Introduction
Chronic inflammatory bone disease has the major
characteristics of osteopenia, mainly including post-menopausal
osteoporosis, rheumatoid arthritis and periodontal disease. The
disease is widely observed on a global scale, and is the primary
cause of fractures and bone loss, and seriously threatens human
health (1). The suppression of
bone-forming ability is the main pathogenesis of the disease.
Methods with which to enhance bone-forming ability are of major
importance in providing a cure for the disease. Bone marrow-derived
mesenchymal stem cells (BMSCs) are adult stem cells with
self-renewal and multi-directional differentiation ability. Under
different induction environments, they can differentiate into
osteoblasts, adipocytes, chondroblasts and neuroblasts (2). Studies have indicated that BMSCs are
the source of osteoblasts and osteocytes (3,4).
Inducing BMSCs to differentiate directionally is of significance to
curing inflammatory bone disease and repairing bone loss (3,4).
However, the specific mechanisms involved remain unclear, as
osteogenic differentiation involves a complex network, including
the regulation of multiple signaling pathways at the
transcriptional and post-transcriptional level (3).
MicroRNAs (miRNAs or miRs) are non-coding RNAs
approximately 18–25 nucleotides in length and have been discovered
in recent years. By targeting the 3′UTR of target genes, they can
restrain the translation and stability of genes (5). It has been demonstrated that miRNAs
plays an important role in regulating the self-renewal and
differentiation ability of stem cells (6). Studies have found that miRNAs exert
a regulatory effect on osteogenic differentiation in different
cells; however, the osteogenic differentiation of stem cells is an
extremely complex process and may involve the mutual coordination
of multiple factors and different signaling pathways on different
levels (6,7). As a result, studying the regulatory
role of miRNAs in the osteogenic differentiation of BMSCs may be
crucial to providing a cure for osteoporosis and bone loss.
As adult stem cells derived from mesoblast, BMSCs
can not only differentiate into adult cells from mesoblasts,
including osteocytes, chondrocytes and adipocytes under the
function of an appropriate inducer for culture in vitro, but
can also can transdifferentiate into nerve cells from the neuroderm
(8). Therefore, in the process of
transdifferentiation, it is necessary to transmit the signal with
induced differentiation into cells through the signal transduction
pathway, so as to induce cell differentiation and impact outcomes.
Mitogen-activated protein kinase (MAPK) is a type of
serine/threonine protein kinase in cells. When extracellular growth
factors are combined with corresponding receptors of the
cytomembrane, this activates the MAPK cascade reaction, so as to
impact the expression of corresponding cellular transcription
factors and impact cell proliferation, differentiation and outcomes
(9,10).
With the development of cytobiology, it has been
realized that cell adhesion is not a pure mechanical connection.
Instead, it also involves complex signal transduction pathways
(8). When studying the
categories, structure and functions of the cell surface for
adhesion molecules, the investigation of the signal transduction
process mediated by these adhesion molecules has already become the
hotspot in the field of life science. In a previous study, it was
demostrated that genistein promotes osteoblastic differentiation
through the p38 mitogen activated protein kinase
(MAPK)-core-binding factor 1 (Cbfa1) pathway. Downstream JUN/p38 is
activated (9). Thus, JUN/p38 may
participate in the process of cell differentiation and
proliferation. However, the mechanisms and function of miR-214 in
the osteogenic differentiation of BMSCs remain unclear. Thus, the
purpose of the present study was to investigate the function and
possible molecular mechanisms of miR-214 in the osteogenic
differentiation of BMSCs.
Materials and methods
Animals, isolation of BMSCs and cell
culture
Male Sprague-Dawley rats (n=6; weighing, 180–210 g)
were purchased from Beijing Vital River (Beijing, China). This
study was approved by the Ethics Committee of Beijing Army General
Hospital, Beijing, China. The rats were then sacrificed by
decollation under 5% chloral hydrate and sterilized using 75% ethyl
alcohol for 3–5 min. The femur of each rat was separated and excess
tissue was removed using surgical scissors. The bone marrow was
flushed out using Dulbeccos modified Eagle's medium (DMEM; HyClone,
Waltham, MA, USA) and the bone marrow cells were washed and
filtered using a 200 mesh sieve. Bone marrow cell were collected
following centrifugation at 500 × g for 10 min. The cells were
washed with PBS and cultivated with red blood cell lysis buffer.
The cell precipitate (1–2×106) was cultivated with DMEM
supplemented with 10% fetal bovine serum (FBS), 100 U/ml penicillin
and 100 lg/ml streptomycin) (all from HyClone) and seeded in a 6
well-plate at 37°C in 5% CO2. The culture medium was
replaced every 3 days. Primary rat BMSCs were digested with
10%trypsin after large colonies had formed and became
confluent.
Transfection of cells with miR-214 and
anti-miR-214 plasmids
miR-214 and the negative control plasmids, and the
anti-miR-214 plasmid were packaged using lentiviral vectors
generated by GeneChem (Shanghai, China). Lentivirus
(1×107 TU/ml; Invitrogen, Carlsbad, CA, USA) was used to
infect the cells according to the manufacturer's instructions.
Untransfected cells were used as controls. The cells were then
cultured in fresh medium for further experiments after the
lentivirus for 8 h.
Osteogenic differentiation of BMSCs,
treatment with p38 and c-Jun N-terminal kinase (JNK) inhibitors and
Oil Red O staining
Osteogenic differentiation was induced using
standard osteoblast induction medium (10−8 M
dexamethasone, 0.2 mM l-ascorbic acid, 10 mM, β-glycerophosphate
and 10 mM 1.25-vitamin D3), as previously described (11). The JNK inhibitor, SP600125 (final
concentration 20 µM; MedChemexpress, Princeton, NJ, USA),
and the p38 inhibitor, SB202190 (final concentration 20 µM;
MedChemexpress), were added to the BMSCs for 7 days. The BMSCs were
fixed in 10% formalin for 30 min at room temperature and stained
with Oil Red O (Beyotime, Jiangsu, China) to assess adipocyte
differentiation. Subsequently, 85% propylene glycol was added to
the cells for 5 min and the cells were observed under a microscope
(LSM510 Meta Confocal Microscope; Carl Zeiss, Oberkochen,
Germany).
Alkaline phosphatase (ALP) activity
assay
The BMSCs transfected with the miR-214 or
anti-miR-214 plasmids, or treated with JNK inhibitor or p38
inhibitor were seeded at a density of 2×105 cells/well
in 24-well plates and subjected to osteogenic differentiation. The
BMSCs were lysed with RIPA lysis buffer (Thermo Fisher Scientific,
Wilmington, DE, USA) supplemented with protease inhibitor. The
protein concentration was determined by Bradford assay (Bio-Rad,
Hercules, CA, USA) on a microplate spectrophotometer (Tecan, Port
Melbourne, Australia). ALP activity was determined using an ALP
activity kit (Nanjing Jiancheng Biotech, China) on a microplate
spectrophotometer (Tecan).
