Introduction
Osteoarthritis (OA), one of the most common chronic
diseases of the joint and highly correlated with increasing age, is
characterized by cartilage degeneration (1,2).
Cartilage is the dense connective tissue located in the joints
between bones, and is composed of chondrocytes and extracellular
matrix (ECM) (3). Cartilage
degradation can lead to broken dynamic equilibrium between normal
cartilage ECM synthesis and degradation. Chondrocytes, the only
cell population of articular cartilage, are involved in maintaining
cartilage homeostasis (4,5). Cartilage degradation occurs in
pathological conditions of OA (6). Therefore, improving and maintaining
the proliferation potential and phenotype of chondrocytes are
essential for the inhibition of cartilage degradation (7). This may potentially be an effective
method with which to delay the development and progression of
OA.
The Wnt-β/catenin signaling pathway is involved in
the regulation of cartilage homeostasis, which plays a crucial role
in the process of cell proliferation and the regulation of the
chondrocyte phenotype (8,9). Activation of the Wnt/β-catenin
signaling pathway is sufficient to induce dedifferentiation of
articular chondrocytes (10).
Following activation of the canonical Wnt/β-catenin signaling
pathway, β-catenin can accumulate in the cytoplasm and translocate
to the nucleus, subsequently interacting with transcription factors
such as T-cell factor and lymphoid enhancer binding factor (LEF)
(11). Finally, these factors
influence chondrocyte proliferation by affecting the expression of
cyclin D1, a crucial factor in the cell cycle.
Psoralen (PSO) is one of the major active
constituents of Fructus Psoraleae (Bu Gu Zhi) which is the dried
ripe fruit of Psoralea corylifolia L. It has been commonly
used in traditional Chinese medicine (TCM) for the treatment of
osteoporosis, osteosarcoma, bone fracture, and osteomalacia
(12). Studies have demonstrated
that PSO stimulates local new bone formation in vivo and
triggers osteogenesis (13,14). A previous study showed that PSO
may be used in the prevention and treatment of OA (15). However, the precise molecular
mechanisms responsible for its effect on chondrocyte proliferation
remain to be elucidated. Thus, our present study aimed to determine
whether PSO promotes chondrocyte proliferation by activating the
Wnt/β-catenin signaling pathway.
Materials and methods
Animals
Male Sprague-Dawley (SD) rats at 6 weeks of age were
purchased from Shanghai SLAC Laboratory Animal Co., Ltd. (Shanghai,
China). Animal experiments were strictly conducted in accordance
with the Guide for the Care and Use of Laboratory Animals of Fujian
University of Traditional Chinese Medicine (Fuzhou, China). The
study protocol was approved by the Animal Care and Use Committee of
Fujian University of Traditional Chinese Medicine. The SD rats were
sacrificed using carbon dioxide (cage size, 7×11×5 inches; flow
rate, 1.3 l/min), according to the Guide for the Care and Use of
Animals.
Preparation of PSO-conditioned culture
medium
PSO (psoralen, NIFDC, batch no. 110739-201115, 99.3%
pure) was first dissolved in phosphate-buffered-saline (PBS;
HyClone Laboratories, Inc., Logan, UT, USA) to a concentration of
10−3 mol/l, and stored at −20°C. The PSO-conditioned
culture medium was prepared by diluting the stock solution in
low-Dulbecco's modified Eagle's medium (Low-DMEM) containing 10%
fetal bovine serum (FBS) (both from HyClone Laboratories, Inc.),
filtering through a 0.22-µm filter and stored at 4°C before
use.
Isolation and identification of
chondrocytes
Articular chondrocytes were isolated and cultured as
previously described (16). The
morphological changes and growth feature of chondrocytes were
recorded under a phase-contrast microscope (Olympus, Tokyo, Japan).
The primary chondrocytes were termed passage 0 (P0); the P2
chondrocytes were identified by immunohistochemical staining of
type II collagen. P2 chondrocytes approximately at 50% were used in
this study.
Cell viability analysis
Chondrocytes were seeded at the 96-well culture
plates with a density of 2×104 cells/ml and incubated
(100 µl/well) for 24 h. The cells were then treated with
PSO-conditioned culture medium at different concentrations (0,
10−8, 10−7, 10−6, 10−5
and 10−4 mol/l) for different periods (24, 48 and 72 h).
At the end of intervention, the supernatant was then removed and
100 µl 1% MTT (Sigma-Aldrich) was added to each well. After
4 h of incubation at 37°C, the supernatant was replaced with 150
µl/well of DMSO (Hengxing Chemical Preparation Co., Ltd.,
Tianjin, China). The absorbance was measured at 490 nm using an
enzyme labeling instrument (model EXL800; BioTek, Winooski, VT,
USA). The optimum conditions of intervention were selected for
further examination. To further investigate whether PSO promotes
chondrocyte proliferation via the canonical Wnt-β/catenin signaling
pathway, the chondrocytes were treated with PSO (10−6
mol/l) in the absence or presence of 0.2 µg/ml Dickkopf-1
(DKK-1; R&D Systems, Minneapolis, MN, USA) for 48 h, as
previously described (17).
RNA extraction and RT-PCR analysis
After treatment, total RNA was extracted from the
cells using TRIzol reagent (Invitrogen, Grand Island, NY, USA). RNA
(1 µg) was reverse transcribed into cDNA using a reverse
transcription kit (Thermo Fisher Scientific, Inc., Rockford, IL,
USA) according to the manufacturer's instructions. Then DNA bands
were analyzed via gel electrophoresis (1.5% agarose) using the Gel
Documentation System (Bio-Rad, Hercules, CA, USA) and normalized to
that of β-actin. The PCR primers were as follows: β-actin forward,
5′-GAG AGG GAA ATC GTG CGT GAC-3′ and reverse, 5′-CAT CTG CTG GAA
GGT GGA CA-3′; Wnt-4 forward, 5′-TCA GCC CAC AGG GTT TCC A-3′ and
reverse, 5′-CGC TCG CCA GCA TGT CTT T-3′; β-catenin forward, 5′-AAG
GAA GCT TCC AGA CAT GC-3′ and reverse, 5′-AGC TTG CTC TCT TGA TTG
CC-3′; Frizzled-2 forward, 5′-TCG AGG CCA ATT CGC AGT A-3′ and
reverse, 5′-CAG GAA GGA TGT GCC GAT G-3′; glycogen synthase
kinase-3β (GSK-3β) forward, 5′-AAA GTG CAT CGC TGG CTT A-3′ and
reverse, 5′-GTC GAC GGT TTG TTT CCA AT-3′; cyclin D1 forward,
5′-AAT GCC AGA GGC GGA TGA GA-3′ and reverse, 5′-GCT TGT GCG GTA
GCA GGA GA-3′; Col-II forward, 5′-CCA GAG TGG AAG AGC GGA GAC-3′
and reverse, 5′-CAG TGG ACA GTA GAC GGA GGA AAG-3′; and β-actin
forward, 5′-CAC CCG CGA GTA CAA CCT TC-3′ and reverse, 5′-CCC ATA
CCC ACC ATC ACA CC-3′.
