Introduction
Temporomandibular joint disorder (TMD) is a common
disease of the stomatognathic system. It is a degenerative disease,
accompanied by internal derangement or osteoarthritis (OA), which
is characterized by progressive articular cartilage destruction and
decreased physical capabilities. Underlying these symptoms is the
limited self-regenerating capacity of cartilage. Certain drugs,
such as etanercept, infliximab and adalimumab, have been used
clinically to treat TMD following the failure of traditional
injection with corticosteroids or hyaluronic acid. However, a
substantial proportion of patients still fail to respond to these
biological therapies (1).
Mesenchymal stem cells (MSCs) are increasingly being
considered attractive biological therapeutic agents for the
treatment of inflammatory diseases and for the stimulation of
tissue regeneration. They possess multi-differentiation potential
and can be isolated from various human tissues, such as the bone
marrow, adipose tissue, the synovium and periosteum. Among these
sources, synovial fluid plays an important role in regulating
temporomandibular joint (TMJ) physiology. Jones et al
(2) were the first to identify
MSCs in the synovial fluid of swollen osteoarthritic knee joints.
These cells may originate from the MSCs in synovial tissue
(3), as both populations express
similar cell surface markers (4).
Whereas the majority of subsequent studies have focused on large
joints, information regarding synovial fluid-derived MSCs (SFMSCs)
in the TMJ is limited. Koyama et al demonstrated that human
pluripotent cells can be isolated from the synovial fluid of
patients with TMD (5). Analogous
to MSCs that are isolated from the fragments of synovial tissue and
synovial fluid, we hypothesized that the SFMSCs most likely
originate from the disrupted intimal synovium (6). SFMSCs of the human TMJ can be
obtained by arthrocentesis, which is a commonly used and less
invasive method compared with arthroscopy or open operation to
obtain sections of the synovium. Therefore, these cells constitute
a promising resource for cell-directed articular cartilage
repair.
Both the genetic background of MSCs and the local
inflammatory milieu may affect the ability of MSCs to modulate
disease activity in vitro (7,8).
The number of SFMSCs reportedly increases in early human OA, and
this increase correlates with cartilage degeneration and the
severity of OA in the knee (9–11).
Thus, these cells may play a potential role in joint homeostasis.
Neidhart et al (12) found
that adherent synovial fluid cells derived from patients with
rheumatoid arthritis (RA) mediated cartilage destruction,
independent of the accompanying synovial tissue hyperplasia.
Heldens et al (13)
demonstrated that human MSC chondrogenesis was inhibited in a
diseased joint environment or upon treatment with pro-inflammatory
cytokines. In another study, the amount of chondrogenic
differentiation of mesenchymal progenitor cell (MPC) pellets in
operated knees was significantly lower than that observed in
control subjects. Additional treatment with interleukin (IL)-1β
also affected the degree of chondrogenic differentiation in cell
pellets derived from bone marrow-derived MSCs (14). To the best of our knowledge,
however, the effect of the inflammatory milieu on SFMSCs delivered
to human TMJs remains poorly understood. To address this issue, in
this study, we character-ized the inflammatory factors that are
secreted by SFMSCs following stimulation with IL-8, IL-1β, IL-6,
IL-10, tumor necrosis factor (TNF)-α and IL-12p in vitro. We
found that the expression of only IL-6 and IL-8 increased
significantly upon IL-1β stimulation, and that this was independent
of IL-8, IL-6, IL-10, TNF-α and IL-12p stimulation.
IL-1β is a pivotal pro-inflammatory cytokine that
mediates a variety of host defense processes, including
inflammation and cellular response to injury, which can also lead
to joint destruction if left unchecked (15). The level of IL-1β is significantly
higher in the synovial fluid from patients with TMD and chondral
damage than in the synovial fluid from asymptomatic volunteers
(16–18). IL-1β reduces the expression of
type II collagen and aggrecan by the increased secretion of matrix
metalloproteinases (MMPs) by chondrocytes (19). This requires the activation of the
nuclear factor-κB (NF-κB) pathway, which has been described as the
link between inflammation and joint cartilage degeneration in OA
(20–22). In this study, we further evaluated
the chondrogenic differentiation potency and intracellular
signaling pathways that are mediated by IL-1β in SFMSCs.
Materials and methods
Ethics statement
This study was approved by the Institutional Ethics
Board of the Hospital of Stomatology, Sun Yat-sen University,
Guangzhou, China. Written informed consent was obtained from the
sample providers prior to participation in this study.
Assay of inflammatory cytokines in the
TMJ
Synovial fluid samples were obtained from 19
patients with TMD (19 laterals) during arthrocentesis of the TMJ.
These patients were aged between 16 and 57 years; 16 were female
and 3 were male, and all were free of other systemic diseases. The
patients with TMD were evaluated using the dysfunction index (DI)
described in the study by Fricton JR and Schiffman (23). The synovial fluid samples were
withdrawn with 2 ml of lidocaine (containing 25% vitamin
B12). We measured the levels of IL-8, IL-1β, IL-6,
IL-10, TNF-α and IL-12p in the synovial fluid samples with a BD
Cytometric Bead Array (CBA) Human Soluble Protein Master Buffer kit
(Human Inflammatory Cytokine kit; Cat. no. 551811; BD Biosciences,
Bedford, MA, USA). The cytokine concentrations in the synovial
fluid were calculated using the following formula: C =
Casp/[(ODinj − ODasp)/ODinj],
Casp refers to the levels of IL-8, IL-1β, IL-6, IL-10,
TNF-α and IL-12p in the synovial fluid samples; ODinj
refers to the optical density of lidocaine containing 25% vitamin
B12 before injection; ODasp refers to the
optical density of lidocaine containing 25% vitamin B12
after injection, as previously described (16).
Human SFMSC isolation and expansion
The collected synovial fluid samples were
centrifuged at 300 × g for 5 min at room temperature, and the
supernatant was used in the inflammatory cytokine assay. The
sediment was then cultured with complete culture medium [α minimum
essential medium supplemented with 10% fetal bovine serum and 1X
GlutaMAX (both from Gibco, Grand Island, NY, USA) at 37°C in a 5%
CO2 incubator. Portions of the cells from each sample
were mixed and expanded in a monolayer culture for further
experimentation. The cells were passaged at a density of 500
cells/cm2.
Surface antigen identification
Surface markers, including CD90 (1:20; Cat. no.
562245; BD Biosciences), CD105 (1:11; Cat. no. 130-094-941;
Miltenyi Biotec, Begisch Gladbach, Germany), CD73 (1:20), CD44
(1:20), and CD45/CD34/CD11b/CD19/HLA-DR (1:20) (Cat. no. 562245;
all from BD Biosciences), were detected using an FC500 flow
cytometer and MXP software, and the data were analyzed using CXP
software (both from Beckman Coulter, Brea, CA, USA).
Osteogenic differentiation
The SFMSCs were seeded in 12-well plates at a
density of 5,000 cells/cm2. Following adherence, the
complete culture medium was replaced with osteogenic induction
medium consisting of H-DMEM, 10% fetal bovine serum (FBS) (both
from Gibco), 10 mM sodium β-glycerophosphate (Santa Cruz
Biotechnology, Inc., Santa Cruz, CA, USA), 10 nM
1,25-dihydroxyvitamin D3 (Sigma-Aldrich, St. Louis, MO, USA) and 50
μg/l ascorbic acid-2-phosphate (Wako, Tokyo, Japan). The
culture medium was replaced every 3 days. Following 28 days of
induction, osteogenic differentiation was assessed by staining with
a fresh 0.1% Alizarin Red solution (Cat. no. 130-22-3; Santa Cruz
Biotechnology, Inc.) for 30 min at 37°C.
Adipogenic differentiation
The SFMSCs were seeded in 12-well plates at a
density of 5,000 cells/cm2. Following adherence, the
complete culture medium was replaced with adipogenic induction
medium consisting of H-DMEM, 10% FBS (both from Gibco), 200 mM
indomethacin (Sigma-Aldrich), 0.5 mM isobutylmethylxanthine, 1 mM
dexamethasone and 10 mg/ml insulin (all from MP Biomedicals,
Irvine, CA, USA). The medium was replaced every 3 days. After 28
days, cell staining was performed with a fresh 0.3% Oil Red O
solution (Cat. no. 1320-06-5, Santa Cruz Biotechnology, Inc.) for 3
min.
Chondrogenic differentiation
The SFMSCs were harvested using 0.25% trypsin and
approximately 3×105 cells were transferred to a 15 ml
centrifuge tube. They were then centrifuged at 450 × g for 8 min,
and 500 μl of chondrocyte differentiation induction medium
[H-DMEM, 1X ITS-A (both from Gibco), 100 nM dexamethasone (MP
Biomedicals), 50 mM ascorbic acid, 40 mg/ml proline (both from
Sigma-Aldrich), and 10 ng/ml transforming growth factor-β1
(PeproTech, Rocky Hill, NJ, USA)] were added. The medium was
refreshed every 3 days. Following culture for 28 days, the
chondrogenic pellets were fixed with 4% formalin and
paraffin-embedded. Chondrogenic differentiation was assessed by
0.1% Safranin O staining (Cat. no. S2255; Seebio, Shanghai, China)
and immunohistochemical staining for collagen II [rabbit anti-human
collagen type II antibodies (Sigma-Aldrich) 1:80]. The control
group was then incubated with complete culture medium.
