Introduction
Colorectal cancer (CRC), one of the most common
malignancies worldwide, is regarded as the second cause of
cancer-related deaths each year (1). The incidence of CRC has rapidly
increased in China among both men and women (2). Although advances in treatment
strategy and extensive investigation of CRC have been accomplished
in the past decade, the mortality rate remains high due to the
metastasis-associated poor prognosis (3). Therefore, in order to improve the
prognosis and reduce the rate of recurrence of CRC patients, the
search for novel potential biomarkers and efficient targets for CRC
treatment is a crucial goal.
MicroRNAs (miRNAs or miRs), small non-coding RNA
molecules approximately 18–25 nucleotides in length, are highly
conserved in most eukaryotic organisms (4). Through binding to the
3′-untranslated regions (3′-UTRs) of their corresponding messenger
RNA (mRNA) targets, miRNAs degrade or inhibit the
posttranscriptional regulation of their (mRNA) targets (5). miRNAs have been reported to be
involved in multiple physiological and pathological processes
including human malignancies (6,7).
Many studies have confirmed that miRNAs can be identified as
oncogenes or tumor suppressors in the initiation and development of
cancers by targeting corresponding abnormally expressed proteins
(8). Consequently, miRNAs have
been proposed to be potential indicators and therapeutic targets in
various types of cancers (9,10).
miR-30d is a member of the miR-30 family that is
responsible for tumor development and progression. Recently,
accumulating evidence has shown that miR-30d plays a role as either
a tumor suppressor or an oncogene in the progression of different
tumor types (11). Zhang et
al found that dysregulation of miR-30d in human anaplastic
thyroid carcinoma resulted in the insensitivity to chemotherapeutic
drugs by enhancing autophagic survival (12). Xuan et al demonstrated that
down-regulation of miR-30d resulted in the pathogenesis of prostate
cancer by targeting B-cell specific moloney leukemia virus
insertion region homologue-1 (Bmi-1) (13). Ye et al indicated that
overexpression of miR-30d blocked transforming growth factor-β
(TGF-β1)-induced epithelial-mesenchymal transition (EMT) by
targeting Snail in ovarian cancer cells (14). However, the exact role and
molecular mechanisms underlying the regulation of the progression
of CRC tumorgenesis by miR-30d remains largely unknown.
Liver receptor homologue-1 (LRH-1), also known as
NR5A2, is a member of the nuclear receptor (NR) subfamily (15). This NR participates in a variety
of biological processes, such as differentiation and development,
reverse cholesterol transport, bile-acid homeostasis and
steroidogenesis (16,17). Evidence indicates that LRH-1 is
responsible for the pathogenesis of multiple types of tumors
(18). Recent studies have
demonstrated high LRH-1 expression in pancreatic cancer cells and
have found that LRH-1 overexpression enhances cell migration and
invasion abilities (19). In
addition, LRH-1 contributes to intestinal tumor proliferation in
gastrointestinal tumors by activating the Wnt/β-catenin pathway
(20,21). These findings indicate that LRH-1
can be involved in the development of cancers.
In the present study, we found that miR-30d was
down-regulated in CRC tissue samples and a decreased expression
level of miR-30d was closely correlated to clinicopathological
parameters including poor prognosis, the degree of differentiation,
invasive depth, TNM stage, distant metastasis, and lymph node
metastasis. LRH-1 was identified as a direct target of miR-30d as
detected by the LRH-1 3′-UTR region and luciferase report assay. In
addition, ectopic expression of miR-30d inhibited CRC cell
proliferation and invasion, as well as the Wnt/β-catenin signaling
pathway by suppressing the expression of LRH-1. Therefore, the
present study indicates that miR-30d may be a potential prognostic
marker for CRC.
Materials and methods
Clinical sample collection
Fresh CRC tissue samples and paired adjacent normal
samples were obtained from 80 patients who underwent routine
surgery at the Department of General Surgery, Shaanxi Province
People's Hospital (Xi'an, China) from March 2005 to July 2010.
Patients who underwent radiotherapy or chemotherapy prior to
surgery were excluded. All patients involved were divided into two
groups according to the median RNA level of miR-30d (high and low).
A five-year follow-up was performed to record the difference in
prognosis between the low and high miR-30d groups. Informed consent
was provided by all patients, and the present study was approved by
the Human Ethics Committee of Shaanxi Provincial People's Hospital
(Xi'an, China) and the 1964 Helsinki Declaration.
Cell culture
The human CRC cell lines (LoVo, HT-29, RKO, HCT8 and
SW480), the normal colorectal mucosa cell line FHC, and the 293T
cell line were purchased from the Cell Center of the Chinese
Academy of Sciences (Shanghai, China). All cells were cultured in
Dulbecco's modified Eagle's medium (DMEM) supplemented with 10%
fetal calf serum (FBS) (Gibco BRL, Gaithersburg, MD, USA) and 1%
penicillin/streptomycin in a humidified incubator containing 5%
CO2 at 37°C.
Cell transfection
The miR-30d mimic (miR-30d) and miR-30d mimic NC
(miR-NC) were purchased from GenePharma Company (Shanghai, China).
LoVo and SW480 cells were selected and seeded into 6-well plates at
the density of 3×105 cells/well and transfected with 100
nM oligonucleotides using Lipofectamine 2000 reagent (Invitrogen
Life Technologies, Carlsbad, CA, USA) according to the
manufacturer's protocol.
Isolation of total RNA and quantitative
RT-PCR
Total RNA or miRNA from the CRC tumor samples and
cell lines was extracted using the RNeasy Mini kit or miRNeasy Mini
kit (Qiagen, Manchester, UK) according to the manufacturer's
instructions. cDNA was synthesized from 10 µg RNA using
M-MLV reverse transcriptase (Clontech Laboratories, Mountain View,
CA, USA) or miScript reverse transcription kit (Qiagen). Real-time
polymerase chain reaction (PCR) was performed with SYBR Premix Ex
Taq (Takara Bio, Dalian, China) to examine the mRNA or miRNA level.
GAPDH and U6 were used for normalization. The primers for LRH-1 and
GAPDH were designed as follows: LRH-1 forward,
5′-CCCAGCATTTTAACACTACCGA-3′ and reverse
5′-GGGCAGGGGAAAGACAACTAA-3′; GAPDH forward,
5′-AAGCGTTGTCCTTACTGTCGT-3′ and reverse
5′-GTCGGTAGTGTTAAAATGCTGGG-3′. ABI 7900 Fast Real-time PCR system
(ABI, USA) was used to perform RT-qPCR. The 2−ΔΔCt
method was used to analyze the relative gene expression level.
Western blot analysis
CRC tissue samples and cells were washed with
ice-cold phosphate-buffered saline (PBS) 3 times and lysed in RIPA
buffer supplemented with protease inhibitor. The protein
concentration was determined using the bicinchoninic acid method.
Equal amounts of proteins (30 µg) were separated by 15%
SDS-PAGE and transferred to a PVDF membrane (Immobilon; Millipore
Corp., Bedford, MA, USA). After blocking with 5% non-fat milk
(diluted with phosphate-buffered saline/Tween-20, TBST) at 37°C for
1 h, the membranes were incubated with primary antibodies:
anti-LRH-1 (1:1000, ab41901), anti-matrix metalloproteinase (MMP)2
(1:1,000, ab80737), anti-MMP9 (1:1,000, ab119906), anti-β-catenin
(1:500, ab92514), cyclin D1 (1:1,000, ab134175), c-Myc (1:500,
ab51156) and GAPDH (1:1,000, ab8245; Abcam, Cambridge, UK) at 4°C
overnight. After rinsing with TBST, they were incubated with
horseradish peroxidase-conjugated goat anti-mouse or rabbit
immunoglobulin G antibodies (1:200; Santa Cruz Biotechnology, Inc.,
Santa Cruz, CA). Protein bands were detected using an enhanced
chemiluminescence reagent (Pierce Biotechnology, Inc., Rockford,
IL, USA). Band intensity was quantified using Image software (Media
Cybernetics, Inc., Rockville, MD, USA).
