Introduction
Gastric cancer (GC) is the second most common type
of malignancy worldwide. In developed western countries, the 5-year
survival rate for GC is as low as 10-19% (1). In China, GC is the most common type
of malignancy according to the number of cases diagnosed annually
(2). It is currently elusive what
factors contribute to the development, progression and metastasis
of GC in geographic areas with highly prevalence, including certain
cities in Northwest China (3,4).
GC includes two distinct morphological subtypes: Gastric intestinal
type adenocarcinoma and diffuse gastric adenocarcinoma (5-7).
In addition to tumor protein 53 mutations, studies have reported
frequent inactivating mutations in genes associated with cell
adhesion and chromatin remodeling (8,9).
Although certain essential factors that may serve as therapeutic
tagets were identified in recent years, effective methods to treat
GC or monitor disease progression in the clinic are still lacking
(10-13).
The LIM domain only (LMO) protein family contains
four members: LMO1, LMO2, LMO3 and LMO4. Although LMO proteins lack
DNA-binding activity, they were reported to be involved in the
transcriptional regulation of specific target genes in
collaboration with other transcription factors (2). Genetic analyses demonstrated that
LMO1 and LMO2 contribute to the genesis of immature and aggressive
T-cell leukemia (14), while LMO4
has been reported to be implicated in the development of breast
cancer (15,16). LMO3 forms a complex with the
neuronal-specific basic helix-loop-helix (HLH) transcription
factor, HLH protein 2, which is also overexpressed in neuroblastoma
with unfavorable prognosis compared with that in neuroblastoma with
favorable prognosis. Furthermore, LMO3 has been reported to have
important roles in numerous cancer types, including neuroblastoma
(17,18) and lung cancer (19,20).
The present study reported that LMO3 expression was
significantly upregulated in GC tissues and was closely associated
with the clinicopathological characteristics and patient prognosis.
Knockdown of LMO3 significantly suppressed the invasion and
proliferation of GC cells. Furthermore, the effects of LMO3 on the
invasion and proliferation of GC cells were determined to be
dependent on Akt/mammalian target of rapamycin (mTOR) and
Akt/glycogen synthase kinase (GSK)3β signaling.
Materials and methods
Cell culture
Human GC cell lines, including AGS, BGC-823, HGC-27,
MGC-803, MKN-45 and SGC-7901, were purchased from the Cell Bank of
the Chinese Academy of Sciences (Shanghai, China). BGC-823,
MGC-803, MKN-45 and SGC-7901 cells were cultured in RPMI-1640
medium (Gibco, Thermo Fisher Scientific, Inc., Waltham, MA, USA)
supplemented with 10% (v/v) fetal calf serum (FCS) (Gibco, Thermo
Fisher Scientific, Inc.) and 1% antibiotics. AGS was cultured in
DMEM F-12 medium (Gibco, Thermo Fisher Scientific, Inc.)
supplemented with 10% FCS and 1% antibiotics. HGC-27 was cultured
in RPMI-1640 medium supplemented with 20% FCS and 1% antibiotics.
All of the GC cells were cultured at 37°C in a humidified
atmosphere containing 5% CO2.
Clinical samples
Human gastric tumor (n=19) and normal tissues
(n=15), in 10 cases of which gastric tumor and normal tissues were
paired, were obtained from patients who were admitted between
January 2012 and December 2016 at the Department of Bone Tumor
(collected form the Department of General Surgery, at which the
surgery was performed), Yantai Mountain Hospital (Yantai, China).
All of the patients provided written informed consent prior to
enrollment, and the study was approved by the Research Ethics
Committee of Yantai Mountain Hospital (Yantai, China). In addition,
a human tissue microarray, which was also obtained from the
Department of Bone Tumor (collected form the Department of General
Surgery), Yantai Mountain Hospital (Yantai, China) was used, which
contained GC samples and paired normal tissues from 164 cases.
KMplot analysis
The Affy ID of the dataset used in the presnet study
was 204424_s_at. The best cut-off value was auto selected using
KMplot (http://kmplot.com/analysis/index.php?p=service&cancer=gastric).
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
Total RNA was extracted using TRIzol (Takara Bio
Inc., Otsu, Japan) and reverse transcribed with a PrimeScript
RT-PCR kit (Perfect Real-Time; Clontech; Takara Bio Inc.).
Real-time qPCR analyses were performed with SYBR Premix Ex Taq
(Takara Bio Inc.) on a 7500 Real-Time PCR System (Applied
Biosystems; Thermo Fisher Scientific, Inc., Waltham, MA, USA) at
the recommended thermal cycling settings: One initial cycle at 95°C
for 30 sec, followed by 40 cycles of 5 sec at 95°C and 31 sec at
60°C. Results were quantified using the 2−ΔΔCq method
was used (21). The primer
sequences used were as follows: LMO3 forward,
5′-TCTGAGGCTCTTTGGTGTAACG-3′ and reverse,
5′-CCAGGTGGTAAACATTGTCCTTG-3′; and β-actin forward,
5′-CTCCATCCTGGCCTCGCTGT-3′ and reverse,
5′-GCTGTCACCTTCACCGTTCC-3′.