Neurogenic differentiation
The cells were plated in 12-well plates and cultured
until they reached 70% confluence, after which the complete culture
medium was replaced with a neuronal induction medium that consisted
of Neurobasal-A media containing 1X B-27 Supplement (both from
Gibco), 20 ng/ml recombinant human epidermalgrowth factor (rhEGF),
40 ng/ml basic fibroblast growth factor (bFGF) (both from
PeproTech), 10 ng/ml brain-derived neurotrophic factor (BDNF;
ProSpec, East Brunswick, NJ, USA), 1 mM
N6,2′-O-dibutyryladenosine 3′:5′ cyclic monophosphate
(dbCAMP; Sigma-Aldrich), 0.5 mM isobutylmethylxanthine (MP
Biomedicals), and 10 ng/ml fibroblast growth factor (FGF)-8
(PeproTech) for 24 h to induce differentiation. The control group
was incubated in complete culture medium. Neuronal differentiation
was assessed by immunofluorescence staining for glial fibrillary
acidic protein (GFAP; mouse anti-human antibody to GFAP, 1:200;
Cat. no. MAB360; Millipore, Billerica, MA, USA).
RT-qPCR
Total RNA from the chondrogenic differentiation
pellets and other cell samples was extracted using RNeasy mini kit
reagents (Qiagen, Valencia, CA, USA) and TRIzol reagent (Roche,
Indianapolis, IN, USA), respectively. cDNA was synthesized using
the Transcriptor First Strand cDNA Synthesis kit (Roche). The
relative expression levels of Runt-related transcription factor 2
(RUNX-2), osteocalcin (OCN), peroxisome proliferator
activated receptor gamma 2 (PPARG2), lipoprotein lipase
(LPL), SRY-box 9 (SOX9) and MMP13 were
quantified using a SYBR-Green qPCR kit (Roche). The primer
nucleotide sequences are listed in Table I. The 2−ΔΔCt method was
used to analyze the relative gene expression levels by normalizing
against the levels of glyceraldehyde 3-phosphate dehydrogenase
(GAPDH) as an endogenous control and calibrating for amplification
efficiency. ΔΔCt was calculated using the following formula:
(Cttarget gene − Ctgapdh)sample −
(Cttarget gene − Ctgapdh) control.
 | Table IOligonucleotide primers used in
RT-qPCR. |
Table I
Oligonucleotide primers used in
RT-qPCR.
| Gene | Primer
sequence | Product size
(bp) |
|---|
| GAPDH | F:
GACAGTCAGCCGCATCTTCT | |
| R:
TTAAAAGCAGCCCTGGTGAC | 178 |
| RUNX2 | F:
TCAACGATCTGAGATTTGTGGG | |
| R:
GGGGAGGATTTGTGAAGACGG | 81 |
| OCN | F:
CCACCGAGACACCATGAGAG | |
| R:
TCAGCCAACTCGTCACAGTC | 267 |
| PPARG2 | F:
GCAAACCCCTATTCCATGCTG | |
| R:
CACGGAGCTGATCCCAAAGT | 167 |
| LPL | F:
CAAGAGTGAGTGAACAAC | |
| R:
AATTATGCTGAAGGACAAC | 189 |
| SOX9 | F:
ACACACAGCTCACTCGACCTTG | |
| R:
AGGGAATTCTGGTTGGTCCTCT | 104 |
| MMP13 | F:
GACTGGTAATGGCATCAAGGGA | |
| R:
CACCGGCAAAAGCCACTTTA | 149 |
Detection of actate dehydrogenase (LDH)
cytotoxicity
The human SFMSCs were seeded into 96-well plates at
10,000 cells/well and then treated with 10 ng/ml IL-1β, 10 ng/ml
TNF-α, 10 ng/ml IL-8/IL-1β/IL-6/IL-10/TNF-α/IL-12p compound, or 10
μM NF-κB inhibitor (BAY11-7082; Cat. no. 19542-67-7; Santa
Cruz Biotechnology, Inc.) for 12 h. We then measured the activity
of LDH released from the cells using the cytotoxicity detection kit
(LDH; Roche) following the manufacturer's instructions.
Detection of cytokines secreted by
SFMSCs
We measured the concentrations of IL-8, IL-1β, IL-6,
IL-10, TNF-α and IL-12p in the culture medium of SFMSCs following
treatment with cytokines in vitro using a BD CBA Human
Soluble Protein Master Buffer kit (Human Inflammatory Cytokine kit;
Cat. no. 551811; BD Biosciences).
Immunofluorescence staining for
NF-κB
The human SFMSCs were plated in 24-well plates and
allowed to grow onto cover-slips placed in each well. At
approximately 50% confluence, the wells received either the same
volume of diluting agent as the control group or 10 ng/ml IL-1β for
2 h. The medium was then aspirated, and the cells were washed twice
with phosphate-buffered saline (PBS). The cells were then fixed in
4% paraformaldehyde for 30 min prior to treatment with 3% Triton
X-100 for 15 min at approximately 26°C. The cells were blocked for
2 h with 5% bovine serum albumin. After washing with PBS, the cells
were stained with rabbit anti-human NF-κB p65 antibody (1:400; Cell
Signaling Technology, Boston, MA, USA) for 18 h at 4°C. After
washing with 1X PBS, the cells were incubated with the secondary
antibody, DyLight 549 goat anti-rabbit IgG antibody (1:100; Cat.
no. E032320-01; EarthOx LLC, San Francisco, CA, USA) for 60 min at
37°C. After washing with PBS, the cells were incubated with
4′,6-diamidino-2-phenylindole (DAPI; Cell Signaling Technology) for
5 min. The cells were viewed under a fluorescent microscope
(Olympus, Tokyo, Japan). As a specificity control, the cells were
treated using the same process, but without incubation with a
primary antibody.
Dual-luciferase reporter assay
The human SFMSCs were seeded into 96-well plates at
5,000 cells/well. pGL3-basic-NF-κB-REVector and
pRL-SV40-Renilla were transfected into the SFMSCs using
ViaFect transfection reagent (both from Promega, Madison, WI, USA)
according to the manufacturer's instructions. Following incubation
for 48 h, cells were treated with the same volume of diluting agent
as the control group or 10 ng/ml IL-1β for 2 h. Luciferase activity
in the SFMSCs was detected using the GloMax-MultiDetection system
and the Dual-Luciferase Reporter assay kit (Cat. no. E4030) (both
from Promega).
CBA assay
The human SFMSCs were seeded into 10-mm plates and
cultured until they reached 80% confluence. The cells were treated
with cytokines and/or inhibitors. Inhibitors [BAY11-7082 (Cat. no.
19542-67-7; Santa Cruz Biotechnology, Inc.), anti-TNF-α (167348;
Ebioscience, San Diego, CA, USA)] were added 1 h prior to cytokine
treatment. The empty plate using the same culture medium, but
without cells as the negative control. IL-6 and IL-8 expression in
the culture medium was determined using a CBA kit (Human
Inflammatory Cytokine kit; Cat. no. 551811; BD Biosciences).
Double-stranded DNA was detected using Quant-iT PicoGreen dsDNA
reagent (Invitrogen, Carlsbad, CA, USA) as an endogenous
control.
Western blot analysis
Cellular proteins were extracted using RIPA buffer
(Cell Signaling Technology) and separated by sodium dodecyl
sulfate-polyacrylamide gel electrophoresis (SDS-PAGE). The tissue
samples were electrophoresed in a stacking gel (5% w/v) for 30 min
and run on a running gel (10% w/v) for 120 min. Proteins were
transferred electrophoretically onto PVDF membranes by applying 200
mA for 75 min. The membranes were first reacted with antibodies
against NF-κB p65 (1:1,000; Cat. no. 3033; Cell Signaling
Technology) and GAPDH (1:2,000; Cat. no. Mab5465; MultiSciences
Biotech Co., Hangzhou, China) overnight at 4°C, and then with IgG
conjugated to horseradish peroxidase (Cat. no. 7076; Cell Signaling
Technology) for 1 h at room temperature. The target bands were
visualized by an enhanced chemiluminescence method. The corrected
optical density of the protein bands was quantified using a Bio-Rad
image analyzer (Bio-Rad, Hercules, CA, USA).
Effect of IL-1β on the chondrogenic
differentiation of SFMSCs
The chondrogenic differentiation procedure was
performed as described above, with the addition of the
corresponding cytokines. In order to evaluate the chondrogenic
differentiation potential upon stimulation with rhIL-1β, rhIL-6 or
rhIL-8 (PeproTech), we measured the production of sulfated
glycosaminoglycans (sGAG) in the chondrogenic pellets stimulated
with rhIL-6 or rhIL-8, and the mRNA expression of SOX9 and
MMP13 in the chondrogenic pellets, mediated by rhIL-1β.
sGAG assay
The samples were digested overnight at 56°C using
100 μl of a 50 μg/ml proteinase K solution in 100 mM
Na2HPO4 (pH 8.0). Proteinase K was then
inactivated by heating the preparation for 10 min at 90°C.
