Introduction
Stroke is a common and serious condition, with
approximately 795,000 individuals having experienced a new or
recurrent stroke and 128,978 stroke-related deaths having occurred
in the United States in 2013. The prevalence of stroke in the
United States was approximately 6.6 million in 2012, with 87% of
these events being ischemic stroke, 10% intracerebral hemorrhage
(ICH) and 3% subarachnoid hemorrhage (SAH) (1). Despite recent advances in acute
therapy, stroke remains the leading cause of severe disability and
the fifth leading cause of mortality (after heart disease, cancer,
chronic lower respiratory disease and accidents) in the United
States (1). Given that stroke is
a life-threatening condition, its prevention is an important goal
for reducing its overall burden. The identification of biomarkers
for stroke risk is key both for risk prediction and for potential
intervention to avert future cerebrovascular events.
Ischemic stroke is a complex multifactorial disorder
that is thought to result from an interaction between a person's
genetic background and various environmental factors (2). The heritability of ischemic stroke
has been estimated to be 40.3% for large-vessel disease, 32.6% for
cardioembolic stroke, 16.1% for small-vessel disease and 37.9% for
ischemic stroke as a whole (3).
Genome-wide association studies (GWASs) in European ancestry
populations have identified various genes and loci that confer
susceptibility to ischemic stroke (4–11),
with one recent large-scale study having identified HDAC9
and chromosome 1p13.2 (near TSPAN2) as susceptibility loci
for large-vessel disease, PITX2 and ZFHX3 as such
loci for cardioembolic stroke, and 12q24 (near ALDH2) as a
susceptibility locus for small-vessel disease, indicating that
ischemic stroke-related loci are subtype specific (12).
ICH accounts for a large proportion of severe or
fatal cases of stroke, with its most important risk factors being
advanced age and hypertension (13). ICH is usually attributed to
hypertensive small-vessel disease, with the most common sites of
hemorrhage including the basal ganglia, cerebellum and pons
(13,14). In some cases, however, the
hemorrhage is lobar in location, such as in the frontal, parietal,
temporal or occipital cortex, and such patients often do not have
hypertension (14). The
occurrence of lobar ICH has been shown to be associated with the ε2
and ε4 alleles of the apolipoprotein E gene (APOE) (15,16). This relation of lobar ICH to
APOE is presumably due to the association of this gene with
cerebral amyloid angiopathy (17). The heritability of deep or lobar
ICH has been estimated to be 34 and 73%, respectively (18), although genetic factors may
influence, not only the development of ICH, but also hypertension
(19). A previous meta-analysis
of GWASs for ICH in European ancestry populations identified
chromosome 12q21.1 (near TRHDE) as a susceptibility locus
for lobar ICH and 1q22 (near PMF1-SCL25A44) as such a locus
for nonlobar ICH (20).
SAH is commonly caused by the rupture of an aneurysm
in an intracranial artery (21,22). Although the incidence of
aneurysmal SAH in the general population is low (~8/100,000
person-years) (23), a young age
at onset and poor prognosis result in the loss of productive
life-years similar to that for ischemic stroke (24). Given that a family history is an
important risk factor for the development of intracranial aneurysm,
genetics may play an important role in the development of this
condition (25). GWASs have
implicated several loci and genes as conferring susceptibility to
intracranial aneurysm (26–31), with a meta-analysis of such
studies having identified 19 genetic variants related to this
condition (32).
Most genetic variants identified by GWASs for
ischemic stroke, ICH, or intracranial aneurysm have a minor allele
frequency (MAF) of >5% and a small individual effect size. Given
that these common variants explain only a fraction of the
heritability of ischemic and hemorrhagic stroke, it is expected
that low-frequency (MAF of 0.5–5%) or rare (MAF of <0.5%)
variants with larger effect sizes contribute to the genetic
architecture of these conditions (33). Although several polymorphisms have
been found to be significantly associated with ischemic stroke
(34–36) or intracranial aneurysm (30) in Japanese individuals, genetic
variants, including low-frequency and rare variants, that
contribute to genetic susceptibility to ischemic stroke, ICH or SAH
in Japanese individuals remain to be identified definitively.
In this study, we performed exome-wide association
studies (EWASs) with the use of exome array-based genotyping
methods to identify single nucleotide polymorphisms (SNPs) and in
particular, low-frequency or rare coding variants with moderate to
high effect sizes, that confer susceptibility to ischemic stroke,
ICH, or SAH in Japanese individuals. Given that most of the known
low-frequency or rare variants were not included in the arrays
adopted in previous GWASs for these conditions, we applied Illumina
arrays that provide coverage of functional SNPs in entire exons,
including such variants.
Materials and methods
Study subjects
For EWAS of ischemic stroke, 1,575 patients with
ischemic stroke and 9,210 control individuals were examined,
whereas for EWASs of hemorrhagic stroke, 673 patients with ICH, 265
patients with SAH and 9,158 controls were examined. The majority of
the control individuals were the same for the studies of ischemic
and hemorrhagic stroke. The subjects were recruited from
individuals who either visited outpatient clinics of or were
admitted to participating hospitals (Gifu Prefectural Tajimi
Hospital, Tajimi; Gifu Prefectural General Medical Center, Gifu;
Japanese Red Cross Nagoya First Hospital, Nagoya; Inabe General
Hospital, Inabe; Hirosaki University Hospital and Hirosaki Stroke
and Rehabilitation Center, Hirosaki, Japan) due to various symptoms
or for an annual health checkup between 2002 and 2014; were
community-dwelling individuals recruited to a population-based
cohort study in Inabe between 2010 and 2014 or in Tokyo or Kusatsu
between 2011 and 2015; or were cases of autopsy performed at the
Tokyo Metropolitan Geriatric Hospital from 1995 to 2012.
The diagnosis of ischemic stroke, ICH, or SAH was
based on the occurrence of a new and abrupt focal neurological
deficit, with neurological symptoms and signs persisting for >24
h, and it was confirmed by positive findings in computed tomography
or magnetic resonance imaging (or both) of the head. The type of
stroke was determined according to the Classification of
Cerebrovascular Diseases III (37). Given that susceptibility loci for
ischemic stroke are subtype-specific (12), we examined subjects with
atherothrombotic cerebral infarction (large-vessel disease).
For the study of ischemic stroke, subjects with
cardiogenic embolic stroke, lacunar infarction alone, transient
ischemic attack, hemorrhagic stroke, cerebrovascular malformations,
moyamoya disease, cerebral venous sinus thrombosis, brain tumors,
or traumatic cerebrovascular diseases were excluded from
enrollment. For the studies of hemorrhagic stroke, individuals with
ischemic stroke, lacunar infarction, transient ischemic attack,
intracranial hemorrhage resulting from cerebrovascular
malformations, moyamoya disease, cerebral venous sinus thrombosis,
brain tumors, traumatic cerebrovascular diseases, or subdural
hematoma were excluded. The control individuals had no history of
ischemic or hemorrhagic stroke; of aortic, coronary, or peripheral
artery disease; or of other thrombotic, embolic or hemorrhagic
disorders. Individuals with unruptured intracranial aneurysm were
also excluded from the controls. The absence of stroke history was
evaluated with a detailed questionnaire and was confirmed by the
absence of a history of neurological deficits. Autopsy cases were
excluded from the controls.
Body mass index was calculated as follows: body mass
index = body weight (kg)/[body height (m)]2. Blood
pressure was measured at least twice with subjects having first
rested in the sitting position for >5 min; the measurements were
taken by a skilled physician or nurse. Venous blood was collected
in the early morning after the subjects had fasted overnight.
Plasma glucose level, blood hemoglobin A1c, and serum
concentrations of triglycerides, high density lipoprotein
(HDL)-cholesterol, low density lipoprotein (LDL)-cholesterol, and
creatinine were measured with automated analyzers at the clinical
laboratory of each hospital. Estimated glomerular filtration rate
(eGFR) was calculated as follows: eGFR (ml/min/1.73 m2)
= 194 × [age (years)]−0.287 × [serum creatinine
(mg/dl)]−1.094 × [0.739 if female]. Chronic kidney
disease was defined as an eGFR of <60 ml/min/1.73
m2.
The study protocol complied with the Declaration of
Helsinki and was approved by the Committees on the Ethics of Human
Research of Mie University Graduate School of Medicine, Tokyo
Metropolitan Institute of Gerontology, Hirosaki University Graduate
School of Medicine, and participating hospitals. Written informed
consent was obtained from each participant or the families of the
deceased subjects.
EWASs
Venous blood (5 or 7 ml) was collected into tubes
containing 50 mmol/l ethylenediaminetetraacetic acid (disodium
salt), peripheral blood leukocytes were isolated and genomic DNA
was extracted from these cells either with a DNA extraction kit
(Genomix; Talent, Trieste, Italy; or SMITEST EX-R&D; Medical
and Biological Laboratories, Nagoya, Japan) or by standard
protocols based on phenol-chloroform extraction and spin columns.
In autopsy cases, genomic DNA was extracted from the kidneys. EWASs
were performed with the use of a HumanExome-12 v1.1 or v1.2 DNA
Analysis BeadChip or Infinium Exome-24 v1.0 BeadChip (Illumina, San
Diego, CA, USA), both of which include putative functional exonic
variants selected from >12,000 individual exome and whole-genome
sequences. The exonic content of ~244,000 SNPs represents diverse
populations, including European, African, Chinese and Hispanic
individuals (38). SNPs contained
in only one of the exome arrays (~3.6% of all SNPs) were excluded
from the analysis. We performed quality control (39) as follows: i) genotyping data with
a call rate of <97% were discarded, with the mean call rate for
the remaining data being 99.9%; ii) sex specification was examined
for all samples, and those for which sex phenotype in the clinical
records was inconsistent with genetic sex were discarded; iii)
duplicated samples and cryptic relatedness were examined by
calculation of identity by descent; all pairs of DNA samples
showing identity by descent of >0.1875 were inspected, and one
sample from each pair was excluded; iv) heterozygosity of SNPs was
calculated for all samples, with those showing extremely low or
high heterozygosity (>3 standard deviations from the mean) being
discarded; v) SNPs in sex chromosomes or in mitochondrial DNA were
excluded from the analysis, as were non-polymorphic SNPs or SNPs
with a MAF of <0.001; vi) SNPs whose genotype distributions
deviated significantly (P<0.001) from Hardy-Weinberg equilibrium
in control individuals were discarded; vii) genotype data were
examined for population stratification by principal components
analysis (40), and population
outliers were excluded from the analysis. A total of 41,339 and
41,332 SNPs that passed quality control for the studies of ischemic
and hemorrhagic stroke, respectively, were subjected to
analysis.
