Introduction
Hepatocellular carcinoma (HCC) is one of the most
frequent and most lethal types of cancer worldwide with hardly any
effective treatment available thus far (1). The major risk factors for the
disease vary, including hepatitis B or C virus infection, alcoholic
liver disease, and non-alcoholic fatty liver disease. There is
still no systemic treatment available to effectively treat HCC
(2). Accumulating evidence
suggests that the loss of certain tumor suppressors and the
aberrant regulation of cellular growth signaling, such as the
extracellular signal-regulated kinase (ERK)/mitogen-activated
protein kinase (MAPK) pathway and Wnt/β-catenin pathway, are
associated with liver tumorigenesis; however, the definite
molecular pathogenesis of HCC remains poorly understood (3). Recently, the aberrant regulation of
non-coding RNAs (ncRNAs) has been proposed to be associated with
hepatocarcinogenesis. These ncRNAs can be further subdivided into
two major categories, long non-coding RNAs and short non-coding
RNAs (4). To date, the most
extensively studied small RNAs in cancer are the microRNAs (miRNAs
or miRs).
miRNAs are short non-coding RNAs, approximately
21–25 nucleotides in length, which target mRNAs for degradation or
translational repression by direct binding to their 3′-untranslated
region (3′-UTR) (5). The
involvement of miRNAs in cancer pathogenesis is well established,
as miRNAs can function as either oncogenes or tumor suppressor
genes, depending on the cellular function of their targets.
Moreover, the activation or suppression of specific miRNAs is
established through oncogenes, such as myc, or tumor suppressor
genes, such as p53, which can induce or inhibit tumorigenesis.
Although their functions remain elusive, it is evident that miRNAs
play an important role in cancer initiation and progression via the
regulation of tumor suppressor genes or oncogenes (6). Previous studies have now linked the
deregulation of miRNA expression to the development of liver
cancer. The downregulation of miRNAs is observed in liver cancer
and the target genes regulated by these miRNAs possibly act as
oncogenes. On the contrary, overexpressed miRNAs may have oncogenic
functions by targeting tumor suppressors in HCC. For instance, some
miRNAs, such as miR-29, miR-21 and miR-221, have been reported to
regulate tumor cell growth, apoptosis, migration and invasion by
targeting proteins involved in those cellular pathways (7–9).
However, the roles of miRNAs in hepatic carcinogenesis are complex;
different studies have reported unique profiles, with only a few
miRNAs in common, indicating the heterogeneity of HCC. For example,
it has been shown that miR-26a inhibits (10), whereas miR-17-5p promotes tumor
growth and metastasis in HCC (11).
miR-31 functions as an antitumor miRNA and presents
distinct expression patterns in different human cancers. miR-31 has
been found to be significantly overexpressed in colorectal cancer
(12), squamous cell carcinoma of
the tongue (13), head and neck
squamous cell carcinoma (14),
lung cancer (15) and esophageal
squamous cell carcinoma (16); it
has been found to be downregulated in bladder tumors (17), prostate carcinoma (18), gastric cancer (19), HCC (20), breast cancer (21) and serous ovarian carcinoma
(22). Studies have suggested
that Kruppel (or Krüppel)-like factor 4 (Klf4), a zinc-finger
transcription factor, functions as a tumor suppressor or an
oncogene. It plays an important role in cell proliferation and
metastasis by regulating the expression of a number of downstream
target genes (23). Accumulating
clinical, experimental and mechanistic evidence suggests that Klf4
functions as a tumor suppressor in various types of cancer,
including HCC (24–27). However, the mechanistic role of
Klf4 in HCC and its causal link to altered miR-31 function remain
undetermined to date.
In this study, we demonstrated that the loss of Klf4
and miR-31 are evident in human HCC, and the altered expression and
function of Klf4 and miR-31 contribute to liver carcinogenesis. In
the present study, we profiled 18 differentially regulated miRNAs,
one of these being miR-31 and aimed to determine the correlation
between Klf4 and miR-31 in HCC. miR-31 may serve as a potential
diagnostic marker and provide new insight into the pathogenesis of
HCC.
Materials and methods
Patients and tissue specimens
Fresh liver cancer tissue samples and neighboring
non-cancerous liver tissue samples were obtained from 52 patients
who underwent primary surgical resection of liver tumors at the
Shanghai Eastern Hepatobiliary Surgery Hospital affiliated to the
Second Military Medical University (Shanghai, China) between March
and September 2015. Non-cancerous tissues were obtained 5–10 cm
away from the primary tumor. Follwoing resection, all the samples
were snap-frozen in liquid nitrogen and stored at −80°C prior to
RNA extraction. Patients who received preoperative treatment were
excluded from the study. All included patients provided written
informed consent prior to enrollment. The protocols involving human
samples were conducted in conformity with the ethical principles of
research and approved by the Human Resources Ethics Committee of
Shanghai East Hospital affiliated to Tongji University (Shanghai,
China). The histopathological diagnosis of all samples was
respectively verified by two pathologists. The clinical staging was
based on the 7th edition of the AJCC Cancer Staging Manual. The
main demographic and clinicopathological characteristics of the
patients are presented in Table
I.
 | Table ICorrelation between the
clinicopathologic characteristics of the patients with
hepatocellular carcinoma and the expression of Klf4 and miR-31. |
Table I
Correlation between the
clinicopathologic characteristics of the patients with
hepatocellular carcinoma and the expression of Klf4 and miR-31.
