Introduction
According to the Centers for Disease Control and
Prevention (CDC) in the USA, malignant melanoma is the most
aggressive and lethal skin cancer (1,2).
Its incidence is increasing (3).
Patients with metastatic melanoma have an extremely poor prognosis
(4). Recent studies have reported
that the abnormal activation of pathways, such as WNT,
mitogen-activated protein kinase (MAPK) and phosphoinositide
3-kinase (PI3K), maybe crucial events responsible for the invasion
and metastasis of melanoma (5–8).
NF-κB is an important signaling pathway which is highly expressed
in a number of tumors. Hu et al proved that NF-κB inhibitor
could inhibit the migration of human uveal melanoma cells (9). Wu et al also found s low
expression of NF-κB p65 protein accompanied by the decreased
migration and invasion of A375.S2 cells (10). These studies show that the
expression of NF-κB in melanoma is closely related to the migration
and invasion of tumors. The SOX4 gene, a transcription factor and s
member of the SOX family, regulates transcription through numerous
methods and mediates both gene activation and repression. It is
overexpressed in a wide variety of malignancies, such as lung
cancer (11), breast cancer
(12,13) and prostate cancer (14), and is closely associated with
cancer migration and invasion. However, in a limited subset of
tumors, SOX4 has been reported to act as a tumor suppressor
(15). Some studies have shown
that the increased expression of SOX4 correlates with prolonged
patient survival and slower cancer invasion and metastasis,
including bladder cancer, melanoma and gallbladder cancer (16–18). Over the past 10 years, scholars
have focused on the role of SOX9 and SOX10 in melanoma. It is not
surprising that both SOX9 and SOX10 are expressed during various
stages of melanoma progression and in established melanoma cell
lines (19).
However, few scholars have investigated the
association between SOX4 and melanoma. In this study, we examined
SOX4 expression in melanoma. The expression of SOX4 was knocked
down or upregulated by small interfering RNA (siRNA) or lentivirus
transfection, respectively to examine the effects of SOX4 on the
invasion and migration of melanoma cells. We also aimed to
elucidate the underlying mechanisms. We found that SOX4 promoted
melanoma cell migration and invasion, and these that these effects
were partly mediated though the activation of the nuclear factor-κB
(NF-κB)/p65 signaling pathway.
Materials and methods
Cell culture, siRNA and lentivirus
transfection
The human melanoma cell lines. A375, A875 and A2058.
were purchased from the Cell Bank of the Chinese Academy of
Sciences (Shanghai, China). All cells were cultured in DMEM
supplemented with 10% fetal bovine serum (FBS) and 1%
penicillin/streptomycin (all from Sigma, Poznan, Poland) at 37°C in
a 5% CO2 incubator. SOX4-specific siRNA (si-SOX4;
sc-38412) was purchased from Santa Cruz Biotechnology, Inc. (Santa
Cruz, CA, USA) and si-SOX4 was transfected into the cells at a
final concentration of 5 or 10 nM using Lipofectamine 2000 reagent
(Invitrogen, Grand Island, NY, USA) according to the manufacturer's
instructions. Scrambled siRNA (sc-37007; Santa Cruz Biotechnology,
Inc.) was used as a negative control. The cells were harvested at
24 or 48 h following transfection. The melanoma cells
(8×104/well) were plated into a 6-well plate and
incubated overnight. The medium was changed with FBS-free medium
containing 6 μg/ml polybrene and SOX4 shRNA viral or
overexpression lentivirus (both from GeneChem, Shanghai, China). A
blank vector lentivirus was used for the control group. After 24 h,
fresh medium with 10% FBS was added to the cells and the cells were
incubated for an additional 48 h. SOX4 expression in cells
following transfection was confirmed by western blot analysis and
real-time PCR.
In this study, we used both siRNA and shRNA. This
was done as the use of siRNA in some cases can lead to mismatches
with the RNA target, and the knockdown of genes other than the
intended target may make the results difficult to interpret. In
order to eliminate the off-target effects of siRNA, we also used
sox4 shRNA viral transfection.
Western blot analysis
For total protein extraction, the A375, A875 and
A2058 cells were lysed in RIPA buffer (Beyotime, Jiangsu, China)
for 15 min and centrifuged for 10 min at 12,000 rpm. The
supernatant was harvested and stored at −80°C. For nuclear protein
extraction, the A2058 cells were suspended in hypotonic buffer (20
mM Tris-HCL pH 7.9, 1.5 mM MgCl2 and 10 mM KCL).
Following incubation on ice for 15 min, 12 μl 10% (v/v)
NP-40 were added, and the combination was vortexed for 10 sec and
kept on ice again for a further 10 min, followed by centrifugation
at 14,000 rpm at 4°C for 1 min. The supernatants, which were
considered to be the cytoplasmic extracts, were harvested. The
pellets were washed once with 100 μl hypotonic buffer, and
resus-pended in high salt buffer (20 mM Tris-HCL pH 7.9, 25%
glycerol, 0.42 M NaCl, 1.5 mM MgCl2 and 0.2 mM EDTA).