Following centrifugation, 500 μl of working DMMB solution
were added to 50 μl of proteinase K-treated or untreated
sample, and the mixture was vigorously vortexed for 30 min to
promote complete GAG/DMMB complexation. The samples were then
centrifuged at 12,000 × g for 10 min. The supernatant was
discarded, and the pellet was dissolved with DMMB decomplexation
solution and vortexed for 30 min. The absorbance at 656 nm was used
to quantify the sGAG levels.
Statistical analysis
In this study, the numerical data are expressed as
the means ± standard deviation (SD). Statistical significance was
evaluated using the Student's t-test, and a value of p<0.05 was
considered to indicate a statistically significant difference.
Results
Cytokine expression in the synovial fluid
of patients with TMD
In the present study, we randomly selected 19
patients with TMD (19 laterals) to evaluate the IL-8, IL-1β, IL-6,
IL-10, TNF-α and IL-12p expression levels in the synovial fluid of
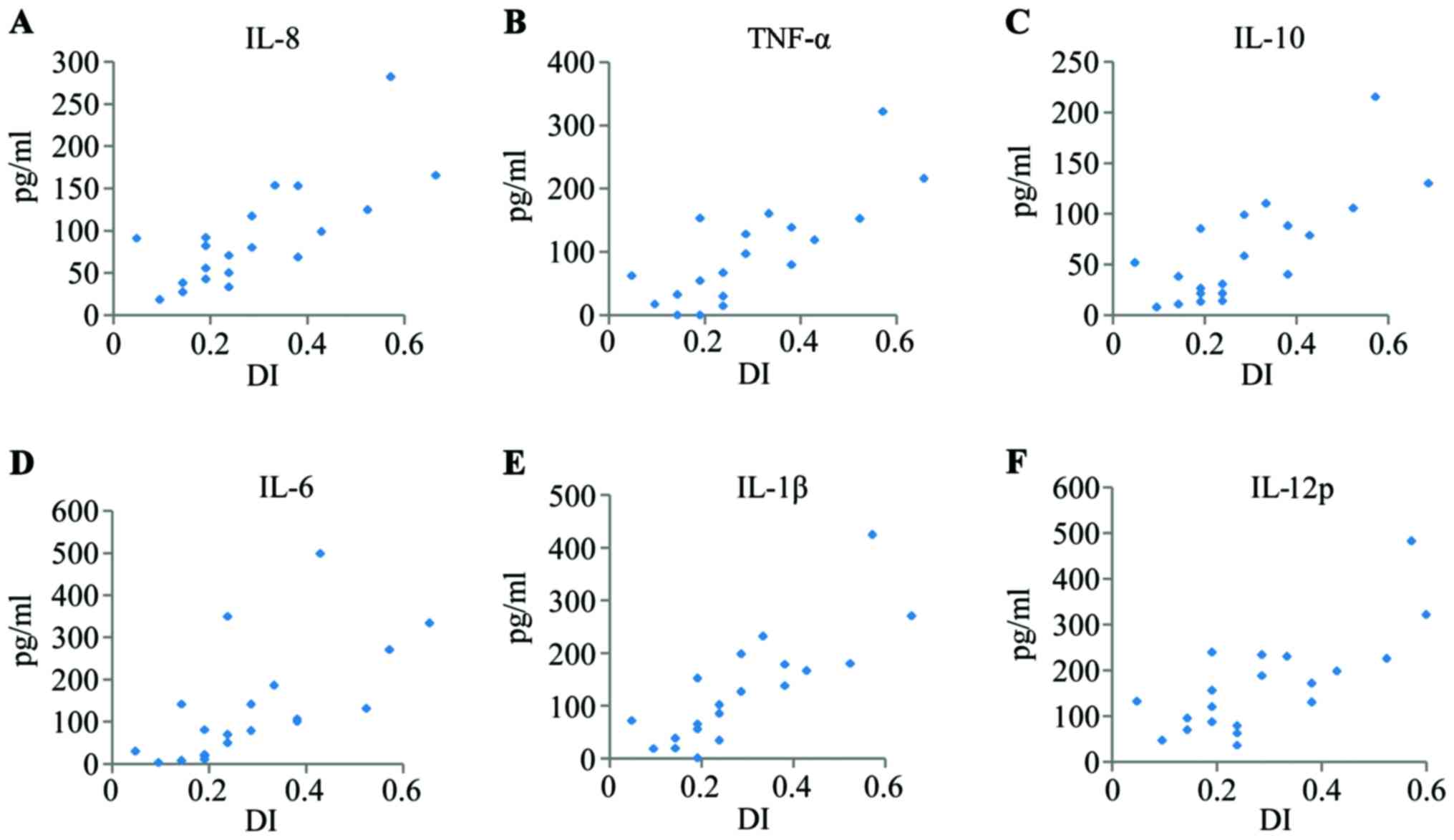
the TMJ. The levels of these cytokines were 18.23–282.24 pg/ml
(average 88.25 pg/ml), 0.57–424.01 pg/ml (average 120.21 pg/ml),
2.80–498.44 pg/ml (average 120.73 pg/ml), 7.63–215.54 pg/ml
(average 58.61 pg/ml), 0.00–321.55 pg/ml (average 85.48 pg/ml), and
36.39–482.77 pg/ml (average 157.32 pg/ml), respectively. The DI of
these patients with TMD ranged from 0.05 to 0.57 (average 0.27). A
more detailed summary of the cytokine expression profiles in the
synovial fluid of the patients with TMD is depicted in Fig. 1.
Characteristics of SFMSCs in vitro
After the synovial fluid samples were plated and
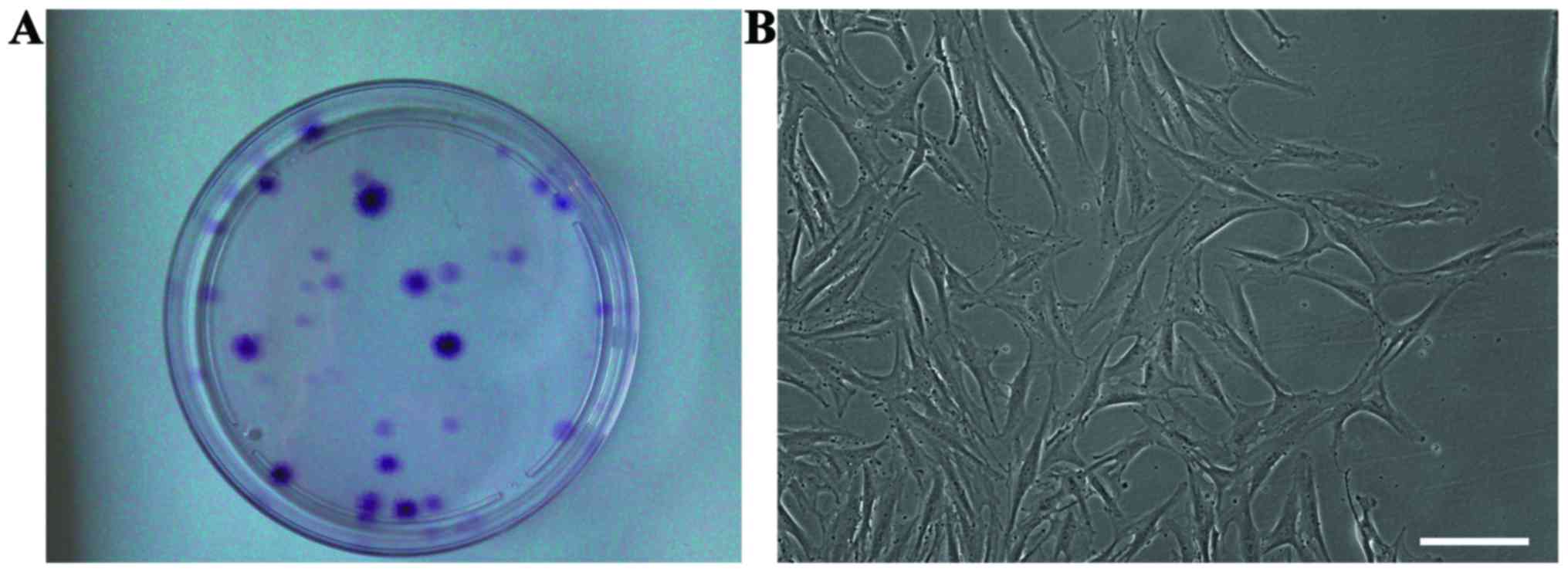
cultured for approximately 2 weeks, cell clones were observed in
culture (Fig. 2A), and the cells
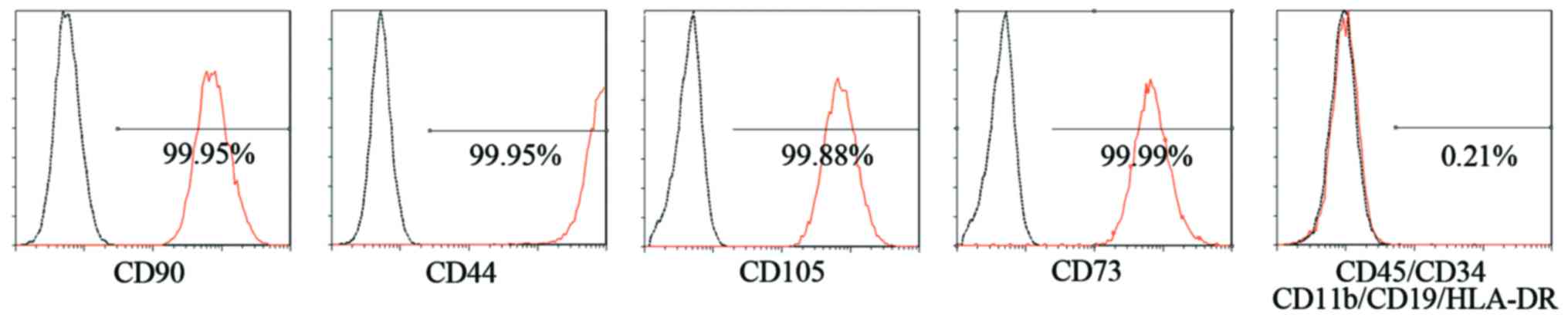
exhibited a typical fibroblastic spindle shape (Fig. 2B). The SFMSCs expressed CD90,
CD105, CD73 and CD44, whereas CD45, CD34, CD11b, CD19 and HLA-DR
were not detected by FACS analysis (Fig. 3).