|
Characteristics | n | Klf4
| miR-31
|
|---|
| High, n (%) | Low, n (%) | P-value | High, n (%) | Low, n (%) | P-value |
|---|
| Gender | | | | | | | |
| Male | 44 | 19 (43.2) | 25 (56.8) | 0.100 | 13 (29.5) | 31 (70.1) | 0.072 |
| Female | 8 | 1
(12.5) | 7
(87.5) | | 5
(62.5) | 3
(37.5) | |
| Age (years) | | | | | | | |
| ≥60 | 13 | 5
(38.5) | 8
(61.5) | 1.000 | 3
(23.1) | 10 (76.9) | 0.016 |
| <60 | 39 | 15 (38.5) | 24 (61.5) | | 24 (61.5) | 15 (38.5) | |
|
Differentiation | | | | | | | |
| Poorly | 45 | 18 (40.0) | 27 (60.0) | 0.563 | 14 (31.1) | 31 (68.9) | 0.178 |
| Moderate/well | 7 | 2
(28.6) | 5
(71.4) | | 4
(57.1) | 3
(42.9) | |
| Clinical stage | | | | | | | |
| I–II | 14 | 9
(64.3) | 5
(35.7) | 0.020 | 9
(64.3) | 3
(21.4) | 0.009 |
| III–IV | 38 | 11 (28.9) | 27 (71.1) | | 13 (34.2) | 27 (71.1) | |
| T
classification | | | | | | | |
| T1-T2 | 11 | 8
(72.7) | 3
(27.3) | 0.029 | 4
(36.4) | 7
(63.6) | 0.535 |
| T3-T4 | 41 | 15 (36.6) | 26 (63.4) | | 11 (26.8) | 30 (73.2) | |
| Hepatitis B | | | | | | | |
| Negative | 6 | 4
(66.7) | 2
(33.4) | 0.043 | 3
(50.0) | 3
(50.0) | 0.400 |
| Positive | 46 | 12 (26.1) | 34 (73.9) | | 15 (32.6) | 31 (67.4) | |
| Pathological
types | | | | | | | |
| HCC | 38 | 16 (42.1) | 22 (57.9) | 0.374 | 11 (28.9) | 27 (71.1) | 0.157 |
| Other types | 14 | 4
(28.6) | 10 (71.4) | | 7
(50.0) | 7
(50.0) | |
| Tumor size | | | | | | | |
| ≥6 cm | 47 | 17 (36.2) | 30 (63.8) | 0.298 | 16 (34.0) | 31 (66.0) | 0.790 |
| <6 cm | 5 | 3
(60.0) | 2
(40.0) | | 2
(40.0) | 3
(60.0) | |
Cell lines and culture
The human HCC cell lines, Bel-7402, Sk-hep-1, Hep3B,
Huh7 and HepG2, were obtained from the Shanghai Cell Bank of
Chinese Academy of Sciences (Shanghai, China). The liver cell line,
L02, was purchased from Li Yandong Research Group Shanghai East
Hospital affiliated to Tongji University School of Medicine.
Each cell line was cultured in minimum essential
medium (MEM) supplemented with 10% fetal bovine serum (FBS;
HyClone, Logan, UT, USA) as well as 100 U/ml penicillin and 100
µg/ml streptomycin. All the above-mentioned cells were
maintained in a humidified incubator at 37°C with 5%
CO2.
Western blot analysis
Protein was extracted from the cells and tissues
using RIPA lysis buffer [1% NP40, 0.1% sodium dodecyl sulfate
(SDS), 100μg/ml phenylmethylsulfonyl fluoride, 0.5% sodium
deoxycholate, in PBS] on ice. The super-natants were collected
following centrifugation at 12,000 × g at 4°C for 20 min. The
protein concentration was determined using a BCA protein assay kit
(Bio-Rad, Shanghai, China), and whole lysates were mixed with 4X
SDS loading buffer [125 mmol/l Tris-HCl, 4% SDS, 20% glycerol, 100
mmol/l dithiothreitol (DTT) and 0.2% bromophenol blue] at a ratio
of 1:3. The samples were heated at 100°C for 5 min and were
separated on SDS-polyacrylamide gels. The separated proteins were
then transferred onto PVDF membranes (Millipore, Bedford, MA, USA).
The membranes were first probed with a primary antibody. After
blocking with 5% skim milk for 2 h, the membranes incubated with
primary antibodies [rabbit anti-KLF4 (1:500, Santa Cruz
Biotechnology, Santa Cruz, CA, USA). The secondary antibody was
mouse anti-GAPDH (1:3000, Santa Cruz Biotechnology).
RNA extraction and reverse
transcription-quantitative PCR (RT-qPCR)
Total RNA was isolated from the tissue samples and
cultured cells using TRIzol reagent (Invitrogen, Carlsbad, CA, USA)
according to the manufacturer's insstructions. The concentration of
extracted RNA was measured using a NanoDrop ND-1000
Spectrophotometer (Agilent, Santa Clara, CA, USA). RNA was reverse
transcribed into complementary DNA (cDNA) using the PrimeScript RT
reagent kit with gDNA Eraser (Takara, Dalian, China). Quantitative
PCR (qPCR) was performed using the SYBR PrimeScript RT-PCR kit
(Takara, Shiga, Japan) and the ABI 7500 System (Applied Biosystems,
Foster City, CA, USA) according to manufacturer's instructions. The
relative expression was calculated via the comparative cycle
threshold (CT) method and was normalized to the expression of U6
small RNA. The primers used are listed in Table II. The differential expression
level was calculated using the 2−ΔΔCt formula. All the
experiments were conducted at least 3 times.
| Table IIPrimer sequences used in this
study. |
Table II
Primer sequences used in this
study.
| Primer name | Primer sequences
(5′→3′) |
|---|
| U6-OF |
CTCGCTTCGGCAGCACA |
| U6-OR |
AACGCTTCACGAATTTGCGT |
| Klf4-OF |
ACCCTGGGTCTTGAGGAAGT |
| Klf4-OR |
CATGAGCTCTTGGTAATGGAGC |
| Klf4-siKlf4 |
AUCGUUGAACUCCUCGGUCUCUCUC |
| Pri-miR-31-OF |
GAACTACCCACAAACCTCCTG |
| Pri-miR-31-OR |
GGTGTGTCCAAGGAATAGCCA |
| miR-31-OF |
TAATACTGCCTGGTAATGATGA |
| miRNA-OR |
GTCGTATCCAGTGCAGGGTCCGAGGTATTCGCACTGGATACGACAGCTAT |
|
miR-31-promoter(−1069To+1)-F(MluI) |
CCGGACGCGTGCACAAAAGTTATACATAATGTCATTATTCTTATG |
|
miR-31-promoter(−1069To+1)-R(HindIII) |
GCCCAAGCTTCAGTTCCAAGTTACAGGAGAATACTATGA |
|
Klf4-miR-31-mut-F1 |
AATTAATGAGTGTGTTTACGCACGCACAGGTGAAAGGAAAAA |
|
Klf4-miR-31-mut-R1 |
TTTTTCCTTTCACCTGTGCGTGCGTAAACACACTCATTAATT |
|
Klf4-miR-31-mut-F2 |
AGTTGCTTGAGAGGCTACGCTCGCACGATTCACCTGAGCCTG |
|
Klf4-miR-31-mut-R2 |
CAGGCTCAGGTGAATCGTGCGAGCGTAGCCTCTCAAGCAACT |
|
Klf4-miR-31-mut-F3 |
GAGCATTATTTTTGGCACGCCTTCGCGGCTCATGCCTGTAAT |
|
Klf4-miR-31-mut-R3 |
ATTACAGGCATGAGCCGCGAAGGCGTGCCAAAAATAATGCTC |
| Negative control
sense |
UUCUCCGAACGUGUCACGUTT |
| Negative control
antisense |
ACGUGACACGUUCGGAGAATT |
| Hsa-miR-31 mimics
sense |
AGGCAAGAUGCUGGCAUAGCU |
| Hsa-miR-31 mimics
antisense |
CUAUGCCAGCAUCUUGCCUUU |
| MircoRNA inhibitor
NC |
CAGUACUUUUGUGUAGUACAA |
| Hsa-miR-31
inhibitors |
AGCUAUGCCAGCAUCUUGCCU |
Plasmids, small interfering RNAs, miRNAs
and transfection
The miR-31 mimic, inhibitor, Klf4 siRNA and negative
controls (mimic NC or inhibitor NC) were synthesized by GenePharma,
Shanghaim, China. The previously described plasmid Klf4 and the
empty control vector plasmid were obtained from Xie Keping Research
Group Shanghai East Hospital affiliated to Tongji University School
of Medicine. The transfection of the plasmids and siRNAs,
respectively, into the HCC cells was performed using Lipofectamine
2000 (Invitrogen) transfection reagent. The relative levels of Klf4
and miR-31 in the transfected cells were examined by RT-qPCR. The
cells were transfected with the plasmids or siRNAs at various
concentrations as indicated 48 h before the performance of
functional assays.