Following 30 min of incubation on ice, the lysates were centrifuged
at 14,000 rpm at 4°C for 10 min; the resultant supernatants were
kept as nuclear extracts. The samples were separated by sodium
dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) and
transferred onto poly-vinylidene fluoride (PVDF) membranes.
Specific proteins were sequentially blotted with primary
antibodies, including SOX4 (ab80261; Abcam, Cambridge, UK) p65
(#8242), matrix metalloproteinase (MMP)2 (#13132) and MMP9 (#2270)
(Cell Signaling Technology, Beverly, MA, USA). After washing, the
blots were incubated with HRP-conjugated secondary antibody (#7074;
Cell Signaling Technology) Immunocomplexes were detected using the
ECL detection system (Amersham Pharmacia Biotechnology, Tokyo,
Japan). Densitometric analysis of the western blot analysis results
was carried out using ImageJ software.
RNA extraction and real-time PCR
RNA from the melanoma cell lines was extracted using
TRIzol reagent (Invitrogen). The resulting RNA was reverse
transcribed into cDNA using PrimeScript™ 1st strand cDNA Synthesis
kit (Takara, Dalian, China) and amplified using specific primers.
The primers were composed by Sangon Co. (Shanghai, China) (Table I). The RNA concentration and
quantity were assessed using a NanoDrop Spectrophotometer.
Real-time PCR was performed on a 25 μl reaction system using
the SYBR® Premix DimerEraser™ kit according to the
manufacturer's instructions (Takara). Denaturation was performed at
95°C for 30 sec, annealing at 60°C for 30 sec, and elongation at
72°C for 30 sec for 39 cycles. PCR was carried out in triplicate
and analyzed using the ABI Prism 7900HT Fast RT-PCR system (Applied
Biosystems Life Technologies, Foster City, CA, USA).
 | Table ISequences of primers used for PCR. |
Table I
Sequences of primers used for PCR.
| Gene name | Forward primer | Reverse primer |
|---|
| SOX4 |
ACAGCGACAAGATCCCTTTC |
CGGACTTCACCTTCTTCCTG |
| p65 |
AGCACAGATACCACCAAGACC |
CGGCAGTCCTTTCCTACAAG |
| MMP2 |
TATGGCTTCTGCCCTGAGAC |
CACACCACATCTTTCCGTCA |
| MMP9 |
AGTCCACCCTTGTGCTCTTC |
ACTCTCCACGCATCTCTGC |
Cell migration and invasion assay
Cell invasion assay and migration assay were both
measured by counting the numbers of cells that invaded through a
well with 8.0-μm pores (Costar, Cambridge, UK). Cell
invasion assay was performed using a well coated with Matrigel™
(1:5 dilution; BD Biosciences, San Jose, CA, USA), while the
migration assay was performed using a well without Martigel
coating. The cells were starved with serum-free DMEM for 12 h,
3×104 cells in serum-free DMEM were then added to the
upper chamber and DMEM with 10% serum was added to the lower
chamber. After 24 or 48 h of incubation, the cells on the surface
of the filter membrane were removed with cotton swabs. The invading
cells on the lower surface of the filter membrane were fixed in 4%
paraformaldehyde, stained with 0.5% crystal violet (46364;
Sigma-Aldrich, St. Louis, MO, USA), and counted in 5 random squares
using a microscope (Olympus CX22, Olympus, Tokyo, Japan). Each
experiment was performed in triplicate.
Immunological staining assay
The cells were transfected with si-SOX4 or scrambled
siRNA, After 48 h, the cells were fixed with 4% paraformaldehyde on
the slides and incubatedin 0.2% Triton X-100 for 2 min. The slides
were then blocked with 5% BSA at room temperature for 30 min,
followed by incubation with rabbit anti-p65 antibody (#8242; Cell
Signaling Technology)at 4°C overnight. The cells were then stained
with a secondary Alexa Fluor 594-labeled goat anti-rabbit IgG
(1:500; ab150080; Abcam) and counterstained with 1 μg/ml
4,6-diamino2-phenylindole dihydrochloride (DAPI; Sigma-Aldrich).
The cells were visualized using an Olympus fluorescence
microscope.
Statistical analysis
All data are presented as the means ± standard
deviation. Data analysis was performed by one-way analysis of
variance (ANOVA). For comparisons between 2 groups, a Student's
t-test was used. Differences with P-values <0.05 were considered
to be statistically significant.