Reverse transcription-quantitative PCR
(RT-qPCR)
Total RNA was extracted from the BMSCs transfected
with miR-214 or anti-miR-214 plasmids, or treated with JNK
inhibitor or p38 inhibitor using TRIzol reagent (Takara, Dalian,
China). A total of 2 µl of total RNA was reverse transcribed
into cDNA using a HiFi-MMLV cDNA kit (Tiangen, Beijing, China). The
Quant SYBR-Green PCR kit (Bio-Rad, Munich, Germany) was used for
qPCR. The temperature profile of the reaction was 95°C for 2 min,
followed by 40 cycles of denaturation at 95°C for 20 sec, annealing
at 60°C for 30 sec and an extension at 72°C for 45 sec. The
relative quantitation was calculated using the 2−ΔΔCt
method. The sequences of the primers are listed in Table I.
 | Table IPrimer sequences use for PCR. |
Table I
Primer sequences use for PCR.
| Gene | Forward primer | Reverse primer |
|---|
| miR-214 |
5′-AGCCGACAGCAGGCACAGACA-3′ |
5′-AGCCGACAGCAGGCACAGACA-3′ |
| U6 |
5′-CGCTTCACGAATTTGCGTGTCAT-3 |
5′-GCTTCGGCAGCACATATACTAAAAT-3 |
| Col I |
5′-CTGCCCAGAAGAATATGTATCACC-3′ |
5′-GAAGCAAAGTTTCCTCCAAGACC-3′ |
| OCN |
5′-GAGGGCAATAAGGTAGTGAA-3′ |
5′-CATAGATGCGTTTGTAGGC-3′ |
| OPN |
5′-CCAAGCGTGGAAACACACAGCC-3′ |
5′-GGCTTTGGAACTCGCCTGACTG-3′ |
| Actin |
5′-GTAAAGACCTCTATGCCAACA-3′ |
5′-GGACTCATCGTACTCCTGCT-3′ |
Western blot analysis
The BMSCs transfected with miR-214 or anti-miR-214
plasmids, or treated with JNK inhibitor or p38 inhibitor were lysed
on ice for 30 min in RIPA lysis buffer (Thermo Fisher Scientific)
supplemented with protease inhibitor. The protein concentration was
determined by Bradford assay (Bio-Rad) on a microplate
spectrophotometer (Tecan). A total of 40 µg proteins was
resolved on a 10% sodium dodecyl sulfate-polyacrylamide gel
electrophoresis (SDS-PAGE) gel and electro-transferred onto
polyvinylidene difluoride (PVDF) membranes (Pall Corporation, New
York, NY, USA). The membranes were incubated with the appropriate
primary antibodies [anti-fibroblast growth factor (FGF; 1:1,000;
9740), anti-phosphorylated (p-)JNK (1:2,000; 4668), anti-p-p38
(1:2,000; 4511) (all from Cell Signaling Technology, Danvers, MA,
USA) and GAPDH (1:5,000; H00002597-D01P; Abnova, Taiwan, China) at
4°C overnight. The membranes were then incubated with an
HRP-conjugated secondary antibody (Xi'an Kehao Biological
Engineering Co. Ltd, Xi'an, China) and developed by enhanced
chemiluminescence (ECL; Millipore, Billerica, MA, USA). Protein
expression was analyzed using an Odyssey Two-Color Infrared Imaging
System (LI-COR Biosciences, Lincoln, NE, USA).
Statistical analysis
All measurements are expressed as the means ± SD.
Comparisons were made using one/two-way analysis of variance
(ANOVA) followed by the Turkey post hoc test for multiple
comparisons. A value of P<0.05 was considered to indicate a
statistically significant difference.
Results
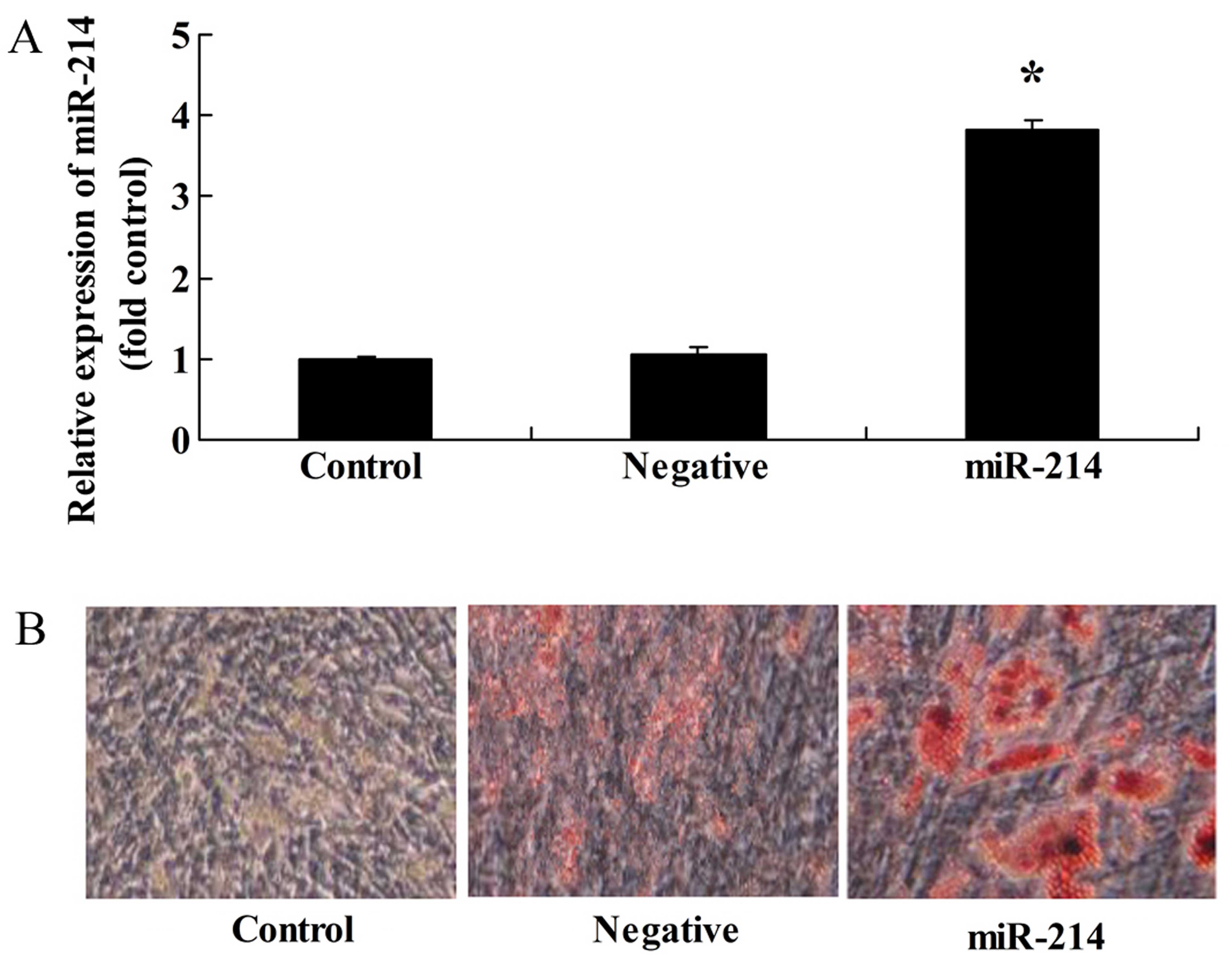
Overexpression of miR-214 promotes the
adipocyte differentiation of BMSCs
To investigate the miRNAs involved in the
differentiation of BMSCs, an miR-214 overexpresion plasmid was
constructed and transfrected into the BMSCs. Transfection with
miR-214 plasmid significantly increased miR-214 expression in the
BMSCs (Fig. 1A) and promoted the
adipocyte differentiation of BMSCs (Fig. 1B).