Western blot analysis
After treatment, total proteins were extracted from
cells using radioimmunoprecipitation assay lysis buffer (RIPA) with
1 mM phenylmethanesulfony fluoride (PMSF) (both from Beyotime
Biotechnology, Shanghai, China) and quantified using a
bicinchoninic acid (BCA) assay. Twenty micrograms of proteins were
separated on sodium dodecyl sulfate-polyacrylamide gel
electrophoresis (SDS-PAGE) gels (12%) and transferred onto a PVDF
membrane. Subsequently, the membrane was blocked with 5% non-fat
milk in TBST solution for 2 h at room temperature (RT). After
incubation with the primary antibodies against Wnt-4 (sc-5214),
Frizzled-2 (sc-68327; Santa Cruz Biotechnology, Inc., Santa Cruz,
CA, USA), β-catenin (9582s), GSK-3β (9315; Cell Signaling
Technology, Inc., Beverly, MA, USA), cyclin D1 (BS6532), Col-II
(BS1071; Bioworld Technology, Natong, China) overnight at 4°C, and
the HRP-conjugated secondary antibodies (ZB-2301; Zhongshan Golden
Bridge Biotechnology Co., Ltd.) for 1 h at RT, the blots were
detected using a Bio-Rad Chemi Doc XRS+ (Bio-Rad), and β-actin was
used as the control.
Immunofluorescence staining
After treatment with PSO at the concentration of
10−6 mol/l, chondrocytes were fixed with ice-cold
methanol at 4°C for 30 min, permeabilized with 0.5% Triton X-100
for 10 min, and blocked in 5% bovine serum albumin (BSA) for 1 h at
room temperature. Then the cells were incubated with rabbit
anti-β-catenin antibody overnight at 4°C, and TRITC-conjugated
secondary antibody (Zymed Laboratories, San Francisco, CA, USA) was
applied for 1 h at room temperature in the dark. Following DAPI
staining, confocal images were acquired with a fluorescence
microscope (Olympus, Tokyo, Japan).
Statistical analysis
All the experiments were repeated three times
independently and all the data were analyzed by one way analysis of
variance (ANOVA) or Student's t-test using SPSS 19.0 software (SPSS
Inc, Chicago, IL, USA). P<0.05 was considered to indicate a
statistically significant difference.
Results
Morphology and identification of
chondrocytes
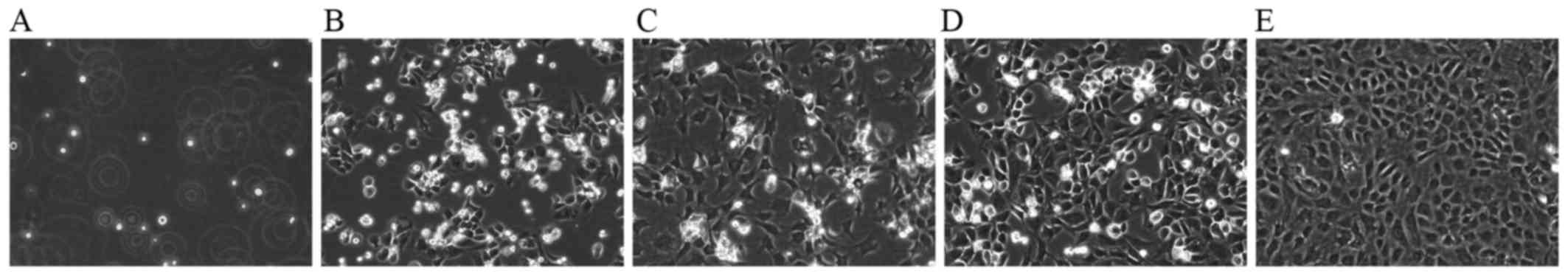
The cultured chondrocytes in this study exhibited
typical morphology with a spherical, fusiform and slab stone shape,
as described in previous studies (17,18) (Fig.
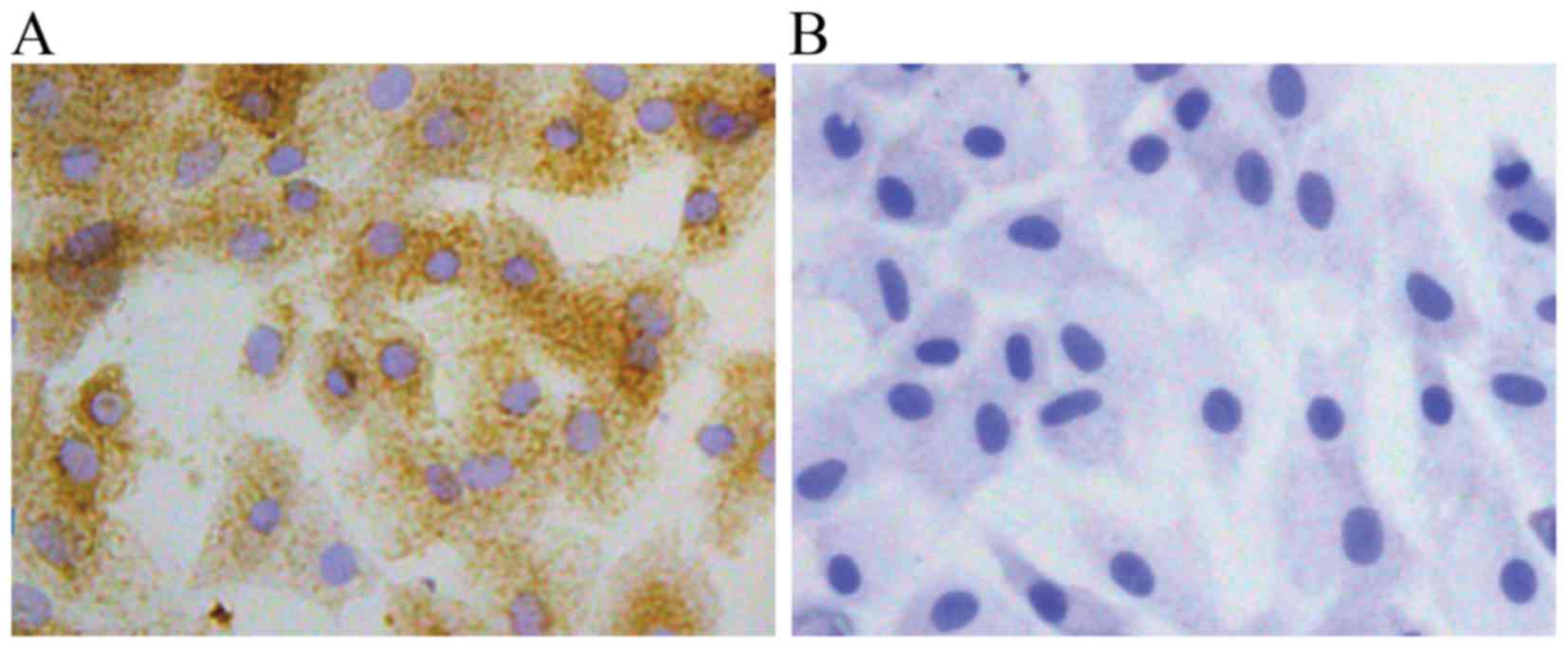
1). The P2 chondrocytes exhibited a typical polygonal or
spherical shape, as shown by the identification of type II collagen
immunohistochemical staining. The cytoplasm of positive
chondrocytes was stained brown, whereas the negative control failed
to stain in the cytoplasm (Fig.