Cell proliferation assay
3-(4,5-Dimethylthiazol-2-yl)-2,
5-diphenyltetrazolium bromide (MTT) assay was used to assess the
CRC cell proliferation. Firstly, CRC cells were seeded into a
96-well plate at a density of 2×103 cells/well and
transfected with miR-30d or miR-NC for 96 h. Fresh medium with 20
µl MTT (5 mg/ml) was added into each well to replace the old
medium and incubation was carried out for another 4 h at 37°C. Then
the medium was removed and 150 µl DMSO was added into each
well. The optical density was recorded at a wavelength of 490 nm
using a microplate reader (Multiskan FC; Thermofisher,
Finland).
Cell cycle analysis
Cell cycle distribution was analyzed by flow
cytometry as previously reported (22). Briefly, cells trans-fected with
miR-30d or miR-NC for 48 h were resuspended and fixed with 70%
ice-cold ethanol at -20°C overnight. Afterwards, the cells were
washed with PBS and stained with propidium iodide (PI)/RNase
fluorescent probe solution (Molecular Probes, Eugene, OR, USA) for
30 min in the dark. DNA contents were assessed with a FACSCalibur
flow cytometer (Becton-Dickinson, San Diego, CA, USA).
Apoptosis assay
Apoptosis was measured using the Annexin V-FITC/PI
apoptosis kit (Abcam, Cambridge, UK) according to the
manufacturer's protocol (22).
Cells were collected after transfection with miR-30d or miR-NC for
48 h, washed twice with cold PBS, and resuspended at
1×105 cells/ml and mixed with a binding buffer
containing Annexin V-FITC and PI. After incubation for 15 min in
the dark, the apoptotic cells were analyzed using a FACSCalibur
flow cytometry.
Migration and invasion assays
Cells were pretreated with miR-30d or miR-NC for 48
h. For the migration assay, 1×104 cells in serum-free
medium were added to the upper chamber of each transwell (Millipore
Corp.) with a non-coated membrane. For the invasion assay,
5×104 cells were added on the upper chamber of each
insert coated with Matrigel (Becton-Dickinson). Medium (500
µl) containing 10% FBS as the chemotactic factor was added
to the lower chambers. After 24 h of incubation for the migration
assay and 48 h of incubation for the invasion assay at 37°C, the
cells remaining on the upper membrane of the filter were carefully
removed with cotton swabs. Cells that had translocated to the lower
chambers were fixed with methanol and stained using 0.1% crystal
violet. The migratory or invasive cells were evaluated and imaged
by an IX71 inverted microscope (Olympus, Tokyo, Japan).
miRNA target prediction and
dual-luciferase reporter assay
The potential targets of miR-30d were predicted
using three online prediction programs: microRNA.org (http://www.microrna.org/microrna/home.do), PicTar
(http://www.pictar.org/cgi-bin/PicTar_vertebrate.cgi)
and TargetScan (http://www.targetscan.org/vert_60/). Based on the
prediction results, the 3′-UTR of LRH-1 containing the miR-30d
putative binding site and mutant site were designed and produced by
GenePharma (Shanghai, China), and subcloned into the pGL3
luciferase vector (Promega Corp., Madison, WI, USA) to construct a
3′-UTR-LRH1-wild-type and mutant 3′-UTR-LRH1. Human 293T cells were
seeded into a 24-well plate and co-transfected with miR-30d or
miR-NC and LRH-1 WT-3′-UTR or the mutant 3′-UTR vector for 48 h.
The cells were then harvested and luciferase activity was assessed
using the dual-luciferase reporter assay system (Promega, USA).
Renilla luciferase activity was used to normalize the luciferase
activity.
Xenograft experiment
Four-week-old BALB/c male nude mice were obtained
from the Animal Center of the Fourth Military Medical University
(Xi'an, China) and divided into three groups (control, miR-30d and
miR-NC, n=10 in each group). SW480 cells with miR-30d or miR-NC
transfection were suspended in PBS and subcutaneously injected into
each mouse at a total of 5×106 cells. Tumor growth was
monitored every week after tumor cell injection. After 7 weeks, all
animals were euthanized by cervical dislocation and xenograft tumor
volumes were measured using the formula: Volume = 1/2 (length ×
weight)2. All the animal experimental procedures were
approved by the Animal Care and Use Committee of Shaanxi Province
People's Hospital (Xi'an, China).
Statistical analysis
All statistical analyses were performed using SPSS
13.0 software. The data are presented as the mean ± standard
deviation (SD). Student's t-test was used for comparisons between
two groups, whereas more than two groups were compared by one-way
analysis of variance followed by Bonferroni test. Pearson
correlation was used to identify the correlation between the
expression of miR-30d and LRH-1 (R=correlation coefficient).
P<0.05 was considered to indicate a statistically significant
difference. All experiments were repeated at least in
triplicate.
Results
miR-30d is downregulated and LRH-1 is
upregulated in human CRC tissues
First, the expression level of miR-30d was detected
in CRC tissue samples and paired adjacent tissues (n=80) using
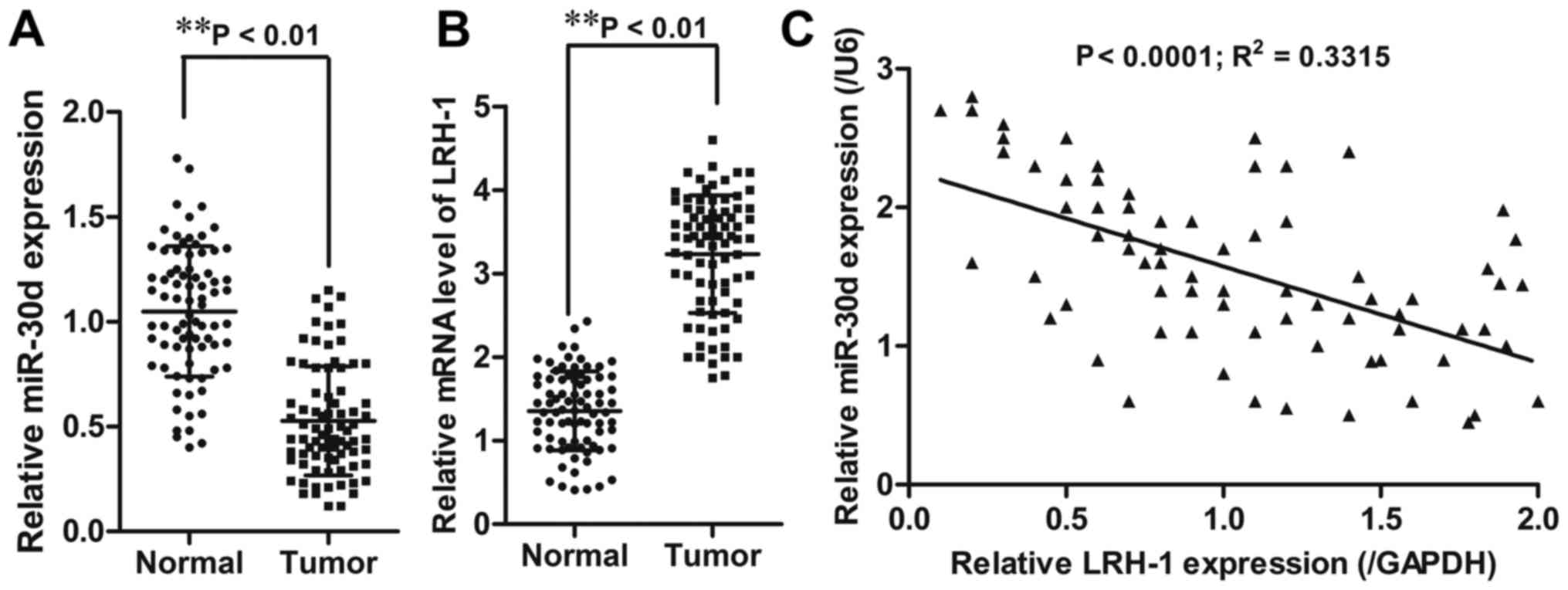
RT-qPCR. The results showed that CRC tissues had significantly
reduced miR-30d mRNA expression (P<0.01) and increased LRH-1
mRNA expression (P<0.01) when compared to the adjacent normal
tissues (Fig. 1A and B).