Results
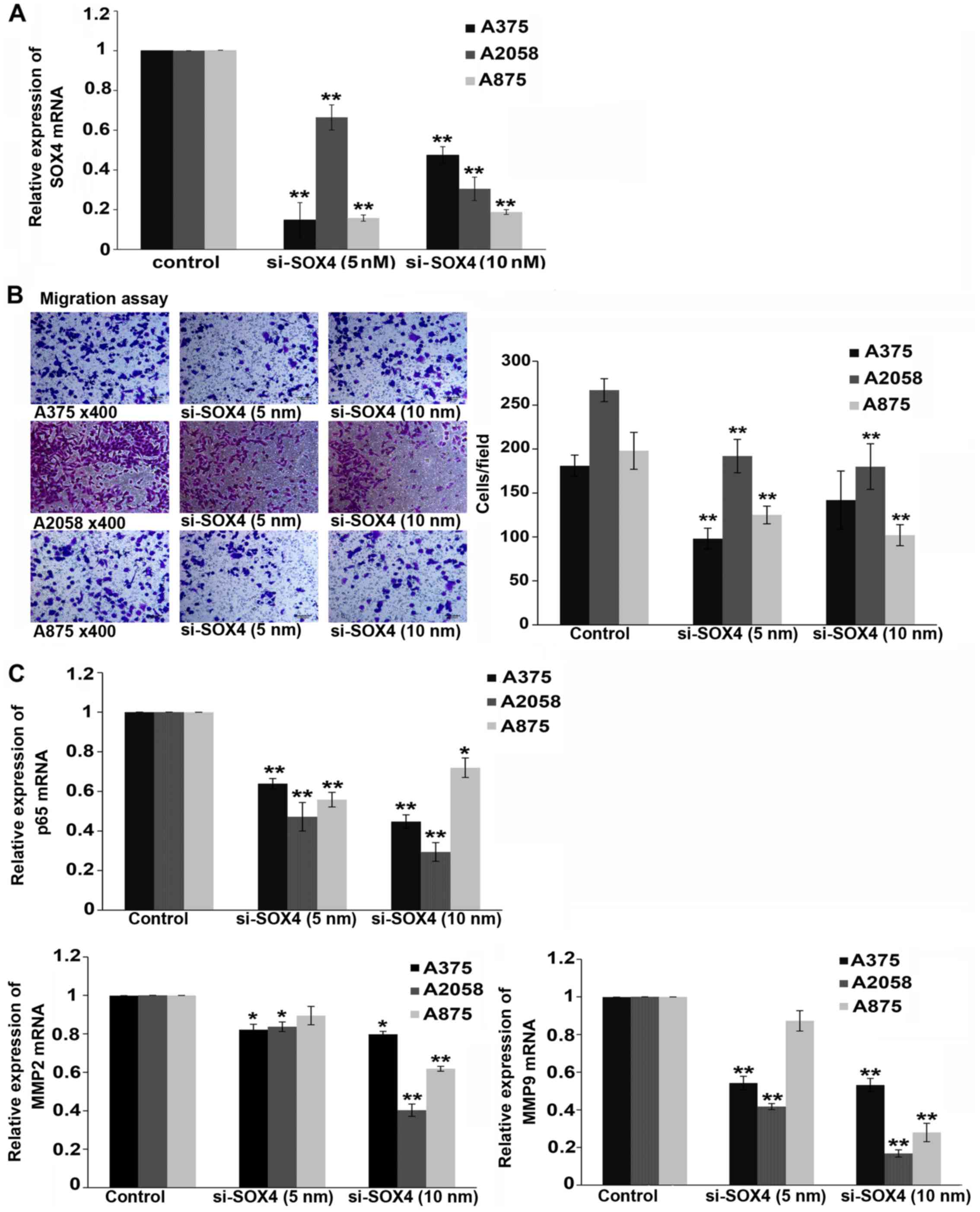
Downregulation of SOX4 induced by SOX4
siRNA inhibits the migration of melanoma cells
We examined the effects of the siRNA-mediated
knockdown of SOX4 in melanoma cell lines (A375, A2058 and A875)
in vitro. Transfection with SOX4-specific siRNA resulted in
a significant decrease in the SOX4 mRNA and protein levels in the
A375, A2058 and A875 cells, as compared with the negative control
(P<0.01 and P<0.05) (Figs.
1A and 2A–C). We then
examined the effects of SOX4 knockdown on cell migration. The
downregulation of SOX4 markedly suppressed the migration of the
melanoma cells (P<0.01; Fig.
1B).