Differentiation of SFMSCs
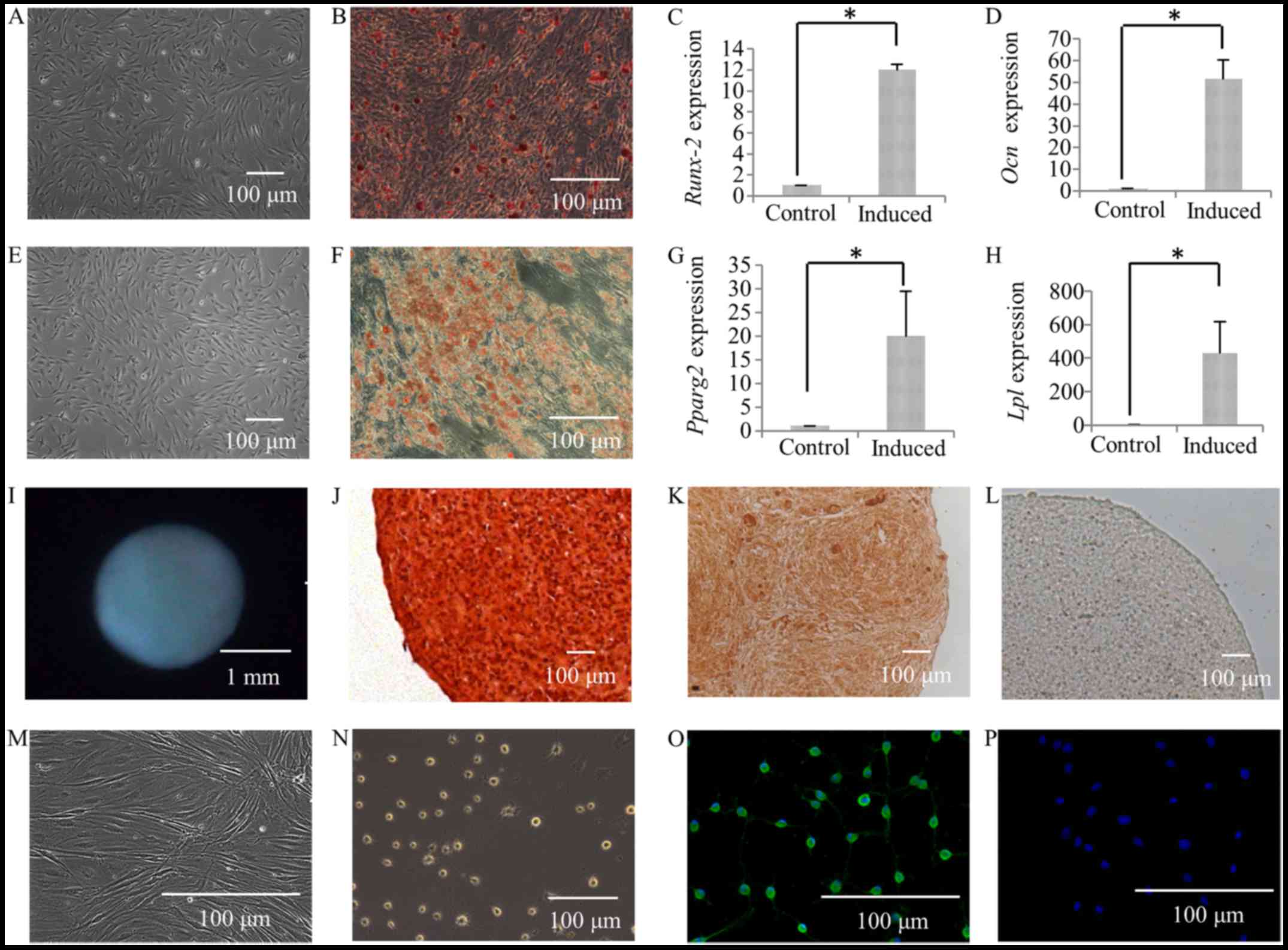
Following culture in osteogenic induction medium for
4 weeks, calcium deposits were observed in the SFMSCs (Fig. 4B), while none were observed in the
control group (Fig. 4A), as shown
by by Alizarin Red staining (Fig. 4A
and B). The expression of the osteogenic transcription factor,
RUNX2, was also found to be higher in the group subjected to
osteogenic differentiation (induced group) compared to the controls
(Fig. 4C). The expression of
OCN, which is a marker for late-stage-matrix-producing
osteoblasts, was significantly increased (Fig. 4D).
Following 28 days of culture in adipogenic induction
medium, the SFMSCs had developed into lipid-laden fat cells that
stained Oil Red O-positive (Fig.
4F), while this was not observed in the control group (Fig. 4E). In addition, PPARG2
expression was significantly increased (Fig. 4G). The gene encoding LPL, a
lipid exchange enzyme that is upregulated during adipogenesis, was
significantly upregulated at day 28 of induction (Fig. 4H).
Following 28 days of culture in chondrogenic
induction medium, the formation of cartilage pellets was evident
(Fig. 4I), and this was not
observed in the controls. The histological sections of the
cartilage pellets were positively stained with Safranin O (Fig. 4J), as also shown in our previous
study (6). In addition, there was
extensive immunoreactivity for collagen type II, a characteristic
collagen found in cartilage (Fig.
4K), whereas no immunostaining was detected in the respective
controls (Fig. 4L).
Following 24 h of culture in neurogenic induction
medium, the cells exhibited a bipolar and stellate morphology
(Fig. 4N), while the cells in the
control group exhibited a typical fibroblastic spindle shape
(Fig. 4M). The expression of the
glia-associated marker, glial fibrillary acidic protein (GFAP, a
specific marker for the astrocyte lineage) in SFMSCs was
demonstrated by immunofluorescence staining (Fig. 4O). No immunostaining was detected
in the respective control cells (Fig.
4P).
Cytotoxicity detection
None of the treatments induced a significant
increase in LDH activity compared to the controls, indicating that
they did not exert cytotoxic effects on the SFMSCs (data not
shown).