2).
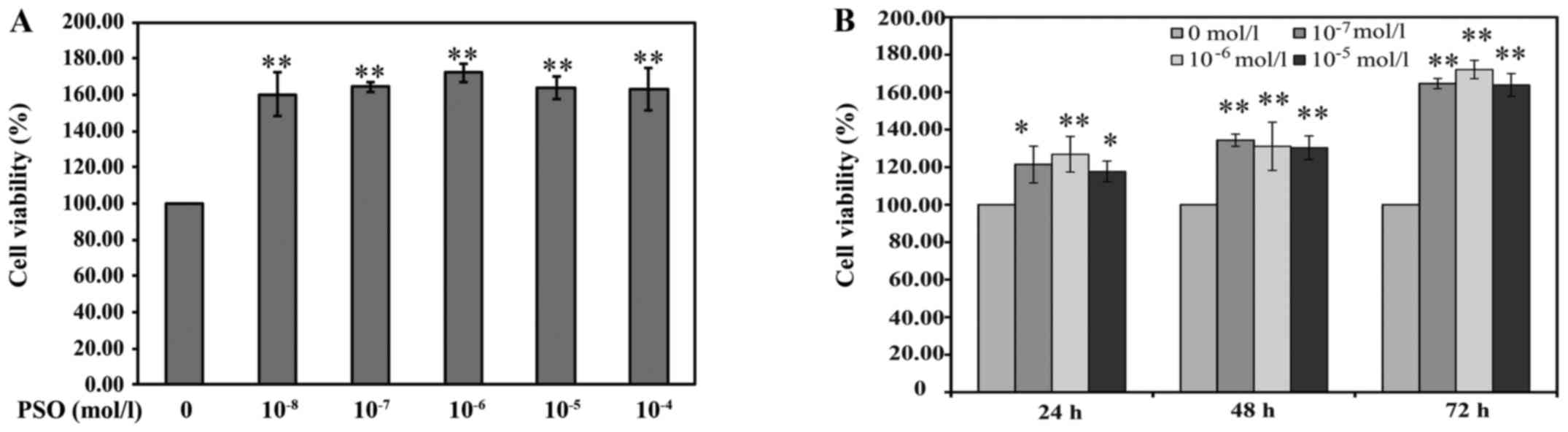
PSO increases chondrocyte viability
The effect of PSO on cell viability was measured by
the MTT assay. As shown by the results of the MTT assay, the
viability of the chondrocytes was increased following treatment
with different PSO dosages of 10−8–10−4 mol/l
and for increasing time periods as compared with the viability
noted in the untreated cells. The cell viability of the
10−6 mol/l PSO concentration treatment group was
slightly higher than that in the other PSO concentration treatment
groups. However, there was no significant difference among the
10−8–10−4 mol/l PSO concentration treatment
groups. Furthermore, treatments with 10−7,
10−6 and 10−5 mol/l of PSO were more
effective on the third day compared with the first day, and all the
comparisons showed significant difference (P<0.01 or P<0.05)
(Fig. 3). These results
preliminarily indicate that PSO promotes the viability of
chondrocytes in a time- and concentration-dependent manner. Thus,
the concentrations of 10−7, 10−6 and
10−5 mol/l (low, medium and high doses, respectively) of
PSO for a 48-h incubation were used for further examination.
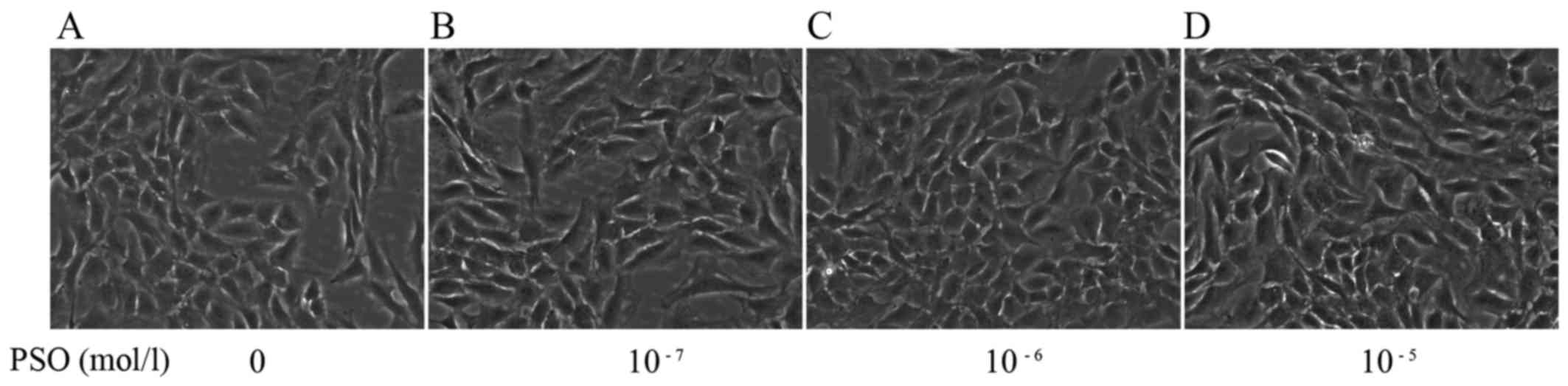
The morphology of the PSO-treated chondrocytes at 48
h was observed by phase-contrast microscope. Compared with the
untreated cells, the PSO-treated chondrocytes exhibited changes in
cell size and shape, and in particular an increase in cell number
(Fig. 4).