NanoString
A total of 2×106 Hep3B cells grown in the
exponential phase were seeded in 6-well plates and cultured for 2
days. The cells were then harvested for total RNA using the
miRNeasy kit (Qiagen). A total of 100 ng of total RNA was assayed
using the Human nCounter miRNA Assay 2.0 kit following the
manufacturer's instructions (NanoString Technologies, Seattle, WA,
USA). Differences in miRNA expression were analyzed using the
NanoSTRIDE software program with default settings. Clustering of
the differentially expressed genes and heatmap generation was
performed using the GenePattern Server (genepattern.broadinstitute.org). The volcano plot
displaying the significance of the miRNA differences was produced
using R version 3.0.2.
Luciferase reporter assays
For the binding of Klf4 to the miR-31 promoter, the
1.5 kb region directly upstream of miR-31 transcription binding
site was amplified by PCR and inserted into the pGL3 vector
(Promega, San Luis Obispo, CA, USA). The Hep3B and Sk-hep-1 cells
were co-transfected with either pcDNA3.1 (Promega) or the
pcDNA3.1-Klf4 and miR-31 promoter. At 48 h after transfection,
luciferase activity was analyzed according to the Dual-Luciferase
reporter assay system (Promega), using a GloMax fluorescence reader
(Promega).
Cell proliferation assay
The Sk-hep-1 and Bel-7402 cells were transfected
with the indicated miR-31 mimic or inhibitor (50 nM), the Klf4
overexpression plasmid or and Klf4 siRNA (siKlf4; 120 ng), plated
in 96-well plates at 3,000 cells/well and maintained in culture
medium. They were assessed at days 1–6 of culture with 0.5 mg/ml
cell counting kit 8 (CCK8; Dojindo Laboratories, Kumamoto, Japan)
at 37°C for 3 h and then shaken for 20 min. The optical density was
determined at 450 nm using an enzyme-linked immunosorbent assay
reader (Dasit, Milan, Italy). Each experiment was conducted for at
least 3 times and the average of the results was analyzed.
Cell migration assay
The transfected HCC cells (5×104) in
serum-free medium were placed into the upper part of a Transwell
chamber in a 24-well format with 8 mm diameters (Corning, Corning,
NY, USA). In the bottom chamber, 800 µl of normal MEM medium
containing 10% FBS were added as a chemoattractant and the chambers
were incubated for 24–48 h at 37°C and 5% CO2. The cells
on the upper part of the Transwell chamber were removed using a
cotton swap, and the cells that had migrated through the membrane
were stained with 0.05% crystal violet for 2 h. Finally, the
migrated cells were counted in 5 random fields under a microscope
(Olympus, Shanghai, China) and the average number of 5 fields was
calculated. All assays were performed in triplicate and repeated 3
times.
Wound healing assay
For the wound healing assay, 3.5×105 of
examined HCC cells were seeded into 6-well plates to achieve 90%
confluence. Wounds were produced in confluent monolayer cells using
a plastic tip, the cell debris were removed using
phosphate-buffered saline (PBS), and 0.5% FBS-containing MEM was
added. The scratched cells were then incubated at 37°C with 5%
CO2. The initial scratched gap breadth (0 h) and the
residual scratched gap breadth (48 h) were measured using a light
microscope (Nikon, Tokyo, Japan).
Statistical analyses
Quantitative data are presented as the means ± SEM.
Parameters of two-tailed, 95% CI were used for statistical
analysis. Only P-values <0.05 were considered to indicate
statistically significant differences.