Western blot analysis
Cells were lysed in lysis buffer (50 mM Tris-HCl,
150 mM NaCl, 1% Triton X-100, 1 mM each MgCl2,
MnCl2 and CaCl2, 1 mM phenylmethylsulfonyl
fluoride and 10 mM sodium fluoride; Sangon, Shanghai, China). The
protein concentration was determined using the BCA method. A total
of 20 μg protein was loaded per lane and proteins were
separated by 6-12% SDS-PAGE under reducing conditions and
transferred onto a nitrocellulose membrane (Thermo Fisher
Scientific, Inc.). The membrane was blocked in PBS/Tween-20
containing 5% bovine serum albumin (Sangon Biotech Co., Ltd.,
Shanghai, China), followed by incubation with the antibodies for
LMO3 (1:1,000, cat. no. ab69573; Abcam, Cambridge, MA, USA),
phospho-Akt (1:2,000, cat. no. 4060), total-Akt (1:2,000, cat. no.
4691), phospho-mTOR (1:2,000, cat. no. 5536), total-mTOR (1:2,000,
cat. no. 2983), phospho-GSK3β (1:2,000, cat. no. 5558), total-GSK3β
(1:2,000, cat. no. 12456; all from Cell Signaling Technology, Inc.,
Danvers, MA, USA) and GAPDH (1:1,000, cat. no. 9001-50-7;
Sigma-Aldrich; Merck KGaA, Darmstadt, Germany) at 4°C overnight.
Subsequently, the membrane was washed and incubated with
IRDye® 680LT Goat anti-Rabbit IgG (H+L, 1:10,000, cat.
no. P/N 925-68021) or IRDye® 800CW Goat anti-Mouse IgG
(H+L, 1:10,000, cat. no. P/N 925-32210; LI-COR Biosciences,
Lincoln, NE, USA) at room temperature for 1 h. The
fluorescently-labeled secondary antibodies were directly detected
with the Odyssey imaging system (LI-COR Biosciences).
Small interfering (si)RNA
transfection
siRNA duplexes for LMO3 were produced by Genepharma
(Shanghai, China). The LMO3 siRNA and scrambled siRNA were
purchased from Abnova (cat. no. H00055885-R01; Abnova, Taiwan,
China). Transfection steps were performed according to the
manufacturer's protocols of the X-tremeGENE siRNA transfection
reagent (cat. no. 4476093001; Sigma-Aldrich; Merck KGaA).
Recombinant (r)LMO3 protein and
inhibitors
rLMO3 protein was purchased from Abnova (Taipei,
Taiwan). The inhibitor of mTOR (dactolisib) and the inhibitor of
GSK3β (CHIR-98014) were purchased from Selleck Chemicals (Houston,
TX, USA). rLMO3 protein was added to BGC-823 and HGC-27 cells, and
dactolisib or CHIR-98014 were added to above cells 2 h later. All
cells were incubated at 37°C.
Invasion assay
MGC-803, SGC-7901, BGC-823 or HGC-27 cells were
detached, resuspended in serum-free RPMI-1640 medium, and seeded at
2×104 cells per well in 100 μl in 8-μm
Transwell inserts (EMD Millipore, Billerica, MA, USA) with
membranes coated in Matrigel (BD Biosciences, Franklin Lakes, NJ,
USA) on top of a 24-well plate. RPMI-1640 medium containing 5% FBS
was added to the bottom chamber. Cells were incubated at 37°C for
48 h. Subsequently, filters were fixed and stained with 0.1% (w/v)
crystal violet at room temperature for 20 min. Non-invading cells
were removed from the upper side of the membrane and invaded cells
on the lower side were counted under a microscope at a
magnification of ×400. At least three grids (each grid was
restricted at a field ~982×722 mm, with a magnification of ×400)
per field were counted and the experiments were repeated twice.
Cell viability assay
Cell viability was determined using a standard Cell
Counting Kit-8 (CKK-8) assay. MGC-803, SGC-7901, BGC-823 or HGC-27
cells were seeded into 96-well plates (100 μl per well) at a
density of 2×104 cells per ml, and the total number of
cells seeded in each well was 2×103. MGC-803 and
SGC-7901 cells were transfected with LMO3 siRNA or scrambled siRNA
before these cells were seeded. However, BGC-823 and HGC-27 cells
were treated with rLMO3 protein, dactolisib or CHIR-98014 after the
cells adhered to the bottom of dishes. After incubation for at 12,
24, 48 and 72 h, 10 μl CKK-8 reagent (Dojindo, Kumamoto,
Japan) was added to each well. After 2 h of incubation at 37°C, the
optical density was measured using a microplate reader at a
wavelength of 450 nm.
Statistical analysis
Values are expressed as the mean ± standard error of
the mean. Statistical analyses were performed using SPSS 16.0 for
windows (SPSS, Inc., Chicago, IL, USA). Survival time was analyzed
with the Kaplan-Meier method. The association between LMO3
expression and the clinicopathological features of patients with
gastric cancer was evaluated using Pearson's Chi-square test.