Statistical analysis
For the analysis of characteristics of the study
subjects, quantitative data were compared between subjects with
ischemic stroke, ICH or SAH and controls with the Mann-Whitney U
test, given that variables exhibited skewed distribution (P<0.01
by the Kolmogorov-Smirnov and Lilliefors test). Categorical data
were compared between two groups with Fisher's exact test. Allele
frequencies were estimated by the gene counting method, and
Fisher's exact test was applied to identify departure from
Hardy-Weinberg equilibrium. Allele frequencies of SNPs were
compared between subjects with ischemic stroke, ICH or SAH and the
controls with Fisher's exact test. Given that the Fisher's exact
test gives exact P-values, it is appropriate to use this test to
examine the relation of low frequency or rare variants to
phenotypes. To compensate for multiple comparisons of genotypes
with ischemic stroke, ICH or SAH, we applied Bonferroni's
correction for statistical significance of association. Given that
41,339 or 41,332 SNPs were analyzed for ischemic and hemorrhagic
stroke, respectively, the significance level was set at
P<1.21×10−6 (0.05/41,339 or 0.05/41,332) for each
EWAS. Quantile-quantile plots for P-values of allele frequencies in
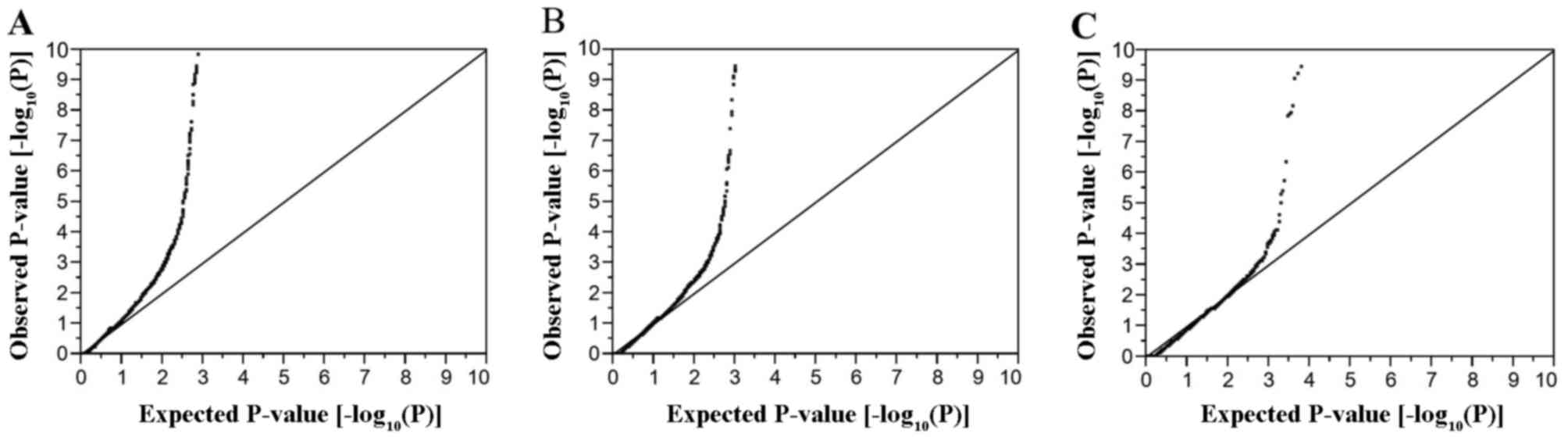
EWASs for ischemic stroke, ICH and SAH are shown in Fig. 1. The inflation factor (λ) was 1.30
for ischemic stroke, 1.52 for ICH, and 1.72 for SAH. Multivariable
logistic regression analysis was performed with ischemic stroke as
a dependent variable and independent variables including age, sex
(0, woman; 1, man), the prevalence of hypertension and diabetes
mellitus (0, no history of these conditions; 1, positive history)
and genotype of each SNP. Similar analysis was performed with ICH
or SAH as a dependent variable and independent variables including
age, sex, the prevalence of hypertension and genotype of each SNP.
Genotypes of each SNP were assessed according to dominant [0, AA;
1, AB + BB (A, major allele; B, minor allele)], recessive (0, AA +
AB; 1, BB) and additive genetic models, and the P-value, odds ratio
and 95% confidence interval were calculated. Additive models
comprised additive 1 (0, AA; 1, AB; 0, BB) and additive 2 (0, AA;
0, AB; 1, BB) scenarios, which were analyzed simultaneously with a
single statistical model. The relation of genotypes of SNPs to
intermediate phenotypes was examined with Fisher's exact test (2×2)
or Pearson's Chi-square test (2×3). Bonferroni's correction was
also applied to other statistical analysis as indicated.
Statistical tests were performed with JMP Genomics version 6.0
software (SAS Institute, Cary, NC, USA).
Results
EWAS for ischemic stroke
The characteristics of the subjects enrolled in the
study are shown in Table I. Age,
the frequency of the male gender, body mass index, and the
prevalence of hypertension, diabetes mellitus and chronic kidney
disease, as well as systolic and diastolic blood pressure, fasting
plasma glucose levels, blood glycosylated hemoglobin (hemoglobin
A1c) content and serum concentrations of triglycerides
and creatinine were greater, whereas the serum concentrations of
HDL-cholesterol and estimated glomerular filtration rate were
lower, in the subjects with ischemic stroke than in the
controls.
 | Table ICharacteristics of the 10,785
subjects in the exome-wide association study for ischemic
stroke. |
Table I
Characteristics of the 10,785
subjects in the exome-wide association study for ischemic
stroke.
| Characteristic | Ischemic
stroke | Controls | P-value |
|---|
| No. of
subjects | 1,575 | 9,210 | |
| Age (years) | 71.8±12.2 | 58.8±13.8 | <0.0001 |
| Sex (male/female,
%) | 58.9/41.1 | 50.4/49.6 | <0.0001 |
| Body mass index
(kg/m2) | 23.5±3.5 | 23.1±3.5 | <0.0001 |
| Current or former
smoker (%) | 33.9 | 37.2 | 0.0450 |
| Hypertension
(%) | 79.2 | 42.4 | <0.0001 |
| Systolic blood
pressure (mmHg) | 147±27 | 125±20 | <0.0001 |
| Diastolic blood
pressure (mmHg) | 82±16 | 75±12 | <0.0001 |
| Diabetes mellitus
(%) | 44.0 | 14.5 | <0.0001 |
| Fasting plasma
glucose (mmol/l) | 7.09±2.88 | 5.80±1.95 | <0.0001 |
| Blood hemoglobin
A1c (%) | 6.34±1.42 | 5.70±0.93 | <0.0001 |
| Dyslipidemia
(%) | 58.7 | 57.6 | 0.5082 |
| Serum triglycerides
(mmol/l) | 1.47±0.96 | 1.38±0.96 | <0.0001 |
| Serum
HDL-cholesterol (mmol/l) | 1.28±0.42 | 1.62±0.44 | <0.0001 |
| Serum
LDL-cholesterol (mmol/l) | 3.10±0.98 | 3.13±0.80 | 0.0208 |
| Chronic kidney
disease (%) | 36.8 | 18.8 | <0.0001 |
| Serum creatinine
(µmol/l) | 90.3±115.5 | 72.9±6804 | <0.0001 |
| eGFR (ml
m−1 1.73 m−2) | 67.7±22.7 | 74.0±18.1 | <0.0001 |
| Hyperuricemia
(%) | 16.9 | 16.1 | 0.4794 |
| Serum uric acid
(µmol/l) | 329±96 | 323±90 | 0.1073 |
We examined the relation of allele frequencies for
41,339 SNPs that passed quality control to ischemic stroke with
Fisher's exact test. A Manhattan plot of EWAS for ischemic stroke
is shown in Fig. 2. After
Bonferroni's correction, 77 SNPs were found to be significantly
(P<1.21×10−6) associated with ischemic stroke
(Table II). The genotype
distributions of these SNPs were in Hardy-Weinberg equilibrium
(P>0.001) among both subjects with ischemic stroke and the
controls (data not shown).
| Table IIThe 77 SNPs significantly
(P<1.21×10−6) associated with ischemic stroke in the
exome-wide association study. |
Table II
The 77 SNPs significantly
(P<1.21×10−6) associated with ischemic stroke in the
exome-wide association study.