Pearson's correlation analyses showed that miR-30d expression was
inversely correlated with LRH-1 expression (R2=0.3315;
P<0.0001) in the CRC tissue samples (Fig. 1C).
Downregulation of miR-30d and
upregulation of LRH-1 is associated with aggressiveness of CRC
progression
To further verify the significance of aberrant
expression of miR-30d and LRH-1 in CRC tissue specimens,
clinicopathological analysis was performed accordingly. Based on
the median levels (0.527, miR-30d; 3.232, LRH-1) in CRC tissue
specimens, all 80 CRC patients were subdivided into a low miR-30d
expression group (<0.527, 43 cases), a high miR-30d expression
group (>0.527, 37 cases) as well as a high LRH-1 expression
group (>3.232, 51 cases) and a low LRH-1 expression group
(<3.232, 29 cases). The correlation between miR-30d expression
and the clinicopathological factors of the patients was determined.
As shown in Table I, the
clinicopathological analysis illustrated that downregulation of
miR-30d was significantly associated with distant metastasis
(P<0.05), differentiation (P<0.05), invasive depth
(P<0.05), TNM stage (P<0.05), and lymph node metastasis
(P<0.05); whereas, upregulation of LRH-1 was markedly correlated
with distant metastasis (P<0.05), TNM stage (P<0.05),
invasive depth (P<0.05), and lymph node metastasis (P<0.05),
but had no correlation with age, gender or tumor size (P>0.05).
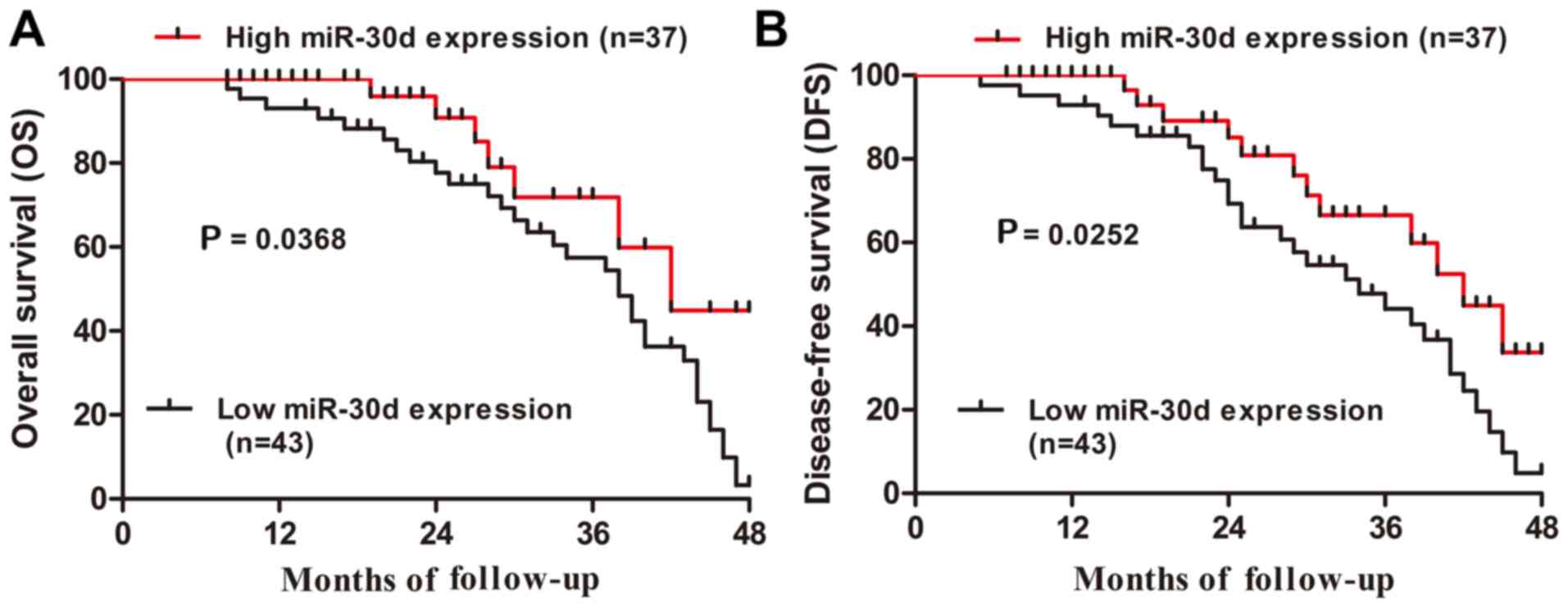
Furthermore, Kaplan-Meier analysis revealed that CRC patients with
low miR-30d expression had markedly shorter overall survival and
disease-free survival rates than those in the high miR-30d
expression group (P=0.0368 and P=0.0252, respectively; Fig. 2A and B). Thus, these data
indicated that downregulation of miR-30d may be involved in the
development of CRC.
 | Table ICorrelation between miR-30d
expression and clinicopathologic parameters in the patients with
CRC. |
Table I
Correlation between miR-30d
expression and clinicopathologic parameters in the patients with
CRC.
| Parameters | No. of cases | miR-30d expression
| P-value | LRH-1 expression
| P-value |
|---|
| High (37) | Low (43) | Low (29) | High (51) |
|---|
| Age (years) | | | | 0.071 | | | 0.169 |
| >60 | 44 | 16 | 28 | | 19 | 25 | |
| ≤60 | 36 | 21 | 15 | | 10 | 26 | |
| Gender | | | | 0.262 | | | 0.486 |
| Male | 41 | 16 | 25 | | 13 | 28 | |
| Female | 39 | 21 | 18 | | 16 | 23 | |
| Distant
metastasis | | | | <0.05a | | | <0.05a |
| M0 | 46 | 30 | 16 | | 25 | 21 | |
| M1 | 34 | 14 | 20 | | 10 | 24 | |
| TNM stage | | | | <0.01b | | | <0.05a |
| I–II | 34 | 22 | 12 | | 7 | 27 | |