Overexpression of miR-214 suppresses ALP
activity and collagen type I (Col I), osteocalcin (OCN) and
osteopontin (OPN) gene expression in BMSCs
We then examined the effects of the overexpression
of miR-214 on ALP activity and Col I, OCN and OPN gene expression
in BMSCs. We performed ALP activity assay and RT-qPCR. As expected,
the overexpression of miR-214 significantly suppressed ALP activity
and inhibited Col I, OCN and OPN gene expression in the BMSCs
(Fig. 2). These results indicated
that the overexpression of miR-214 suppressed the osteogenic
differentiation of BMSCs.
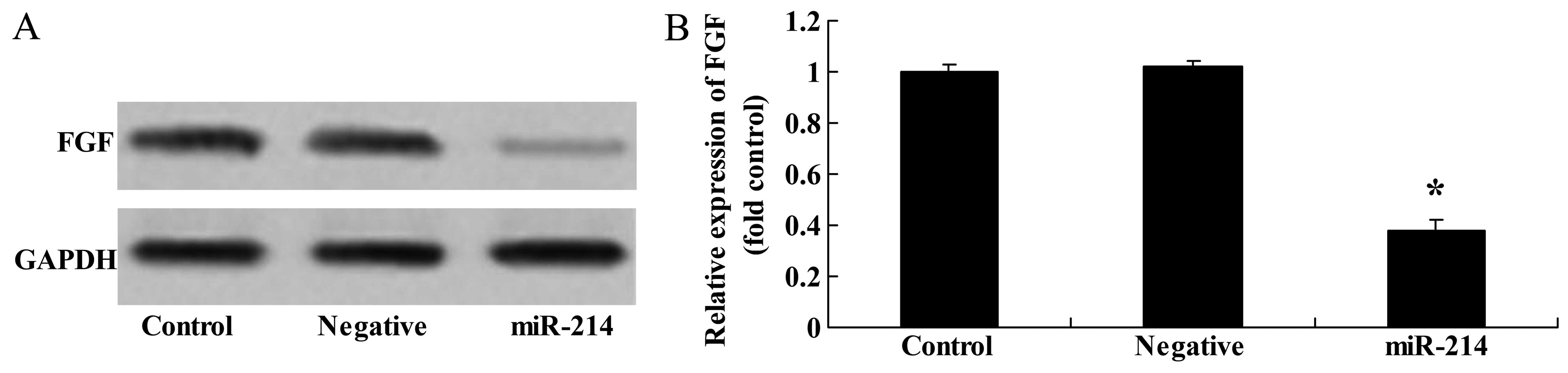
Overexpression of miR-214 suppresses FGF
protein expression in BMSCs
To determine whether FGF is a direct target of
miR-214 in BMSCs, we measured FGF protein expression in the BMSCs
following the overexpression of miR-214. As shown in Fig. 3, there was a significant
inhibition of FGF protein expression in the BMSCs following the
overexpression of miR-214.
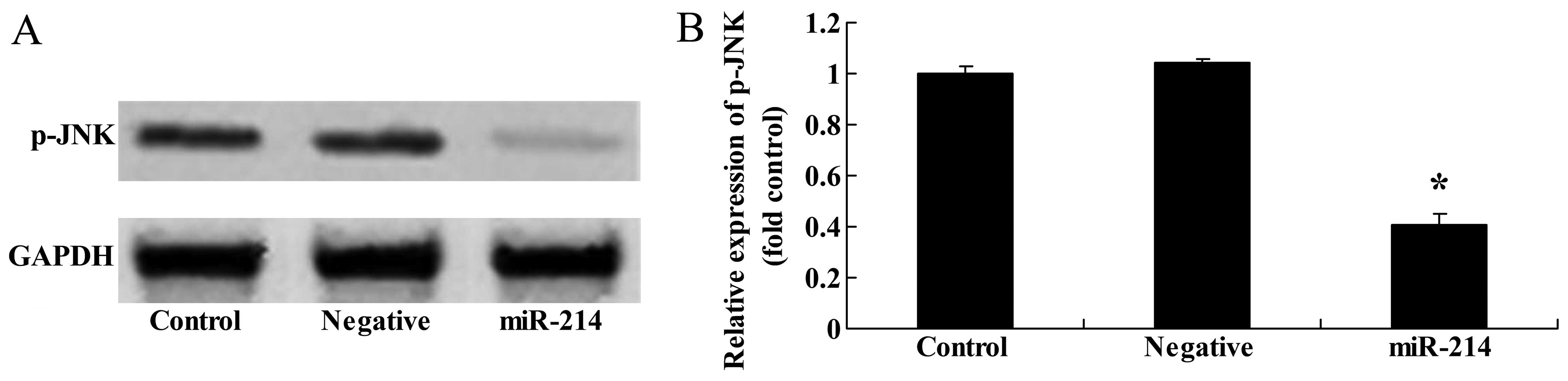
Overexpression of miR-214 suppresses
p-JNK protein expression in BMSCs
In addition, western blot analysis was applied to
examine the effects of miR-214 on JNK protein expression. Compared
with the controls, the overexpression of miR-214 significantly
suppressed p-JNK protein expression in the BMSCs (Fig. 4).
Overexpression of miR-214 suppresses
p-p38 protein expression in BMSCs
To determine whether the overexpression of miR-214
affects p38 protein expression in BMSCs, western blot analysis was
then performed to examine the effects of the overexpression of
miR-214 expression on p-p38 protein expression in BMSCs. The
overexpression of miR-214 significantly inhibited p-p38 protein
expression in the BMSCs (Fig.
5).
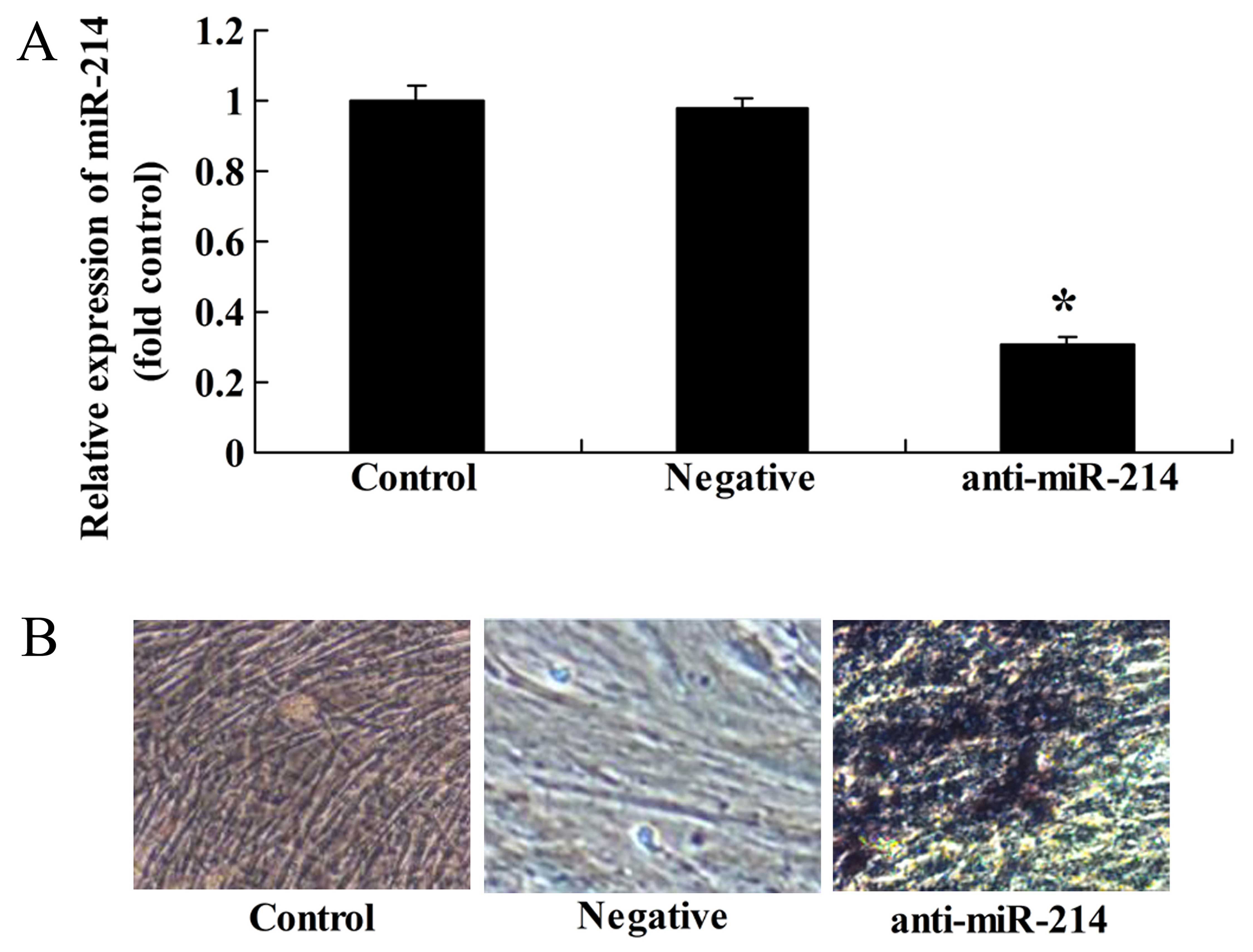
Downregulation of miR-214 promotes the
osteogenic differentiation of BMSCs
Conversely, we examined whether the downregulation
of miR-214 affects the osteogenic differentiation of BMSCs.