PSO upregulates the expression of Wnt-4,
Frizzled-2, β-catenin, cyclin D1 and downregulates the expression
of GSK-3β
To evaluate the effect of PSO on the Wnt/β-catenin
signaling pathway in chondrocytes, RT-PCR and western blot analysis
were performed to determine the expression levels of Wnt-4,
Frizzled-2, β-catenin, cyclin D1 and GSK-3β in chondrocytes.
Compared with the control group, PSO efficiently upregulated the
mRNA expression of Wnt-4, Frizzled-2, β-catenin, cyclin D1
(P<0.01 or P<0.05), but downregulated the mRNA expression of
GSK-3β (P<0.01 or P<0.05) (Fig.
5). Similarly, the protein levels, respectively, corresponded
to the mRNA expression (P<0.01 or P<0.05) (Fig. 6).
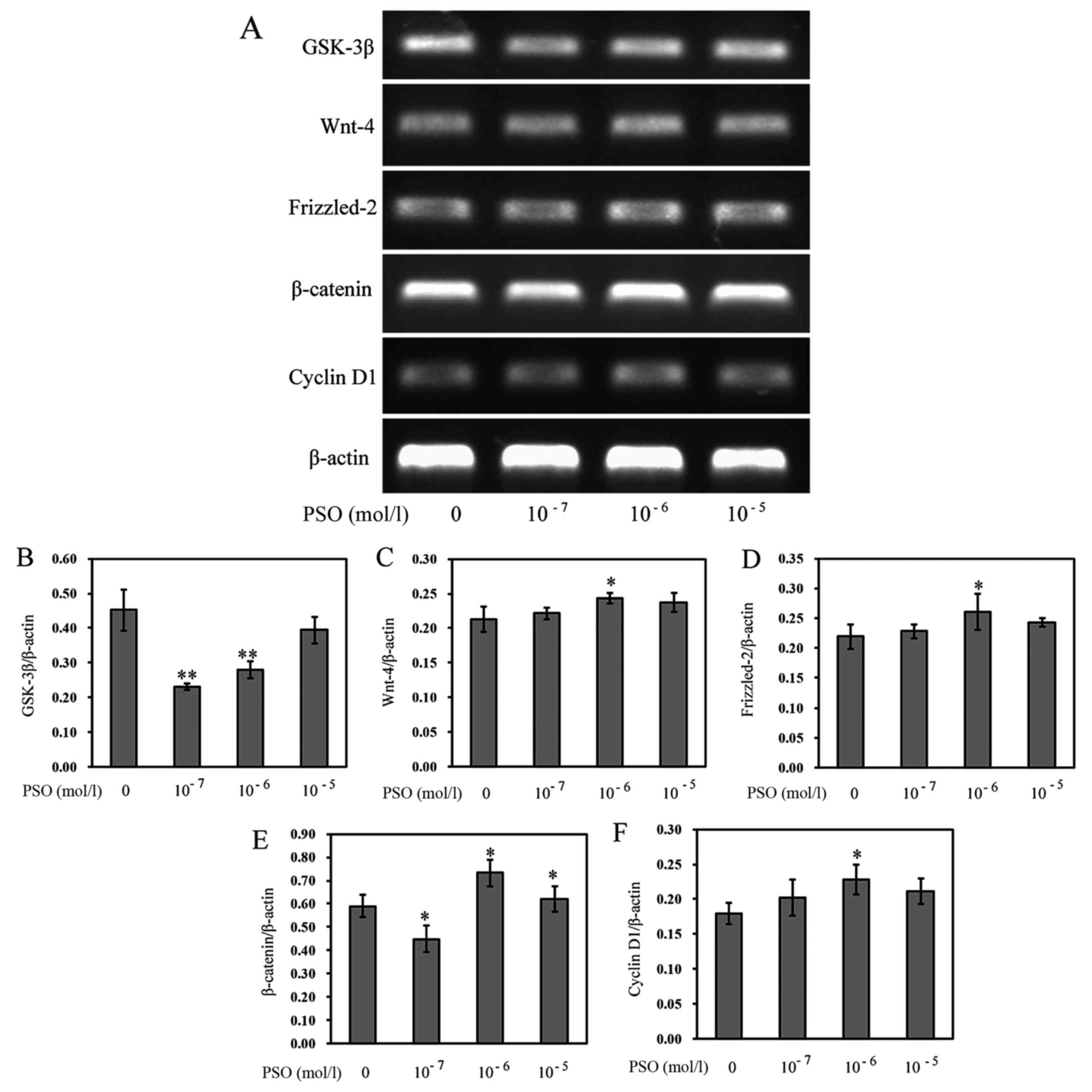 | Figure 5Psoralen (PSO) increases the mRNA
expression of Wnt-4, Frizzled-2, β-catenin, cyclin D1, and
decreases the expression of glycogen synthase kinase-3β (GSK-3β).