| Gene | dbSNP | Nucleotide (amino
acid) substitutiona | Chromosome:
position | MAF (%) | P-value
(allele) | Allele OR | 95% CI |
|---|
| TTLL5 | rs2833270 | C/T | 21: 3115036 | 4.1 |
5.45×10−112 | 1.05 | 0.87–1.27 |
| rs605066 | C/T | 6: 139508529 | 25.6 |
1.93×10−89 | 1.02 | 0.94–1.11 |
| rs146036604 | A/C (L879F) | 14: 75783181 | 1.4 |
1.09×10−73 | 0.90 | 0.65–1.25 |
| rs12615742 | T/C | 2: 37768584 | 47.3 |
3.80×10−62 | 1.04 | 0.96–1.12 |
| TCEB3B | rs2010834 | A/C (F254C) | 18: 47034504 | 24.5 |
2.40×10−61 | 1.02 | 0.94–1.11 |
| TCF19 | rs3130933 | G/A | 6: 31164308 | 5.2 |
1.05×10−53 | 1.18 | 1.00–1.39 |
| GABRB3 | rs3212335 | G/A | 15: 26766994 | 38.9 |
8.21×10−53 | 1.02 | 0.94–1.11 |
| NRXN3 | rs6574433 | G/A | 14: 78319816 | 26.5 |
6.14×10−49 | 0.98 | 0.90–1.06 |
| UBASH3A | rs11203203 | G/A | 21: 42416077 | 3.9 |
7.70×10−44 | 1.13 | 0.93–1.37 |
| TBC1D32 | rs79221470 | T/A (C505S) | 6: 121281639 | 17.8 |
5.10×10−37 | 1.01 | 0.92–1.11 |
| DBT | rs140308307 | T/C (N13D) | 1: 100249784 | 0.3 |
2.69×10−36 | 1.59 | 0.86–2.94 |
| IL20RA | rs191996643 | T/C (I43V) | 6: 137017065 | 0.8 |
2.63×10−31 | 0.74 | 0.47–1.18 |
| MMP28 | rs117651561 | G/A | 17: 35766644 | 13.6 |
1.80×10−29 | 1.01 | 0.90–1.12 |
|
C15orf57 | rs3803354 | T/C | 15: 40564790 | 8.9 |
3.88×10−27 | 1.11 | 0.98–1.27 |
| TMPRSS7 | rs147783135 | C/T
(R692*) | 3: 112081004 | 0.8 |
3.41×10−26 | 0.54 | 0.31–0.94 |
| rs7752978 | A/G | 6: 114869897 | 47.4 |
1.29×10−24 | 1.00 | 0.92–1.08 |
| HMGCS2 | rs181428774 | G/T (L59M) | 1: 119764556 | 0.3 |
1.56×10−24 | 0.49 | 0.21–1.14 |
| DPF3 | rs757572 | T/C | 14: 72665321 | 41.2 |
1.28×10−21 | 1.07 | 0.99–1.15 |
| SLC6A4 | rs56316081 | T/C (I108V) | 17: 30221637 | 0.1 |
2.54×10−21 | ND | ND |
| OSGEPL1 | rs75321854 | C/T (A98T) | 2: 189755490 | 4.8 |
2.31×10−20 | 1.10 | 0.93–1.31 |
| S100A3 | rs36022742 | C/T (R3K) | 1: 153548478 | 1.6 |
4.16×10−20 | 0.77 | 0.55–1.09 |
| ALMS1 | rs3820700 | G/A (S2576N) | 2: 73489683 | 26.1 |
6.70×10−20 | 0.98 | 0.90–1.08 |
| OR2D2 | rs1965209 | A/G (S148P) | 11: 6892059 | 12.2 |
9.24×10−20 | 1.11 | 0.99–1.24 |
| rs9494145 | T/C | 6: 135111414 | 31.8 |
9.77×10−20 | 0.96 | 0.88–1.04 |
| AXDND1 | rs41267592 | C/T (T627M) | 1: 179468524 | 0.3 |
1.92×10−19 | 0.61 | 0.28–1.33 |
| SCMH1 | rs182666831 | A/T (S48C) | 1: 41151649 | 0.1 |
5.23×10−19 | 0.58 | 0.18–1.92 |
| ELMO1 | rs11984075 | A/G | 7: 37397251 | 1.1 |
3.61×10−18 | 0.78 | 0.53–1.16 |
| OTOL1 | rs12633334 | G/A | 3: 161452823 | 45.1 |
2.34×10−17 | 0.97 | 0.89–1.04 |
| rs5026743 | T/G | 6: 32472187 | 29.7 |
4.27×10−17 | 1.12 | 1.03–1.22 |
| MUC16 | rs12459532 | G/A (P9422L) | 19: 8948505 | 1.8 |
4.29×10−17 | 1.42 | 1.10–1.85 |
| TMPRSS6 | rs2235321 | G/A | 22: 37066886 | 43.7 |
7.08×10−17 | 1.09 | 1.01–1.18 |
| rs11624336 | G/A | 14: 96727175 | 14.9 |
2.40×10−16 | 0.99 | 0.88–1.10 |
|
KIAA1324 | rs1052878 | C/T (P922L) | 1: 109202996 | 4.9 |
6.66×10−16 | 1.05 | 0.89–1.24 |
| MMP28 | rs79742527 | G/A | 17: 35766662 | 13.6 |
1.04×10−15 | 1.00 | 0.89–1.11 |
| SERINC1 | rs11964202 | G/A | 6: 122444993 | 0.6 |
1.64×10−15 | 0.87 | 0.51–1.47 |
| KANK4 | rs2258470 | C/T (R822H) | 1: 62263166 | 19.3 |
8.84×10−15 | 1.13 | 1.03–1.24 |
| PRR22 | rs201451364 | C/T (R78H) | 19: 5784014 | 0.1 |
1.42×10−14 | 1.54 | 0.57–4.13 |
| PLCB1 | rs6140742 | C/G | 20: 8838465 | 36.3 |
3.52×10−14 | 1.01 | 0.94–1.09 |
| AHNAK2 | rs150385420 | A/G (S4485P) | 14: 104941998 | 2.6 |
6.43×10−14 | 0.98 | 0.77–1.23 |
| FBXO16 | rs3735726 | C/T (R63Q) | 8: 28463730 | 3.3 |
4.02×10−13 | 1.14 | 0.93–1.39 |
| CALR3 | rs3810198 | T/C | 19: 16490383 | 28.6 |
1.45×10−12 | 1.03 | 0.95–1.12 |
| PDGFD | rs10895547 | C/T | 11: 103937424 | 41.0 |
1.74×10−12 | 0.99 | 0.92–1.07 |
|
LOC101928877 | rs6844558 | T/C | 4: 185060282 | 38.5 |
4.86×10−11 | 1.02 | 0.94–1.10 |
| MBD2 | rs1145315 | T/C | 18: 54162585 | 33.3 |
1.47×10−10 | 0.95 | 0.88–1.03 |
| LRTM1 | rs182902370 | C/T | 3: 54918891 | 0.9 |
3.41×10−10 | 1.10 | 0.74–1.62 |
| SEC16B | rs202187751 | G/C (D41H) | 1: 177967861 | 0.2 |
4.18×10−10 | 0.53 | 0.16–1.72 |
| NEURL4 | rs117553236 | T/G (M1048L) | 17: 7321417 | 0.3 |
5.35×10−10 | 0.97 | 0.41–2.33 |
| PPFIA1 | rs546502 | G/A (V71I) | 11: 70272383 | 15.9 |
6.09×10−10 | 1.18 | 1.08–1.30 |
| SLC18A1 | rs1390938 | G/A (T136I) | 8: 20179202 | 25.5 |
6.98×10−10 | 1.06 | 0.97–1.15 |
| DDOST | rs74526704 | C/T (R396Q) | 1: 20652655 | 0.2 |
9.25×10−10 | 0.86 | 0.34–2.20 |
| ZNF16 | rs139521477 | C/T (R669Q) | 8: 144930781 | 0.1 |
9.85×10−10 | 0.73 | 0.22–2.43 |
| TTC16 | rs142193455 | G/A (R450Q) | 9: 127726328 | 1.1 |
1.05×10−9 | 0.60 | 0.38–0.93 |
| SEMA4D | rs13295305 | C/T (R713K) | 9: 89363482 | 12.7 |
1.24×10−9 | 1.07 | 0.96–1.20 |
| TRH | rs13306057 | G/A (G3S) | 3: 129975823 | 0.2 |
1.47×10−9 | 0.44 | 0.14–1.41 |
| MAST4 | rs56337909 | G/A (S1863N) | 5: 67164767 | 2.7 |
3.19×10−9 | 1.01 | 0.79–1.28 |
| PTPRR | rs10784867 | C/T | 12: 70740947 | 48.3 |
4.83×10−9 | 0.98 | 0.91–1.06 |
| PREX1 | rs6095241 | G/A | 20: 48692260 | 41.5 |
4.95×10−9 | 1.09 | 1.01–1.18 |
| rs12402711 | G/A | 1: 41413409 | 27.1 |
6.95×10−9 | 1.00 | 0.92–1.09 |
| rs10277516 | C/A | 7: 22869155 | 3.3 |
2.30×10−8 | 1.08 | 0.88–1.33 |
| ROR2 | rs200805854 | C/A (E703D) | 9: 91724385 | 0.2 |
3.93×10−8 | 0.89 | 0.35–2.27 |
| DUS4L | rs4730250 | A/G | 7: 107567250 | 9.4 |
4.04×10−8 | 0.91 | 0.80–1.04 |
| rs11185362 | A/G | 1: 104018866 | 24.2 |
4.37×10−8 | 1.04 | 0.96–1.14 |
| IL7R | rs3194051 | A/G (I356V) | 5: 35876172 | 7.4 |
6.06×10−8 | 1.01 | 0.87–1.16 |
| FAM200A | rs75129401 | T/C (H35R) | 7: 99548304 | 1.5 |
6.08×10−8 | 0.64 | 0.44–0.93 |
| PLXNC1 | rs75674989 | G/T (R614S) | 12: 94226656 | 3.8 |
6.12×10−8 | 0.84 | 0.68–1.03 |
| rs7131744 | G/A | 12: 4387114 | 44.7 |
7.92×10−8 | 1.09 | 1.01–1.18 |
| PDIA5 | rs2292661 | C/T (T391M) | 3: 123150263 | 0.7 |
1.03×10−7 | 0.47 | 0.26–0.88 |
| GOLGB1 | rs3732407 | G/C (S911T) | 3: 121697776 | 3.5 |
1.11×10−7 | 1.15 | 0.94–1.41 |
| ARPC1B | rs1045012 | G/C (K37N) | 7: 99386731 | 1.7 |
1.97×10−7 | 0.94 | 0.70–1.27 |
| ALMS1 | rs138921247 | G/C (V807L) | 2: 73448943 | 1.8 |
2.71×10−7 | 0.87 | 0.65–1.16 |
| TRIM67 | rs1998027 | A/G | 1: 231190340 | 33.7 |
2.80×10−7 | 0.98 | 0.90–1.06 |
|
C16orf95 | rs3748393 | A/C (S26A) | 16: 87317167 | 41.7 |
3.24×10−7 | 1.06 | 0.98–1.14 |
| CES5A | rs145397395 | C/T (V220M) | 16: 55866010 | 0.9 |
4.76×10−7 | 0.85 | 0.76–1.79 |
| ACTR5 | rs3752289 | C/T (P580L) | 20: 38771731 | 1.4 |
5.15×10−7 | 0.98 | 0.71–1.36 |
| rs3095354 | A/G | 6: 30868334 | 26.7 |
5.75×10−7 | 1.12 | 1.03–1.22 |
| CYP4F12 | rs191885206 | T/C (C402R) | 19: 15696024 | 0.3 |
6.57×10−7 | 2.11 | 1.23–3.57 |
| CELSR2 | rs117684956 | G/A (V598M) | 1: 109251871 | 1.3 |
9.21×10−7 | 1.25 | 0.93–1.69 |
Multivariable logistic regression
analysis of the relation of SNPs to ischemic stroke
The relation of the 77 SNPs identified by EWAS to
ischemic stroke was examined further by multivariable logistic
regression analysis with adjustment for age, sex and the prevalence
of hypertension and diabetes mellitus (Table III). Four SNPs, rs3212335 (G/A)
of γ-aminobutyric acid type A receptor β3 subunit (GABRB3),
rs147783135 [C/T (R692*)] of transmembrane protease,
serine 7 gene (TMPRSS7), rs2292661 [C/T (T391M)] of protein
disulfide isomerase family A member 5 gene (PDIA5), and
rs191885206 [T/C (C402R)] of cytochrome P450 family 4 subfamily F
member 12 gene (CYP4F12) were related (P<0.01 in at least
one genetic model) to ischemic stroke, although no SNP was
significantly [P<1.62×10−4 (0.05/308)] associated
with this condition. The minor A and C alleles of rs3212335 and
rs191885206, respectively, were risk factors for ischemic stroke,
whereas the minor T alleles of rs147783135 and rs2292661 were
protective against this condition.
| Table IIIRelation of SNPs to ischemic stroke
as determined by multivariable logistic regression analysis. |
Table III
Relation of SNPs to ischemic stroke
as determined by multivariable logistic regression analysis.
| SNP | Dominant
| Recessive
| Additive 1
| Additive 2
|
|---|
| P-value | OR (95% CI) | P-value | OR (95% CI) | P-value | OR (95% CI) | P-value | OR (95% CI) |
|---|
| rs3212335 | G/A | 0.6428 | | 0.0036 | 1.29
(1.09–1.53) | 0.5896 | | 0.0150 | 1.26
(1.05–1.52) |
| rs147783135 | C/T
(R692*) | 0.0024 | 0.37
(0.16–0.72) | 0.4322 | | 0.0029 | 0.38
(0.17–0.74) | 0.4296 | |
| rs2292661 | C/T (T391M) | 0.0054 | 0.35
(0.14–0.76) | ND | | 0.0054 | 0.35
(0.14–0.76) | ND | |
| rs191885206 | T/C (C402R) | 0.0082 | 2.60
(1.30–4.93) | ND | | 0.0082 | 2.60
(1.30–4.93) | ND | |
EWAS for ICH
The characteristics of the subjects for EWAS for ICH
are shown in Table IV. Age, the
frequency of the male gender, and the prevalence of hypertension
and diabetes mellitus were greater in the subjects with ICH than in
the controls.