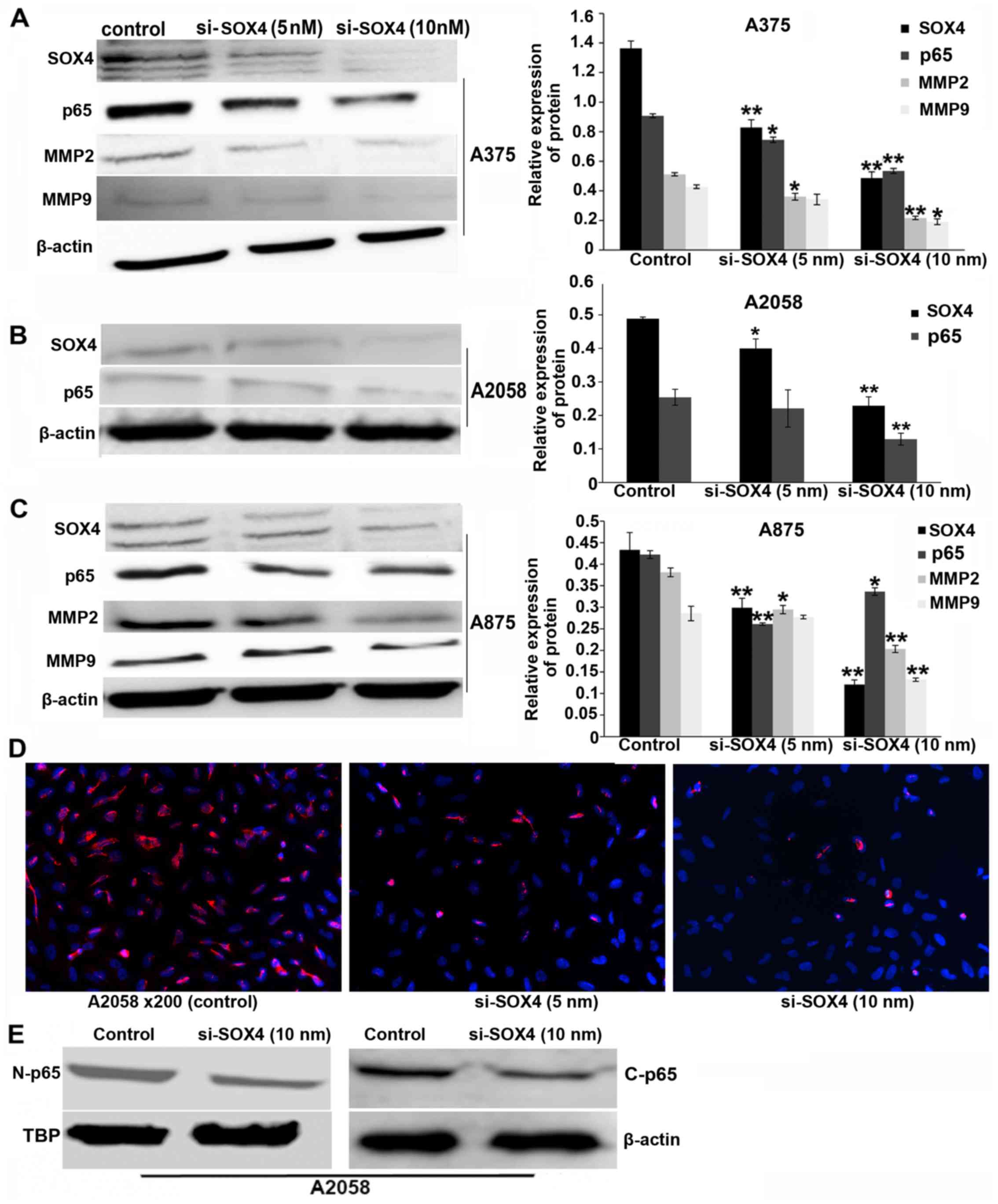
NF-κB p65 is downregulated in
SOX4-siRNA-transfected cells
NF-κB is one of the key modulators of tumor cell
migration and invasion (9,10).
In this study, we observed a marked decrease in the level of NF-κB
p65 mRNA (P<0.01 and P<0.05) (Fig. 1C) and protein (P<0.01 and
P<0.05; Fig. 2A–C) in the SOX4
siRNA-transfected cells compared to the controls. NF-κB is a
transcription factor. The NF-κB p65 expression level was analyzed
by immunological staining. As demonstrated by immunological
staining assay, the expression of nuclear NF-κB was decreased in
the SOX4 siRNA-transfected melanoma cells, as compared with that in
the scrambled siRNA-transfected cells (Fig. 2D). A similar tendency was observed
by western blot analysis (Fig.
2E).
Expression levels of MMP2 and MMP9 are
decreased in SOX4 siRNA-transfected melanoma cells
MMP2 and MMP9 are two of the NF-κB target genes.
Thus, we performed real-time PCR to determine the mRNA expression
of MMP2 and MMP9 following transfection of the melanoma cells with
SOX4 siRNA for 24 h. Our results revealed a marked downregulation
in the mRNA levels of MMP2 and MMP9 in the A375, A2058 and A875
cells transfected with SOX4 siRNA (P<0.01 and P<0.05;
Fig. 1C). Similar results were
observed for MMP2 and MMP9 protein expression after 48 h of SOX4
siRNA transfection (P<0.01 and P<0.05; Fig. 2A and C).