(A) mRNA expression of GSK-3β, Wnt-4, Frizzled-2, β-catenin and
cyclin D1 as analyzed by RT-PCR. (B-F) mRNA expression of (B)
GSK-3β, (C) Wnt-4, (D) Frizzled-2, (E) β-catenin, and (F) cyclin D1
in chondrocytes treated with or without PSO. *P<0.05
and **P<0.01 compared with the untreated chondrocytes
(treated with 0 mol/l PSO). |
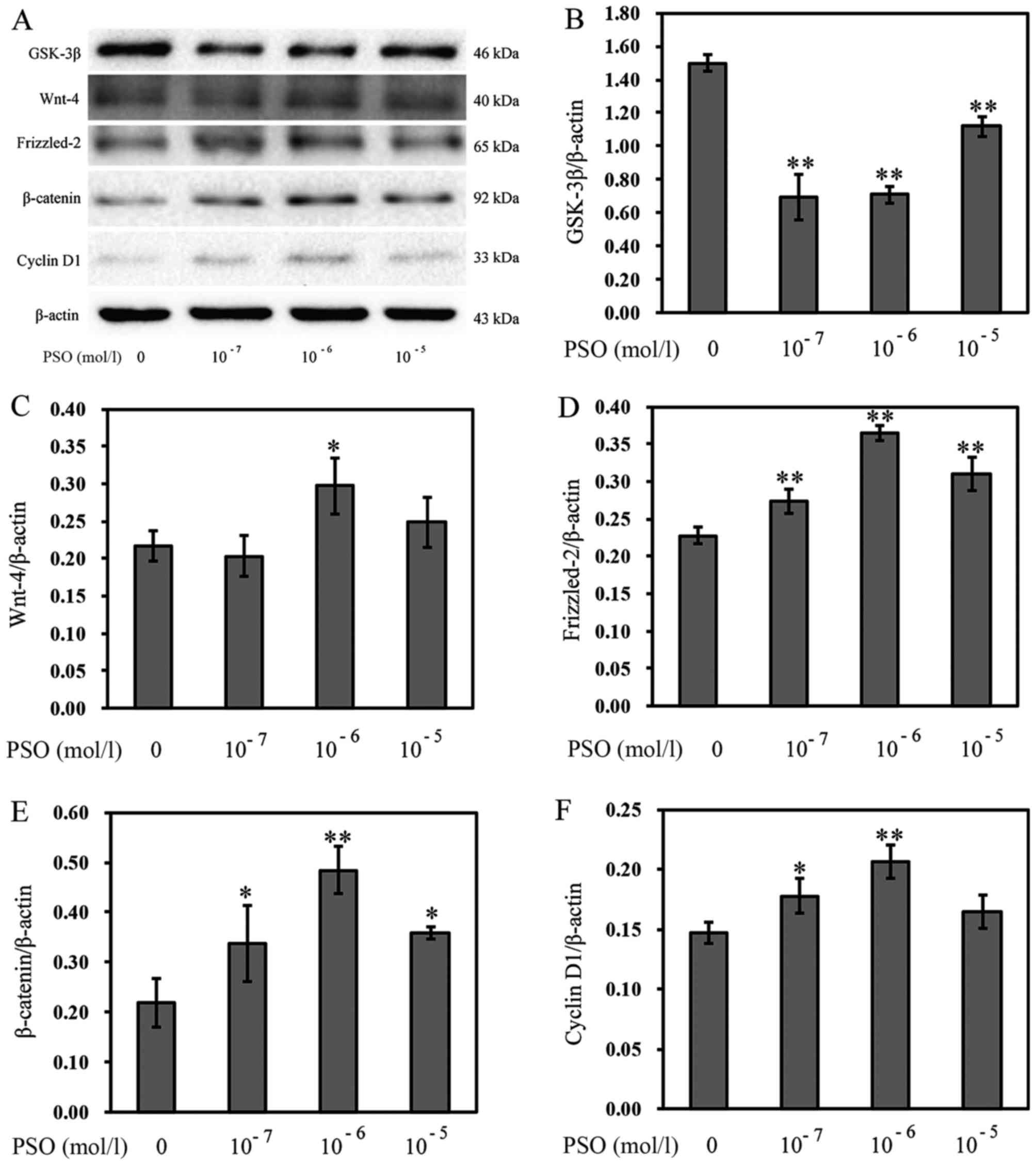 | Figure 6Psoralen (PSO) increases the protein
levels of Wnt-4, Frizzled-2, β-catenin, cyclin D1, and decreases
the protein level of glycogen synthase kinase-3β (GSK-3β). (A) The
protein levels of GSK-3β, Wnt-4, Frizzled-2, β-catenin and cyclin
D1 as analyzed by western blotting. (B-E) The protein levels of (B)
GSK-3β, (C) Wnt-4, (D) Frizzled-2, (E) β-catenin and (F) cyclin D1
in chondrocytes treated with or without PSO. *P<0.05
and **P<0.01 compared with the untreated chondrocytes
(treated with 0 mol/l PSO). |
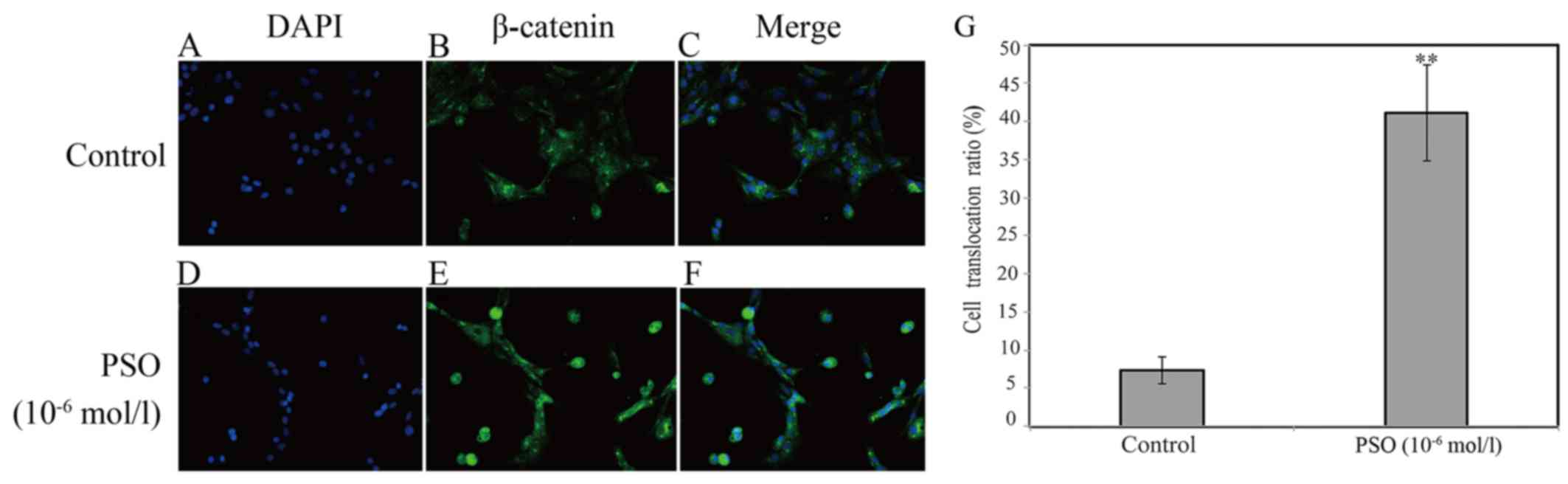
PSO promotes β-catenin nuclear
translocation
To gain insight into the effect of PSO on the
promotion of β-catenin nuclear translocation, immunofluorescence
staining was applied. According to the staining results, we found
that PSO markedly promoted the translocation of β-catenin into the
nucleus (P<0.01 or P<0.05) (Fig. 7). β-catenin is a protein that
mainly localizes in the membrane with minimal localization in the
cytoplasm. It can enter into the nucleus to transactivate target
gene expression upon activation in the cytoplasm (19). These results indicate that PSO
activates the Wnt/β-catenin signaling pathway by promoting
β-catenin nuclear translocation.