| Table IVCharacteristics of the subjects in
the exome-wide association studies for ICH or subarachnoid
hemorrhage. |
Table IV
Characteristics of the subjects in
the exome-wide association studies for ICH or subarachnoid
hemorrhage.
| Characteristic | Controls | ICH | P-value | SAH | P-value |
|---|
| No. of
subjects | 9158 | 673 | | 265 | |
| Age (years) | 58.8±13.9 | 69.4±13.9 | <0.0001 | 59.4±13.2 | 0.8515 |
| Sex (male/female,
%) | 50.4/49.6 | 62.6/37.4 | <0.0001 | 41.1/58.9 | 0.0033 |
| Body mass index
(kg/m2) | 23.1±3.5 | 23.1±3.8 | 0.9986 | 23.1±3.3 | 0.9672 |
| Current or former
smoker (%) | 37.1 | 33.3 | 0.1836 | 31.5 | 0.1590 |
| Hypertension
(%) | 42.4 | 81.4 | <0.0001 | 60.5 | <0.0001 |
| Diabetes mellitus
(%) | 14.5 | 38.1 | <0.0001 | 19.6 | 0.0345 |
| Dyslipidemia
(%) | 57.5 | 51.5 | 0.0152 | 41.2 | <0.0001 |
| Chronic kidney
disease (%) | 18.8 | 21.2 | 0.2656 | 28.0 | 0.0052 |
| Hyperuricemia
(%) | 16.0 | 15.6 | 0.8915 | 9.4 | 0.0060 |
We examined the relation of allele frequencies for
41,332 SNPs to ICH with Fisher's exact test. A Manhattan plot for
EWAS for ICH is shown in Fig. 3A.
After Bonferroni's correction, 48 SNPs were significantly
(P<1.21×10−6) associated with ICH (Table V). The genotype distributions of
these SNPs were in Hardy-Weinberg equilibrium (P>0.001) both
among subjects with ICH and among controls (data not shown).
| Table VThe 48 SNPs significantly
(P<1.21×10−6) associated with intracerebral
hemorrhage in the exome-wide association study. |
Table V
The 48 SNPs significantly
(P<1.21×10−6) associated with intracerebral
hemorrhage in the exome-wide association study.
| Gene | dbSNP | Nucleotide (amino
acid) substitutiona | Chromosome:
position | MAF (%) | P-value
(allele) | Allele OR | 95% CI |
|---|
| RECQL | rs146924988 | A/G (M1T) | 12: 21499569 | 0.3 |
8.57×10−103 | 0.93 | 0.29–3.00 |
| ATF7IP2 | rs13335336 | A/G | 16: 10465406 | 17.0 |
3.91×10−93 | 1.15 | 1.00–1.33 |
| SHCBP1 | rs11545690 | A/C (M60R) | 16: 46618297 | 5.3 |
3.58×10−76 | 0.87 | 0.67–1.13 |
| rs1405262 | T/C | 2: 5994808 | 41.3 |
6.36×10−70 | 0.98 | 0.88–1.10 |
| CAT | rs7943316 | A/T | 11: 34438925 | 33.7 |
1.49×10−53 | 1.02 | 0.91–1.15 |
| EXOC6B | rs1517182 | C/A | 2: 72508184 | 1.7 |
8.40×10−52 | 0.91 | 0.59–1.41 |
| RNASE10 | rs202109789 | G/A (G87S) | 14: 20510730 | 0.2 |
3.52×10−49 | 1.00 | 0.31–3.25 |
| GKN2 | rs146849599 | C/T (V130M) | 2: 68946388 | 2.7 |
6.04×10−49 | 0.82 | 0.57–1.18 |
| IGSF10 | rs78090556 | G/A (T981M) | 3: 151447039 | 0.2 |
1.49×10−48 | 2.57 | 1.08–6.25 |
| GFM2 | rs77099085 | G/T (H92N) | 5: 74759397 | 6.4 |
5.68×10−46 | 0.93 | 0.74–1.17 |
| ADRB2 | rs1042713 | G/A (G16R) | 5: 148826877 | 49.1 |
4.57×10−36 | 1.02 | 0.92–1.15 |
| SPN | rs3764276 | C/T | 16: 29661882 | 37.3 |
5.46×10−36 | 1.13 | 1.01–1.26 |
| rs3135365 | T/G | 6: 32421478 | 18.9 |
4.03×10−32 | 0.99 | 0.85–1.14 |
| NAA25 | rs12231744 | C/T (R876K) | 12: 112039251 | 35.1 |
1.00×10−30 | 1.14 | 1.02–1.28 |
| DNAH11 | rs78763603 | G/A | 7: 21698150 | 15.3 |
3.69×10−24 | 0.99 | 0.85–1.15 |
|
C15orf57 | rs3803354 | T/C | 15: 40564790 | 8.9 |
6.29×10−24 | 0.92 | 0.76–1.12 |
| BDP1 | rs34529158 | C/A (P1669Q) | 5: 71522303 | 0.5 |
3.02×10−23 | 0.94 | 0.44–2.04 |
| FLG | rs2184953 | G/A (H2194Y) | 1: 152308306 | 36.2 |
9.10×10−21 | 0.98 | 0.88–1.10 |
| rs563694 | T/G | 2: 168917561 | 2.1 |
1.75×10−20 | 1.24 | 0.85–1.79 |
|
LOC100996813 | rs2453589 | G/A | 17: 19585538 | 26.0 |
8.53×10−19 | 1.09 | 0.96–1.23 |
| DPF3 | rs757572 | T/C | 14: 72665321 | 41.2 |
2.14×10−17 | 1.03 | 0.93–1.16 |
| CCDC18 | rs3820059 | G/A (S172F) | 1: 169421916 | 7.1 |
2.24×10−15 | 0.98 | 0.79–1.22 |
| rs11624336 | G/A | 14: 96727175 | 14.9 |
1.56×10−14 | 0.99 | 0.84–1.15 |
| AXDND1 | rs41267592 | C/T (T627M) | 1: 179468524 | 0.3 |
1.65×10−14 | 0.62 | 0.19–1.97 |
| rs13234712 | G/A | 7: 119939419 | 38.8 |
1.39×10−13 | 0.98 | 0.88–1.10 |
| MAPT | rs3785879 | C/A | 17: 45908270 | 40.8 |
8.89×10−12 | 0.95 | 0.85–1.06 |
| NLRP13 | rs17711239 | T/C (N781S) | 19: 55907897 | 9.3 |
1.57×10−11 | 1.05 | 0.87–1.27 |
| AIF1 | rs2857697 | A/G | 6: 31617442 | 34.1 |
1.84×10−11 | 1.19 | 1.06–1.34 |
| STYK1 | rs138533962 | G/A (R379C) | 12: 10620278 | 2.0 |
3.90×10−11 | 401.7 | 100.0–995.0 |
| CCDC169 | rs9546897 | T/C (K120R) | 13: 36254100 | 34.6 |
3.74×10−10 | 1.04 | 0.93–1.16 |
| HMHA1 | rs150294461 | G/A (G654E) | 19: 1080682 | 1.4 |
4.63×10−10 | 1.12 | 0.69–1.79 |
| TJP3 | rs1046268 | C/T (T898M) | 19: 3750617 | 29.5 |
5.56×10−10 | 1.00 | 0.88–1.13 |
| PAX5 | rs2297105 | A/C | 9: 37020625 | 48.3 |
7.69×10−10 | 0.98 | 0.88–1.09 |
| rs4996815 | G/T | 13: 105999312 | 12.6 |
8.66×10−10 | 1.03 | 0.88–1.22 |
|
HLA-DPB1 | rs9277471 | A/G | 6: 33085905 | 43.6 |
1.55×10−9 | 1.01 | 0.90–1.12 |
| SSC4D | rs10227141 | C/T (R505G) | 7: 76390273 | 14.4 |
4.91×10−9 | 0.94 | 0.80–1.10 |
| ZCCHC11 | rs138145860 | A/G (I1270T) | 1: 52445800 | 0.2 |
1.04×10−8 | 0.93 | 0.29–2.99 |
| MOGAT1 | rs35959734 | G/A (A13T) | 2: 222671822 | 1.2 |
1.11×10−8 | 0.82 | 0.49–1.39 |
| rs6534076 | C/T | 4: 118030971 | 38.4 |
1.38×10−8 | 0.96 | 0.85–1.07 |
| SPATC1L | rs113710653 | C/T (E231K) | 21: 46161921 | 1.9 |
3.75×10−8 | 3.64 | 2.43–5.44 |
| LRRC17 | rs3800939 | A/G (K119E) | 7: 102934268 | 14.0 |
3.90×10−8 | 0.97 | 0.82–1.13 |
| PATE1 | rs2114084 | A/G (Q47R) | 11: 125747715 | 36.4 |
2.19×10−7 | 1.10 | 0.98–1.23 |
| LY6G6C | rs117894946 | G/C (G75A) | 6: 31719250 | 9.5 |
2.77×10−7 | 1.06 | 0.88–1.28 |
| VWA5B1 | rs139281890 | G/A (R92Q) | 1: 20312971 | 0.2 |
3.26×10−7 | 0.44 | 0.06–3.23 |
| PABPC4 | rs4660293 | A/G | 1: 39562508 | 15.2 |
3.99×10−7 | 1.10 | 0.95–1.28 |
| ELMO1 | rs11984075 | A/G | 7: 37397251 | 1.1 |
5.49×10−7 | 1.04 | 0.62–1.74 |
| C2 | rs511294 | A/C | 6: 31921092 | 0.6 |
7.29×10−7 | 1.20 | 0.63–2.30 |
| rs2823962 | G/A | 21: 16673913 | 32.8 |
8.40×10−7 | 0.99 | 0.88–1.11 |
Multivariable logistic regression
analysis of the relation of SNPs to ICH
The relation of the 48 SNPs identified by EWAS to
ICH was examined further by multivariable logistic regression
analysis with adjustment for age, sex and the prevalence of
hypertension. A total of 6 SNPs were related (P<0.05 in at least
one genetic model) to ICH (Table
VI). Among these SNPs, rs138533962 [G/A (R379C)] of
serine/threonine/tyrosine kinase 1 gene (STYK1) (dominant
and additive 1 models) was significantly [P<2.60×10−4
(0.05/192)] associated with ICH, with the minor A allele
representing a risk factor for this condition.
| Table VIRelation of SNPs to intracerebral
hemorrhage as determined by multivariable logistic regression
analysis. |
Table VI
Relation of SNPs to intracerebral
hemorrhage as determined by multivariable logistic regression
analysis.
| SNP | Dominant
| Recessive
| Additive 1
| Additive 2
|
|---|
| P-value | OR (95% CI) | P-value | OR (95% CI) | P-value | OR (95% CI) | P-value | OR (95% CI) |
|---|
| rs3764276 | C/T | 0.0377 | 1.21
(1.01–1.45) | 0.3181 | | 0.0634 | | 0.1001 | |
| rs3820059 | G/A (S172F) | 0.6828 | | 0.0091 | 2.6×10−8
(0–0.55) | 0.4378 | | 0.0095 | 2.6×10−8
(0–5.5×10−7) |
| rs2857697 | A/G | 0.0444 | 1.20
(1.00–1.43) | 0.0176 | 1.36
(1.06–1.74) | 0.1876 | | 0.0073 | 1.46
(1.11–1.90) |
| rs138533962 | G/A (R379C) |
<1.0×10−23 | 111.3
(33.0–694.6) | ND | |
<1.0×10−23 | 111.3
(33.0–694.6) | ND | |
| rs113710653 | C/T (E231K) | 0.0020 | 2.35
(1.39–3.78) | 0.6760 | | 0.0018 | 2.38
(1.41–3.83) | 0.6804 | |
| rs3800939 | A/G (K119E) | 0.8551 | | 0.0414 | 0.44
(0.15–0.97) | 0.5338 | | 0.0466 | 0.44
(0.16–0.99) |
EWAS for SAH
The characteristics of the subjects in the EWAS for
SAH are shown in Table IV. The
prevalence of hypertension was greater, whereas that of
dyslipidemia was lower, in the subjects with SAH than in the
controls.
