiglia L, Colonna-Romano G, Duro G, Nuzzo D, Iemolo F, Lio D,
Pellicanò M, et al: Immune-inflammatory responses and oxidative
stress in Alzheimer's disease: therapeutic implications. Curr Pharm
Des. 16:684–691. 2010. View Article : Google Scholar : PubMed/NCBI
|