Anti-miR-214 plasmid was tansfected into the BMSCs. We found that
the inhibition of miR-214 significantly decreased miR-214
expression in the BMSCs (Fig. 6A)
and promoted the osteogenic differentiation of BMSCs (Fig. 6B).
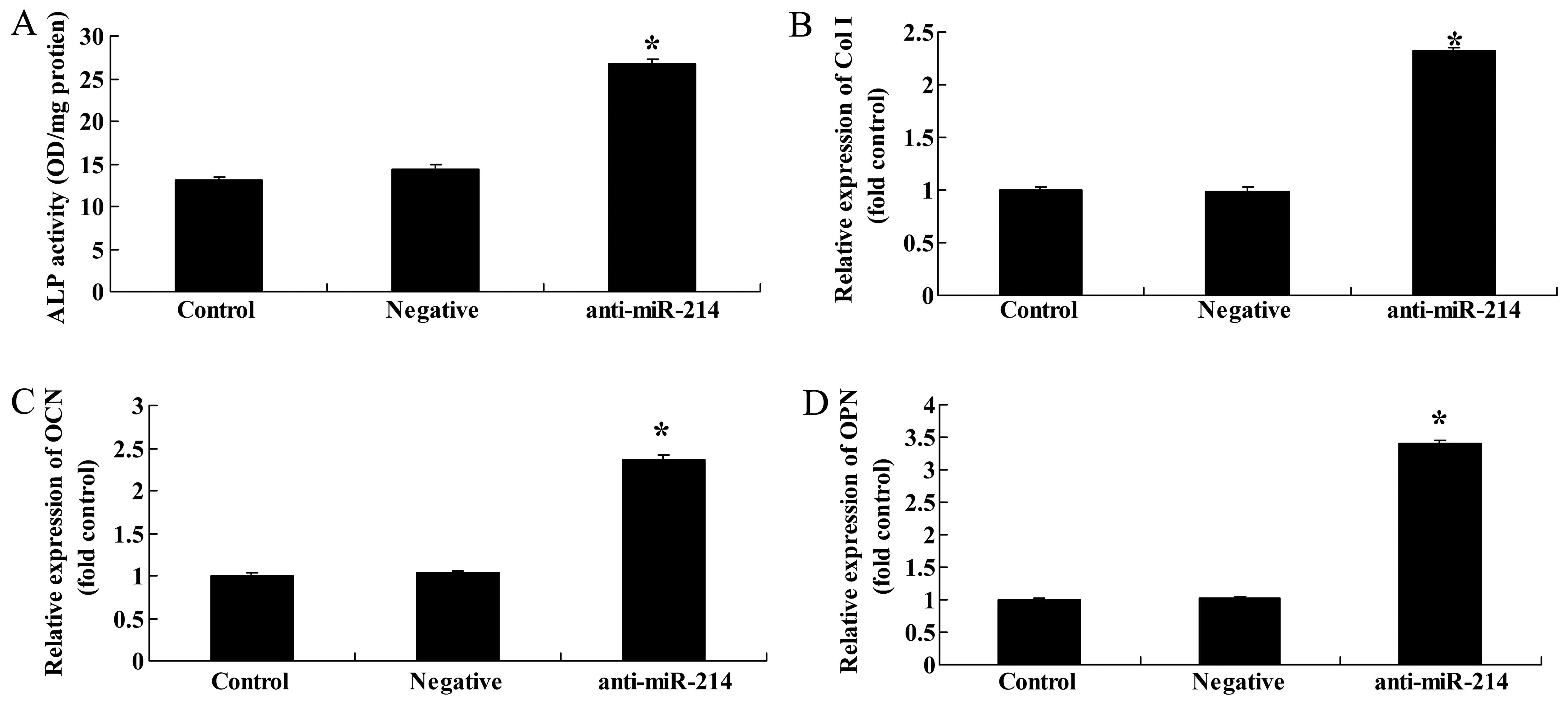
Downregulation of miR-214 enhances ALP
activity, and increases Col I, OCN and OPN gene expression in
BMSCs
We then examined whether the downregulation of
miR-214 also affects ALP activity, and Col I, OCN and OPN gene
expression in BMSCs. The downregulation of miR-214 significantly
increased ALP activity and promoted Col I, OCN and OPN gene
expression in the BMSCs (Fig.
7).
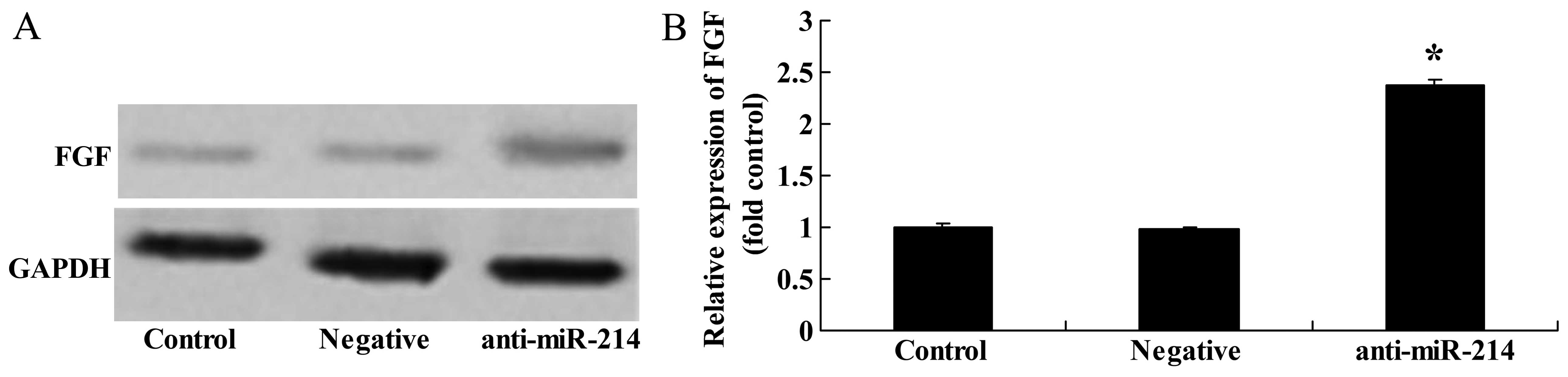
Downregulation of miR-214 increases FGF
protein expression in BMSCs
To further determine whether FGF is a direct target
of miR-214 in BMSCs, we measured FGF protein expression in BMSCs
following the downregulation of miR-214. There was a significant
increase in FGF protein expression in the BMSCs following the
downregulation of miR-214 (Fig.