Results
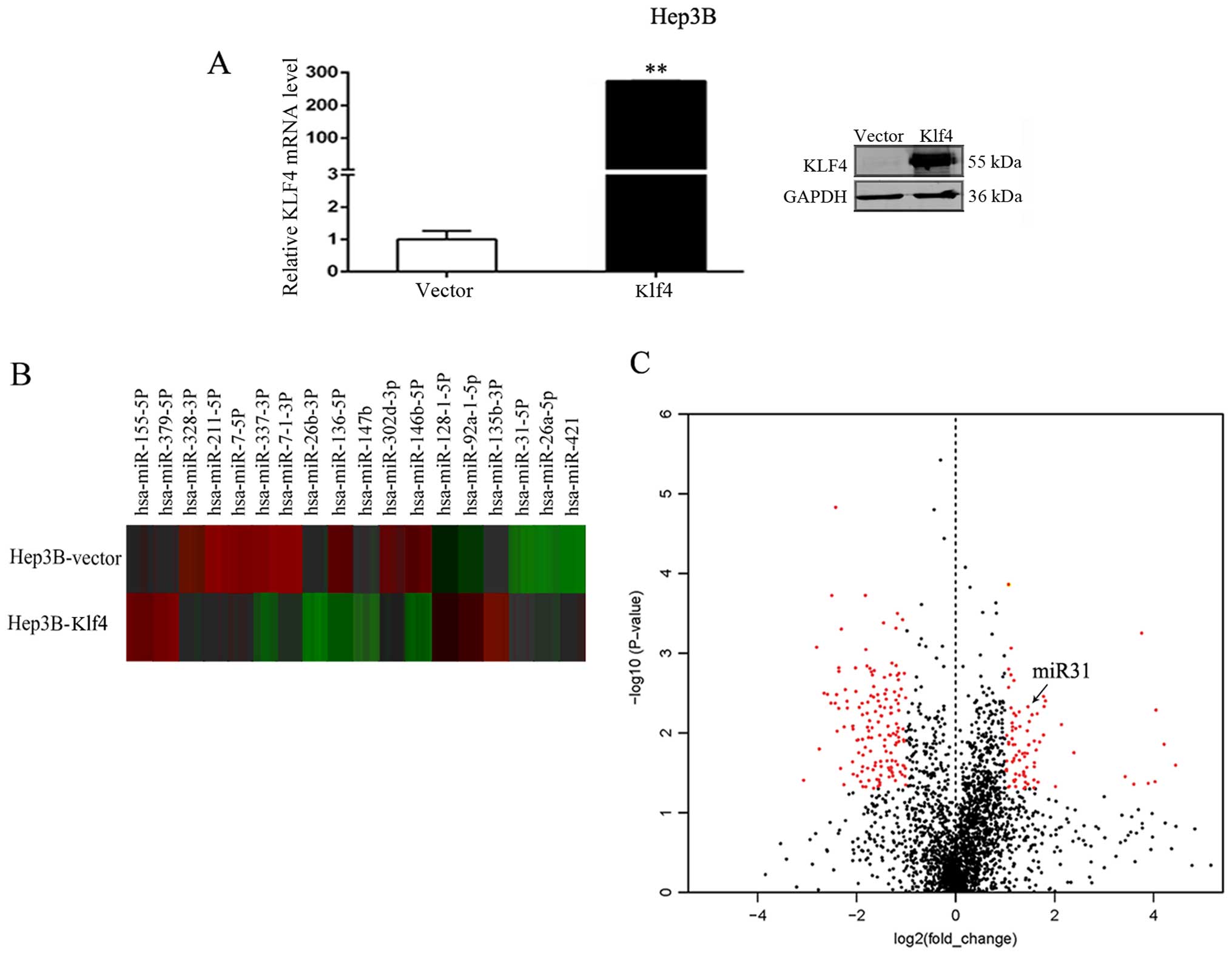
miRNAs are differentially expressed in
the Hep3B cells transfected with the empty vector or the Klf4
overexpression plasmid
To identify candidate miRNAs that may play a role in
HCC, RT-qPCR and western blot analysis of Klf4 expression in the
Hep3B cells transfected with the Klf4 plasmid were initially
performed (Fig. 1A). We found
that the mRNA and protein expression of Klf4 was significantly
increased following the overexpression of Klf4. We then compared
miRNA expression levels between the empty vector- and Klf4
overexpression vector-transfected Hep3B cells using the nCounter
NanoString platform. Using RNA derived from the Hep3B cells
transfected with the empty vector or Klf4 overexpression vector
under standard culture conditions, we found that a number of miRNAs
were differentially expressed. Of the 2,565 miRNAs assayed by this
method, a significant difference in expression was observed in 436
miRNAs (17.0%; P<0.05, 1.5-fold), including 216 that were
downregulated and 220 that were upregulated in the Hep3B cells
transfected with the empty vector or Klf4 overexpression vector.
The heatmap identified the 18 most differentially regulated miRNAs
from the miRNA array of Hep3B cells transfected with the empty
vector or Klf4 overexpression vector, including 8 upregulated and
10 downregulated miRNAs (Fig.
1B). We found that the miR-31 cluster was significantly
upregulated in the Hep3B cells (fold change, 1.92; P-value, 0.009).
At last, Volcano plot revealed the profile of the differentially
expressed miRNAs in the Hep3B cells. This plot outlines the fold
change (x-axis) and significance level of expression as the
log10 P-value (y-axis). The red dots on the left side
represent the downregulated miRNAs in the Hep3B cells transfected
with the Klf4 overexpression vector compared to the cells
transfected with the empty vector; the red dots on the right side
represent the upregulated miRNAs in the Hep3B cells transfected
with the Klf4 overexpression plasmid compared with the cells
transfected with the empty vector. The black dots indicate miRNAs