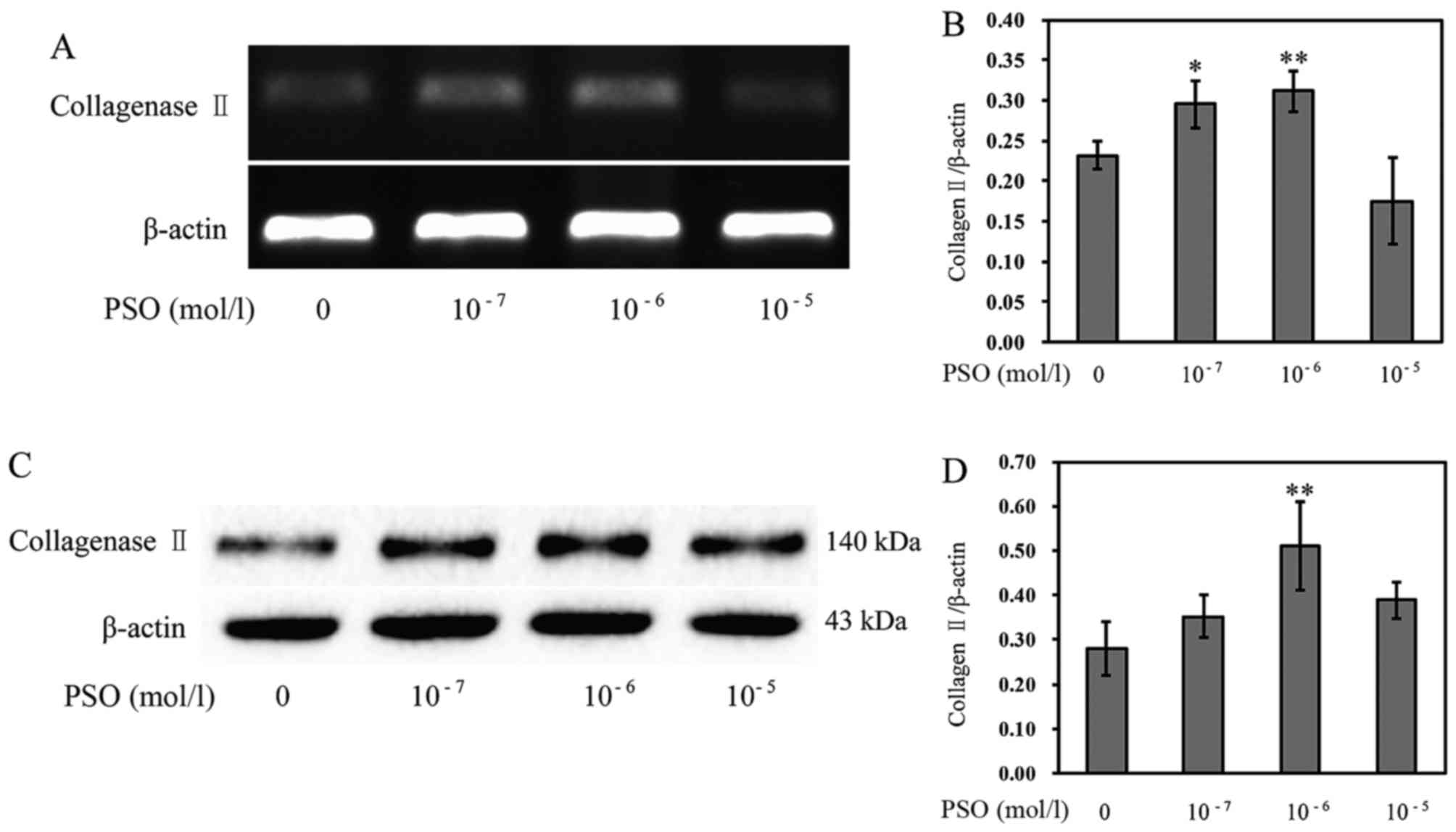
PSO increases the expression of
Col-II
The loss of type II collagen is a characteristic of
articular cartilage degradation. Thus, we investigated the
influence of PSO on Col-II expression. The results revealed that
the mRNA expression and protein level of Col-II were markedly
enhanced in the PSO-treated chondrocytes (P<0.01 or P<0.05)
(Fig. 8), compared with levels in
the untreated cells, indicating that PSO upregulates the expression
of Col-II to promote cartilage ECM synthesis, and confer a positive
effect on chondrocyte proliferation.
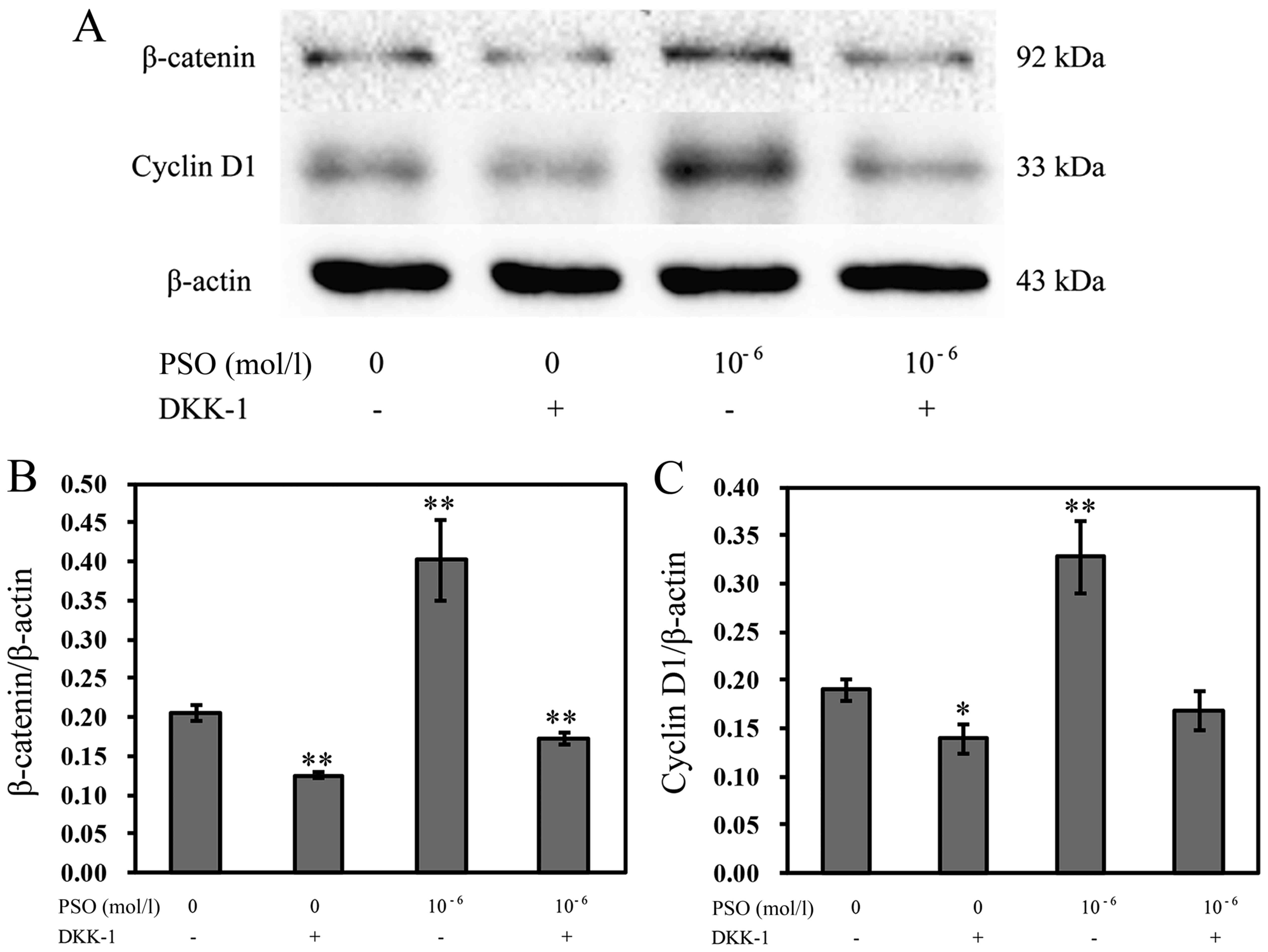
Expression of β-catenin and cyclin D1 in
the Wnt/β-catenin signaling pathway is inhibited by DKK-1
To further confirm the effects of PSO on chondrocyte
proliferation by activating the Wnt/β-catenin signaling pathway, we
also investigated the influences of Wnt/β-catenin signaling
inhibition on β-catenin and cyclin D1 expression. The expression of
β-catenin and cyclin D1 was decreased in cells pre-treated with
DKK-1 compared with that without DKK-1 treatment (P<0.01 or
P<0.05), implying that PSO is involved in the regulation of
chondrocyte proliferation by activating the Wnt/β-catenin signaling
pathway (Fig. 9).
Discussion
OA is a progressively degenerative joint disorder
that is gradually becoming a major health issue among the aged
population worldwide (20).
Chondrocytes are responsible for the production and maintenance of
the ECM. In OA, dysfunction of articular chondrocytes causes
degradation of the ECM exceeding its synthesis, leading to the
degradation of cartilage (21).
Thus, the development of effective agents for the protection of
chondrocytes to treat OA is crucial. However, currently no
effective medical therapy is supported for OA except physiotherapy
and surgery. In the present study, we found that PSO enhanced
chondrocyte viability in a dose-and time-dependent manner as
determined by MTT assay, indicating that PSO is a potential
therapeutic agent for the treatment of OA. Thus, we further
investigated the mechanisms involved in the chondrocyte
proliferation mediated by PSO.
The Wnt/β-catenin signaling pathway plays a vital
role in a number of cellular events including cell proliferation,
migration and differentiation. The Wnt/β-catenin signaling pathway
is important in the regulation of proliferation. It regulates
chondrogenes at different stages in different ways, and plays a
crucial role in the physiopathology of OA (22–24). Altering the expression of genes
and proteins in articular chondrocytes is correlated with the
pathological process of OA (25).
Several molecular components of the Wnt/β-catenin signaling pathway
have been shown to be associated with OA, including Wnt-4,
Frizzled-2, β-catenin and GSK-3β. The Wnt/β-catenin signaling