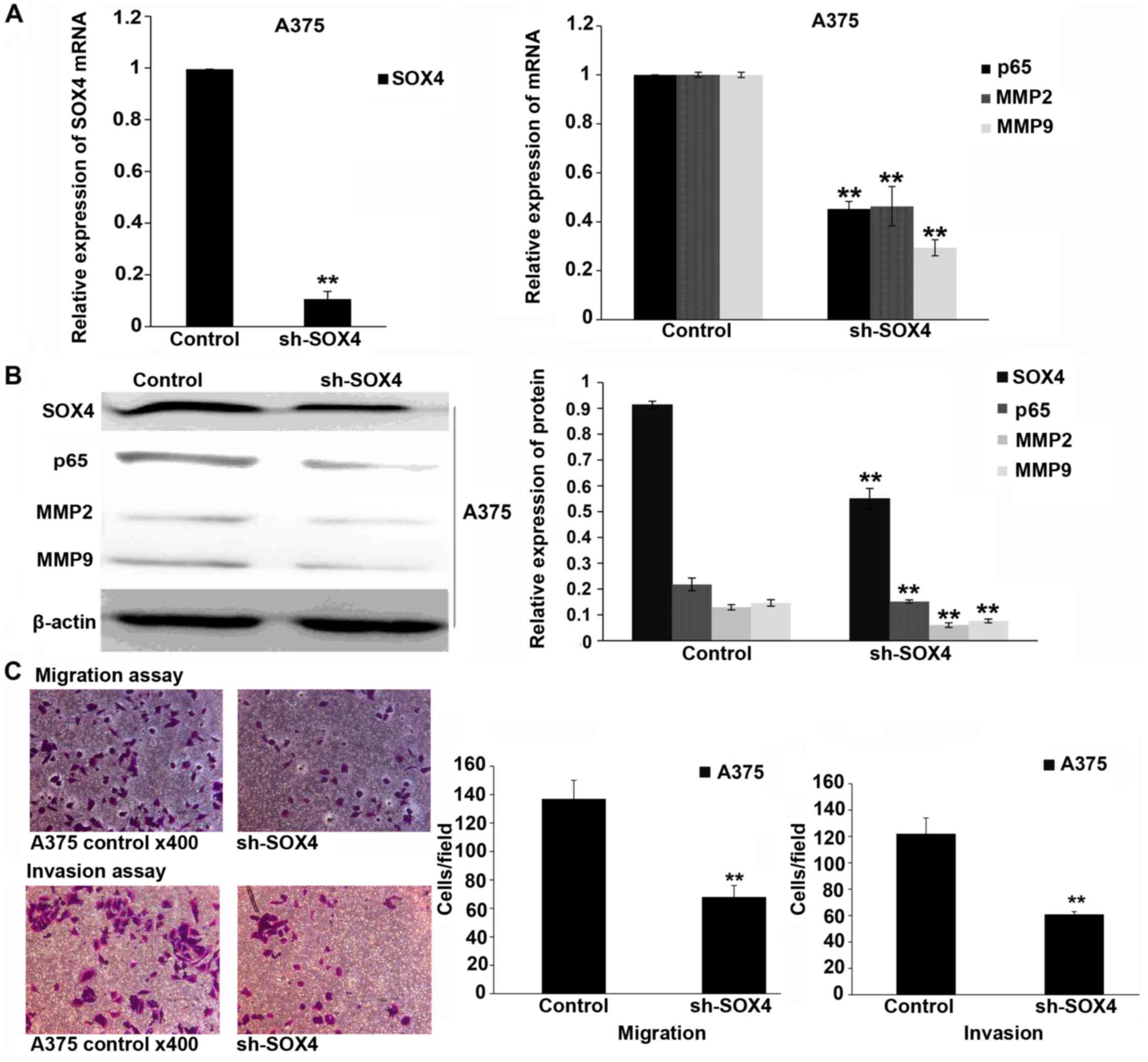
SOX4 shRNA viral transfection decreases
melanoma cell migration and invasion, and inhibits the expression
of MMP2, MMP9 and NF-κB p65
To verify the results observed with SOX4 siRNA
transfection, we performed the experiments using shRNA viral
transfection. After 72 h, SOX4 expression was suppressed by SOX4
shRNA viral transfection. The mRNA and protein levels of p65, MMP2
and MMP9 were also inhibited in the SOX4 shRNA-transfected cells
(P<0.01; Fig. 3A and B). The
downregulation of induced by shRNA viral transfection significantly
suppressed melanoma cell migration and invasion (P<0.01;
Fig. 3C).