which did not exhibit a significant change in expression.
Significance was determined with a P-value cut off of 0.05 and a
1.5-fold change (Fig. 1C). We
found that miR-31 (the red dots on the right) was significantly
upregulated in the Hep3B cells.
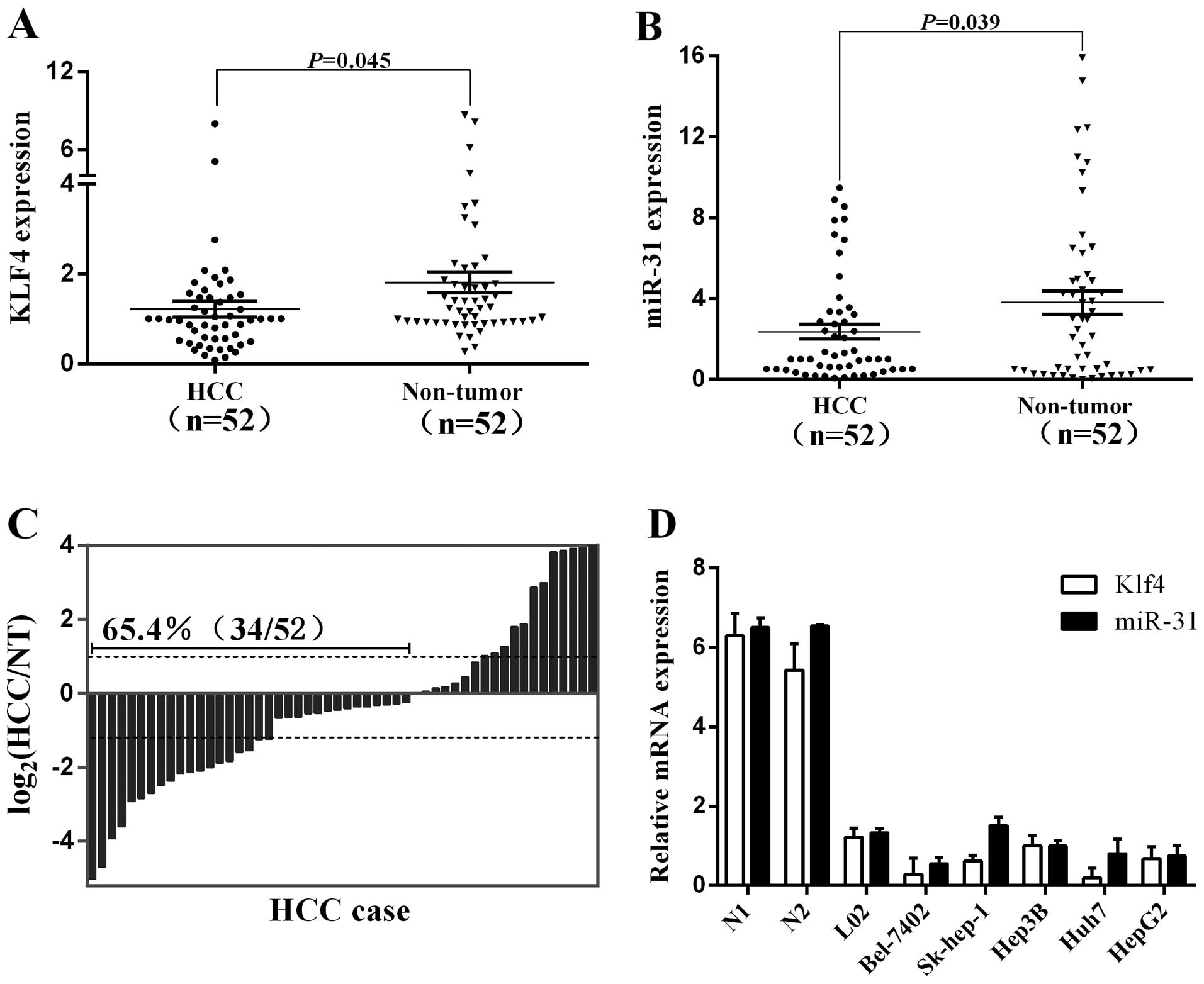
Klf4 and miR-31 are downregulated in HCC
tissues
In this study, we explored the relative expression
levels of Klf4 and miR-31 in 52 pairs of HCC tissues and
neighboring noncancerous liver tissues by RT-qPCR. The expression
of Klf4 and miR-31 was significantly downregulated in the cancerous
compared to the non-cancerous tissues (Fig. 2A and B). The downregulation of
Klf4 was found to inversely correlate with the clinical stage, T
classification and hepatitis B infection, whereas the
downregulation of miR-31 was directly associated with the clinical
stage in patients with HCC (Table
I). Of note, the decreased expression of miR-31 in HCC was
observed in 65.4% (34/52) of the cases (Fig. 2C). Additionally, the endogenous
expression of Klf4 and miR-31 was analyzed by qRT-PCR in six chosen
human liver cell lines, including an immortalized normal hepatic
cell line (Fig. 2D). The liver
cancer cell lines (Bel-7402, Sk-hep-1, Hep3B, Huh7 and HepG2)
exhibited relatively low Klf4 and miR-31 expression levels compared
to the non-cancer cell line, L02. These findings again suggest that
the expression of Klf4 and miR-31 is suppressed in HCC and that a
low expression of Klf4 and miR-31 is associated with the biological
process of tumorigenesis in HCC.
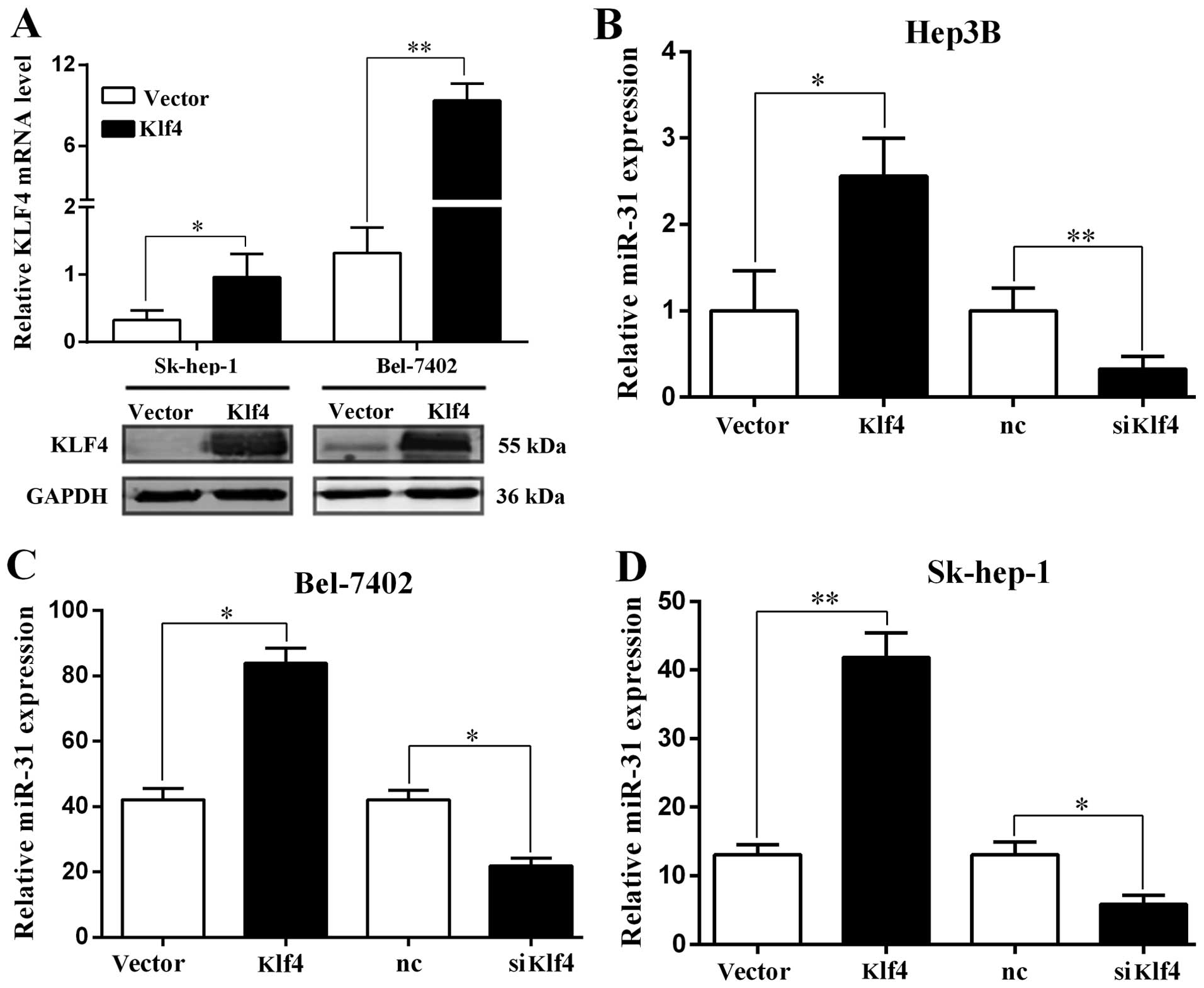
miR-31 is directly regulated by the
transcription factor Klf4
The expression of Klf4 in HCC was examined by
RT-qPCR and western blot analysis. The Klf4 mRNA and protein levels
were significantly increased when the Sk-hep-1 and Bel-7402 cells
were transfected with the Klf4 overexpression plasmid (Fig. 3A). We then examined the expression
of miR-31 by RT-qPCR, and found that it was significantly increased
and decreased when the Hep3B, Bel-7402 and Sk-hep-1 cells were
transfected with the Klf4 overexpression plasmid and or siKlf4,
respectively (Fig. 3B–D).