8).
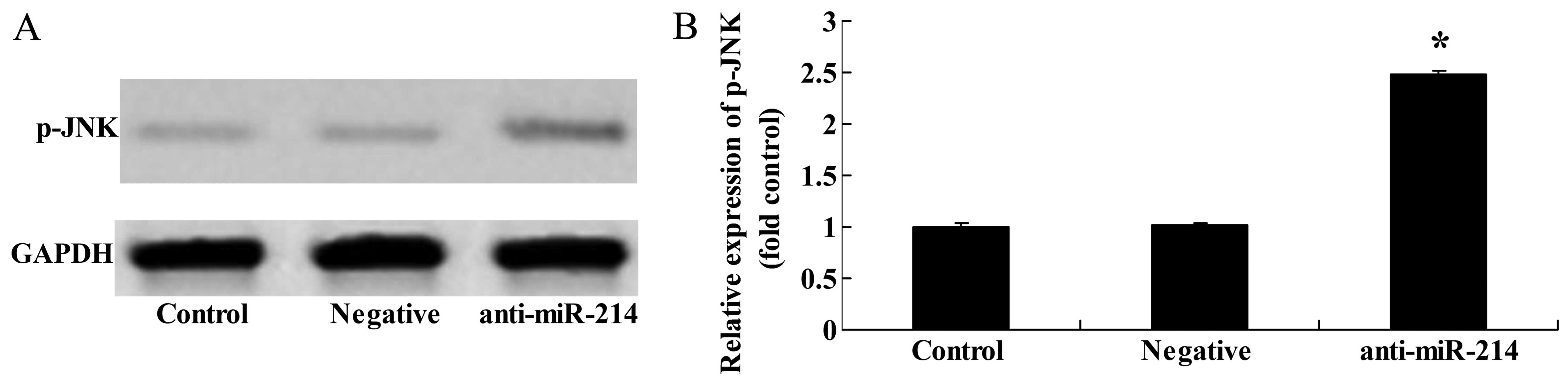
Downregulation of miR-214 increases p-JNK
protein expression in BMSCs
When miR-214 expression was downregulated, western
blot analysis was applied to examine the effects of miR-214
inhibition on JNK protein expression. The downregulation of miR-214
significantly increased p-JNK protein expression in BMSCs (Fig. 9).
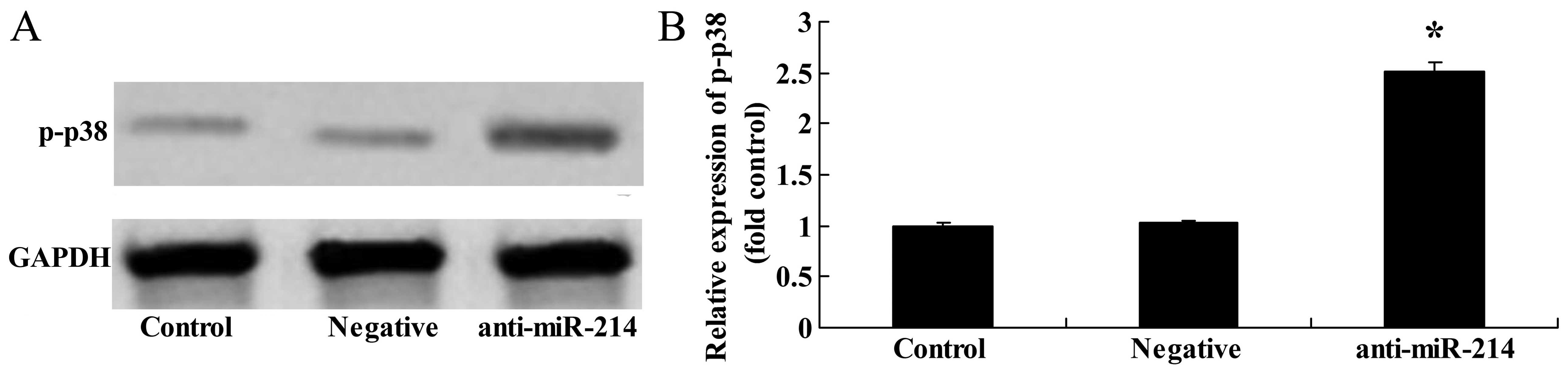
Downregulation of miR-214 increases p-p38
protein expression in BMSCs
To further determine whether the downregulation of
miR-214 affects p38 protein expression in BMSCs, western blot
analysis was then performed to examine the effects of the
downregulation of miR-214 expression on p-p38 protein expression in
BMSCs. p-p38 protein expression in the BMSCs was significantly
increased by the downregulation of miR-214 (Fig. 10).
Effect of the inhibition of JNK
expression on the adipocyte differentiation of BMSCs following the
overexpression of miR-214
To assess the significance of the inhibition of JNK
expression on the osteogenic differentiation of BMSCs following the
overexpression of miR-214, we treated the BMSCs with JNK inhibitor
during the course of osteogenic differentiation following the
overexpression of miR-214. The use of the JNK inhibitor, SP600125,
significantly suppressed the protein expression of p-JNK (Fig. 11A and B) and enhanced the
promoting effects of miR-214 on the adipocyte differentiation of
BMSCs (Fig. 11C).