| III–IV | 46 | 15 | 31 | | 22 | 24 | |
| Degree of
differentiation | | | | <0.05a | | | 0.345 |
| Well/moderate | 27 | 17 | 10 | | 14 | 13 | |
| Poor | 53 | 20 | 33 | | 21 | 32 | |
| Invasive depth | | | | <0.05a | | | <0.05a |
| T1-T2 | 32 | 20 | 12 | | 17 | 15 | |
| T3-T4 | 48 | 17 | 31 | | 12 | 36 | |
| Lymph node
metastasis | | | | <0.01b | | | <0.05a |
| Yes | 36 | 25 | 11 | | 18 | 18 | |
| No | 44 | 12 | 32 | | 11 | 33 | |
| Tumor size
(cm) | | | | 0.262 | | | 0.647 |
| >5 | 41 | 19 | 22 | | 16 | 25 | |
| ≤5 | 39 | 13 | 26 | | 13 | 26 | |
Overexpression of miR-30d suppresses CRC
cell proliferation and invasion
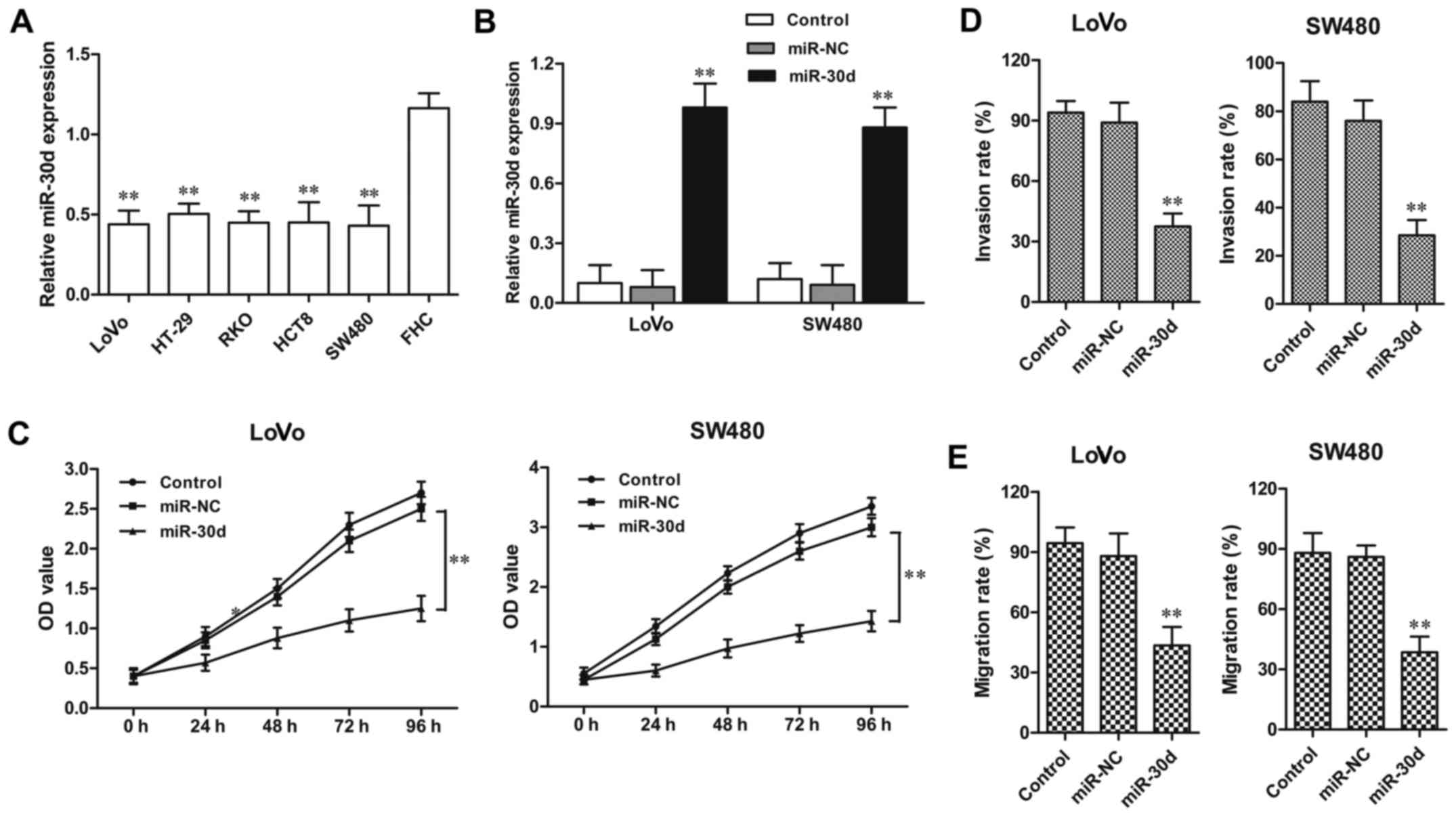
The miR-30d expression level was detected in four
CRC cell lines. Consistent with the tissue sample results, the
miR-30d expression level was also distinctly downregulated in the
CRC HT29, LoVo, RKO, HCT8, SW480 cell lines when compared to the
level in the normal colorectal cell line FHC (Fig. 3A). Then LoVo and SW480 CRC cell
lines were randomly selected for functional analysis in the
following experiments. miR-30d mimic or miR-NC was transfected into
the LoVo and SW480 cells for 48 h, respectively. The efficiency of
transfection was confirmed by RT-qPCR (Fig. 3B). MTT assay showed that in both
the LoVo and SW620 cell lines, the cell proliferation was
significantly inhibited by miR-30d overexpression when compared to
the NC groups, respectively (Fig.
2C). In addition, the invasion assay demonstrated that the
miR-30d mimic markedly suppressed the migratory and invasive
abilities of the LoVo and SW480 cells when compared with the NC
groups, respectively (Fig. 3D and
E). These findings indicate that overexpression of miR-30d
suppressed cell proliferation and invasion in CRC cells.
Upregulation of miR-30d causes cell cycle
arrest and induces cell apoptosis in CRC cells
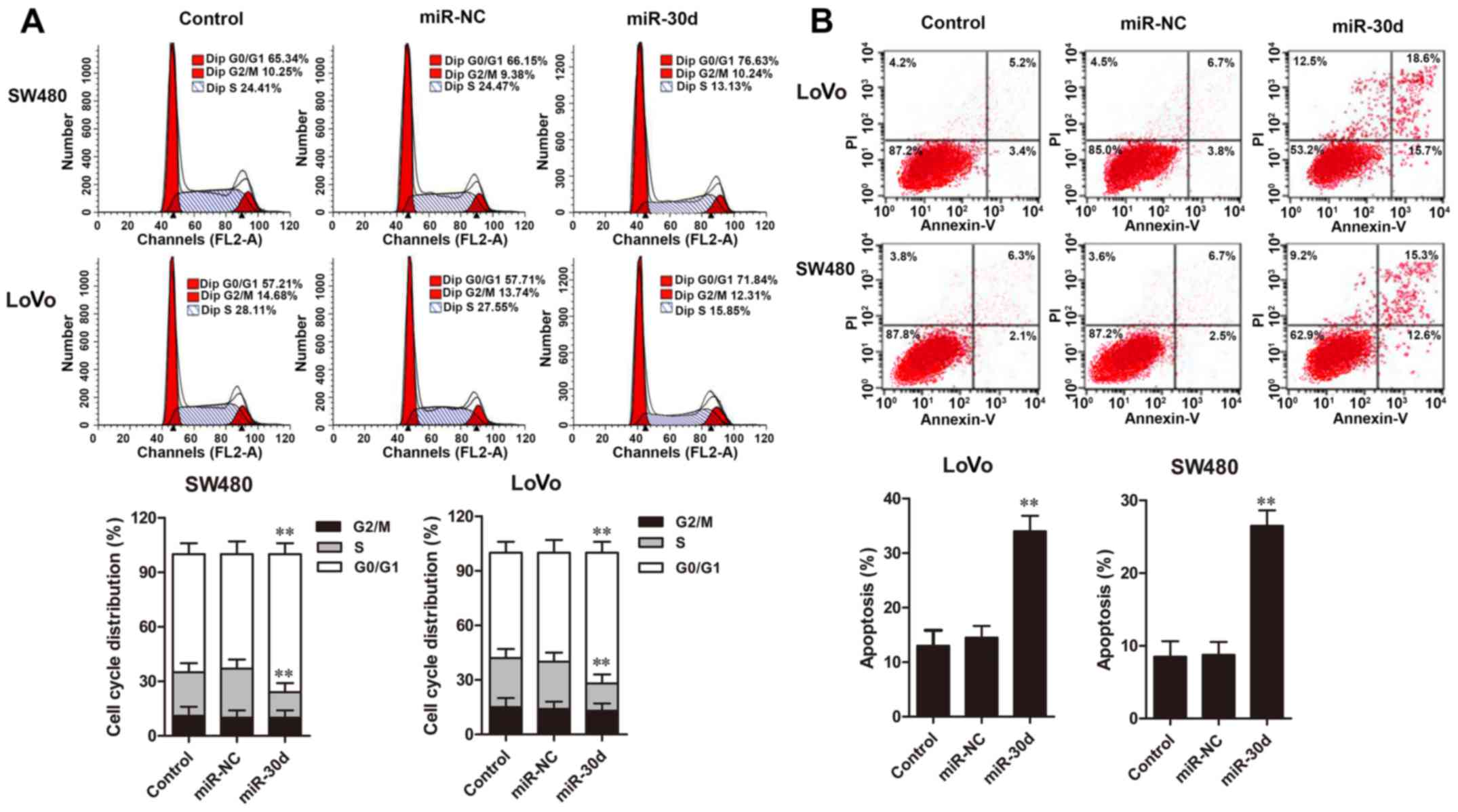
Cell cycle distribution and apoptosis of CRC cells
were assessed by flow cytometric analysis. The results showed that
overexpression of miR-30d in LoVo and SW480 cells resulted in
significant cell accumulation in the G0/G1 phase and a decreased
percentage of cells in the S phase when compared to the NC groups
(Fig. 4A). Furthermore, flow
cytometric analysis revealed that cell apoptosis rates in the LoVo
and SW480 cells with miR-30d mimic transfection were significantly
promoted compared with the rates noted in the NC groups (Fig. 4B). Taken together, these results
demonstrated that upregulation of miR-30d caused cell cycle arrest
at the G0/G1 phase and induced cell apoptosis in the CRC cells.