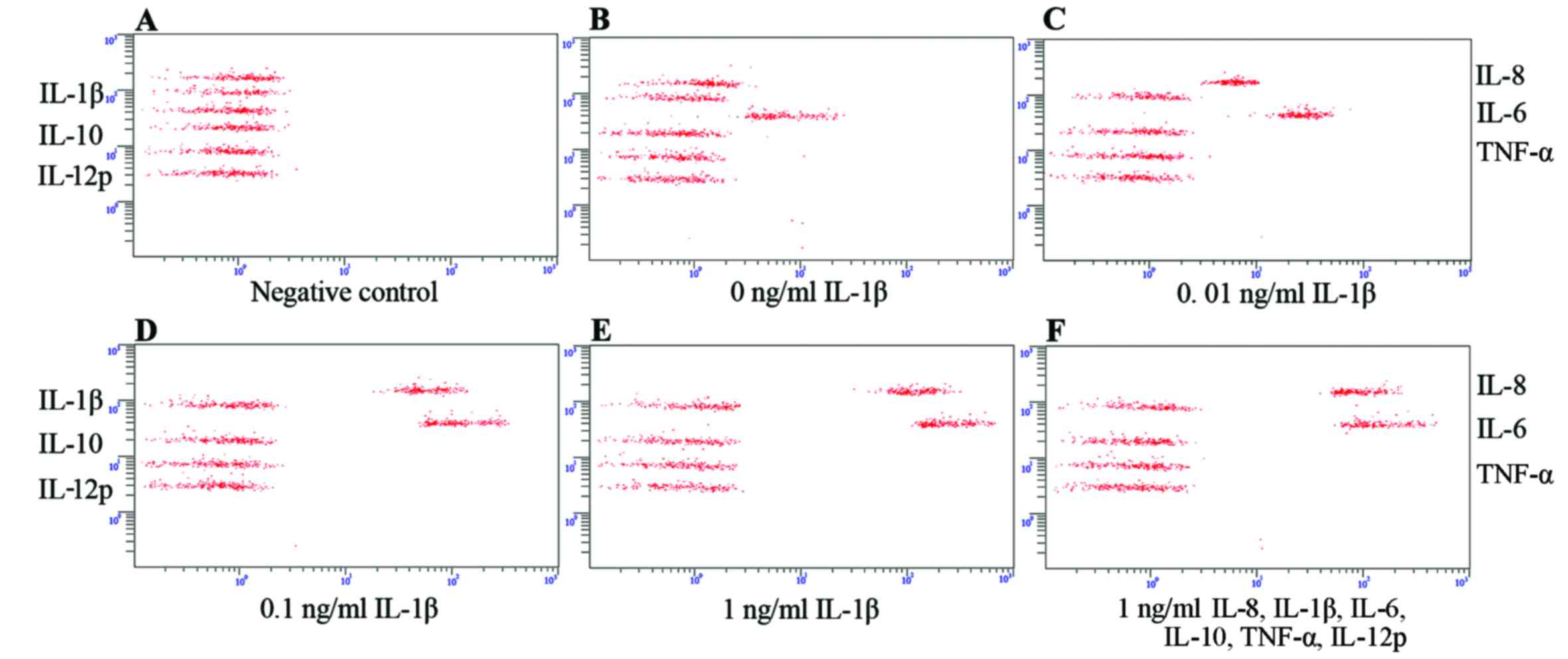
Cytokine secretion from SFMSCs stimulated
with the inflammatory cytokines, IL-8, IL-1β, IL-6, IL-10, TNF-α
and IL-12p
We then determined which cytokines were secreted by
SFMSCs upon exposure to the inflammatory cytokines, IL-8, IL-1β,
IL-6, IL-10, TNF-α and IL-12p. We found that only the IL-8 and IL-6
levels increased significantly following stimulation with IL-1β,
independent of IL-8/IL-1β/IL-6/IL-10/TNF-α/IL-12p compound
stimulation (Fig. 5E and F). This
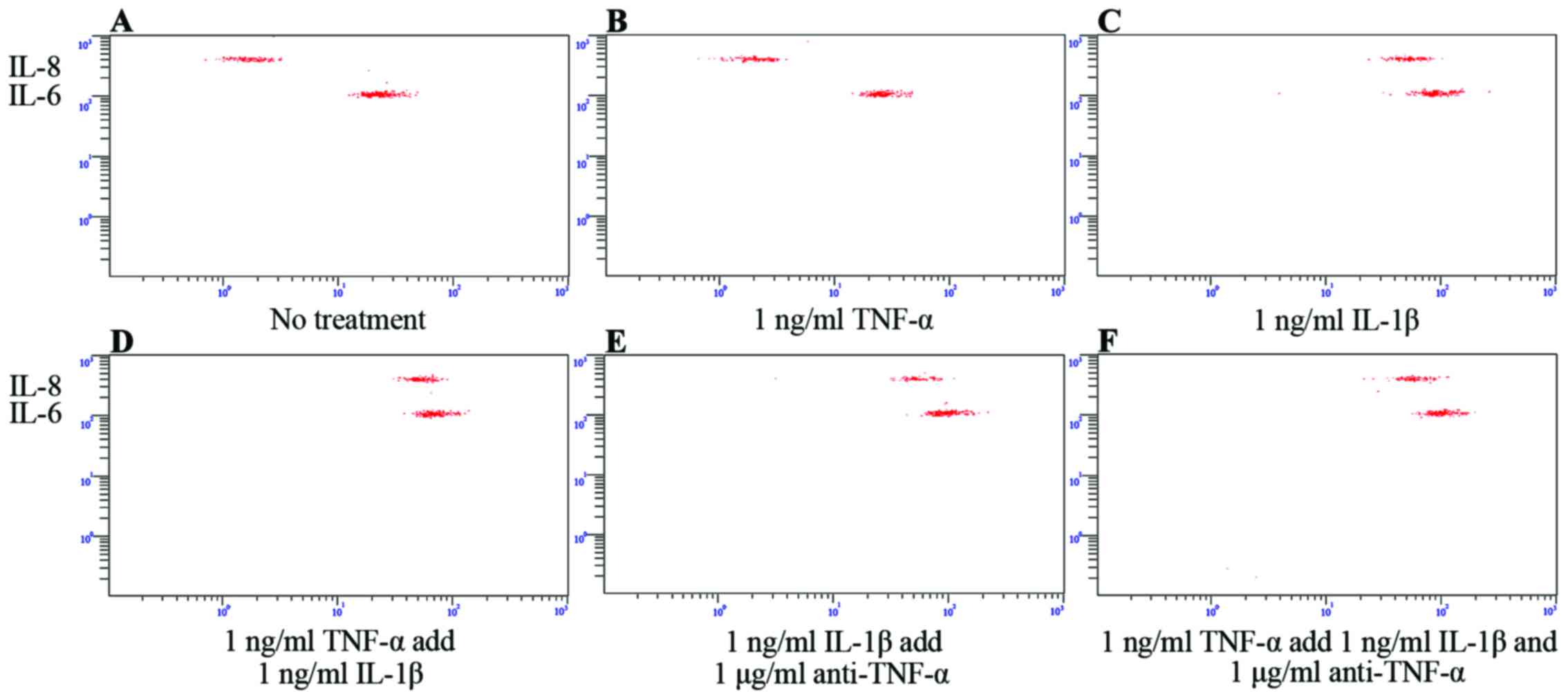
secretion profile was also observed following stimulation with, or
the blockade of, TNF-α signaling (Fig. 6). IL-8 and IL-6 expression was
markedly dependent on the concentration of IL-1β used (Fig. 5A–E).
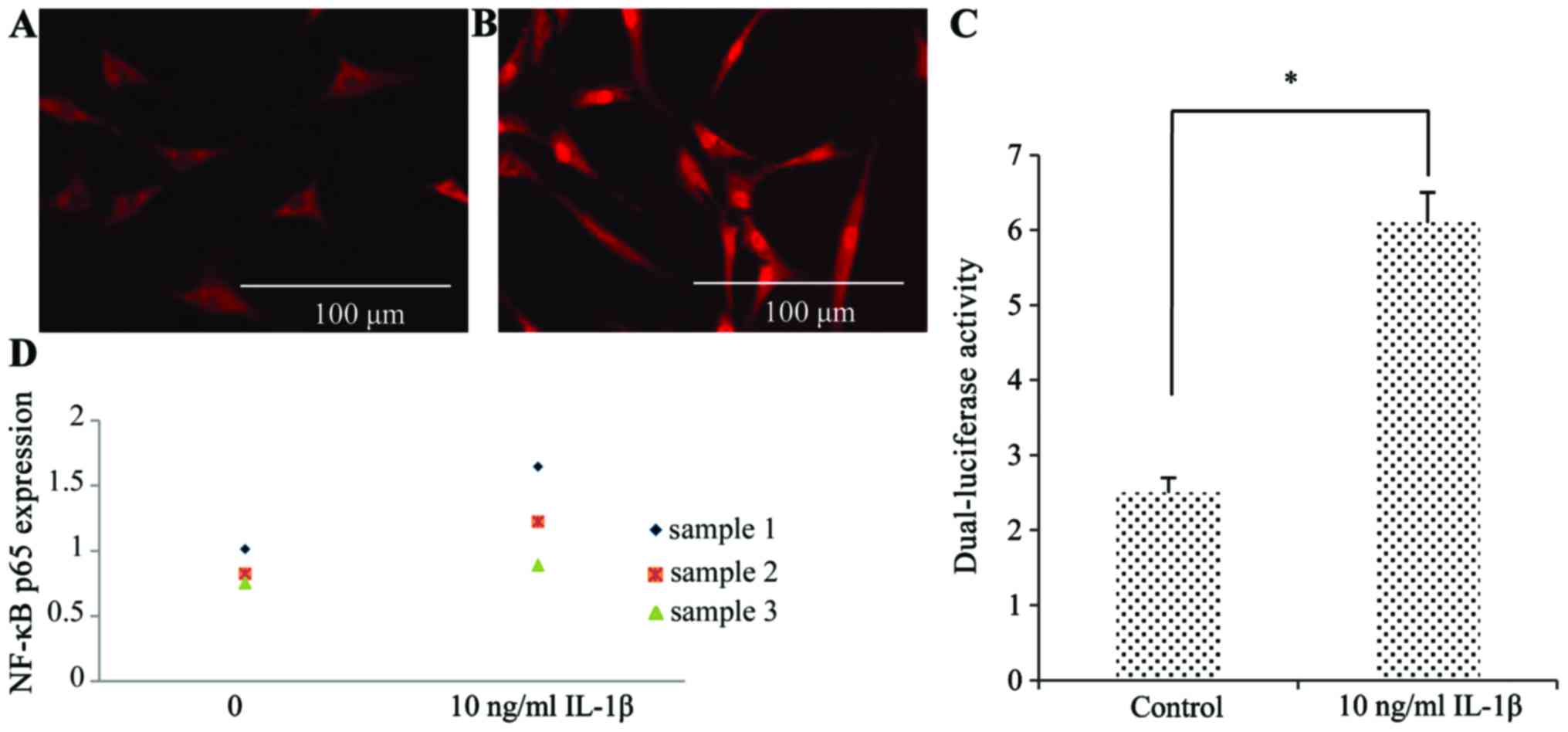
To determine whether NF-κB was responsible for the
effects of IL-1β, we performed immunofluorescence staining using an
antibody to the NF-κB p65 subunit (Fig. 7A and B). Compared to the control
group, the nuclei of the SFMSCs stimulated with IL-1β were strongly
stained, indicating the induction of NF-κB activity. The results of
western blot analysis revealed that NF-κB p65 expression increased
in response to IL-1β (Fig. 7D).