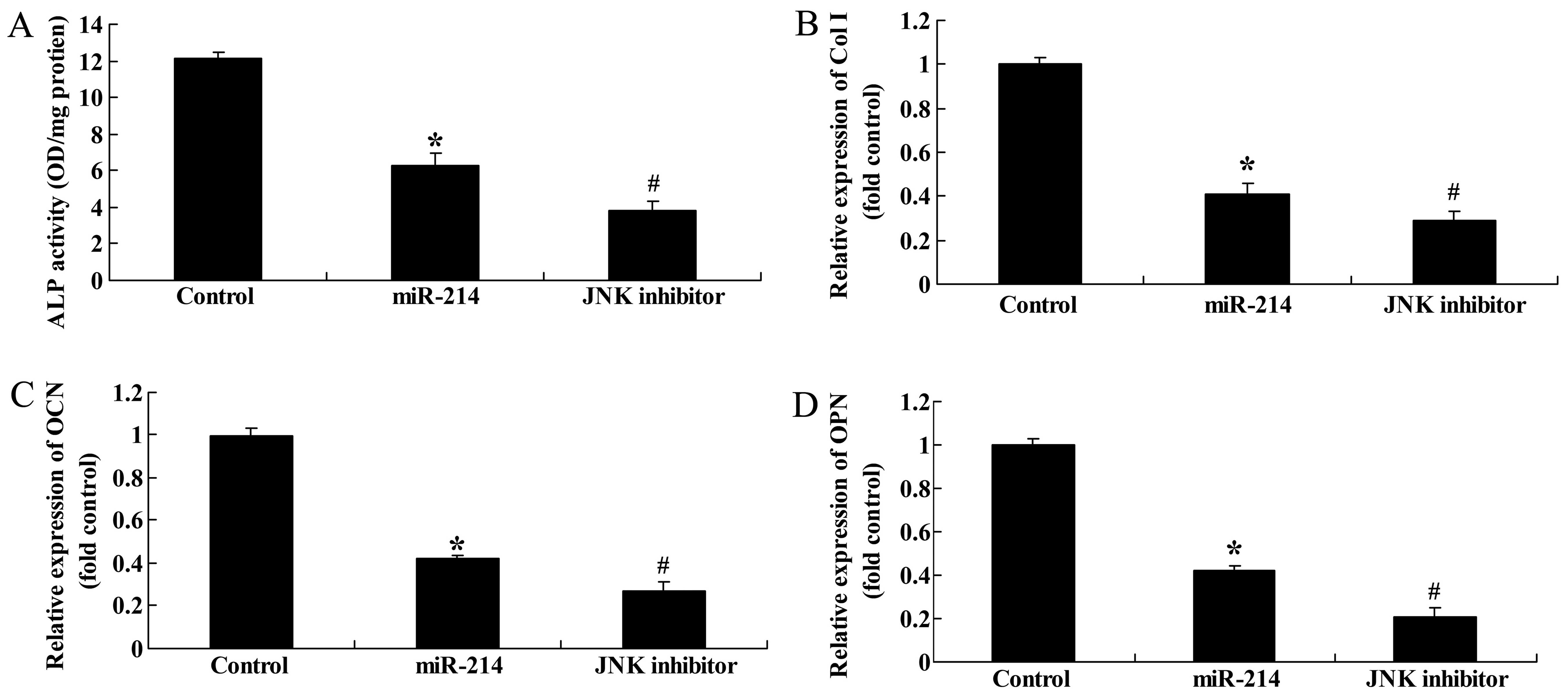
Effect of the inhibition of JNK
expression on ALP activity, and Col I, OCN and OPN gene expression
in BMSCs following the overexpression of miR-214
We then examined the effects of the inhibition of
JNK expression on ALP activity, and Col I, OCN and OPN gene
expression in BMSCs following the overexpression of miR-214. The
use of the JNK inhibitor, SP600125, significantly inhibited ALP
activity, and decreased Col I, OCN and OPN gene expression in the
BMSCs following the overexpression of miR-214 (Fig. 12).
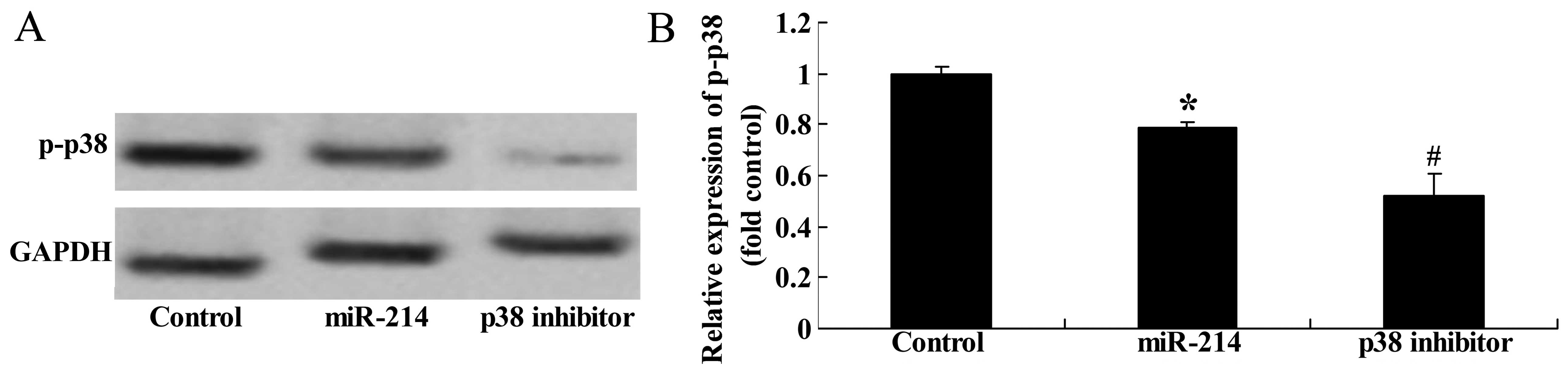
Effect of the inhibition of p38
expression on the adipocyte differentiation of BMSCs following the
overexpression of miR-214
To further assess the significance of the inhibition
of p38 expression on the adipocyte differentiation of BMSCs
following the overexpression of miR-214, we treated the BMSCs with
p38 inhibitor during the course of differentiation following the
overexpression of miR-214. The use of the p38 inhibitor, SB202190,
significantly suppressed the protein expression of p-p38 (Fig. 13A and B) and enhanced the
promoting effect of miR-214 on the adipocyte differentiation of
BMSCs (Fig. 13C).
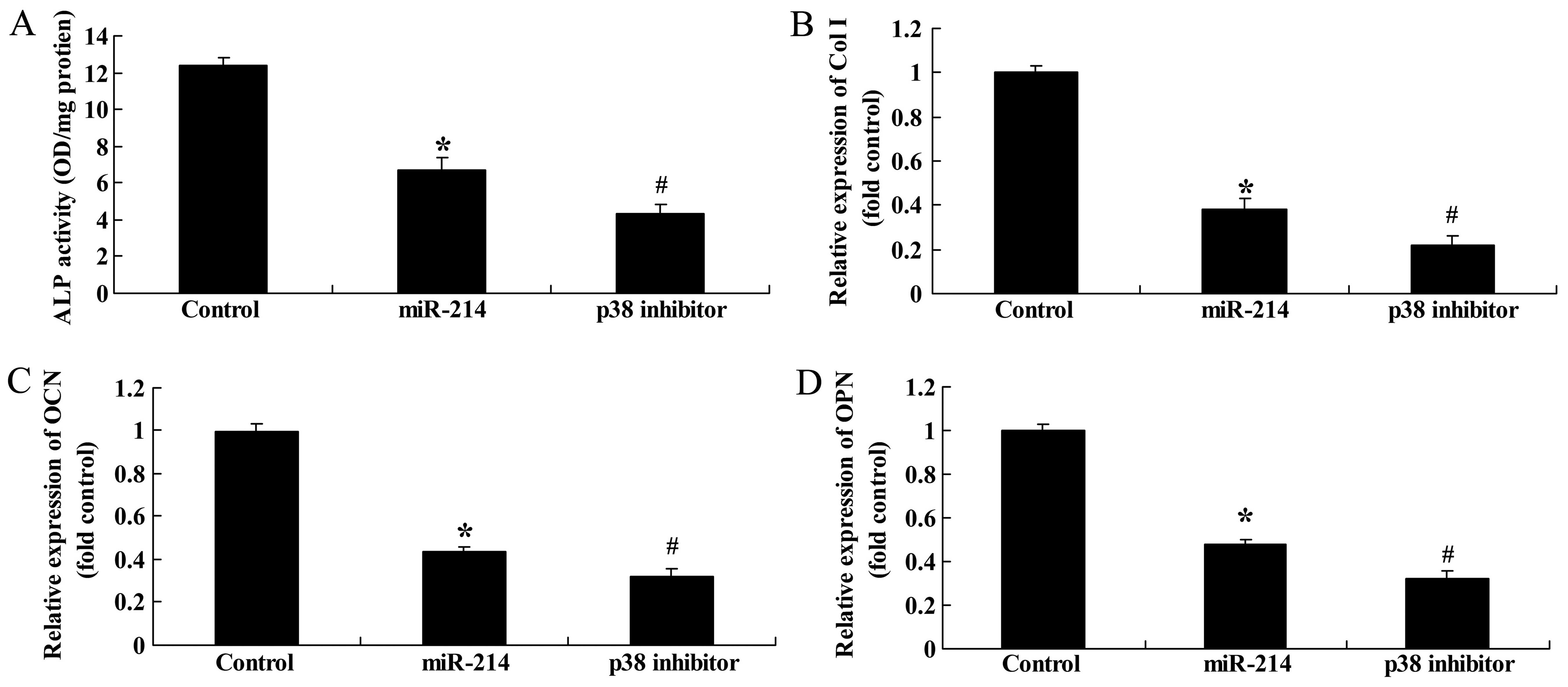
Effect of the inhibition of of p38
expression on ALP activity, and Col I, OCN and OPN gene expression
in BMSCs following the overexpression of miR-214
Finally, we examined the effect of the inhibition of
p38 expression on ALP activity, and Col I, OCN and OPN gene
expression in BMSCs following the overexpression of miR-214. The
use of the p38 inhibitor, SB202190, significantly inhibited ALP
activity, and decreased Col I, OCN and OPN gene expression in the
BMSCs following the overexpression of miR-214 (Fig. 14). Thee findings indicated that
the use of the p38 inhibitor enhanced the inhibitory effects of
miR-214 on the osteogenic differentiation of BMSCs.