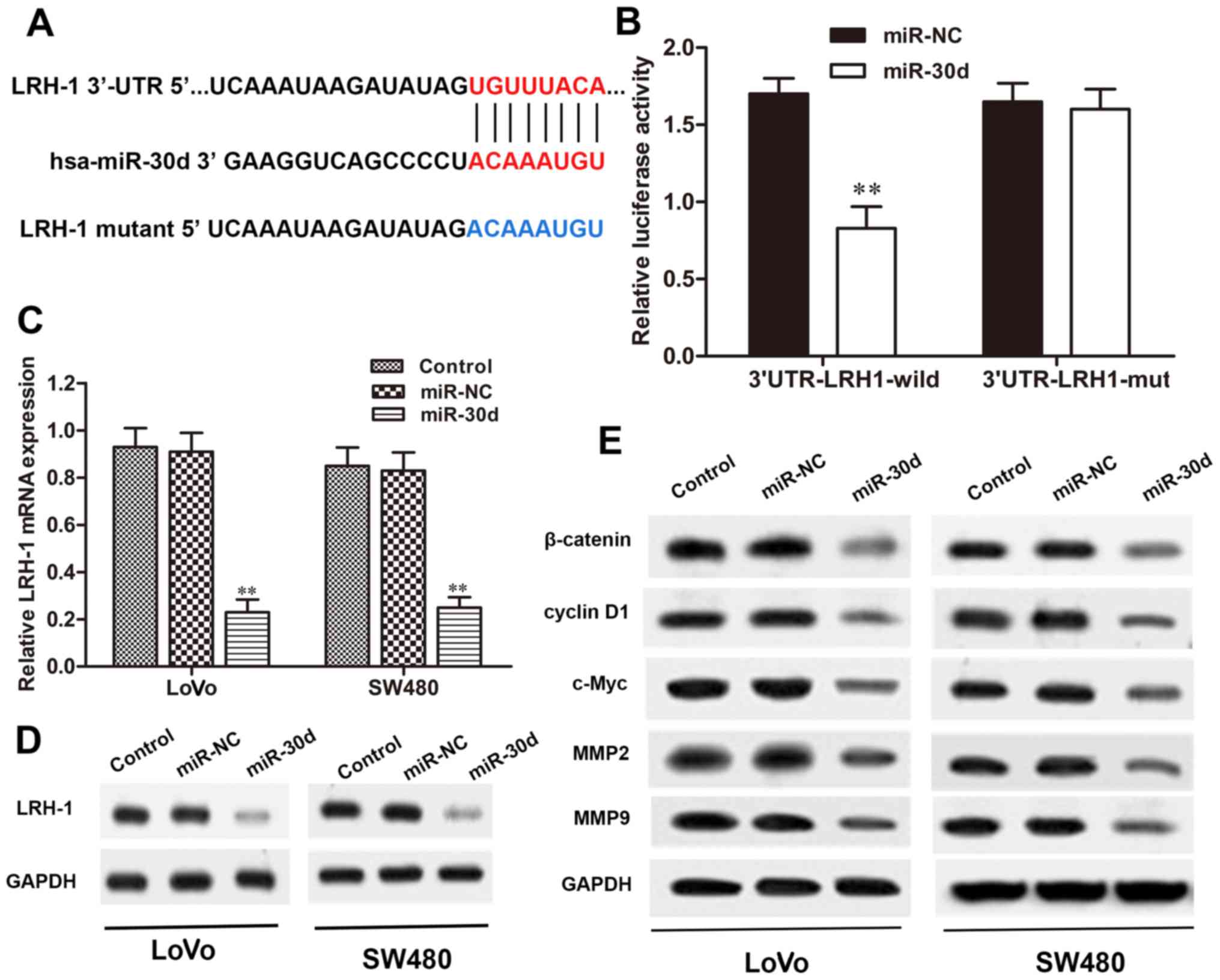
miR-30d directly targets the 3′-UTR of
LRH-1 in CRC cells
To verify whether LRH-1 is a direct target of
miR-30d, we searched for potential targets of miR-30d using
microRNA. org, PicTar, and TargetScan. The binding regions between
the miR-30d and LRH-1 3′-UTR were found (Fig. 5A), indicating that LRH-1 may be a
potential target of miR-30d. To validate that miR-30d directly
targets the 3′-UTR region of LRH-1, a dual-luciferase reporter
assay was performed. When 293T cells were transfected with the
wild-type LRH-1 3′-UTR vector, the luciferase activity was
significantly reduced in the miR-30d mimic co-transfected cells
when compared with the NC group (P<0.01). Conversely, the mutant
3′-UTR of LRH-1 caused no significant changes in the luciferase
activity (Fig. 5B). We next
investigated the interaction between miR-30d and LRH-1 in the LoVo
and SW480 cells. As shown in Fig. 5C
and D, overexpression of miR-30d significantly reduced LRH-1
expression both at the mRNA and protein level in the LoVo and SW480
cells with miR30d mimic transfection, but not in the
miR-NC-transfected cells. These results revealed that miR-30d
directly binds to the site of the 3′-UTR region of LRH-1 and
regulated LRH-1 expression in the CRC cells.
miR-30d regulates the Wnt/β-catenin
signaling pathway through LRH-1
It has been reported that miRNAs play crucial roles
in regulating tumor cell proliferation, differentiation, and
invasion through various signaling pathways, including the
Wnt/β-catenin pathway (23).
Here, to determine the signaling pathway involved in the regulatory
effects of miR-30d on CRC, the Wnt/β-catenin signaling pathway was
detected in LoVo and SW480 cells transfected with the miR-30d mimic
by western blot analysis. The results showed that the expression
levels of Wnt signaling-associated proteins, β-catenin, cyclin D1,
c-Myc, MMP2 and MMP9, were markedly reduced in the miR-30d mimic
group compared to the levels noted in the NC group (Fig. 5E). Taken together, we conclude
that overexpression of miR-30d suppressed activation of the
Wnt/β-catenin signaling pathway by targeting LRH-1 to inhibit CRC
tumorgenesis.