pathway is triggered by binding of Wnt ligands to frizzled
receptors and co-receptors, low-density lipoprotein receptors
(LRP)5/6. GSK-3 phosphorylates LRP5/6 which results in recruitment
of the axin complex to this co-receptor and leads to reduced
degradation of β-catenin. Next, the resultant accumulated
cytoplasmic β-catenin translocates to the nucleus (26,27). Finally, this activates target
genes, such as cyclin D1, a positive effector of the G1/S
transition that is particularly involved in cell proliferation
(11,28). In the present study, we
investigated the activity of PSO in the promotion of chondrocyte
proliferation. Our results showed that the expression of Wnt-4,
Frizzled-2, β-catenin and cyclin D1 in PSO-treated chondrocytes was
markedly increased, whereas the expression of GSK-3β was
significantly decreased. Additionally, PSO contributed to increased
β-catenin nuclear translocation in the chondrocytes. These results
revealed that PSO activates the Wnt/β-catenin signaling pathway to
promote chondrocyte proliferation.
DKK-1, an inhibitor of the Wnt/β-catenin signaling
pathway, antagonizes WNT signaling by interacting with co-receptor
LRP leading to LRP endocytosis (29). DKK-1 inhibits the Wnt signaling
pathway by binding to LRP5/6 on target cells. Following the
inhibition, GSK-3 directly phosphorylates the transcriptional
regulator β-catenin, marking it for proteosomal degradation, and
then partly decreasing the expression of β-catenin and cyclin D1
(30). Our results revealed that
the expression of β-catenin and cyclin D1 was partly inhibited by
DKK-1, which further demonstrated that PSO promoted chondrocyte
proliferation via the Wnt/β-catenin signaling pathway.
In conclusion, the present study demonstrated that
PSO promoted chondrocyte proliferation by regulating the
Wnt/β-catenin signaling pathway. Further studies using animal
models should be carried out to investigate the detailed mechanisms
of the Wnt/β-catenin signaling pathway in the pathogenesis of OA.
In addition, the present study also found that PSO increased the
expression of Col-II in chondrocytes, while this was partly
inhibited by DKK-1. This result indicated that PSO has a positive
effect on preventing cartilage degradation by increasing the
expression of Col-II, a major component of the cartilage matrix.
Future studies using animal models should investigate the detailed
mechanism of the Wnt/β-catenin signaling pathway in the
pathogenesis of OA.
Acknowledgments
The present study was supported by the National
Natural Science Foundation of China (grant no. 81373818), the Key
Project of Fujian Provincial Department of Science and Technology
Department (grant no. 2014Y0064), the Natural Science Foundation of
Fujian Province (grant nos. 2014J01357 and 2016J01395) and the
Developmental Fund of Chen Keji Integrative Medicine (grant no.
CKJ2015009).