The increased NF-κB activity was also confirmed by dual-luciferase
reporter assays (Fig. 7C).
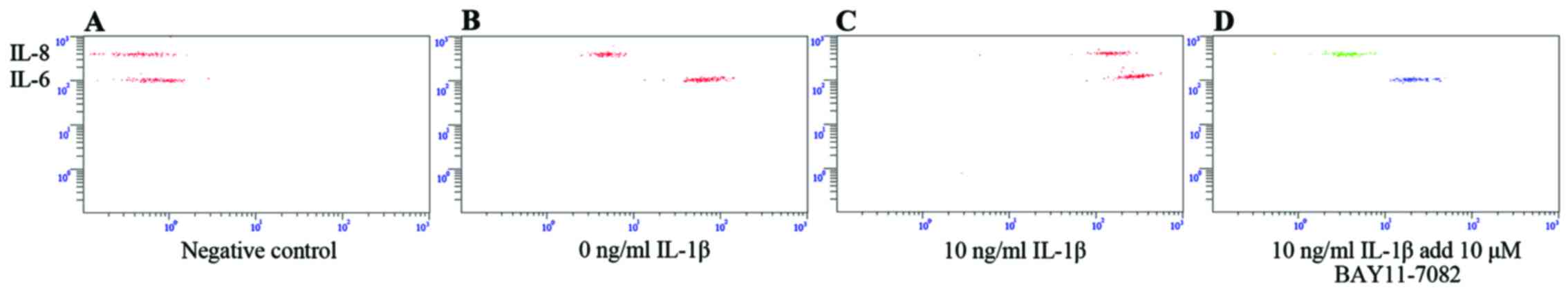
Compared with the untreated control group, the levels of IL-8 and
IL-6 were much higher in the SFMSCs stimulated with 10 ng/ml IL-1β
for 2 h (Fig. 8A–C).
Pre-treatment with NF-κB inhibitor for 1 h significantly attenuated
the IL-1β-dependent induction of IL-8 and IL-6 expression (Fig. 8C and D). Taken together, these
data indicate that the secretion of IL-6 and IL-8 by SFMSCs upon
stimulation with IL-1β in vitro requires the activation of
the NF-κB pathway.
Influence of cytokines on the
chondrogenesis of SFMSCs in vitro
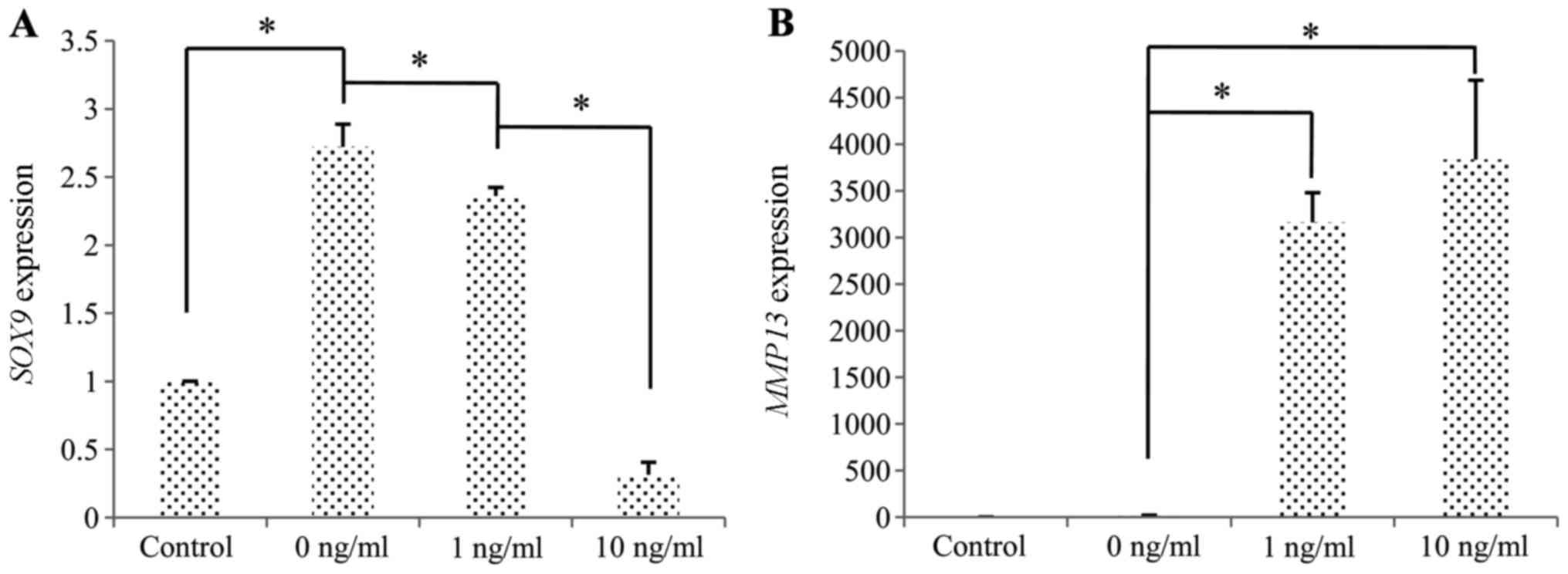
Compared to the untreated cells, the induction of
chondrogenic differentiation in the cells stimulated with IL-1β was
associated with the downregulation of SOX9 and the
upregulation of MMP13 (Fig.
9).
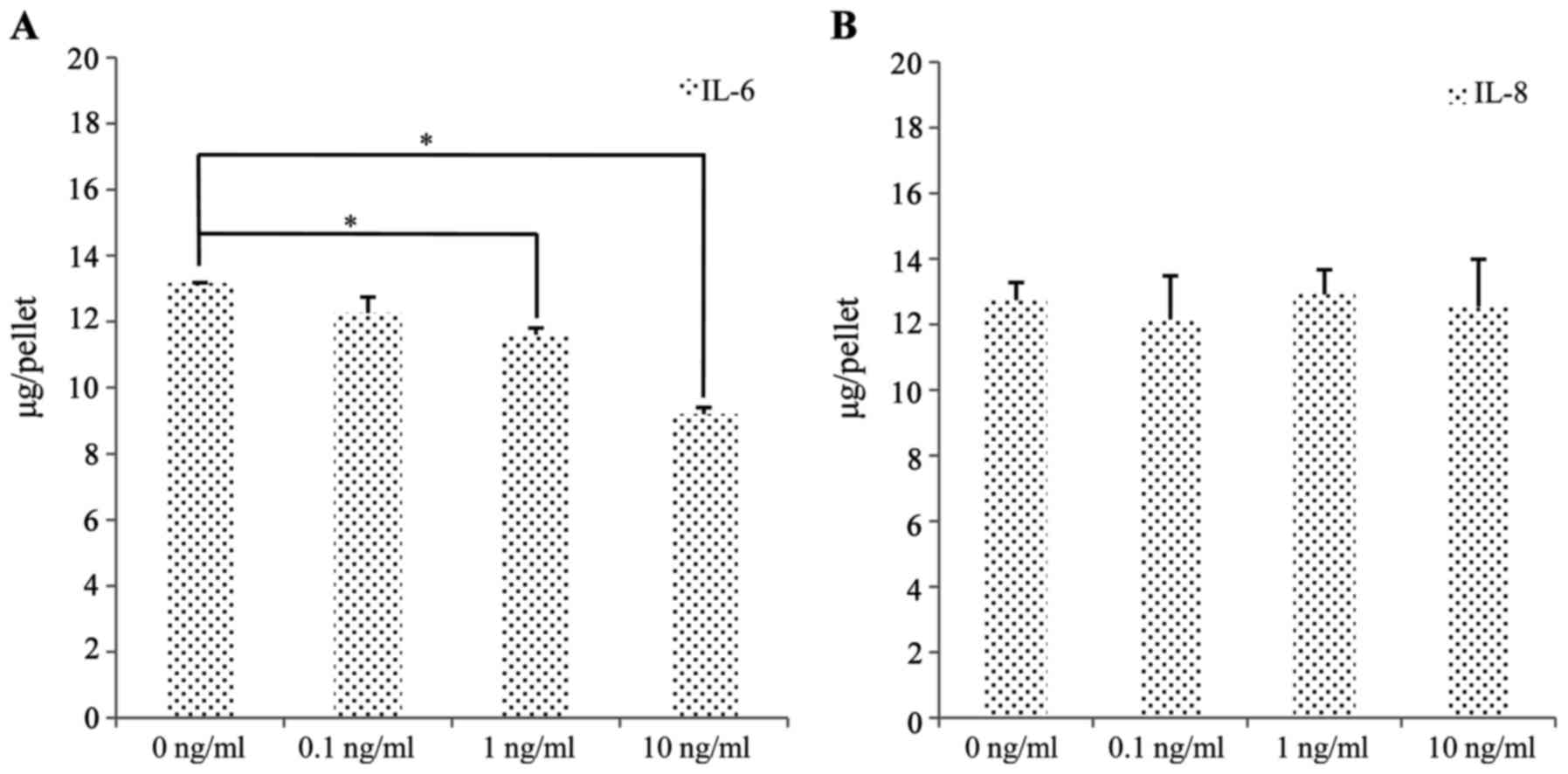
To determine the relative contributions of IL-6 and
IL-8 to the inhibition of chondrogenesis, we exposed the SFMSCs to
IL-6 and IL-8 and quantitatively analyzed the sGAG content. The
sGAG levels of the cartilage pellets exposed to 0, 0.1, 1 and 10
ng/ml IL-6 decreased from 13.17±0.48 to 9.20±0.39 μg/pellet
(300,000 cells/pellet) (Fig.