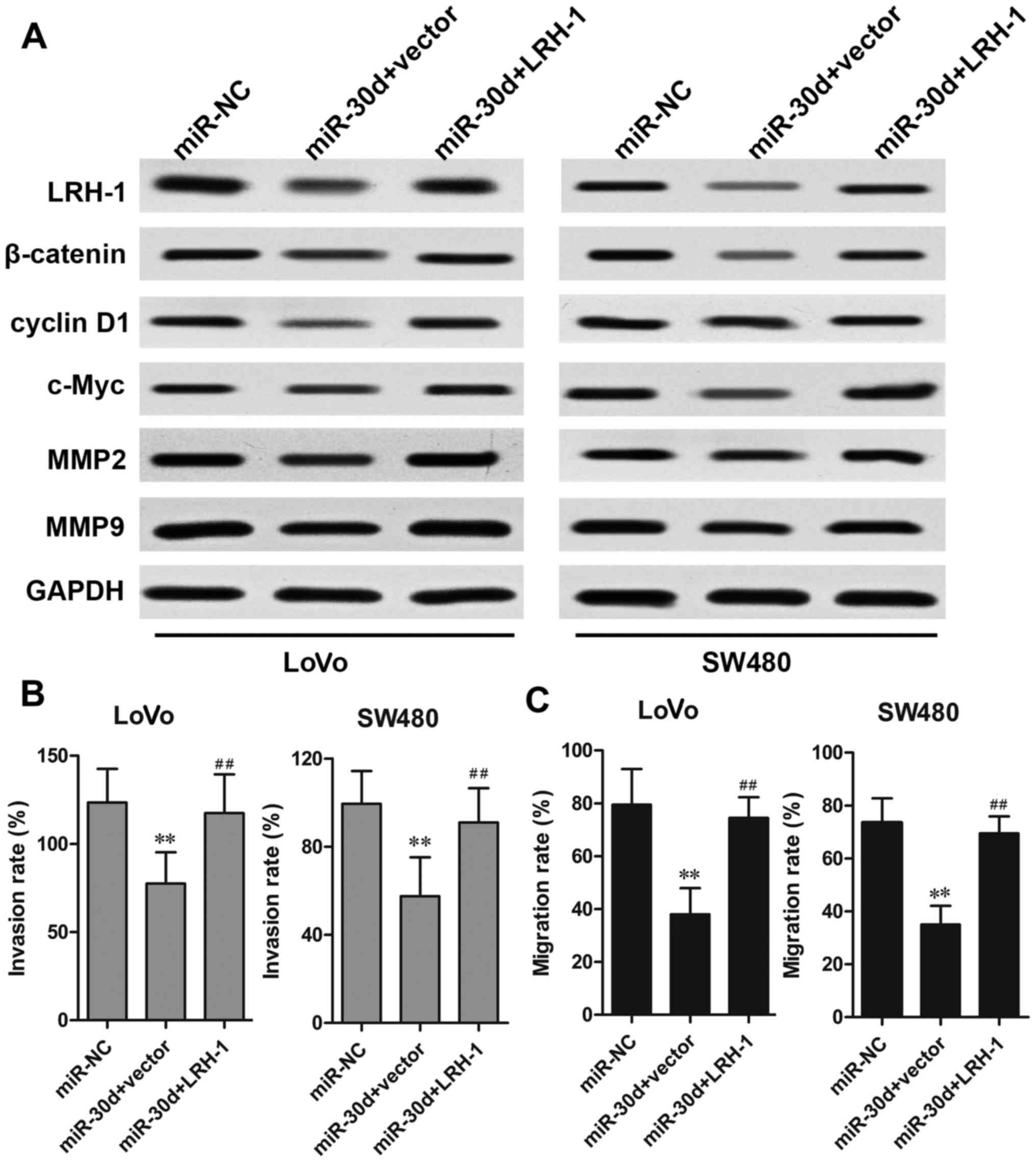
Overexpression of LRH-1 rescues the
effects of miR-30d on CRC cells
To investigate the functional relevance of LRH-1
targeting by miR-30d, a rescue assay was performed to determine
whether LRH-1 overexpression could rescue the suppressive effects
induced by miR-30d. CRC cells were co-transfected with the miR-30d
mimics and LRH-1-overexpressing plasmid. The results showed that
the reduced protein expression caused by miR-30d mimics was
markedly restored by transfection of the LRH-1 overexpressing
plasmid in the LoVo and SW480 cells (Fig. 6A). Overexpression of LRH-1 also
rescued the inhibitory effect of miR-30d on Wnt
signaling-associated proteins, β-catenin, cyclin D1 and c-Myc. In
addition, the inhibitory effects of miR-30d on CRC cell metastasis
regulatory factors MMP2 and MMP9 were significantly reversed by
LRH-1 overexpression, which was further confirmed by invasion and
migration assays (Fig. 6B and C).
These results indicated that miR-30d suppressed CRC tumorigenesis
by inhibiting LRH-1 expression.
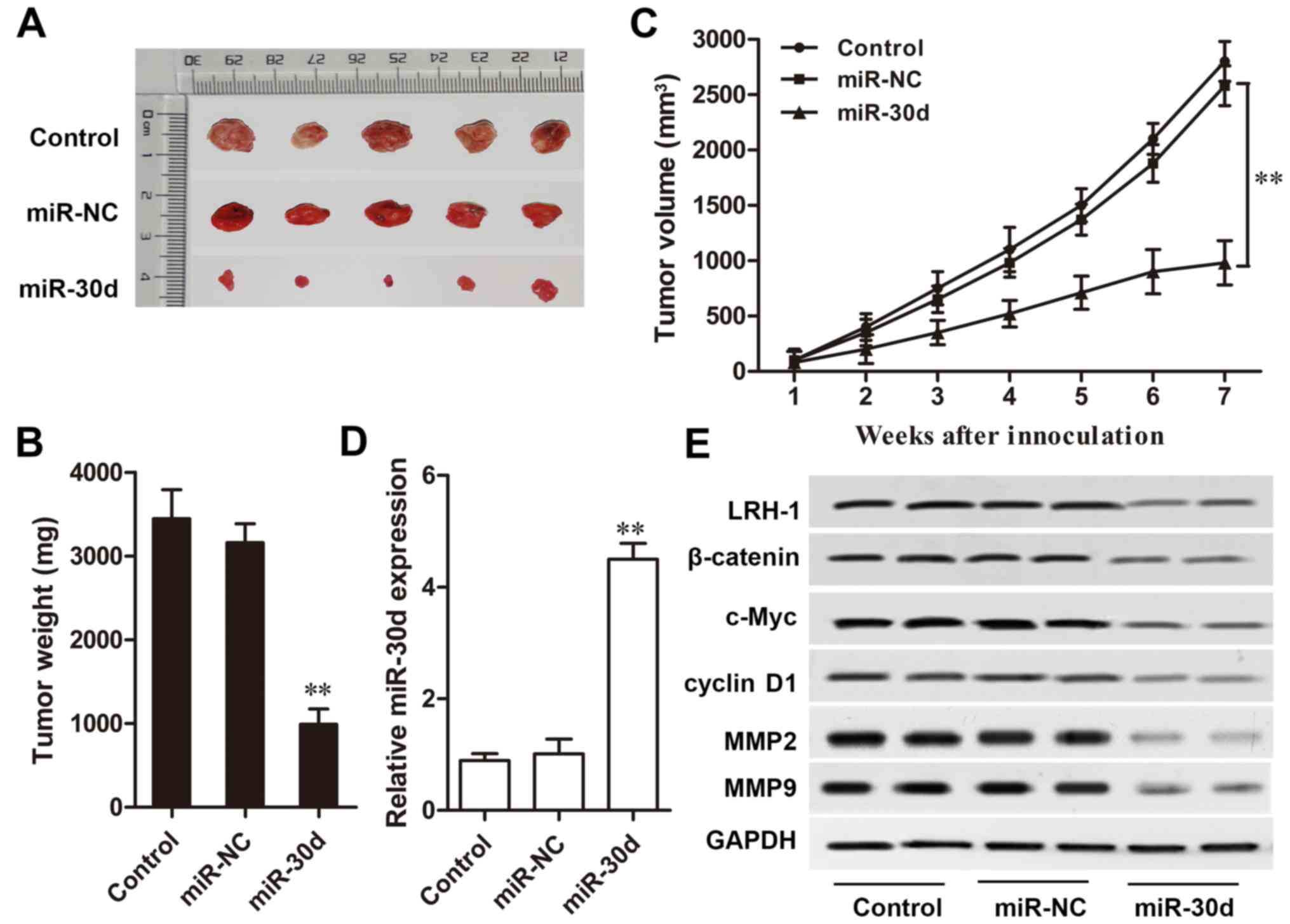
miR-30d inhibits CRC tumor growth in
vivo
A CRC mouse model was designed to investigation the
effect of miR-30d on CRC tumor growth in vivo. Compared with
the NC group, mice injected with miR-30d mimic-transfected cells
showed a markedly slow tumor growth rate during the 7-week
detection (P<0.01, Fig. 7A–C).
Furthermore, RT-qPCR and western blot analysis results showed that
the expression level of miR-30d in the xenograft tumors derived
from miR-30d mimic-transfected cells was markedly upregulated
compared to that in the tumors derived from the miR-NC-transfected
cells (P<0.01, Fig. 7D).
Western blot analysis results also revealed that the expression
levels of key components of the Wnt/β-catenin signaling pathway,
β-catenin, c-Myc and cyclin D1; metastasis regulatory factors, MMP2
and MMP9; as well as LRH-1 were significantly reduced in the
xenograft tumor tissues with miR-30d overexpression (Fig. 7E). These results indicated that
miR-30d suppressed CRC tumor growth in vivo by targeting
LRH-1 by regulating the Wnt/β-catenin signaling pathway.
Discussion
CRC is one of the most prevalent malignancies in the
world, with high morbidity and mortality. Patients with advanced
stage CRC have a poor survival rate (1). Searching for more efficient
prognostic biomarkers for patients with poor prognosis and cancer
metastasis is a critical issue for achieving better outcomes
(24). Recently, focus on the
deregulation of miRNAs during CRC progression has becoming a
promising research field, which provides reliable information for
the distinct understanding of molecular mechanisms underlying CRC
tumorgenesis and development (25). Abnormal miRNA expression is
responsible for alterations in CRC cell proliferation, invasion,
apoptosis and migration (26).