Discussion
As adult stem cells from mesoblast, BMSCs do not
only differentiate into adult cells from mesoblasts, including
osteocytes, chondrocytes and adipocytes under the function of an
appropriate inducer for culture in vitro, but can also be
transdifferentiated into nerve cells from the neuroderm (4). However, whether BMSCs can
differentiate into nerve cells has become a top of extensive
investigation. The differentiation mechanisms can be studied from
different angles, which can be summarized as: cellular mechanisms
of toxicity, cellular fusion mechanism and transdifferentiation
(5). Cellular toxicity refers to
certain oxidative stress reaction and results in tearing of the
cytoskeleton of BMSCs, shrinking of the cytoplasm, and cell
shrinking as regards morphology (12). Cytoskeletal proteins are extended
to form vimineous and prominent structures similar to nerve cells
(13). Changes in biology and
molecular remodeling are derived from the toxicity of the inducer
on the BMSCs. Thus, they do not really differentiate into
neuronal-like cells (13).
Cellular fusion mechanism refers to when BMSCs and nerve cells are
cultured together, and thus two cells become fused, resulting in
gene fusion and the regulation of the gene expression of BMSCs,
making the morphology and functions similar to those of nerve cells
(14). In this study, the
overexpression of miR-214 suppressed osteogenic differentiation and
the downregulation of miR-214 activated thez osteogenic
differentiation of BMSCs.
ALP is a type of exoenzyme of osteoblasts and a
marker of osteogenic differentiation at the early stage. Its
expressive activity is an obvious characteristic of osteoblast
differentiation (15). ALP can be
used for identifying the degree of osteogenic differentiation at
the early stage for BMSCs (16).
During the later period of differentiation, OCN (also known as bone
γ-carboxylic glutamic acid containing protein) is a type of protein
composed and secreted by osteoblasts. Through the gene expression
level of OCN, we can identify osteoblasts, particularly at the
active stage of newly-formed osteoblasts. This study demonstrated
that the overexpression of miR-214 suppressed ALP activity, and Col
I, OCN and OPN gene expression. The downregulation of miR-214
enhanced ALP activity, and increased Col I, OCN and OPN gene
expression in the BMSCs. Ramazzotti et al (17) indicated that the inhibition of
miR-214 enhanced ALP activity and osterix expression in C2C12
cells.
Integrin is a heterosexual cell adhesion molecule.
In combination with the extracellular matrix, they form adhesion
plaque compounds with other molecules (FAK, talin, vinculi and
paxillin) to mediate interactions between cells and the
extracellular matrix, as well as mutual communications between
intracellular signals and extracellular cells, including force
signals. Adhesion plaque is an important signaling molecular
compound connecting with intracellular and extracellular
environments. FAK is a star molecule in adhesion plaque. Studies
have shown that FAK plays an important regulatory role in related
gene expression and the fate of cell differentiation. The study by
Torsoni et al (18)
demonstrated that mechanical stretching can regulate Tyr
phosphorylation at FAK 397 sites through integrin. Moreover, FAK
phosphorylation is an important regulatory signaling molecule of
β-MHC induced by mechanical stretching. Moreover, FAK is also an
important induction molecule of force signals relating to cellular
feeling matrix hardness. It can induce different matrix hardness
and adjust BMSCs induced by matrix hardness to differentiate into
osteoblast. FAK thus plays a decisive role in the process of force
signal transduction and the induction of stem cell differentiation.
In the present study, the upregulation of miR-214 suppressed FAK
protein expression and the downregulation of miR-214 activated FAK
protein expression in BMSCs. Yang et al suggested that
miR-214 inhibits osteogenic differentiation by targeting the
FGFR1/FGF signaling pathway in mesenchymal stem cells (19).
MAPK is a type of serine/threonine protein kinase,
and is distributed in cell plasma and is equipped with the double
phosphorylation of serine and tyrosine. The MAPK pathway in
eukaryocytes is extremely abundant (10). The pathway participates in
cellular activities through interaction, such as cellular genetic
expression, mitosis, metabolism, autonomic activities,
proliferation, apoptosis, or even differentiation. MAPK is a type
of serine/threonine protein kinase in cells, mainly including ERKs,
JNKs and p38 MAPK (8,10). Generally speaking, the ERK signal
pathway is activated by mitogens, such as the growth factor, IGFs
and FGFs (20). On the other
hand, the JNK signaling pathway and p38 MAPK signaling pathway are
mainly activated through bone morphogenetic proteins (BMPs) and
oxidative stress. Three signaling pathways of MAPKs participate in
the proliferation and differentiation of osteoblasts (10). In a previous study, following the
co-culture of BMSCs, it was found that the phosphorylation level of
p38 MAPK and ERK1/2 protein kinase increased (20). In the present study, we found that
the upregulation of miR-214 suppressed p-JNK and p-p38 protein
expression, and the downregulation of miR-214 increased p-JNK and
p-p38 protein expression in BMSCs.
The JNK/p38 MAPK signaling pathway exist in all
mammals, and participate in and regulate multiple biological
reactions (21). After MEK
identifies JNK/p38 MAPK selectively, the conservative
threonine-glycine-tyrosine sequence in JNK/p38 MAPK subtype is
phosphorylated (22). The
corresponding subtype of JNK/p38 MAPK is activated, so as to
activate JNK/p38 MAPK signaling. JNK/p38 MAPK in rats provides a
safeguard for the normal development of the skeleton (22). If coding gene in any MAPK signal
transduction pathway is deficient, such as MKK3, MKK6, p38a or
p38b, this results in the reduction of bone mass and osteoblast
differentiation defects (23). In
osteoblasts, Transforming growth factor-β (TGF-β)-activated kinase
1 (TAK1) TGF-β is the key site of activating upstream JNK/p38 MAPK
(24). In this study, we found
that the inhibition of JNK/p38 MAPK enhanced the inhibitory effect
of miR-214 on osteogenic differentiation and inhibited ALP activity
and Col I, OCN and OPN gene expression in BMSCs. This result
indicated that JNK/p38 MAPK plays a role in the effects of miR-214
on the osteogenic differentiation of BMSCs.