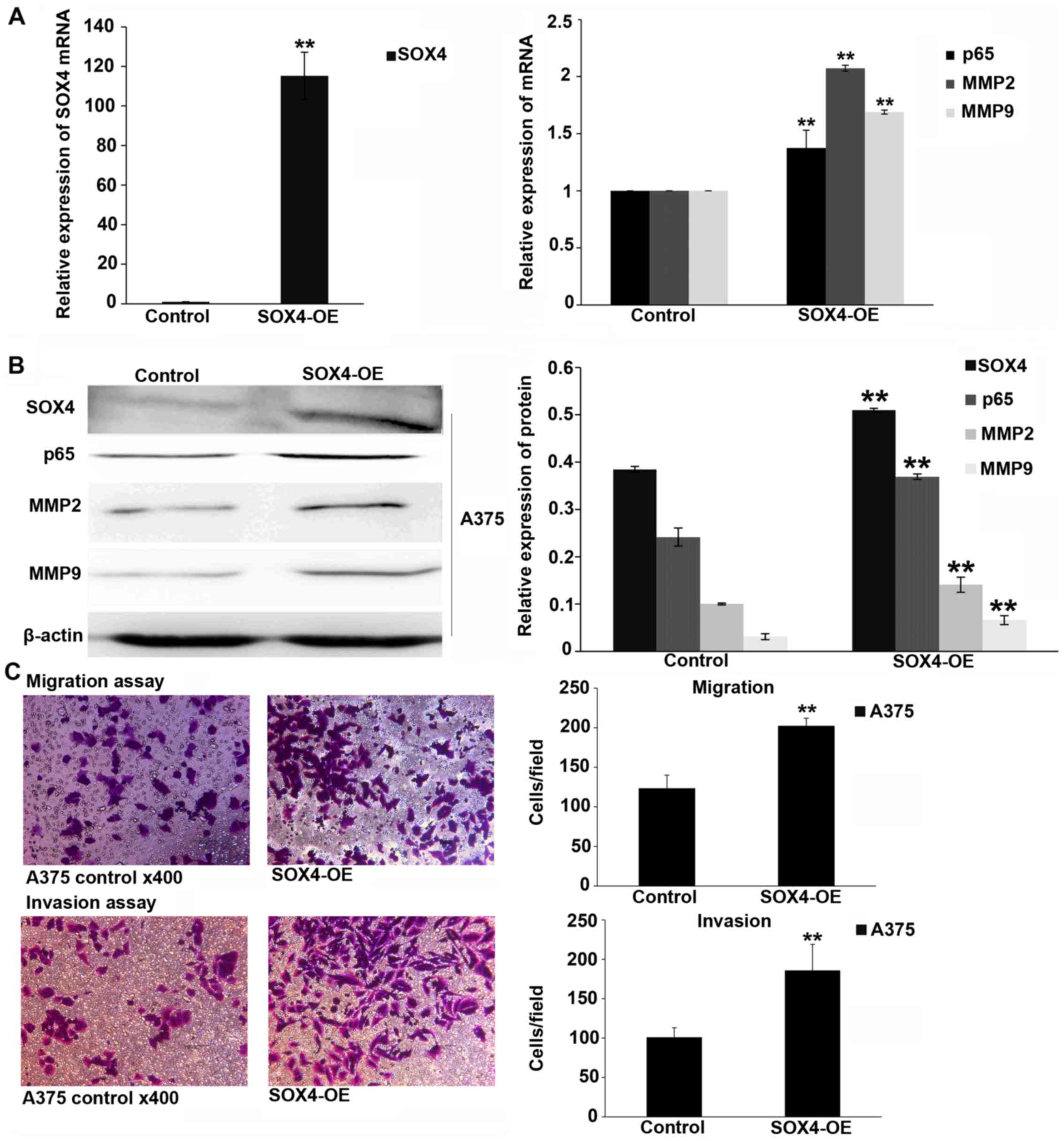
Overexpression of SOX4 promotes melanoma
cell migration and invasion, and upregulates the expression of
MMP2, MMP9 and NF-κB p65
SOX4 was overexpressed by lentiviral transfection.
The mRNA and protein levels of p65, MMP2 and MMP9 were upregulated
in the melanoma cells after 72 h of transfection (P<0.01;
Fig. 4A and B). Migration and
invasion assays demonstrated that the overexpression of SOX4
promoted melanoma cell migration and invasion (P<0.01; Fig. 4C).
Discussion
SOX4 is highly expressed in many tumors and plays an
important role in the occurrence and progression of tumors
(20,21). It has been proposed to act as
either an oncogene or a tumor suppressor (16). Aaboe et al found that SOX4
was overex-pressed in bladder cancer tissues compared to normal
tissues, but a strong SOX4 expression was found to correlate with
increased patient survival (16).
Similar results were observed by Zhang et al (22). Jafarnejad et al reported
that the knockdown of SOX4 enhanced melanoma cell invasion and
migration (17). In this study,
we found that SOX4 siRNA transfection decreased melanoma (A375,
A2058 and A875) cell invasion and migration. Similar anti-invasive
and anti-migratory effects were observed in the SOX4 shRNA
transfected melanoma cells. Different melanoma cell lines were used
in the study by Jafarnejad et al (17) and our study. SOX4 expressed in
different cell lines may bind to different proteins, which
regulates different target genes and lead to different biological
functions.