Moreover we examined the pri-miR-31 levels by RT-qPCR, and the
results revealed that the levels were significantly increased and
decreased when the Hep3B, Bel-7402 and Sk-hep-1 cells were
transfected with the Klf4 overexpression plasmid and siKlf4,
respectively (Fig. 3E–G).
To explore this regulation of miR-31 expression, we
analyzed the 1.5 kb region of miR-31 upstream and found 3 binding
sites of Klf4 (mut1-3). We then examined the effects of the
transcription factor Klf4 on miR-31 promoter-driven luciferase
activity in Hep3B cells (Fig.
3H). The mutation of the Klf4 binding site 3 (mut3) abolished
the effects of Klf4 on the relative luciferase activity in the
Hep3B and Sk-hep-1 cells, while mutations of other binding sites
had no significant effect (Fig. 3I
and J). Thus, Klf4 regulates the expression of miR-31 by
binding to site 3, indicating that Klf4 is involved in the
transcriptional regulation of miR-31 by directly binding to the
miR-31 promoter. A positive correlation of Klf4 and miR-31 was also
revealed in the HCC tissues (Fig.
3K). Taken together, these findings suggest that the
transcription factor Klf4 is a functional regulator of miR-31 in
HCC.
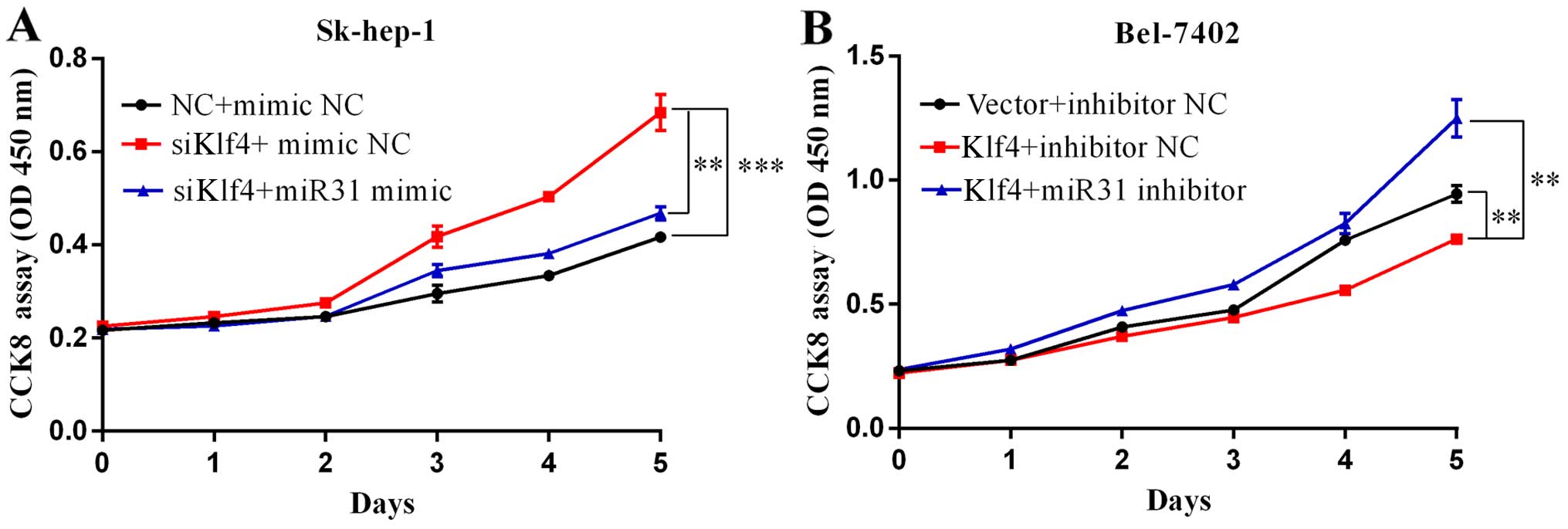
Klf4 and miR-31 inhibit HCC cell
proliferation and migration in vitro
To ascertain the biological functions of Klf4 and
miR-31 in HCC, following the ectopic expression of Klf4 and miR-31,
we examined the growth rate of two different liver cancer cell
lines, Sk-hep-1 and Bel-7402, by CCK8 assay. Transfection with
siKlf4 promoted Sk-hep-1 cell proliferation, and transfection with
miR-31 mimic attenuated the growth-promoting effects induced by
transfection with siKlf4 (Fig.
4A). Klf4 overexpression suppressed Bel-7402 cell
proliferation, while transfection with miR-31 inhibitor attenuated
the growth inhibitory effects of Klf4 (Fig. 4B). The Sk-hep-1 and Bel-7402 cells
were seeded onto 96-well plates and respectively transfected with
Klf4 or siKlf4, miR-31 mimic or inhibitor for 1–6 days. Cell
viabilities were determined by CCK8 assays. Cell growth was
measured every 24 h. Mimic NC and inhibitor NC represents negative
control miRNA.
As shown in Fig.
4C, transfection with miR-31 mimic decreased the number of
Sk-hep-1 migrating cells, which had been increased by transfection
with siKlf4. Similarly, transfection with miR-31 inhibitor
increased the number of Bel-7402 migrating cells, which had been
decreased by transfection with Klf4 overexpression vector.