References
|
1
|
Pereira D, Ramos E and Branco J:
Osteoarthritis. Osteoarthritis Acta Med Port. 28:99–106. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Blalock D, Miller A, Tilley M and Wang J:
Joint instability and osteoarthritis. Clin Med Insights Arthritis
Musculoskelet Disord. 8:15–23. 2015.PubMed/NCBI
|
|
3
|
Thomas JT, Schneider BS, Frank EL and
Krizan SJ: Cartilage repair and replacement in the knee. Trends
Biotechnol. 31:665–667. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Chen C, Tambe DT, Deng L and Yang L:
Biomechanical properties and mechanobiology of the articular
chondrocyte. Am J Physiol Cell Physiol. 305:C1202–C1208. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Tonge DP, Pearson MJ and Jones SW: The
hallmarks of osteoarthritis and the potential to develop
personalised disease-modifying pharmacological therapeutics.
Osteoarthritis Cartilage. 22:609–621. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Schulze-Tanzil G: Activation and
dedifferentiation of chondrocytes: Implications in cartilage injury
and repair. Ann Anat. 191:325–338. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Pap T and Korb-Pap A: Cartilage damage in
osteoarthritis and rheumatoid arthritis - two unequal siblings. Nat
Rev Rheumatol. 11:606–615. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Aman A, Nguyen M and Piotrowski T:
Wnt/β-catenin dependent cell proliferation underlies segmented
lateral line morphogenesis. Dev Biol. 349:470–482. 2011. View Article : Google Scholar
|
|
9
|
Monroe DG, McGee-Lawrence ME, Oursler MJ
and Westendorf JJ: Update on Wnt signaling in bone cell biology and
bone disease. Gene. 492:1–18. 2012. View Article : Google Scholar :
|
|
10
|
Ryu JH, Kim SJ, Kim SH, Oh CD, Hwang SG,
Chun CH, Oh SH, Seong JK, Huh TL and Chun JS: Regulation of the
chondrocyte phenotype by beta-catenin. Development. 129:5541–5550.
2002. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Bougault C, Priam S, Houard X, Pigenet A,
Sudre L, Lories RJ, Jacques C and Berenbaum F: Protective role of
frizzled-related protein B on matrix metalloproteinase induction in
mouse chon-drocytes. Arthritis Res Ther. 16:R1372014. View Article : Google Scholar
|
|
12
|
Lu H, Zhang L, Liu D, Tang P and Song F:
Isolation and purification of psoralen and isopsoralen and their
efficacy and safety in the treatment of osteosarcoma in nude rats.
Afr Health Sci. 14:641–647. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Wong RW and Rabie AB: Effect of psoralen
on bone formation. J Orthop Res. 29:158–164. 2011. View Article : Google Scholar
|
|
14
|
Tang DZ, Yang F, Yang Z, Huang J, Shi Q,
Chen D and Wang YJ: Psoralen stimulates osteoblast differentiation
through activation of BMP signaling. Biochem Biophys Res Commun.
405:256–261. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Li J, Zhu A, Xie W and Zheng H: Study of
the mechanism of benefiting kidney herbs in the prevention and
treatment of osteoarthritis in vitro. Chin J Osteoporos.
22:877–882. 2016.In Chinese.
|
|
16
|
Li H, Li X, Liu G, Chen J, Weng X, Liu F,
Xu H, Liu X and Ye H: Bauhinia championi (Benth.) Benth.
polysaccharides upregulate Wnt/β-catenin signaling in chondrocytes.
Int J Mol Med. 32:1329–1336. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Weng X, Lin P, Liu F, Chen J, Li H, Huang
L, Zhen C, Xu H, Liu X, Ye H, et al: Achyranthes bidentata
polysaccharides activate the Wnt/β-catenin signaling pathway to
promote chondrocyte proliferation. Int J Mol Med. 34:1045–1050.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Yu F, Li X, Cai L, Li H, Chen J, Wong X,
Xu H, Zheng C, Liu X and Ye H: Achyranthes bidentata
polysaccharides induce chon-drocyte proliferation via the promotion
of the G1/S cell cycle transition. Mol Med Rep. 7:935–940. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Maturana JL, Niechi I, Silva E, Huerta H,
Cataldo R, Härtel S, Barros LF, Galindo M and Tapia JC:
Transactivation activity and nucleocytoplasmic transport of
β-catenin are independently regulated by its C-terminal end. Gene.
573:115–122. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Hunter DJ and Felson DT: Osteoarthritis.
BMJ. 332:639–642. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Dreier R: Hypertrophic differentiation of
chondrocytes in osteoarthritis: The developmental aspect of
degenerative joint disorders. Arthritis Res Ther. 12:2162010.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Tao HY, He B, Liu SQ, Wei AL, Tao FH, Tao
HL, Deng WX, Li HH and Chen Q: Effect of carboxymethylated chitosan
on the biosynthesis of NGF and activation of the Wnt/β-catenin
signaling pathway in the proliferation of Schwann cells. Eur J
Pharmacol. 702:85–92. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Sassi N, Laadhar L, Allouche M, Achek A,
Kallel-Sellami M, Makni S and Sellami S: WNT signaling and
chondrocytes: From cell fate determination to osteoarthritis
physiopathology. J Recept Signal Transduct Res. 34:73–80. 2014.
View Article : Google Scholar
|
|
24
|
Sassi N, Laadhar L, Allouche M,
Zandieh-Doulabi B, Hamdoun M, Klein-Nulend J, Makni S and Sellami
S: The roles of canonical and non-canonical Wnt signaling in human
de-differentiated articular chondrocytes. Biotech Histochem.
89:53–65. 2014. View Article : Google Scholar
|
|
25
|
Li X, Chen J, Liang W, Li H, Liu F, Weng
X, Lin P, Chen W, Zheng C, Xu H, et al: Bushen Zhuangjin Decoction
promotes chondrocyte proliferation by stimulating cell cycle
progression. Exp Ther Med. 9:839–844. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Zeng X, Tamai K, Doble B, Li S, Huang H,
Habas R, Okamura H, Woodgett J and He X: A dual-kinase mechanism
for Wnt co-receptor phosphorylation and activation. Nature.
438:873–877. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Doble BW, Patel S, Wood GA, Kockeritz LK
and Woodgett JR: Functional redundancy of GSK-3alpha and GSK-3beta
in Wnt/beta-catenin signaling shown by using an allelic series of
embryonic stem cell lines. Dev Cell. 12:957–971. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
MacDonald BT, Tamai K and He X:
Wnt/β-catenin signaling: Components, mechanisms, and diseases. Dev
Cell. 17:9–26. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Mao B and Niehrs C: Kremen2 modulates
Dickkopf2 activity during Wnt/LRP6 signaling. Gene. 302:179–183.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Liu C, Li Y, Semenov M, Han C, Baeg GH,
Tan Y, Zhang Z, Lin X and He X: Control of beta-catenin
phosphorylation/degradation by a dual-kinase mechanism. Cell.
108:837–847. 2002. View Article : Google Scholar : PubMed/NCBI
|