In the present study, we demonstrate that miR-214
inhibits the osteogenic differentiation of BMSCs, and results in
the suppression of the JNK and p38 pathways. Importantly, our data
indicate that miR-214 may have potential for use in chronic
inflammatory bone diseases.
References
|
1
|
Huang S, Xu L, Zhang Y, Sun Y and Li G:
Systemic and local administration of allogeneic bone marrow-derived
mesenchymal stem cells promotes fracture healing in rats. Cell
Transplant. 24:2643–2655. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Carvalho A, Pelaez-Vargas A, Hansford DJ,
Fernandes MH and Monteiro FJ: Effects of line and pillar array
microengineered SiO2 thin films on the osteogenic
differentiation of human bone marrow-derived mesenchymal stem
cells. Langmuir. 32:1091–1100. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Wang W, Du Z, Yan J, Ma D, Shi M, Zhang M,
Peng C and Li H: Mesenchymal stem cells promote liver regeneration
and prolong survival in small-for-size liver grafts: Involvement of
C-Jun N-terminal kinase, cyclin D1, and NF-κB. PLoS One.
9:e1125322014. View Article : Google Scholar
|
|
4
|
Baniwal SK, Shah PK, Shi Y, Haduong JH,
Declerck YA, Gabet Y and Frenkel B: Runx2 promotes both
osteoblastogenesis and novel osteoclastogenic signals in ST2
mesenchymal progenitor cells. Osteoporos Int. 23:1399–1413. 2012.
View Article : Google Scholar
|
|
5
|
Xu JF, Yang GH, Pan XH, Zhang SJ, Zhao C,
Qiu BS, Gu HF, Hong JF, Cao L, Chen Y, et al: Altered microRNA
expression profile in exosomes during osteogenic differentiation of
human bone marrow-derived mesenchymal stem cells. PLoS One.
9:e1146272014. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Skårn M, Namløs HM, Noordhuis P, Wang MY,
Meza-Zepeda LA and Myklebost O: Adipocyte differentiation of human
bone marrow-derived stromal cells is modulated by microRNA-155,
microRNA-221, and microRNA-222. Stem Cells Dev. 21:873–883. 2012.
View Article : Google Scholar
|
|
7
|
Qadir AS, Um S, Lee H, Baek K, Seo BM, Lee
G, Kim GS, Woo KM, Ryoo HM and Baek JH: miR-124 negatively
regulates osteogenic differentiation and in vivo bone formation of
mesenchymal stem cells. J Cell Biochem. 116:730–742. 2015.
View Article : Google Scholar
|
|
8
|
Dai Z, Li Y, Quarles LD, Song T, Pan W,
Zhou H and Xiao Z: Resveratrol enhances proliferation and
osteoblastic differentiation in human mesenchymal stem cells via
ER-dependent ERK1/2 activation. Phytomedicine. 14:806–814. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Liao QC, Xiao ZS, Qin YF and Zhou HH:
Genistein stimulates osteoblastic differentiation via p38
MAPK-Cbfa1 pathway in bone marrow culture. Acta Pharmacol Sin.
28:1597–1602. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Wu X, Chen S, Orlando SA, Yuan J, Kim ET,
Munugalavadla V, Mali RS, Kapur R and Yang FC: p85alpha regulates
osteoblast differentiation by cross-talking with the MAPK pathway.
J Biol Chem. 286:13512–13521. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Soleimani M and Nadri S: A protocol for
isolation and culture of mesenchymal stem cells from mouse bone
marrow. Nat Protoc. 4:102–106. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Laitinen A, Oja S, Kilpinen L, Kaartinen
T, Möller J, Laitinen S, Korhonen M and Nystedt J: A robust and
reproducible animal serum-free culture method for clinical-grade
bone marrow-derived mesenchymal stromal cells. Cytotechnology.
68:891–906. 2016. View Article : Google Scholar :
|
|
13
|
Da Silva JS and Hare JM: Cell-based
therapies for myocardial repair: Emerging role for bone
marrow-derived mesenchymal stem cells (MSCs) in the treatment of
the chronically injured heart. Methods Mol Biol. 1037:145–163.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Lv S, Liu G, Sun A, Wang J, Cheng J, Wang
W, Liu X, Nie H and Guan G: Mesenchymal stem cells ameliorate
diabetic glomerular fibrosis in vivo and in vitro by inhibiting
TGF-β signalling via secretion of bone morphogenetic protein 7.
Diab Vasc Dis Res. 11:251–261. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Tao K, Xiao D, Weng J, Xiong A, Kang B and
Zeng H: Berberine promotes bone marrow-derived mesenchymal stem
cells osteogenic differentiation via canonical Wnt/β-catenin
signaling pathway. Toxicol Lett. 240:68–80. 2016. View Article : Google Scholar
|
|
16
|
Kim SE, Jeon O, Lee JB, Bae MS, Chun HJ,
Moon SH and Kwon IK: Enhancement of ectopic bone formation by bone
morphogenetic protein-2 delivery using heparin-conjugated PLGA
nanoparticles with transplantation of bone marrow-derived
mesenchymal stem cells. J Biomed Sci. 15:771–777. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Ramazzotti G, Bavelloni A, Blalock W,
Piazzi M, Cocco L and Faenza I: BMP-2 Induced expression of PLCβ1
that is a positive regulator of osteoblast differentiation. J Cell
Physiol. 231:623–629. 2016. View Article : Google Scholar
|
|
18
|
Torsoni AS, Constancio SS, Nadruz W Jr,
Hanks SK and Franchini KG: Focal adhesion kinase is activated and
mediates the early hypertrophic response to stretch in cardiac
myocytes. Circ Res. 93:140–147. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Yang L, Ge D, Cao X, Ge Y, Chen H, Wang W
and Zhang H: miR-214 attenuates osteogenic differentiation of
mesenchymal stem cells via targeting FGFR1. Cell Physiol Biochem.
38:809–820. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Martinez-Lopez N and Singh R: ATGs:
Scaffolds for MAPK/ERK signaling. Autophagy. 10:535–537. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Dong J, Wang J, He Y, Li C, Zhou A, Cui J,
Xu W, Zhong L, Yin Y, Zhang X and Wang H: GHIP in Streptococcus
pneumoniae is involved in antibacterial resistance and elicits a
strong innate immune response through TLR2 and JNK/p38 MAPK. FEBS
J. 281:3803–3815. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Lee CM, Onésime D, Reddy CD, Dhanasekaran
N and Reddy EP: JLP: A scaffolding protein that tethers JNK/p38
MAPK signaling modules and transcription factors. Proc Natl Acad
Sci USA. 99:14189–14194. 2002. View Article : Google Scholar
|
|
23
|
Pang HY, Liu G and Liu GT: Compound FLZ
inhibits lipopolysaccharide-induced inflammatory effects via
downregulation of the TAK-IKK and TAK-JNK/p38 MAPK pathways in
RAW264.7 macrophages. Acta Pharmacol Sin. 30:209–218. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Kuroyanagi G, Otsuka T, Yamamoto N,
Matsushima-Nishiwaki R, Kozawa O and Tokuda H: Resveratrol
suppresses TGF-β-induced VEGF synthesis in osteoblasts: Inhibition
of the p44/p42 MAPKs and SAPK/JNK pathways. Exp Ther Med.
9:2303–2310. 2015.PubMed/NCBI
|