Consistent with the Transwell assay results, following transfection
with the miR-31 mimic, the Sk-hep-1 cells migrated more slowly than
the controls and the siKlf4-transfected cells, as shown by wound
healing assays (Fig. 4D). By
contrast, the Bel-7402 cells transfected with miR-31 inhibitor
exhibited a more rapid migration rate compared with the controls
and the cells transfected with the Klf4 overexpression plasmid, as
shown by wound healing assays (Fig.
4D). These results demonstrate that miR-31 contributes to the
regulation of liver cancer cell motility and progression.
Discussion
Over the past decade, the aberrant expression of
miRNAs has been increasingly reported in human cancers (5). Numerous studies have shed light on
tumor-targeting therapies using miRNAs as a novel diagnostic and
therapeutic tools (7,8,12,25,28). In the present study, the
downregulation of Klf4 and miR-31 was consistently observed in HCC
tissues and they may thus serve as potential diagnostic markers and
therapeutic targets for HCC. Furthermore, the ectopic expression of
miR-31 suppressed HCC cell proliferation and migration which was
increased by transfection with siKlf4 in vitro. Moreover,
the transcription factor Klf4 regulated the expression of miR-31,
which in turn affected the progression of HCC. Our data suggest an
important role of miR-31 in the progression of HCC.
Klf4 is a zinc finger transcription factor. Previous
studies have indicated that Klf4 expression is reduced in different
types of cancer (29,30). Accumulating clinical evidence also
suggests that Klf4 functions as a tumor suppressor gene (31,32). Klf4 knockdown has been shown to
promote cell growth, migration and adhesion, while Klf4
overexpression inhibits the proliferation of HCC cells (27). This study revealed that Klf4 was
decreased in liver cancer tissues and cells compared with normal
liver tissues. Therefore, the low expression of Klf4 in liver
cancer may contribute to the reduced expression of miR-31 and may
also play a crucial role in the progression of HCC.
miRNAs are post-transcription regulators of many
genes, and their deregulation is related to cancer initiation,
development and progression (33). Proliferation and metastasis, two
hallmarks of malignancy, are the leading causes for cancer-related
death (34). It has been
demonstrated that miRNAs are associated with these two events. For
example, it has been shown that miR-17-5p promotes HCC cell
migration and proliferation by targeting the p38-HSP27 pathway
(35), and miR-26a has been shown
to inhibit HCC growth and metastasis by suppressing IL-6 (10). In the present study, we
demonstrated that miR-31 expression was decreased in liver cancer
tissues and cells. However, the underlying mechanisms involved in
the deregulation of miR-31 are still unknown. Therefore, we
investigated the correlation between the nuclear transcription
factor Klf4 and miR-31 in human HCC and its effect on liver cancer
cell proliferation and metastasis.
Collectively, our results provide experimental
evidence that the transcription factor Klf4 suppresses cell growth
and metastasis by directly targeting miR-31 in HCC. The newly
identified Klf4/miR-31 signalling axis may provide new insight into
the pathogenesis of HCC and represents a potential therapeutic
target for the treatment of HCC.
Acknowledgments
The present study was supported in part by grants
from the National Natural Science Foundation of China (nos.
81272714, 81572310 and 81472576) and the KeyDisciplines Group
Construction Project of Pudong Health Bureau of Shanghai (no.
PWZxq2014-04).
References
|
1
|
Maluccio M and Covey A: Recent progress in
understanding, diagnosing, and treating hepatocellular carcinoma.
CA Cancer J Clin. 62:394–399. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Bhayani NH, Jiang Y, Hamed O, Kimchi ET,
Staveley-O'Carroll KF and Gusani NJ: Advances in the pharmacologic
treatment of hepatocellular carcinoma. Curr Clin Pharmacol.
10:299–304. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Moeini A, Cornellà H and Villanueva A:
Emerging signaling pathways in hepatocellular carcinoma. Liver
Cancer. 1:83–93. 2012. View Article : Google Scholar
|
|
4
|
Lujambio A and Lowe SW: The microcosmos of
cancer. Nature. 482:347–355. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Di Leva G, Garofalo M and Croce CM:
MicroRNAs in cancer. Annu Rev Pathol. 9:287–314. 2014. View Article : Google Scholar :
|
|
6
|
Fang W and Bartel DP: The menu of features
that define primary microRNAs and enable de novo design of dicroRNA
genes. Mol Cell. 60:131–145. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Parpart S, Roessler S, Dong F, Rao V,
Takai A, Ji J, Qin LX, Ye QH, Jia HL, Tang ZY and Wang XW:
Modulation of miR-29 expression by α-fetoprotein is linked to the
hepatocellular carcinoma epigenome. Hepatology. 60:872–883. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Najafi Z, Sharifi M and Javadi G:
Degradation of miR-21 induces apoptosis and inhibits cell
proliferation in human hepatocellular carcinoma. Cancer Gene Ther.
22:530–535. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
He XX, Guo AY, Xu CR, Chang Y, Xiang GY,
Gong J, Dan ZL, Tian DA, Liao JZ and Lin JS: Bioinformatics
analysis identifies miR-221 as a core regulator in hepatocellular
carcinoma and its silencing suppresses tumor properties. Oncol Rep.
32:1200–1210. 2014.PubMed/NCBI
|
|
10
|
Yang X, Liang L, Zhang XF, Jia HL, Qin Y,
Zhu XC, Gao XM, Qiao P, Zheng Y, Sheng YY, et al: MicroRNA-26a
suppresses tumor growth and metastasis of human hepatocellular
carcinoma by targeting interleukin-6-Stat3 pathway. Hepatology.
58:158–170. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Baek S, Cho KJ, Ju HL, Moon H, Choi SH,
Chung SI, Park JY, Choi KH, Kim Y, Ahn SH, et al: Analysis of miRNA
expression patterns in human and mouse hepatocellular carcinoma
cells. Hepatol Res. 45:1331–1340. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Mlcochova J, Faltejskova-Vychytilova P,
Ferracin M, Zagatti B, Radova L, Svoboda M, Nemecek R, John S, Kiss
I, Vyzula R, et al: MicroRNA expression profiling identifies
miR-31-5p/3p as associated with time to progression in wild-type
RAS metastatic colorectal cancer treated with cetuximab.
Oncotarget. 6:38695–38704. 2015.PubMed/NCBI
|
|
13
|
Tseng SH, Yang CC, Yu EH, Chang C, Lee YS,
Liu CJ, Chang KW and Lin SC: K14-EGFP-miR-31 transgenic mice have
high susceptibility to chemical-induced squamous cell tumorigenesis
that is associating with Ku80 repression. Int J Cancer.