10A). By contrast, there were no significant changes observed
in sGAG synthesis following exposure to IL-8 (Fig. 10B).
Discussion
MSCs are adult stem cells with notable self-renewal
and multilineage differentiation capacities (24). MSC-based technology is considered
one of the most promising cell-based treatment options for several
diseases, including those associated with tissue injury (25). Various stem cell sources have been
investigated for the repair of damaged tissue. Synovium-derived
MSCs exist in the same microenvironment as articular cartilage and
display the optimal potential for both proliferation and
chondrogenesis when compared with other MSC sources (26–28).
In this study, cells isolated from synovial fluid
samples exhibited a fibroblastic spindle-like shape and expressed
CD90, CD105, CD73 and CD44; they did not express CD45, CD34, CD11b,
CD19 or HLA-DR. These cells also exhibited multilineage potential
and thus match the criteria for human MSCs proposed by the
Mesenchymal and Tissue Stem Cell Committee of the International
Society for Cellular Therapy (ISCT) (24). SFMSCs are a promising cellular
resource for autologous articular cartilage repair, as they are
simple to isolate and culture, and their introduction into the host
does not cause much trauma. Although the existence of SFMSCs has
been documented in some studies (2,4,5),
research into the biological characteristics of SFMSCs in TMD is
limited. However, particular attention should be paid to the way in
which SFMSCs are used in the clinic, as the safety and efficacy of
the procedure are paramount.
Pro-inflammatory cytokines are key contributors to
TMD pathology. Indeed, cytokine regulatory networks are
well-documented features of arthritis. The concentrations of the
pro-inflammatory cytokines, IL-1β, IL-6, IL-8, IL-10, IL-12p and
TNF-α, in patients with TMD are much higher than those in
non-affected individuals; this leads to synovitis and the
production of MMPs. The effects of individual cytokines are
target-tissue-dependent. Fibroblast-like synoviocytes in the joint
are considered major effectors of cartilage destruction, and they
perpetuate inflammation by producing cytokines and degradative
enzymes (29). In this study, we
simulated a pro-inflammatory environment by treating SFMSCs with
IL-1β, IL-6, IL-8, IL-10, IL-12p and TNF-α in vitro.
We observed a robust secretion of IL-6 and IL-8 by
SFMSCs following stimulation with IL-1β in a
concentration-dependent manner, and this was independent of IL-8,
IL-6, IL-10, TNF-α and IL-12p stimulation. IL-8 is one of the most
potent chemoattractants for neutrophils and T lymphocytes (30). IL-6 is important not only in the
orchestration of the overall cytokine network, but also as it plays
a direct role in destruction of the joint (30). Therefore, SFMSCs appear to be an
alternative source of inflammatory cytokines.
Pro-inflammatory cytokines in the inflammatory
milieu can modulate some of the fundamental characteristics of MSCs
(31–33). There are studies on the beneficial
effects of BMSCs on cartilage function in OA (34,35). However, Djouad et al
observed a switch in the behavior of MSCs depending on the
inflammatory environment. Specifically, the injection of MSCs into
the inflamed tissue in a collagen-induced arthritis model had no
beneficial effects on arthritis severity, and in some cases it even
aggravated the collagen-induced arthritis symptoms (36). Augello et al (34) attributed these effects to the
allogeneic C3H10T1/2 cells used by Djouad et al, as it is an
immortalized cell line that may have lost the expression of
immunosuppressors. Liu and Hwang reported that the continuous
exposure of MSCs from cord blood to IL-1β facilitated adipogenic
maturation (37). However,
whether or not SFMSCs in the TMD microenvironment have beneficial
effects on cartilage repair remains unclear.
We further evaluated the chondrogenic
differentiation potential of SFMSCs following stimulation with
IL-1β, a critical cytokine that mediates the activation of
inflammatory and degradative pathways that are associated with TMD.
We found that the expression of SOX9, which is a
chondrogenic marker, was significantly decreased following exposure
to IL-1β. By contrast, we observed an increased expression of the
gene encoding MMP13, the major protease involved in
degradation of the cartilage extracellular matrix, and a factor in
OA progression. These results indicate that IL-1β impedes the
chondrogenic differentiation of SFMSCs in the human TMJ. However,
further studies are required to elucidate the molecular mechanisms
that underlie this process.
Djouad et al reported that MSC-like cells
from the synovial membrane expressed higher levels of transcripts
for macrophage inflammatory protein (MIP2)-α, IL-6 and IL-8
(38). IL-6 and IL-8, which are
produced mainly in response to IL-1β, are significant contributors
to TMJ disorders, such as synovitis, arthralgia and OA (30,39,40). We further demonstrated that IL-6
significantly inhibited the SFMSC-dependent sGAG production that
accompanies the induction of chondrogenesis. By contrast, IL-8 did
not have this effect, indicating that IL-6 is a key mediator of the
IL-1β-dependent inhibition of chondrogenic differentiation.
The NF-κB pathway plays a major role in transducing
inflammatory and catabolic signals in joint degeneration (41). In this study, to further determine
whether NF-κB plays a pivotal role in IL-1β-mediated IL-6
upregulation, we first examined the association between NF-κB
activity and the presence of IL-6 and IL-8. We found that NF-κB
activity in the SFMSCs increased significantly upon exposure to
IL-1β. Furthermore, the IL-1β-dependent expression of IL-6 and IL-8
was reduced to the basal levels when the cells were pre-treated
with an NF-κB inhibitor, confirming that the NF-κB pathway indeed
contributes to the IL-1β-mediated IL-6 upregulation. Thus, the
detailed regulatory mechanisms involved in the NF-κB pathway and
the biological effects of SFMSCs in vivo require further
investigation.
In conclusion, in this study, we found that IL-1β
plays an important role in regulation of the biological behavior of
SFMSCs in an inflammatory milieu. In doing so, IL-1β impedes the
chondrogenic differentiation of SFMSCs. The upregulation of IL-6
and the activation of the NF-κB pathway contribute to this
biological behavior. Our results indicate the potential adverse
effects of IL-1β on the chondrogenic differentiation of SFMSCs,
which may provide new insight into the pathogenesis of TMD. Our
findings also provide an experimental basis for the existence of
MSCs that initiate self-renewal in the synovial fluid.
Acknowledgments
We would like to thank all the nurses and physicians
for their valuable assistance in this study. This study was
supported by grants from the National Science Foundation of China
(81271115), The founders had no role in the study design, data
collection and analysis, decision to publish, or preparation of
this manuscript.
Glossary
Abbreviations
Abbreviations:
|
TMJ
|
temporomandibular joint
|
|
MSCs
|
mesenchymal stem cells
|
|
TMD
|
temporomandibular joint disorders
|
|
SFMSCs
|
synovial fluid-derived mesenchymal
stem cells
|
|
MPC
|
mesenchymal progenitor cell
|
|
OA
|
osteoarthritis
|
|
RA
|
rheumatoid arthritis
|
|
NF-κB
|
nuclear factor-κB
|
|
DI
|
dysfunction index
|
References
|
1
|
Gauer RL and Semidey MJ: Diagnosis and
treatment of temporomandibular disorders. Am Fam Physician.
91:378–386. 2015.PubMed/NCBI
|
|
2
|
Jones EA, English A, Henshaw K, Kinsey SE,
Markham AF, Emery P and McGonagle D: Enumeration and phenotypic
characterization of synovial fluid multipotential mesenchymal
progenitor cells in inflammatory and degenerative arthritis.
Arthritis Rheum. 50:817–827. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Zhang S, Muneta T, Morito T, Mochizuki T
and Sekiya I: Autologous synovial fluid enhances migration of
mesenchymal stem cells from synovium of osteoarthritis patients in
tissue culture system. J Orthop Res. 26:1413–1418. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Harvanová D, Tóthová T, Sarišský M,
Amrichová J and Rosocha J: Isolation and characterization of
synovial mesenchymal stem cells. Folia Biol (Praha). 57:119–124.
2011.
|
|
5
|
Koyama N, Okubo Y, Nakao K, Osawa K,
Fujimura K and Bessho K: Pluripotency of mesenchymal cells derived
from synovial fluid in patients with temporomandibular joint
disorder. Life Sci. 89:741–747. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Sun YP, Zheng YH, Liu WJ, Zheng YL and
Zhang ZG: Synovium fragment-derived cells exhibit characteristics
similar to those of dissociated multipotent cells in synovial fluid
of the temporomandibular joint. PLoS One. 9:e1018962014. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Sullivan C, Murphy JM, Griffin MD, Porter
RM, Evans CH, O'Flatharta C, Shaw G and Barry F: Genetic mismatch
affects the immunosuppressive properties of mesenchymal stem cells
in vitro and their ability to influence the course of
collagen-induced arthritis. Arthritis Res Ther. 14:R1672012.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Papadopoulou A, Yiangou M, Athanasiou E,
Zogas N, Kaloyannidis P, Batsis I, Fassas A, Anagnostopoulos A and
Yannaki E: Mesenchymal stem cells are conditionally therapeutic in
preclinical models of rheumatoid arthritis. Ann Rheum Dis.