We examined the relation of allele frequencies for
41,332 SNPs to SAH with Fisher's exact test. A Manhattan plot for
EWAS for SAH is shown in Fig. 3B.
After Bonferroni's correction, 12 SNPs were significantly
(P<1.21×10−6) associated with SAH (Table VII). The genotype distributions
of these SNPs were in Hardy-Weinberg equilibrium (P>0.001) both
among subjects with SAH and among controls (data not shown).
| Table VIIThe 12 SNPs significantly
(P<1.21×10−6) associated with subarachnoid hemorrhage
in the exome-wide association study. |
Table VII
The 12 SNPs significantly
(P<1.21×10−6) associated with subarachnoid hemorrhage
in the exome-wide association study.
| Gene | dbSNP | Nucleotide (amino
acid) substitutiona | Chromosome:
position | MAF (%) | P-value
(allele) | Allele OR | 95% CI |
|---|
| CDL6 | rs2972146 | A/C | 2: 226235982 | 8.7 |
1.28×10−18 | 0.98 | 0.72–1.33 |
| rs3135365 | T/G | 6: 32421478 | 18.9 |
9.71×10−15 | 0.86 | 0.68–1.09 |
| rs2282978 | T/C | 7: 92635096 | 10.6 |
3.91×10−12 | 1.23 | 0.95–1.59 |
| rs2639889 | A/G | 16: 61089243 | 32.3 |
4.23×10−11 | 0.88 | 0.72–1.06 |
| ANKFN1 | rs12449568 | T/C | 17: 56352794 | 42.0 |
3.64×10−10 | 0.92 | 0.77–1.09 |
| CHRDL2 | rs79893604 | G/A (P395L) | 11: 74697229 | 0.8 |
5.70×10−10 | 2.45 | 1.23–4.76 |
| CTNNA3 | rs12256826 | C/T | 10: 66214832 | 13.6 |
8.96×10−10 | 0.83 | 0.63–1.09 |
| SLC4A5 | rs10177833 | A/C | 2: 74230591 | 46.8 |
7.03×10−9 | 0.99 | 0.84–1.18 |
| TEX41 | rs2381683 | A/G | 2: 144981989 | 2.7 |
1.15×10−8 | 1.14 | 0.67–1.91 |
| PYGM | rs589691 | T/C | 11: 64757744 | 42.2 |
1.25×10−8 | 1.06 | 0.89–1.27 |
| rs34429154 | A/C | 1: 4303675 | 40.2 |
1.47×10−8 | 1.04 | 0.87–1.24 |
| COL17A1 | rs117564807 | C/T (D919N) | 10: 104040357 | 1.1 |
4.83×10−7 | 0.00 | ND |
Multivariable logistic regression
analysis of the relation of SNPs to SAH
The relation of the 12 SNPs identified by EWAS to
SAH was examined further by multivariable logistic regression
analysis with adjustment for age, sex and the prevalence of
hypertension. Three SNPs were related (P<0.05) to SAH (Table VIII). Among these SNPs,
rs117564807 [C/T (D919N)] of collagen type XVII α1 chain gene
(COL17A1) (dominant and additive 1 models) was significantly
[P<0.0010 (0.05/48)] associated with SAH, with the minor T
allele being protective against this condition.
| Table VIIIRelation of SNPs to subarachnoid
hemorrhage as determined by multivariable logistic regression
analysis. |
Table VIII
Relation of SNPs to subarachnoid
hemorrhage as determined by multivariable logistic regression
analysis.
| SNP | Dominant
| Recessive
| Additive 1
| Additive 2
|
|---|
| P-value | OR (95% CI) | P-value | OR (95% CI) | P-value | OR (95% CI) | P-value | OR (95% CI) |
|---|
| rs2639889 | A/G | 0.0203 | 0.74
(0.58–0.96) | 0.9267 | | 0.0129 | 0.71
(0.55–0.93) | 0.5252 | |
| rs12256826 | C/T | 0.5019 | | 0.0024 | 2.2×10−8
(0–6.1×10−4) | 0.8687 | | 0.0024 | 2.2×10−8
(0–1.1×10−4) |
| rs117564807 | C/T (D919N) | 0.0009 | 2.2×10−8
(0–3.7×10−4) | 0.6615 | | 0.0009 | 2.2×10−8
(0–1.1×10−4) | 0.6583 | |
Relation of SNPs to intermediate
phenotypes
We examined the relation of 6 SNPs (rs3212335 of
GABRB3, rs147783135 of TMPRSS7, rs2292661 of
PDIA5, rs191885206 of CYP4F12, rs138533962 of
STYK1 and rs117564807 of COL17A1) to intermediate
phenotypes of ischemic or hemorrhagic stroke, including
hypertension, diabetes mellitus, hypertriglyceridemia,
hypo-HDL-cholesterolemia, hyper-LDL-cholesterolemia, chronic kidney
disease, obesity, and hyperuricemia. The rs138533962 SNP of
STYK1 was significantly [P<0.0010 (0.05/48)] associated
with the prevalence of hypertension, diabetes mellitus,
hypertriglyceridemia and hypo-HDL-cholesterolemia, whereas the
other 5 SNPs were not related to any of these intermediate
phenotypes (Table IX).
| Table IXRelation of SNPs to intermediate
phenotypes of ischemic or hemorrhagic stroke. |
Table IX
Relation of SNPs to intermediate
phenotypes of ischemic or hemorrhagic stroke.
| SNP | | Hypertension | DM | Hyper-TG | Hypo-HDL | Hyper-LDL | CKD | Obesity | HU |
|---|
| Related to ischemic
stroke | | | | | | | | |
| rs3212335 | G/A | 0.1945 | 0.7378 | 0.3226 | 0.3391 | 0.2901 | 0.8774 | 0.1276 | 0.0084 |
| rs147783135 | C/T
(R692*) | 0.5725 | 0.0957 | 0.6348 | 0.9054 | 0.4731 | 0.6812 | 0.5797 | 0.8922 |
| rs2292661 | C/T (T391M) | 0.2069 | 0.2699 | 0.9279 | 0.8970 | 0.4782 | 0.2028 | 0.7069 | 0.3671 |
| rs191885206 | T/C (C402R) | 1.0000 | 0.8703 | 0.1862 | 0.4469 | 0.5154 | 0.8764 | 1.0000 | 0.1129 |
| Related to ICH | | | | | | | | | |
| rs138533962 | G/A (R379C) |
4.55×10−13 |
7.12×10−5 | 0.0002 |
1.81×10−13 | 0.0765 | 1.0000 | 0.5188 | 1.0000 |
| Related to SAH | | | | | | | | | |
| rs117564807 | C/T (D919N) | 0.7052 | 0.5715 | 0.2072 | 0.2934 | 0.1943 | 0.1056 | 0.5019 | 0.1484 |
Relation of genes and SNPs identified in
the present study to phenotypes previously examined in GWASs
We examined the 6 genes and SNPs identified in the
present study to phenotypes previously examined by GWASs available
in public databases [GWAS Catalog (http://www.ebi.ac.uk/gwas) and GWAS Central
(http://www.gwascentral.org/browser)].
None of these genes or SNPs was found to be associated with
ischemic or hemorrhagic stroke in previous GWASs (data not
shown).
Discussion
The pathogenesis of ischemic stroke is predominantly
attributed to atherothrombosis of the cerebral arteries, with its
treatable risk factors including hypertension, diabetes mellitus
and chronic kidney disease (1).
In addition to these conventional risk factors, genetic variants
are important in the development of ischemic stroke (2). ICH is typically a manifestation of
underlying small-vessel disease. Long-standing hypertension can
thus lead to hypertensive vasculopathy and consequent microscopic
degenerative changes in the walls of small to medium penetrating
vessels (13). Cerebral amyloid
angiopathy is characterized by the deposition of the β-amyloid
peptide in the walls of small leptomeningeal and cortical vessels,
which can also ultimately result in degenerative changes in the
vessel wall including loss of smooth muscle cells, wall thickening,
luminal narrowing, microaneurysm formation and microhemorrhages
(14–17). Spontaneous SAH usually results
from a ruptured intracranial aneurysm. Blood in the subarachnoid
space gives rise to chemical meningitis that commonly increases
intracranial pressure for days or a few weeks. Secondary vasospasm
may then cause focal brain ischemia. Brain edema accelerates
vasospasm and subsequent infarction (21,22). Given the serious nature of both
ischemic and hemorrhagic stroke, prediction of the risk for these
conditions on the basis of genetic variants would be of benefit for
decision-making with regard to how aggressively to target the
clinical risk factors that are currently amenable to treatment.
In this study, we demonstrated that rs3212335 (G/A)
of GABRB3, rs147783135 [C/T (R692*)] of
TMPRSS7, rs2292661 [C/T (T391M)] of PDIA5 and
rs191885206 [T/C (C402R)] of CYP4F12 were related to
ischemic stroke in Japanese individuals. The minor A allele of
rs3212335 and the C allele of rs191885206 were risk factors for
ischemic stroke, whereas the minor T alleles of rs147783135 and
rs2292661 were protective against this condition. We also found
that rs138533962 [G/A (R379C)] of STYK1 was significantly
associated with ICH, with the minor A allele representing a risk
factor for ICH, and that rs117564807 [C/T (D919N)] of
COL17A1 was significantly associated with SAH, with the
minor T allele being protective against this condition.