136:1263–1275. 2015. View Article : Google Scholar
|
|
14
|
Lu WC, Liu CJ, Tu HF, Chung YT, Yang CC,
Kao SY, Chang KW and Lin SC: miR-31 targets ARID1A and enhances the
oncogenicity and stemness of head and neck squamous cell carcinoma.
Oncotarget. Aug 9–2016.Epub ahead of print.
|
|
15
|
Gao W, Liu L, Xu J, Shao Q, Liu Y, Zeng H
and Shu Y: A systematic analysis of predicted MiR-31-targets
identifies a diagnostic and prognostic signature for lung cancer.
Biomed Pharmacother. 68:419–427. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Taccioli C, Garofalo M, Chen H, Jiang Y,
Tagliazucchi GM, Di Leva G, Alder H, Fadda P, Middleton J, Smalley
KJ, et al: Repression of esophageal neoplasia and inflammatory
signaling by anti-miR-31 delivery in vivo. J Natl Cancer Inst.
107:djv2202015. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Wang S, Li Q, Wang K, Dai Y, Yang J, Xue
S, Han F, Zhang Q, Liu J and Wu W: Decreased expression of
microRNA-31 associates with aggressive tumor progression and poor
prognosis in patients with bladder cancer. Clin Transl Oncol.
15:849–854. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Schaefer A, Jung M, Mollenkopf HJ, Wagner
I, Stephan C, Jentzmik F, Miller K, Lein M, Kristiansen G and Jung
K: Diagnostic and prognostic implications of microRNA profiling in
prostate carcinoma. Int J Cancer. 126:1166–1176. 2010.
|
|
19
|
Ruoming W, Zhen Y, Tengteng Z and Jisheng
H: Tumor suppressor microRNA-31 inhibits gastric carcinogenesis by
targeting Smad4 and SGPP2. Cancer Gene Ther. 22:564–572. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Kim HS, Lee KS, Bae HJ, Eun JW, Shen Q,
Park SJ, Shin WC, Yang HD, Park M, Park WS, et al: MicroRNA-31
functions as a tumor suppressor by regulating cell cycle and
epithelial-mesenchymal transition regulatory proteins in liver
cancer. Oncotarget. 6:8089–8102. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Viré E, Curtis C, Davalos V, Git A, Robson
S, Villanueva A, Vidal A, Barbieri I, Aparicio S, Esteller M, et
al: The breast cancer oncogene EMSY represses transcription of
antimetastatic microRNA miR-31. Mol Cell. 53:806–818. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Creighton CJ, Fountain MD, Yu Z, Nagaraja
AK, Zhu H, Khan M, Olokpa E, Zariff A, Gunaratne PH, Matzuk MM and
Anderson ML: Molecular profiling uncovers a p53-associated role for
microRNA-31 in inhibiting the proliferation of serous ovarian
carcinomas and other cancers. Cancer Res. 70:1906–1915. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Zhang N, Zhang J, Shuai L, Zha L, He M,
Huang Z and Wang Z: Krüppel-like factor 4 negatively regulates
β-catenin expression and inhibits the proliferation, invasion and
metastasis of gastric cancer. Int J Oncol. 40:2038–2048.
2012.PubMed/NCBI
|
|
24
|
Li Q, Gao Y, Jia Z, Mishra L, Guo K, Li Z,
Le X, Wei D, Huang S and Xie K: Dysregulated Krüppel-like factor 4
and vitamin D receptor signaling contribute to progression of
hepatocellular carcinoma. Gastroenterology. 143:799–810.e1. 22012.
View Article : Google Scholar
|
|
25
|
Yu T, Chen X, Lin T, Liu J, Li M, Zhang W,
Xu X, Zhao W, Liu M, Napier DL, et al: KLF4 deletion alters gastric
cell lineage and induces MUC2 expression. Cell Death Dis.
7:e22552016. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Lv H, Zhang Z, Wang Y, Li C, Gong W and
Wang X: MicroRNA-92a promotes colorectal cancer cell growth and
migration by inhibiting KLF4. Oncol Res. 23:283–290. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Liu S, Yang H, Chen Y, He B and Chen Q:
Krüppel-like factor 4 enhances sensitivity of cisplatin to lung
cancer cells and inhibits regulating epithelial-to-mMesenchymal
transition. Oncol Res. 24:81–87. 2016. View Article : Google Scholar
|
|
28
|
Gandellini P, Giovannetti E and Nicassio
F: MicroRNAs in cancer management: Big challenges for small
molecules. Biomed Res Int. 2015:9821562015. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Zhou J, Lai PB and Tsui SK: Identification
of a non-coding KLF4 transcript generated from intron retention and
downregulated in human hepatocellular carcinoma. Int J Oncol.
47:1554–1562. 2015.PubMed/NCBI
|
|
30
|
Tien YT, Chang MH, Chu PY, Lin CS, Liu CH
and Liao AT: Downregulation of the KLF4 transcription factor
inhibits the proliferation and migration of canine mammary tumor
cells. Vet J. 205:244–253. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Mao Q, Quan T, Luo B, Guo X, Liu L and
Zheng Q: MiR-375 targets Klf4 and impacts the proliferation of
colorectal carcinoma. Tumour Biol. 37:463–471. 2016. View Article : Google Scholar
|
|
32
|
Lin ZS, Chu HC, Yen YC, Lewis BC and Chen
YW: Krüppel-like factor 4, a tumor suppressor in hepatocellular
carcinoma cells reverts epithelial mesenchymal transition by
suppressing slug expression. PLoS One. 7. pp. e435932012,
View Article : Google Scholar
|
|
33
|
Fiorenza A, Lopez-Atalaya JP, Rovira V,
Scandaglia M, Geijo-Barrientos E and Barco A: Blocking miRNA
biogenesis in adult forebrain neurons enhances seizure
susceptibility, fear memory, and food intake by increasing neuronal
responsiveness. Cereb Cortex. 26:1619–1633. 2016. View Article : Google Scholar
|
|
34
|
Meehan K and Vella LJ: The contribution of
tumour-derived exosomes to the hallmarks of cancer. Crit Rev Clin
Lab Sci. 53:121–131. 2016. View Article : Google Scholar
|
|
35
|
Yang F, Yin Y, Wang F, Wang Y, Zhang L,
Tang Y and Sun S: miR-17-5p Promotes migration of human
hepatocellular carcinoma cells through the p38 mitogen-activated
protein kinase-heat shock protein 27 pathway. Hepatology.
51:1614–1623. 2010. View Article : Google Scholar : PubMed/NCBI
|