Several oncogenic or tumor-suppressive miRNAs such as miR-21,
miR-182, miR-137, and miR-143 have been demonstrated to be
associated with the initiation of CRC (27,28), Thus, further investigation of the
miRNAs involved in the progression of CRC may contribute to
improved diagnostics and more effective therapeutic strategies for
CRC patients.
Recently, deregulated miR-30d has been shown to be
involved in the development of various types of cancer. Yao et
al reported that miR-30d overexpression enhanced tumor invasion
and metastasis by targeting galphai 2 in hepatocellular carcinoma
(29). Kobayashi et al
found that miR-30d was upregulated in prostate cancer cells and
appeared to be a potential prognostic marker of prostate
progression (30). Moreover, Chen
et al recently demonstrated that miR-30d was downregulated
in non-small cell lung cancer tissues and serves as a tumor
suppressor by inhibiting cell proliferation and motility by
directly targeting cyclin E2 (11). Wu et al found that miR-30d
induced the apoptosis of renal cell carcinoma through the Akt/FOXO
signal transduction pathway (31). In the present study, to avoid the
adverse influence of radiotherapy or chemotherapy on the molecular
expression and distribution in the tumor tissues, which might
indirectly mislead the research results in the study, 80 patients
including stage IV patients with distant metastasis who received
only routine curative surgery were enrolled. We found that miR-30d
expression was significantly downregulated in the CRC tissue
specimens and cell lines. Analysis of the clinicopathological
parameters showed that the miR-30d expression level was closely
associated with poor prognosis, tumor differentiation, invasive
depth, lymph node metastasis, distant metastasis, and TNM stage.
Functional studies also verified the inhibitory role of miR-30d on
cell proliferation, invasion and migration, induction of cell cycle
at the G0/G1 phase and increased apoptosis of CRC in vitro.
Furthermore, we also found that miR-30d overexpression
significantly suppressed tumor growth in vivo, which might
suggest the crucial role of miR-30d as a tumor suppressor in CRC
development and provide valuable evidence for the development of
miR-30d as a novel therapeutic target for CRC treatment.
LRH-1 classified as an orphan NR, is critical to
numerous biological processes and is involved in multiple types of
cancer, including breast, pancreatic, gastrointestinal and colon
cancer. Jiang et al demonstrated that in non-small cell lung
cancer, LRH-1 was a direct target of miR-376c to suppress cell
proliferation and invasion through the Wnt signaling pathway
(32). Liang et al
demonstrated that miR-381 promoted colon cell proliferation and
growth via upregulation of its target gene LRH-1 (33). A recent investigation also
illustrated that LRH-1 functions as a direct target gene of miR-381
in suppressing cell growth and metastasis in hepatocellular
carcinoma (34). However, the
relationship between miR-30d and LRH-1 has not been elucidated in
CRC. Here, we demonstrated that the expression of LRH-1 was
negatively correlated with miR-30d expression in CRC tumor tissues,
and was verified as a direct target of miR-30d using luciferase
reporter assay. Furthermore, restoring the expression of LRH-1
significantly rescued the inhibitory effect of miR-30d
overexpression on CRC tumorigenesis. These results imply that
miR-30d regulates the progression of CRC by directly binding to
LRH-1.
Aberrant activation of the Wnt/β-catenin signaling
pathway contributes to the development and progression of various
cancers including CRC, and is thus regarded as a potential target
in cancer treatment (35,36). The stabilization of β-catenin is
the crucial component of the Wnt signaling pathway. During
activation of Wnt signaling, the accumulation of β-catenin occurs
in the cytoplasm, and then translocates to the nucleus to activate
the transcription of Wnt target genes, such as cyclin D1 and c-Myc
(37,38). It has been reported that LRH-1
functions as a coactivator of the Wnt signaling pathway during
cancer progressions, while c-Myc is required for cell
proliferation, which is also shown to be controlled by LRH-1
(23). For example, by
upregulating cyclin D1/E1 and c-Myc genes, LRH-1 promoted cancer
cell proliferation and tumor growth in pancreatic cancer (19). Furthermore, overexpression of
LRH-1 also significantly enhanced cancer cell migration and
invasion by upregulating its downstream target genes MMP2 and MMP9
in pancreatic cancer cell lines (39). In the present study, we found that
overexpression of miR-30d significantly suppressed the expression
of β-catenin, cyclin D1, c-Myc, as well as MMP2 and MMP9 in CRC
cell lines and xenograft tumors. These results revealed that
upregulation of miR-30d inhibited CRC proliferation and invasion by
regulating LRH-1 and its downstream Wnt/β-catenin signaling
pathway.
Admittedly, there are limitations concerning the
tumor specimen sample size in our study. Thus, in further
investigations, a larger sample size involving more patients with
various statuses in regards to distant metastasis, TNM stage, and
invasive depth are needed to avoid any potential bias effect. In
conclusion, our findings demonstrated that the expression level of
miR-30d was downregulated in CRC tissues and cell lines, and its
expression level was associated with poor prognosis, degree of
tumor differentiation, invasive depth, TNM stage, distant
metastasis and lymph node metastasis. We also discovered that
miR-30d overexpression inhibited cell proliferation, migration and
invasion, induced cell cycle arrest at the G0/G1 phase and
apoptosis, as well as suppressed tumor growth in vivo. In
addition, we identified LRH-1 as a direct target gene of miR-30d
and found that miR-30d regulates Wnt/β-catenin signaling in CRC
progression. These findings demonstrate that miR-30d plays a
tumor-suppressor role in CRC development and inhibits tumorigenesis
by regulating LRH-1 and its downstream Wnt/β-catenin signaling
pathways. Thus, miR-30d is a potential novel therapeutic target for
the treatment of CRC in the clinic.
Acknowledgments
The authors would like to thank Dr L.K. Yan for his
valuable suggestions.
References
|
1
|
Fedewa SA, Ma J, Sauer AG, Siegel RL,
Smith RA, Wender RC, Doroshenk MK, Brawley OW, Ward EM and Jemal A:
How many individuals will need to be screened to increase
colorectal cancer screening prevalence to 80% by 2018? Cancer.
121:4258–4265. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Zheng ZX, Zheng RS, Zhang SW and Chen WQ:
Colorectal cancer incidence and mortality in China, 2010. Asian Pac
J Cancer Prev. 15:8455–8460. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Gellad ZF and Provenzale D: Colorectal
cancer: National and international perspective on the burden of
disease and public health impact. Gastroenterology. 138:2177–2190.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Chen S, Chen X, Xiu YL, Sun KX and Zhao Y:
MicroRNA-490 3P targets CDK1 and inhibits ovarian epithelial
carcinoma tumorigenesis and progression. Cancer Lett. 362:122–130.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Šulc M, Marín RM, Robins HS and Vaníček J:
PACCMIT/PACCMIT-CDS: Identifying microRNA targets in 3′ UTRs and
coding sequences. Nucleic Acids Res. 43:W474–9. 2015. View Article : Google Scholar
|
|
6
|
Fang Z, Tang J, Bai Y, Lin H, You H, Jin
H, Lin L, You P, Li J, Dai Z, et al: Plasma levels of microRNA-24,
microRNA-320a, and microRNA-423 5p are potential biomarkers for
colorectal carcinoma. J Exp Clin Cancer Res. 34:862015. View Article : Google Scholar
|
|
7
|
Nishikawa R, Goto Y, Kurozumi A,
Matsushita R, Enokida H, Kojima S, Naya Y, Nakagawa M, Ichikawa T
and Seki N: MicroRNA-205 inhibits cancer cell migration and
invasion via modulation of centromere protein F regulating pathways
in prostate cancer. Int J Urol. 22:867–877. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Chen W, Fan XM, Mao L, Zhang JY, Li J, Wu
JZ and Tang JH: MicroRNA-224: As a potential target for miR-based
therapy of cancer. Tumour Biol. 36:6645–6652. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Tyagi N, Arora S, Deshmukh SK, Singh S,
Marimuthu S and Singh AP: Exploiting nanotechnology for the
development of microRNA-based cancer therapeutics. J Biomed
Nanotechnol. 12:28–42. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Barger JF and Nana-Sinkam SP: MicroRNA as
tools and therapeutics in lung cancer. Respir Med. 109:803–812.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Chen D, Guo W, Qiu Z, Wang Q, Li Y, Liang
L, Liu L, Huang S, Zhao Y and He X: MicroRNA-30d 5p inhibits tumour
cell proliferation and motility by directly targeting CCNE2 in
non-small cell lung cancer. Cancer Lett. 362:208–217. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Zhang Y, Yang WQ, Zhu H, Qian YY, Zhou L,
Ren YJ, Ren XC, Zhang L, Liu XP, Liu CG, et al: Regulation of
autophagy by miR-30d impacts sensitivity of anaplastic thyroid
carcinoma to cisplatin. Biochem Pharmacol. 87:562–570. 2014.