SNPs associated with ischemic stroke
The GABRB3 gene is located at chromosomal
region 15q12 (NCBI Gene, https://www.ncbi.nlm.nih.gov/gene) and is highly
expressed in the brain (The Human Protein Atlas, http://www.proteinatlas.org). The GABRB3 protein is a
component of a multisub-unit Cl− channel that serves as
a receptor for γ-aminobutyric acid (GABA), a major inhibitory
neurotransmitter of the mammalian nervous system (41). Mutations of GABRB3 have
been associated with several disorders, including Angelman syndrome
(42), Prader-Willi syndrome
(43), non-syndromic orofacial
clefts (44), epilepsy (45) and autism (46). In this study, we demonstrated that
rs3212335 (G/A) of GABRB3 was related to ischemic stroke,
with the minor A allele being a risk factor for this condition,
although the molecular mechanism underlying this association
remains unclear.
TMPRSS7 is located at chromosomal region
3q13.2 (NCBI Gene) and is highly expressed in the testis (The Human
Protein Atlas). The TMPRSS7 protein belongs to the type II
transmembrane serine protease (TTSP) family, the 17 human members
of which degrade components of the extracellular matrix (47) and play physiological and
pathological roles in digestion, cardiac function, blood pressure
regulation, hearing, iron metabolism, and epithelial homeostasis
(48,49). They have also been implicated in
tumor growth, invasion and metastasis (48,49), and the genetic variants of
TMPRSS7 have been associated with the risk for and prognosis
of breast cancer (50). In this
study, we demonstrated that rs147783135 [C/T (R692*)] of
TMPRSS7 was related to ischemic stroke, with the minor T
allele being protective against this condition. Given the potential
roles of TMPRSS7 in tumor growth and blood pressure
regulation (48,49) the association of this gene with
ischemic stroke may reflect an effect on atherosclerosis or blood
pressure.
PDIA5 is located at chromosomal region 3q21.1
(NCBI Gene) and is expressed widely including in the brain and
vascular smooth muscle (The Human Protein Atlas). PDIA5
encodes one of the protein disulfide isomerases that catalyze
protein folding and thiol-disulfide interchange reactions in the
endoplasmic reticulum (ER). The PDIA5 protein contains an
NH2-terminal ER signal sequence, three catalytically
active thioredoxin domains, a thioredoxin-like domain, and a
COOH-terminal ER retention sequence. The thioredoxin-like domain is
the primary binding site for the major ER chaperone calreticulin
(51,52). Recent GWASs indicated that a SNP
in PDIA5 was associated with plasma fibrinogen concentration
(53) and platelet count
(54). In this study, we
demonstrated that rs2292661 [C/T (T391M)] of PDIA5 was
related to ischemic stroke, with the minor T allele being
protective against this condition. Given its potential role as a
determinant of fibrinogen concentration and platelet count, both of
which are important in the development of atherosclerotic
thrombosis, the association of PDIA5 with ischemic stroke
may reflect an effect of this gene on arterial thrombosis.
CYP4F12 is located at chromosomal region
19p13.12 (NCBI Gene) and is expressed in various tissues and organs
including the brain and vascular smooth muscle (The Human Protein
Atlas). CYP4F12 encodes a member of the cytochrome P450
superfamily of monooxygenases that catalyze many reactions
including those related to drug metabolism, as well as to the
synthesis of cholesterol, steroids and other lipids. The CYP4F12
protein is likely localized to the ER (55,56). Given that human CYP4F enzymes play
a role in the metabolism of endogenous compounds such as
inflammatory mediators, they likely contribute to regulation of
inflammatory processes (57).
Arachidonic acid and prostaglandin H2 (PGH2)
have been found to serve as substrates of CYP4F12, which also
metabolizes to a lesser extent PGE2, PGF2α
and leukotriene B4 (57,58). In this study, we demonstrated that
rs191885206 [T/C (C402R)] of CYP4F12 was related to ischemic
stroke, with the minor C allele representing a risk factor for this
condition. The association of CYP4F12 with ischemic stroke
may reflect an effect of this gene on vascular inflammation.
A SNP associated with ICH. The STYK1
gene is located at chromosomal region 12p13.2 (NCBI Gene) and is
expressed in various tissues and organs, including the brain (The
Human Protein Atlas). STYK1 plays important roles in diverse
cellular and developmental processes, including cell proliferation,
differentiation and survival (59,60). The upregulation of STYK1
expression has been detected in many types of tumor, including
breast cancer (61), lung cancer
(62), ovarian cancer (63), prostate cancer (64), colorectal cancer (65) and hepatocellular carcinoma
(66). The STYK1 protein has been
shown to promote cell transformation, tumorigenesis and metastasis
by activating the phosphoinositide 3-kinase (PI3K)-AKT signaling
pathway and inactivating glycogen synthase kinase 3β signaling
(67). It has also been found to
enhance angiogenesis and to change the morphology of blood vessels
during tumor growth (68). In
this study, we demonstrated that rs138533962 [G/A (R379C)] of
STYK1 was significantly associated with ICH, with the minor
A allele representing a risk factor for this condition. The
relation of STYK1 to ICH may be attributable to an effect of
this gene on the remodeling of blood vessels in the brain, although
the molecular mechanism underlying this association remains to be
determined.
A SNP associated with SAH. The
COL17A1 gene is located at chromosomal region 10q25.1 (NCBI
Gene) and is highly expressed in skin (The Human Protein Atlas).
Collagen XVII is a homotrimer of three 180-kDa α1 chains, each of
which possesses a long intracellular NH2-terminal
domain, a short transmembrane region, and an extracellular
COOH-terminus. It is a structural component of hemidesmosomes,
which mediate the adhesion of epidermal keratinocytes and certain
other types of epithelial cell to the underlying basement membrane
(69,70). Mutations in COL17A1 that
result in the generation of premature stop codons are associated
with non-Herlitz junctional epidermolysis bullosa (69,70), which is characterized by
generalized blistering of the skin and mucous membranes together
with atrophic scarring and nail dystrophy. The majority of patients
with such mutations lack type XVII collagen mRNA as a result of
nonsense-mediated mRNA decay (71). In addition, an autoimmune response
to collagen XVII is responsible for epidermal impairment in
individuals with bullous pemphigoid (72). A population-based study revealed
an association of bullous pemphigoid with neurological disorders,
in particular stroke, for which the odds ratio was 3.3 (73). This association is likely due to
the fact that the inflammatory state present in individuals with
bullous pemphigoid is not confined to the skin but also involves
the vascular endothelium (74,75). It has been suggested that
intracranial aneurysm is a chronic inflammatory disease at
bifurcation sites of cerebral arterial walls (76). Indeed, many inflammatory cells
have been detected in specimens of surgically dissected
intracranial vessel walls affected by aneurysm (77). Chronic vascular inflammation,
acting together with the structural properties of the cerebral
arterial wall and hemodynamic factors, may therefore accelerate the
development of intracranial aneurysm and subsequent aneurysm
rupture. We have now shown that rs117564807 [C/T (D919N)] of
COL17A1 was significantly associated with SAH, with the
minor T allele being protective against this condition. The
genotype distribution of rs117564807 was 265/265 (100%) CC in
subjects with SAH and 8958/9158 (97.82%) CC, 197/9158 (2.15%) CT,
and 3/9158 (0.03%) TT in control individuals, giving an allele odds
ratio of 0. The association of COL17A1 with SAH may be
attributable to the effect of this gene on cerebrovascular
inflammation, although the molecular mechanism remains to be
determined.
General considerations
In previous meta-analyses of GWASs for ischemic
stroke, the MAF of most SNPs ranged from 6 to 49% and the odds
ratio from 0.8 to 2.0 (4–12,34–36). In our study, we identified four
SNPs related to ischemic stroke, with the allele odds ratio (MAF,
%) of rs3212335 of GABRB3, rs147783135 of TMPRSS7,
rs2292661 of PDIA5, and rs191885206 of CYP4F12 being
1.02 (38.9%), 0.54 (0.8%), 0.47 (0.7%), and 2.11 (0.3%),
respectively. Whereas rs3212335 of GABRB3 was thus a common
variant with a small effect size, the other three SNPs were
low-frequency or rare variants with a moderate effect size.
A meta-analysis of GWASs identified rs11179580 at
12q21.1 and rs156197380 at 1q22 as susceptibility loci for lobar
and nonlobar ICH, respectively, in European ancestry populations
(20). The odds ratio (MAF, %)
was 1.56 (24%) for rs11179580 and 1.44 (32%) for rs156197380.
Previous GWASs (26–31) and a meta-analysis of GWASs
(32) also identified SNPs
associated with intracranial aneurysm. The MAFs of these SNPs
ranged from 15 to 48% and the odds ratios from 0.47 to 2.22.
Another recent GWAS of intracranial aneurysm identified
low-frequency genetic variants with MAFs of 3 to 5% and odds ratios
of ~2.0 (78). We have now
identified two SNPs significantly associated with hemorrhagic
stroke, with the odds ratio (MAF, %) of rs138533962 of STYK1
and rs117564807 of COL17A1 being 111.3 (2.0%) and
2.23×10−8 (1.1%), respectively. Both of these SNPs were
thus low-frequency variants with a large effect size.
There are several limitations to the present study:
i) The inflation factors for ischemic stroke, ICH, and SAH were
relatively high, which may be attributable to the relatively small
numbers of subjects with ischemic and hemorrhagic stroke. Given
that our results were not replicated, they will require validation
in other independent subject panels or in other ethnic groups; ii)
it is possible that rs3212335 of GABRB3, rs147783135 of
TMPRSS7, rs2292661 of PDIA5, rs191885206 of
CYP4F12, rs138533962 of STYK1 or rs117564807 of
COL17A1 is in linkage disequilibrium with other
polymorphisms in the same gene or in other nearby genes that are
actually responsible for the development of ischemic or hemorrhagic
stroke; iii) the functional relevance of these SNPs to the
pathogenesis of ischemic stroke, ICH, or SAH remains to be
elucidated.
In conclusion, our results suggest that rs3212335
(G/A) of GABRB3, rs147783135 [C/T (R692*)] of
TMPRSS7, rs2292661 [C/T (T391M)] of PDIA5, and
rs191885206 [T/C (C402R)] of CYP4F12 may be novel
susceptibility loci for ischemic stroke, whereas rs138533962 [G/A
(R379C)] of STYK1 and rs117564807 [C/T (D919N)] of
COL17A1 may be such loci for ICH and SAH, respectively, in
Japanese individuals. Determination of genotypes for these SNPs may
prove informative for assessment of the genetic risk for ischemic
or hemorrhagic stroke in Japanese individuals.
Acknowledgments
This study was supported by the CREST (grant
number, JPMJCR1302), the Japan Science and Technology Agency (to
Y.Y., J.S. and I.T.) and by the Japan Society for the Promotion of
Science KAKENHI grants JP15H04772 (to Y.Y.), JP25242062 (to M.T.)
and JP16H01872 (to M.T.).
References
|
1
|
Mozaffarian D, Benjamin EJ, Go AS, Arnett
DK, Blaha MJ, Cushman M, Das SR, de Ferranti S, Després JP,
Fullerton HJ, et al Writing Group Members; American Heart
Association Statistics Committee; Stroke Statistics Subcommittee:
Heart Disease and Stroke Statistics-2016 Update: A Report From the
American Heart Association. Circulation. 133:e38–e360. 2016.