NF-κB is a major transcription factor which is
present in the cytoplasm in an inactive complex and can be
activated by various stimuli, and is linked to various cellular
processes in cancer, including inflammation, invasion and
metastasis (23). There are 5
members of the NF-κB family in mammals. The most abundant activated
form of NF-κB is a heterodimer composed of a p50 and p65 (24). Aggarwal and Sung demonstrated that
the inhibition of NF-κB in melanoma cells can sensitize tumors to
chemotherapeutic agents (25).
NF-κB has been shown to be upregulated in melanoma and has been
specifically identified as being a potential 'master regulator' of
melanoma invasion (26,27). In this study, we found that the
NF-κB p65 expression level was significantly decreased in the
melanoma cells in which SOX4 was knocked down. In addition, the
NF-κB p65 expression level was elevated in the melanoma cells
overexpressing SOX4.
Activated NF-κB translocates to the nucleus and can
induce the expression of MMPs (23). MMPs are the most important
metastasis-promoting genes (28)
and are responsible for breaking down the extracellular matrix
(ECM) microstructure. The excess degradation of the surrounding ECM
is one of the hallmarks of tumor invasion and metastasis. MMP2 and
MMP9 can degrade several ECM proteins and correlate with tumor
invasion and metastasis; they are known to be regulated by several
intracellular signaling pathways (29). Highly aggressive melanoma cells
express high levels of MMP2 and MMP9 (30). We observed that the knockdown of
SOX4 significantly downregulated MMP2 and MMP9 expression, whereas
the overexpression of SOX4 upregulated MMP2 and MMP9
expression.
In conclusion, this study demonstrates that SOX4
promotes melanoma cell migration and invasion. These effects were
partly mediated through the activation of the NF-κB/p65 signaling
pathway. Further in vivo studies are required in order to
verify the function of SOX4 in melanoma cell migration and
invasion.
Acknowledgments
This study was funded by a grant d from the Shanghai
Science and Technology Committee of China (no. 13JC1401401).
References
|
1
|
Romano S, Xiao Y, Nakaya M, D'Angelillo A,
Chang M, Jin J, Hausch F, Masullo M, Feng X, Romano MF, et al:
FKBP51 employs both scaffold and isomerase functions to promote
NF-κB activation in melanoma. Nucleic Acids Res. 43:6983–6993.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Peterson M, Albertini MR and Remington P:
Incidence, Survival, and Mortality of Malignant Cutaneous Melanoma
in Wisconsin, 1995–2011. WMJ. 114:196–201. 2015.
|
|
3
|
Zehavi L, Schayek H, Jacob-Hirsch J, Sidi
Y, Leibowitz-Amit R and Avni D: MiR-377 targets E2F3 and alters the
NF-κB signaling pathway through MAP3K7 in malignant melanoma. Mol
Cancer. 14:682015. View Article : Google Scholar
|
|
4
|
Li J, Zhang Z and Li G: Patient outcome
prediction using multiple biomarkers in human melanoma: A
clinicopathological study of 118 cases. Exp Ther Med. 2:131–135.
2011.PubMed/NCBI
|
|
5
|
Gajos-Michniewicz A and Czyz M: Modulation
of WNT/β-catenin pathway in melanoma by biologically active
components derived from plants. Fitoterapia. 109:283–292. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Webter MR, Kuqel CH III and Weeraratna AT:
The Wnts of change: How Wnts regulate phenotype switching in
melanoma. Biochim Biophys Acta. 1856:244–251. 2015.
|
|
7
|
Daphu I, Horn S, Stieber D, Varughese JK,
Spriet E, Dale HA, Skaftnesmo KO, Bjerkvig R and Thorsen F: In
vitro treatment of melanoma brain metastasis by simultaneously
targeting the MAPK and PI3K signaling pathways. Int J Mol Sci.
15:8773–8794. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Yajima I, Kumasaka MY, Thang ND, Goto Y,
Takeda K, Yamanoshita O, Iida M, Ohgami N, Tamura H, Kawamoto Y, et
al: RAS/RAF/MEK/ERK and PI3K/PTEN/AKT signaling in malignant
melanoma progression and therapy. Dermatol Res Pract.
2012:3541912012.
|
|
9
|
Hu S, Luo Q, Cun B, Hu D, Ge S, Fan X and
Chen F: The pharmacological NF-κB inhibitor BAY11-7082 induces cell
apoptosis and inhibits the migration of human uveal melanoma cells.
Int J Mol Sc. 13:15653–15667. 2012. View Article : Google Scholar
|
|
10
|
Wu ZY, Lien JC, Huang YP, Liao CL, Lin JJ,
Fan MJ, Ko YC, Hsiao YP, Lu HF and Chung JG: Casticin inhibits
A375.S2 human melanoma cell migration/invasion through
downregulating NF-κB and matrix metalloproteinase-2 and -1.