71:1733–1740. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Jones EA, Crawford A, English A, Henshaw
K, Mundy J, Corscadden D, Chapman T, Emery P, Hatton P and
McGonagle D: Synovial fluid mesenchymal stem cells in health and
early osteoarthritis: Detection and functional evaluation at the
single-cell level. Arthritis Rheum. 58:1731–1740. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Sekiya I, Ojima M, Suzuki S, Yamaga M,
Horie M, Koga H, Tsuji K, Miyaguchi K, Ogishima S, Tanaka H and
Muneta T: Human mesenchymal stem cells in synovial fluid increase
in the knee with degenerated cartilage and osteoarthritis. J Orthop
Res. 30:943–949. 2012. View Article : Google Scholar
|
|
11
|
Lee DH, Sonn CH, Han SB, Oh Y, Lee KM and
Lee SH: Synovial fluid CD34− CD44+
CD90+ mesenchymal stem cell levels are associated with
the severity of primary knee osteoarthritis. Osteoarthritis
Cartilage. 20:106–109. 2012. View Article : Google Scholar
|
|
12
|
Neidhart M, Seemayer CA, Hummel KM, Michel
BA, Gay RE and Gay S: Functional characterization of adherent
synovial fluid cells in rheumatoid arthritis: Destructive potential
in vitro and in vivo. Arthritis Rheum. 48:1873–1880. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Heldens GT, Blaney Davidson EN, Vitters
EL, Schreurs BW, Piek E, van den Berg WB and van der Kraan PM:
Catabolic factors and osteoarthritis-conditioned medium inhibit
chondrogenesis of human mesenchymal stem cells. Tissue Eng Part A.
18:45–54. 2012. View Article : Google Scholar
|
|
14
|
Boeuf S, Graf F, Fischer J, Moradi B,
Little CB and Richter W: Regulation of aggrecanases from the ADAMTS
family and aggrecan neoepitope formation during in vitro
chondrogenesis of human mesenchymal stem cells. Eur Cell Mater.
23:320–332. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Li J, Long X, Ke J, Meng QG, Lee WC,
Doocey JM and Zhu F: Regulation of HAS expression in human synovial
lining cells of TMJ by IL-1beta. Arch Oral Biol. 53:60–65. 2008.
View Article : Google Scholar
|
|
16
|
Alstergren P, Ernberg M, Kvarnström M and
Kopp S: Interleukin-1beta in synovial fluid from the arthritic
temporomandibular joint and its relation to pain, mobility, and
anterior open bite. J Oral Maxillofac Surg. 56:1059–1066. 1998.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Kristensen KD, Alstergren P, Stoustrup P,
Küseler A, Herlin T and Pedersen TK: Cytokines in healthy
temporomandibular joint synovial fluid. J Oral Rehabil. 41:250–256.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Marks PH and Donaldson ML: Inflammatory
cytokine profiles associated with chondral damage in the anterior
cruciate ligament-deficient knee. Arthroscopy. 21:1342–1347. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Kobayashi M, Squires GR, Mousa A, Tanzer
M, Zukor DJ, Antoniou J, Feige U and Poole AR: Role of
interleukin-1 and tumor necrosis factor alpha in matrix degradation
of human osteoarthritic cartilage. Arthritis Rheum. 52:128–135.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Fan Z, Söder S, Oehler S, Fundel K and
Aigner T: Activation of interleukin-1 signaling cascades in normal
and osteoarthritic articular cartilage. Am J Pathol. 171:938–946.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Goldring MB, Otero M, Plumb DA, Dragomir
C, Favero M, El Hachem K, Hashimoto K, Roach HI, Olivotto E, Borzì
RM, et al: Roles of inflammatory and anabolic cytokines in
cartilage metabolism: Signals and multiple effectors converge upon
MMP-13 regulation in osteoarthritis. Eur Cell Mater. 21:202–220.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Tang J, Cui W, Song F, Zhai C, Hu H, Zuo Q
and Fan W: Effects of mesenchymal stem cells on
interleukin-1β-treated chon-drocytes and cartilage in a rat
osteoarthritic model. Mol Med Rep. 12:1753–1760. 2015.PubMed/NCBI
|
|
23
|
Fricton JR and Schiffman EL: Reliability
of a craniomandibular index. J Dent Res. 65:1359–1364. 1986.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Dominici M, Le Blanc K, Mueller I,
Slaper-Cortenbach I, Marini F, Krause D, Deans R, Keating A,
Prockop DJ and Horwitz E: Minimal criteria for defining multipotent
mesenchymal stromal cells. The International Society for Cellular
Therapy position statement. Cytotherapy. 8:315–317. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Keating A: Mesenchymal stromal cells: New
directions. Cell Stem Cell. 10:709–716. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Yoshimura H, Muneta T, Nimura A, Yokoyama
A, Koga H and Sekiya I: Comparison of rat mesenchymal stem cells
derived from bone marrow, synovium, periosteum, adipose tissue, and
muscle. Cell Tissue Res. 327:449–462. 2007. View Article : Google Scholar
|
|
27
|
Ogata Y, Mabuchi Y, Yoshida M, Suto EG,
Suzuki N, Muneta T, Sekiya I and Akazawa C: Purified human synovium
mesenchymal stem cells as a good resource for cartilage
regeneration. PLoS One. 10:e01290962015. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Suzuki S, Muneta T, Tsuji K, Ichinose S,
Makino H, Umezawa A and Sekiya I: Properties and usefulness of
aggregates of synovial mesenchymal stem cells as a source for
cartilage regeneration. Arthritis Res Ther. 14:R1362012. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Ritchlin C: Fibroblast biology. Effector
signals released by the synovial fibroblast in arthritis. Arthritis
Res. 2:356–360. 2000. View
Article : Google Scholar : PubMed/NCBI
|
|
30
|
Kaneyama K, Segami N, Nishimura M, Suzuki
T and Sato J: Importance of proinflammatory cytokines in synovial
fluid from 121 joints with temporomandibular disorders. Br J Oral
Maxillofac Surg. 40:418–423. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Guttridge DC, Mayo MW, Madrid LV, Wang CY
and Baldwin AS Jr: NF-kappaB-induced loss of MyoD messenger RNA:
Possible role in muscle decay and cachexia. Science. 289:2363–2366.
2000. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Li X and Makarov SS: An essential role of
NF-kappaB in the 'tumor-like' phenotype of arthritic synoviocytes.
Proc Natl Acad Sci USA. 103:17432–17437. 2006. View Article : Google Scholar
|
|
33
|
Sitcheran R, Cogswell PC and Baldwin AS
Jr: NF-kappaB mediates inhibition of mesenchymal cell
differentiation through a posttranscriptional gene silencing
mechanism. Genes Dev. 17:2368–2373. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Augello A, Tasso R, Negrini SM, Cancedda R
and Pennesi G: Cell therapy using allogeneic bone marrow
mesenchymal stem cells prevents tissue damage in collagen-induced
arthritis. Arthritis Rheum. 56:1175–1186. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
van Buul GM, Villafuertes E, Bos PK,
Waarsing JH, Kops N, Narcisi R, Weinans H, Verhaar JA, Bernsen MR
and van Osch GJ: Mesenchymal stem cells secrete factors that
inhibit inflammatory processes in short-term osteoarthritic
synovium and cartilage explant culture. Osteoarthritis Cartilage.
20:1186–1196. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Djouad F, Fritz V, Apparailly F,
Louis-Plence P, Bony C, Sany J, Jorgensen C and Noël D: Reversal of
the immunosuppressive properties of mesenchymal stem cells by tumor
necrosis factor alpha in collagen-induced arthritis. Arthritis
Rheum. 52:1595–1603. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Liu CH and Hwang SM: Cytokine interactions
in mesenchymal stem cells from cord blood. Cytokine. 32:270–279.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Djouad F, Bony C, Häupl T, Uzé G, Lahlou
N, Louis-Plence P, Apparailly F, Canovas F, Rème T, Sany J, et al:
Transcriptional profiles discriminate bone marrow-derived and
synovium-derived mesenchymal stem cells. Arthritis Res Ther.
7:R1304–R1315. 2005. View
Article : Google Scholar : PubMed/NCBI
|
|
39
|
Nishimura M, Segami N, Kaneyama K, Suzuki
T and Miyamaru M: Proinflammatory cytokines and arthroscopic
findings of patients with internal derangement and osteoarthritis
of the temporomandibular joint. Br J Oral Maxillofac Surg.
40:68–71. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Fu K, Ma X, Zhang Z, Pang X and Chen W:
Interleukin-6 in synovial fluid and HLA-DR expression in synovium
from patients with temporomandibular disorders. J Orofac Pain.
9:131–137. 1995.PubMed/NCBI
|
|
41
|
Roman-Blas JA and Jimenez SA: NF-kappaB as
a potential therapeutic target in osteoarthritis and rheumatoid
arthritis. Osteoarthritis Cartilage. 14:839–848. 2006. View Article : Google Scholar : PubMed/NCBI
|