View Article : Google Scholar
|
|
2
|
Falcone GJ, Malik R, Dichgans M and Rosand
J: Current concepts and clinical applications of stroke genetics.
Lancet Neurol. 13:405–418. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Bevan S, Traylor M, Adib-Samii P, Malik R,
Paul NL, Jackson C, Farrall M, Rothwell PM, Sudlow C, Dichgans M,
et al: Genetic heritability of ischemic stroke and the contribution
of previously reported candidate gene and genomewide associations.
Stroke. 43:3161–3167. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Gretarsdottir S, Thorleifsson G, Manolescu
A, Styrkarsdottir U, Helgadottir A, Gschwendtner A, Kostulas K,
Kuhlenbäumer G, Bevan S, Jonsdottir T, et al: Risk variants for
atrial fibrillation on chromosome 4q25 associate with ischemic
stroke. Ann Neurol. 64:402–409. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Gudbjartsson DF, Holm H, Gretarsdottir S,
Thorleifsson G, Walters GB, Thorgeirsson G, Gulcher J, Mathiesen
EB, Njølstad I, Nyrnes A, et al: A sequence variant in ZFHX3 on
16q22 associates with atrial fibrillation and ischemic stroke. Nat
Genet. 41:876–878. 2009. View
Article : Google Scholar : PubMed/NCBI
|
|
6
|
Ikram MA, Seshadri S, Bis JC, Fornage M,
DeStefano AL, Aulchenko YS, Debette S, Lumley T, Folsom AR, van den
Herik EG, et al: Genomewide association studies of stroke. N Engl J
Med. 360:1718–1728. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Holliday EG, Maguire JM, Evans TJ, Koblar
SA, Jannes J, Sturm JW, Hankey GJ, Baker R, Golledge J, Parsons MW,
et al Australian Stroke Genetics Collaborative; International
Stroke Genetics Consortium; Wellcome Trust Case Control Consortium
2: Common variants at 6p21.1 are associated with large artery
atherosclerotic stroke. Nat Genet. 44:1147–1151. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Bellenguez C, Bevan S, Gschwendtner A,
Spencer CC, Burgess AI, Pirinen M, Jackson CA, Traylor M, Strange
A, Su Z, et al International Stroke Genetics Consortium (ISGC);
Wellcome Trust Case Control Consortium 2 (WTCCC2): Genome-wide
association study identifies a variant in HDAC9 associated with
large vessel ischemic stroke. Nat Genet. 44:328–333. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Traylor M, Farrall M, Holliday EG, Sudlow
C, Hopewell JC, Cheng YC, Fornage M, Ikram MA, Malik R, Bevan S, et
al Australian Stroke Genetics Collaborative, Wellcome Trust Case
Control Consortium 2 (WTCCC2); International Stroke Genetics
Consortium: Genetic risk factors for ischaemic stroke and its
subtypes (the METASTROKE collaboration): A meta-analysis of
genome-wide association studies. Lancet Neurol. 11:951–962. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Kilarski LL, Achterberg S, Devan WJ,
Traylor M, Malik R, Lindgren A, Pare G, Sharma P, Slowik A, Thijs
V, et al GARNET Collaborative Research Group, Wellcome Trust Case
Control Consortium 2; Australian Stroke Genetic Collaborative; the
METASTROKE Consortium; the International Stroke Genetics
Consortium: Meta-analysis in more than 17,900 cases of ischemic
stroke reveals a novel association at 12q24.12. Neurology.
83:678–685. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Neurology Working Group of the Cohorts for
Heart and Aging Research in Genomic Epidemiology (CHARGE)
Consortium; Stroke Genetics Network (SiGN); International Stroke
Genetics Consortium (ISGC): Identification of additional risk loci
for stroke and small vessel disease: a meta-analysis of genome-wide
association studies. Lancet Neurol. 15:695–707. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
NINDS Stroke Genetics Network:
International Stroke Genetics Consortium (ISGC): Loci associated
with ischaemic stroke and its subtypes (SiGN): A genome-wide
association study. Lancet Neurol. 15:174–184. 2016. View Article : Google Scholar
|
|
13
|
Qureshi AI, Tuhrim S, Broderick JP, Batjer
HH, Hondo H and Hanley DF: Spontaneous intracerebral hemorrhage. N
Engl J Med. 344:1450–1460. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Sacco RL: Lobar intracerebral hemorrhage.
N Engl J Med. 342:276–279. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
O'Donnell HC, Rosand J, Knudsen KA, Furie
KL, Segal AZ, Chiu RI, Ikeda D and Greenberg SM: Apolipoprotein E
genotype and the risk of recurrent lobar intracerebral hemorrhage.
N Engl J Med. 342:240–245. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Woo D, Kaushal R, Chakraborty R, Woo J,
Haverbusch M, Sekar P, Kissela B, Pancioli A, Jauch E, Kleindorfer
D, et al: Association of apolipoprotein E4 and haplotypes of the
apolipo-protein E gene with lobar intracerebral hemorrhage. Stroke.
36:1874–1879. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Greenberg SM, Vonsattel JP, Segal AZ, Chiu
RI, Clatworthy AE, Liao A, Hyman BT and Rebeck GW: Association of
apolipoprotein E epsilon2 and vasculopathy in cerebral amyloid
angiopathy. Neurology. 50:961–965. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Devan WJ, Falcone GJ, Anderson CD,
Jagiella JM, Schmidt H, Hansen BM, Jimenez-Conde J,
Giralt-Steinhauer E, Cuadrado-Godia E, Soriano C, et al
International Stroke Genetics Consortium: Heritability estimates
identify a substantial genetic contribution to risk and outcome of
intracerebral hemorrhage. Stroke. 44:1578–1583. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Falcone GJ, Biffi A, Devan WJ, Jagiella
JM, Schmidt H, Kissela B, Hansen BM, Jimenez-Conde J,
Giralt-Steinhauer E, Elosua R, et al International Stroke Genetics
Consortium: Burden of risk alleles for hypertension increases risk
of intracerebral hemorrhage. Stroke. 43:2877–2883. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Woo D, Falcone GJ, Devan WJ, Brown WM,
Biffi A, Howard TD, Anderson CD, Brouwers HB, Valant V, Battey TW,
et al International Stroke Genetics Consortium: Meta-analysis of
genome-wide association studies identifies 1q22 as a susceptibility
locus for intracerebral hemorrhage. Am J Hum Genet. 94:511–521.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Macdonald RL and Schweizer TA: Spontaneous
subarachnoid haemorrhage. Lancet. 389:655–666. 2017. View Article : Google Scholar
|
|
22
|
van Gijn J, Kerr RS and Rinkel GJ:
Subarachnoid haemorrhage. Lancet. 369:306–318. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Linn FHH, Rinkel GJE, Algra A and van Gijn
J: Incidence of subarachnoid hemorrhage: role of region, year, and
rate of computed tomography: A meta-analysis. Stroke. 27:625–629.
1996. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Johnston SC, Selvin S and Gress DR: The
burden, trends, and demographics of mortality from subarachnoid
hemorrhage. Neurology. 50:1413–1418. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Tromp G, Weinsheimer S, Ronkainen A and
Kuivaniemi H: Molecular basis and genetic predisposition to
intracranial aneurysm. Ann Med. 46:597–606. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Bilguvar K, Yasuno K, Niemelä M, Ruigrok
YM, von Und Zu Fraunberg M, van Duijn CM, van den Berg LH, Mane S,
Mason CE, Choi M, et al: Susceptibility loci for intracranial
aneurysm in European and Japanese populations. Nat Genet.
40:1472–1477. 2008. View
Article : Google Scholar : PubMed/NCBI
|
|
27
|
Yasuno K, Bilguvar K, Bijlenga P, Low SK,
Krischek B, Auburger G, Simon M, Krex D, Arlier Z, Nayak N, et al:
Genome-wide association study of intracranial aneurysm identifies
three new risk loci. Nat Genet. 42:420–425. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Yasuno K, Bakırcıoğlu M, Low SK, Bilgüvar
K, Gaál E, Ruigrok YM, Niemelä M, Hata A, Bijlenga P, Kasuya H, et
al: Common variant near the endothelin receptor type A (EDNRA) gene
is associated with intracranial aneurysm risk. Proc Natl Acad Sci
USA. 108:19707–19712. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Foroud T, Koller DL, Lai D, Sauerbeck L,
Anderson C, Ko N, Deka R, Mosley TH, Fornage M, Woo D, et al FIA
Study Investigators: Genome-wide association study of intracranial
aneurysms confirms role of Anril and SOX17 in disease risk. Stroke.
43:2846–2852. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Low SK, Takahashi A, Cha PC, Zembutsu H,
Kamatani N, Kubo M and Nakamura Y: Genome-wide association study
for intracranial aneurysm in the Japanese population identifies
three candidate susceptible loci and a functional genetic variant
at EDNRA. Hum Mol Genet. 21:2102–2110. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Foroud T, Lai D, Koller D, Van't Hof F,
Kurki MI, Anderson CS, Brown RD Jr, Connolly ES, Eriksson JG,
Flaherty M, et al Familial Intracranial Aneurysm Study
Investigators: Genome-wide association study of intracranial
aneurysm identifies a new association on chromosome 7. Stroke.
45:3194–3199. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Alg VS, Sofat R, Houlden H and Werring DJ:
Genetic risk factors for intracranial aneurysms: A meta-analysis in
more than 116000 individuals. Neurology. 80:2154–2165. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Manolio TA, Collins FS, Cox NJ, Goldstein
DB, Hindorff LA, Hunter DJ, McCarthy MI, Ramos EM, Cardon LR,
Chakravarti A, et al: Finding the missing heritability of complex
diseases. Nature. 461:747–753. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Kubo M, Hata J, Ninomiya T, Matsuda K,
Yonemoto K, Nakano T, Matsushita T, Yamazaki K, Ohnishi Y, Saito S,
et al: A nonsynonymous SNP in PRKCH (protein kinase C eta)
increases the risk of cerebral infarction. Nat Genet. 39:212–217.
2007. View
Article : Google Scholar : PubMed/NCBI
|
|
35
|
Hata J, Matsuda K, Ninomiya T, Yonemoto K,
Matsushita T, Ohnishi Y, Saito S, Kitazono T, Ibayashi S, Iida M,
et al: Functional SNP in an Sp1-binding site of AGTRL1 gene is
associated with susceptibility to brain infarction. Hum Mol Genet.