Molecules. 21:3842016. View Article : Google Scholar
|
|
11
|
Li Y, Zu L, Wang Y, Wang M, Chen P and
Zhou Q: miR-132 inhibits lung cancer cell migration and invasion by
targeting SOX4. J Thorac Dis. 7:1563–1569. 2015.PubMed/NCBI
|
|
12
|
Song GD, Sun Y, Shen H and Li W: SOX4
overexpression is a novel biomarker of malignant status and poor
prognosis in breast cancer patients. Tumour Biol. 36:4167–4173.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Zhang J, Liang Q, Lei Y, Yao M, Li L, Gao
X, Feng J, Zhang Y, Gao H, Liu DX, et al: SOX4 induces
epithelial-mesenchymal transition and contributes to breast cancer
progression. Cancer Res. 72:4597–4608. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Wang L, Zhang J, Yang X, Chang YW, Qi M,
Zhou Z, Zhang J and Han B: SOX4 is associated with poor prognosis
in prostate cancer and promotes epithelial-mesenchymal transition
in vitro. Prostate Cancer Prostatic Dis. 16:301–307. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Vervoort SJ, van Boxtel R and Coffer PJ:
The role of SRY-related HMG box transcription factor 4 (SOX4) in
tumorigenesis and metastasis: Friend or foe? Oncogene.
32:3397–3409. 2013. View Article : Google Scholar
|
|
16
|
Aaboe M, Birkenkamp-Demtroder K, Wiuf C,
Sørensen FB, Thykjaer T, Sauter G, Jensen KM, Dyrskjøt L and
Ørntoft T: SOX4 expression in bladder carcinoma: Clinical aspects
and in vitro functional characterization. Cancer Res. 66:3434–3442.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Jafarnejad SM, Wani AA, Martinka M and Li
G: Prognostic significance of Sox4 expression in human cutaneous
melanoma and its role in cell migration and invasion. Am J Pathol.
177:2741–2752. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Wang C, Zhao H, Lu J, Yin J, Zang L, Song
N, Dong R, Wu T and Du X: Clinicopathological significance of SOX4
expression in primary gallbladder carcinoma. Diagn Pathol.
7:412012. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Harris ML, Baxter LL, Loftus SK and Pavan
WJ: Sox proteins in melanocyte development and melanoma. Pigment
Cell Melanoma Res. 23:496–513. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Rhodes DR, Yu J, Shanker K, Deshpande N,
Varambally R, Ghosh D, Barrette T, Pandey A and Chinnaiyan AM:
Large-scale meta-analysis of cancer microarray data identifies
common transcriptional profiles of neoplastic transformation and
progression. Proc Natl Acad Sci USA. 101:9309–9314. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Lefebvre V and Bhattaram P: SOXC genes and
the control of skeletogenesis. Curr Osteoporos Rep. 14:32–38. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Zhang J, Jiang H, Shao J, Mao R, Liu J, Ma
Y, Fang X, Zhao N, Zheng S and Lin B: SOX4 inhibits GBM cell growth
and induces G0/G1 cell cycle arrest through Akt-p53 axis. BMC
Neurol. 14:2072014. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Kumar A, Takada Y, Boriek AM and Aggarwal
BB: Nuclear factor-kappaB: Its role in health and disease. J Mol
Med (Berl). 82:434–448. 2004. View Article : Google Scholar
|
|
24
|
Emdad L, Sarkar D, Su ZZ, Randolph A,
Boukerche H, Valerie K and Fisher PB: Activation of the nuclear
factor kappaB pathway by astrocyte elevated gene-1: Implications
for tumor progression and metastasis. Cancer Res. 66:1509–1516.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Aggarwal BB and Sung B: NF-κB in cancer: A
matter of life and death. Cancer Discov. 1:469–471. 2011.
View Article : Google Scholar
|
|
26
|
Amiri KI and Richmond A: Role of nuclear
factor-kappa B in melanoma. Cancer Metastasis Rev. 24:301–313.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Ryu B, Kim DS, Deluca AM and Alani RM:
Comprehensive expression profiling of tumor cell lines identifies
molecular signatures of melanoma progression. PLoS One. 2:e5942007.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Kim A, Kim MJ, Yang Y, Kim JW, Yeom YI and
Lim JS: Suppression of NF-kappaB activity by NDRG2 expression
attenuates the invasive potential of highly malignant tumor cells.
Carcinogenesis. 30:927–936. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Hong IK, Kim YM, Jeoung DI, Kim KC and Lee
H: Tetraspanin CD9 induces MMP-2 expression by activating p38 MAPK,
JNK and c-Jun pathways in human melanoma cells. Exp Mol Med.
37:230–239. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Seftor RE, Seftor EA, Koshikawa N, Meltzer
PS, Gardner LM, Bilban M, Stetler-Stevenson WG, Quaranta V and
Hendrix MJ: Cooperative interactions of laminin 5 gamma2 chain,
matrix metalloproteinase-2, and membrane
type-1-matrix/metalloproteinase are required for mimicry of
embryonic vasculogenesis by aggressive melanoma. Cancer Res.
61:6322–6327. 2001.PubMed/NCBI
|