View Article : Google Scholar :
|
|
13
|
Xuan H, Xue W, Pan J, Sha J, Dong B and
Huang Y: Downregulation of miR-221, -30d, and -15a contributes to
pathogenesis of prostate cancer by targeting Bmi-1. Biochemistry
(Mosc). 80:276–283. 2015. View Article : Google Scholar
|
|
14
|
Ye Z, Zhao L, Li J, Chen W and Li X:
miR-30d blocked transforming growth factor β1-induced
epithelial-mesenchymal transition by targeting snail in ovarian
cancer cells. Int J Gynecol Cancer. 25:1574–1581. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Sablin EP, Blind RD, Uthayaruban R, Chiu
HJ, Deacon AM, Das D, Ingraham HA and Fletterick RJ: Structure of
Liver Receptor Homolog-1 (NR5A2) with IP3 hormone bound in the
ligand binding pocket. J Struct Biol. 192:342–348. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Fayard E, Auwerx J and Schoonjans K:
LRH-1: An orphan nuclear receptor involved in development,
metabolism and steroidogenesis. Trends Cell Biol. 14:250–260. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Stein S and Schoonjans K: Molecular basis
for the regulation of the nuclear receptor LRH-1. Curr Opin Cell
Biol. 33:26–34. 2015. View Article : Google Scholar
|
|
18
|
Schoonjans K, Dubuquoy L, Mebis J, Fayard
E, Wendling O, Haby C, Geboes K and Auwerx J: Liver receptor
homolog 1 contributes to intestinal tumor formation through effects
on cell cycle and inflammation. Proc Natl Acad Sci USA.
102:2058–2062. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Benod C, Vinogradova MV, Jouravel N, Kim
GE, Fletterick RJ and Sablin EP: Nuclear receptor liver receptor
homologue-1 (LRH-1) regulates pancreatic cancer cell growth and
proliferation. Proc Natl Acad Sci USA. 108:16927–16931. 2011.
View Article : Google Scholar
|
|
20
|
Bayrer JR, Mukkamala S, Sablin EP, Webb P
and Fletterick RJ: Silencing LRH-1 in colon cancer cell lines
impairs proliferation and alters gene expression programs. Proc
Natl Acad Sci USA. 112:2467–2472. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Wang SL, Zheng DZ, Lan FH, Deng XJ, Zeng
J, Li CJ, Wang R and Zhu ZY: Increased expression of hLRH-1 in
human gastric cancer and its implication in tumorigenesis. Mol Cell
Biochem. 308:93–100. 2008. View Article : Google Scholar
|
|
22
|
Ray RS, Rana B, Swami B, Venu V and
Chatterjee M: Vanadium mediated apoptosis and cell cycle arrest in
MCF7 cell line. Chem Biol Interact. 163:239–247. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Botrugno OA, Fayard E, Annicotte JS, Haby
C, Brennan T, Wendling O, Tanaka T, Kodama T, Thomas W, Auwerx J
and Schoonjans K: Synergy between LRH-1 and beta-catenin induces G1
cyclin-mediated cell proliferation. Mol Cell. 15:499–509. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Gupta S, Sussman DA, Doubeni CA, Anderson
DS, Day L, Deshpande AR, Elmunzer BJ, Laiyemo AO, Mendez J and
Somsouk M: Challenges and possible solutions to colorectal cancer
screening for the underserved. J Natl Cancer Inst. 106:dju0322014.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Weng W, Feng J, Qin H, Ma Y and Goel A: An
update on miRNAs as biological and clinical determinants in
colorectal cancer: A bench-to-bedside approach. Future Oncol.
11:1791–1808. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Ress AL, Perakis S and Pichler M:
microRNAs and colorectal cancer. Adv Exp Med Biol. 889:89–103.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Yamada A, Horimatsu T, Okugawa Y, Nishida
N, Honjo H, Ida H, Kou T, Kusaka T, Sasaki Y, Yagi M, et al: Serum
miR-21, miR-29a, and miR-125b are promising biomarkers for the
early detection of colorectal neoplasia. Clin Cancer Res.
21:4234–4242. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Yamada A, Cox MA, Gaffney KA, Moreland A,
Boland CR and Goel A: Technical factors involved in the measurement
of circulating microRNA biomarkers for the detection of colorectal
neoplasia. PLoS One. 9:e1124812014. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Yao J, Liang L, Huang S, Ding J, Tan N,
Zhao Y, Yan M, Ge C, Zhang Z, Chen T, et al: MicroRNA-30d promotes
tumor invasion and metastasis by targeting Galphai2 in
hepatocellular carcinoma. Hepatology. 51:846–856. 2010.PubMed/NCBI
|
|
30
|
Kobayashi N, Uemura H, Nagahama K, Okudela
K, Furuya M, Ino Y, Ito Y, Hirano H, Inayama Y, Aoki I, et al:
Identification of miR-30d as a novel prognostic maker of prostate
cancer. Oncotarget. 3:1455–1471. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Wu C, Jin B, Chen L, Zhuo D, Zhang Z, Gong
K and Mao Z: MiR-30d induces apoptosis and is regulated by the
Akt/FOXO pathway in renal cell carcinoma. Cell Signal.
25:1212–1221. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Jiang W, Tian Y, Jiang S, Liu S, Zhao X
and Tian D: MicroRNA-376c suppresses non-small-cell lung cancer
cell growth and invasion by targeting LRH-1-mediated Wnt signaling
pathway. Biochem Biophys Res Commun. 473:980–986. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Liang Y, Zhao Q, Fan L, Zhang Z, Tan B,
Liu Y and Li Y: Down-regulation of microRNA-381 promotes cell
proliferation and invasion in colon cancer through upregulation of
LRH-1. Biomed Pharmacother. 75:137–141. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Zhang Q, Zhao S, Pang X and Chi B:
MicroRNA-381 suppresses cell growth and invasion by targeting the
liver receptor homolog-1 in hepatocellular carcinoma. Oncol Rep.
35:1831–1840. 2016.
|
|
35
|
Anastas JN and Moon RT: WNT signalling
pathways as therapeutic targets in cancer. Nat Rev Cancer.
13:11–26. 2013. View
Article : Google Scholar
|
|
36
|
Luu HH, Zhang R, Haydon RC, Rayburn E,
Kang Q, Si W, Park JK, Wang H, Peng Y, Jiang W and He TC:
Wnt/beta-catenin signaling pathway as a novel cancer drug target.
Curr Cancer Drug Targets. 4:653–671. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Dong Y, Cao B, Zhang M, Han W, Herman JG,
Fuks F, Zhao Y and Guo M: Epigenetic silencing of NKD2, a major
component of Wnt signaling, promotes breast cancer growth.
Oncotarget. 6:22126–22138. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Ozaki S, Ikeda S, Ishizaki Y, Kurihara T,
Tokumoto N, Iseki M, Arihiro K, Kataoka T, Okajima M and Asahara T:
Alterations and correlations of the components in the Wnt signaling
pathway and its target genes in breast cancer. Oncol Rep.
14:1437–1443. 2005.PubMed/NCBI
|
|
39
|
Li C, Heidt DG, Dalerba P, Burant CF,
Zhang L, Adsay V, Wicha M, Clarke MF and Simeone DM: Identification
of pancreatic cancer stem cells. Cancer Res. 67:1030–1037. 2007.
View Article : Google Scholar : PubMed/NCBI
|