16:630–639. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Matsushita T, Ashikawa K, Yonemoto K,
Hirakawa Y, Hata J, Amitani H, Doi Y, Ninomiya T, Kitazono T,
Ibayashi S, et al: Functional SNP of ARHGEF10 confers risk of
atherothrombotic stroke. Hum Mol Genet. 19:1137–1146. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Sacco RL, Kasner SE, Broderick JP, Caplan
LR, Connors JJ, Culebras A, Elkind MS, George MG, Hamdan AD,
Higashida RT, et al American Heart Association Stroke Council;
Council on Cardiovascular Surgery and Anesthesia; Council on
Cardiovascular Radiology and Intervention; Council on
Cardiovascular and Stroke Nursing; Council on Epidemiology and
Prevention; Council on Peripheral Vascular Disease; Council on
Nutrition; Physical Activity and Metabolism: An updated definition
of stroke for the 21st century: A statement for healthcare
professionals from the American Heart Association/American Stroke
Association. Stroke. 44:2064–2089. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Grove ML, Yu B, Cochran BJ, Haritunians T,
Bis JC, Taylor KD, Hansen M, Borecki IB, Cupples LA and Fornage M:
Best practices and joint calling of the HumanExome BeadChip: The
CHARGE Consortium. PLoS One. 8:e680952013. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Anderson CA, Pettersson FH, Clarke GM,
Cardon LR, Morris AP and Zondervan KT: Data quality control in
genetic case-control association studies. Nat Protoc. 5:1564–1573.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Price AL, Patterson NJ, Plenge RM,
Weinblatt ME, Shadick NA and Reich D: Principal components analysis
corrects for stratification in genome-wide association studies. Nat
Genet. 38:904–909. 2006. View
Article : Google Scholar : PubMed/NCBI
|
|
41
|
Kang JQ and Barnes G: A common
susceptibility factor of both autism and epilepsy: Functional
deficiency of GABA A receptors. J Autism Dev Disord. 43:68–79.
2013. View Article : Google Scholar
|
|
42
|
Sinnett D, Wagstaff J, Glatt K, Woolf E,
Kirkness EJ and Lalande M: High-resolution mapping of the
gamma-aminobutyric acid receptor subunit beta 3 and alpha 5 gene
cluster on chromosome 15q11–q13, and localization of breakpoints in
two Angelman syndrome patients. Am J Hum Genet. 52:1216–1229.
1993.PubMed/NCBI
|
|
43
|
Glatt K, Sinnett D and Lalande M: The
human gamma-amino-butyric acid receptor subunit beta 3 and alpha 5
gene cluster in chromosome 15q11–q13 is rich in highly polymorphic
(CA)n repeats. Genomics. 19:157–160. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Scapoli L, Martinelli M, Pezzetti F,
Carinci F, Bodo M, Tognon M and Carinci P: Linkage disequilibrium
between GABRB3 gene and nonsyndromic familial cleft lip with or
without cleft palate. Hum Genet. 110:15–20. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Macdonald RL, Kang JQ and Gallagher MJ:
Mutations in GABAA receptor subunits associated with genetic
epilepsies. J Physiol. 588:1861–1869. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Delahanty RJ, Kang JQ, Brune CW, Kistner
EO, Courchesne E, Cox NJ, Cook EH Jr, Macdonald RL and Sutcliffe
JS: Maternal transmission of a rare GABRB3 signal peptide variant
is associated with autism. Mol Psychiatry. 16:86–96. 2011.
View Article : Google Scholar
|
|
47
|
Hooper JD, Clements JA, Quigley JP and
Antalis TM: Type II transmembrane serine proteases. Insights into
an emerging class of cell surface proteolytic enzymes. J Biol Chem.
276:857–860. 2001. View Article : Google Scholar
|
|
48
|
Bugge TH, Antalis TM and Wu Q: Type II
transmembrane serine proteases. J Biol Chem. 284:23177–23181. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Antalis TM, Buzza MS, Hodge KM, Hooper JD
and Netzel-Arnett S: The cutting edge: Membrane-anchored serine
protease activities in the pericellular microenvironment. Biochem
J. 428:325–346. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Luostari K, Hartikainen JM, Tengström M,
Palvimo JJ, Kataja V, Mannermaa A and Kosma VM: Type II
transmembrane serine protease gene variants associate with breast
cancer. PLoS One. 9:e1025192014. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Hayano T and Kikuchi M: Molecular cloning
of the cDNA encoding a novel protein disulfide isomerase-related
protein (PDIR). FEBS Lett. 372:210–214. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Vinaik R, Kozlov G and Gehring K:
Structure of the non-catalytic domain of the protein disulfide
isomerase-related protein (PDIR) reveals function in protein
binding. PLoS One. 8:e620212013. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
de Vries PS, Chasman DI, Sabater-Lleal M,
Chen MH, Huffman JE, Steri M, Tang W, Teumer A, Marioni RE,
Grossmann V, et al: A meta-analysis of 120 246 individuals
identifies 18 new loci for fibrinogen concentration. Hum Mol Genet.
25:358–370. 2016. View Article : Google Scholar :
|
|
54
|
Gieger C, Radhakrishnan A, Cvejic A, Tang
W, Porcu E, Pistis G, Serbanovic-Canic J, Elling U, Goodall AH,
Labrune Y, et al: New gene functions in megakaryopoiesis and
platelet formation. Nature. 480:201–208. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Bylund J, Bylund M and Oliw EH: cDna
cloning and expression of CYP4F12, a novel human cytochrome 450.
Biochem Biophys Res Commun. 280:892–897. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Hashizume T, Imaoka S, Hiroi T, Terauchi
Y, Fujii T, Miyazaki H, Kamataki T and Funae Y: cDNA cloning and
expression of a novel cytochrome 450 (cyp4f12) from human small
intestine. Biochem Biophys Res Commun. 280:1135–1141. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Kikuta Y, Kusunose E and Kusunose M:
Prostaglandin and leukotriene omega-hydroxylases. Prostaglandins
Other Lipid Mediat. 68–69:345–362. 2002. View Article : Google Scholar
|
|
58
|
Cauffiez C, Klinzig F, Rat E, Tournel G,
Allorge D, Chevalier D, Pottier N, Lovecchio T, Colombel JF,
Lhermitte M, et al: Human CYP4F12 genetic polymorphism:
Identification and functional characterization of seven variant
allozymes. Biochem Pharmacol. 68:2417–2425. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Ye X, Ji C, Huang Q, Cheng C, Tang R, Xu
J, Zeng L, Dai J, Wu Q, Gu S, et al: Isolation and characterization
of a human putative receptor protein kinase cDNA STYK1. Mol Biol
Rep. 30:91–96. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Liu L, Yu XZ, Li TS, Song LX, Chen PL, Suo
TL, Li YH, Wang SD, Chen Y, Ren YM, et al: A novel protein tyrosine
kinase NOK that shares homology with platelet-derived growth
factor/fibroblast growth factor receptors induces tumorigenesis and
metastasis in nude mice. Cancer Res. 64:3491–3499. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Moriai R, Kobayashi D, Amachika T, Tsuji N
and Watanabe N: Diagnostic relevance of overexpressed NOK mRNA in
breast cancer. Anticancer Res. 26:4969–4973. 2006.
|
|
62
|
Amachika T, Kobayashi D, Moriai R, Tsuji N
and Watanabe N: Diagnostic relevance of overexpressed mRNA of novel
oncogene with kinase-domain (NOK) in lung cancers. Lung Cancer.
56:337–340. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Jackson KA, Oprea G, Handy J and Kimbro
KS: Aberrant STYK1 expression in ovarian cancer tissues and cell
lines. J Ovarian Res. 2:152009. View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Chung S, Tamura K, Furihata M, Uemura M,
Daigo Y, Nasu Y, Miki T, Shuin T, Fujioka T, Nakamura Y, et al:
Overexpression of the potential kinase serine/threonine/tyrosine
kinase 1 (STYK1) in castration-resistant prostate cancer. Cancer
Sci. 100:2109–2114. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Hu L, Chen HY, Cai J, Zhang Y, Qi CY, Gong
H, Zhai YX, Fu H, Yang GZ and Gao CF: Serine threonine tyrosine
kinase 1 is a potential prognostic marker in colorectal cancer. BMC
Cancer. 15:2462015. View Article : Google Scholar : PubMed/NCBI
|
|
66
|
Wang Z, Qu L, Deng B, Sun X, Wu S, Liao J,
Fan J and Peng Z: STYK1 promotes epithelial-mesenchymal transition
and tumor metastasis in human hepatocellular carcinoma through
MEK/ERK and PI3K/AKT signaling. Sci Rep. 6:332052016. View Article : Google Scholar : PubMed/NCBI
|
|
67
|
Li J, Wu F, Sheng F, Li YJ, Jin D and
Dingxand Zhang S: NOK/STYK1 interacts with GSK-3β and mediates Ser9
phosphorylation through activated Akt. FEBS Lett. 586:3787–3792.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
68
|
Liu Y, Li T, Hu D and Zhang S: NOK/STYK1
promotes the genesis and remodeling of blood and lymphatic vessels
during tumor progression. Biochem Biophys Res Commun. 478:254–259.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
69
|
Giudice GJ, Emery DJ and Diaz LA: Cloning
and primary structural analysis of the bullous pemphigoid
autoantigen BP180. J Invest Dermatol. 99:243–250. 1992. View Article : Google Scholar : PubMed/NCBI
|
|
70
|
Pulkkinen L and Uitto J and Uitto J:
Mutation analysis and molecular genetics of epidermolysis bullosa.
Matrix Biol. 18:29–42. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
71
|
Bauer JW and Lanschuetzer C: Type XVII
collagen gene mutations in junctional epidermolysis bullosa and
prospects for gene therapy. Clin Exp Dermatol. 28:53–60. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
72
|
Kasperkiewicz M, Zillikens D and Schmidt
E: Pemphigoid diseases: Pathogenesis, diagnosis, and treatment.
Autoimmunity. 45:55–70. 2012. View Article : Google Scholar
|
|
73
|
Chen YJ, Wu CY, Lin MW, Chen TJ, Liao KK,
Chen YC, Hwang CY, Chu SY, Chen CC, Lee DD, et al: Comorbidity
profiles among patients with bullous pemphigoid: A nationwide
population-based study. Br J Dermatol. 165:593–599. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
74
|
Ameglio F, D'Auria L, Cordiali-Fei P,
Mussi A, Valenzano L, D'Agosto G, Ferraro C, Bonifati C and
Giacalone B: Bullous pemphigoid and pemphigus vulgaris: Correlated
behaviour of serum VEGF, sE-selectin and TNF-alpha levels. J Biol
Regul Homeost Agents. 11:148–153. 1997.
|
|
75
|
Seppänen A: Collagen XVII: A shared
antigen in neurodermatological interactions? Clin Dev Immunol.
2013:2405702013. View Article : Google Scholar : PubMed/NCBI
|
|
76
|
Aoki T and Nishimura M: Molecular
mechanism of cerebral aneurysm formation focusing on NF-κB as a key
mediator of inflammation. J Biorheol. 24:16–21. 2010. View Article : Google Scholar
|
|
77
|
Chyatte D, Bruno G, Desai S and Todor DR:
Inflammation and intracranial aneurysms. Neurosurgery.
45:1137–1147. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
78
|
Kurki MI, Gaál EI, Kettunen J, Lappalainen
T, Menelaou A, Anttila V, van't Hof FN, von Und Zu Fraunberg M,
Helisalmi S and Hiltunen M: High risk population isolate reveals
low frequency variants predisposing to intracranial aneurysms. PLoS
Genet. 10:e10041342014. View Article : Google Scholar : PubMed/NCBI
|