One-way analysis of variance was used for comparison between
groups. Multiple comparison between the groups was performed using
Student-Newman-Keuls post hoc method. P<0.05 was considered to
indicate a statistically significant difference.
Results
LMO3 expression is closely associated
with clinicopatho-logical characteristics and prognosis of GC
patients
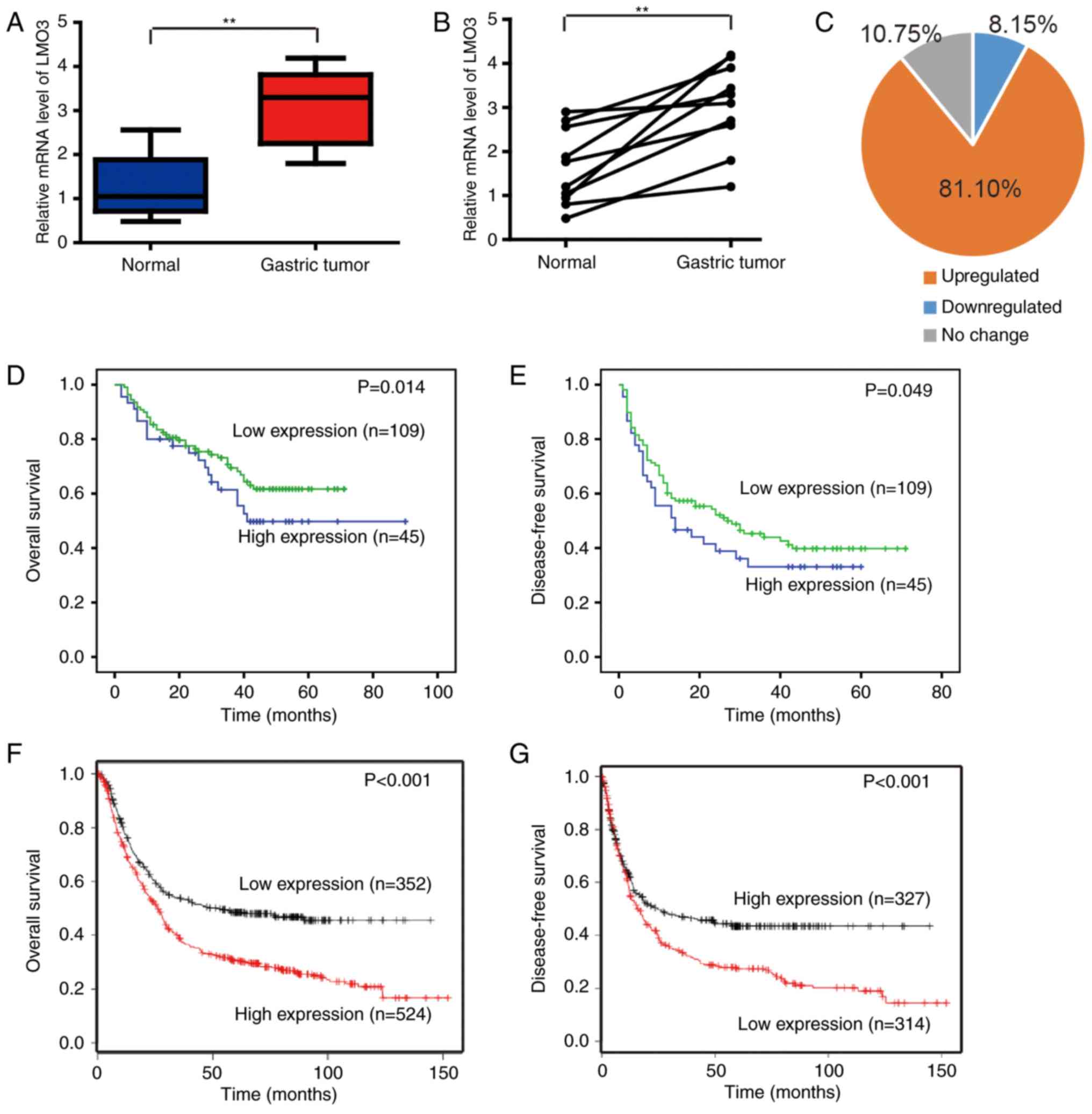
To investigate the expression of LMO3 in GC tissues,
19 gastric tumor and 15 normal gastric tissues were collected and
analyzed by RT-qPCR. The expression levels of LMO3 were
significantly upregulated in GC tissues (Fig. 1A). In 10 paired gastric tumor and
normal tissues within this cohort, LMO3 expression was also
upregulated in GC tissues (Fig.
1B).
A GC tissue microarray (n=164) was then used to
investigate the correlation between LMO3 expression and
clinicopathological characteristics and patient prognoses. Scoring
of immunohistochemical staining was conducted according to the
ratio and intensity of positive-staining cells: 0-35% was
designated as the low expression group; >36% was designated as
the high expression group. The age of 62 years, the median age of
this patient cohort, was set as a cut-off, based on which the
patients were stratified into two groups. Notably, no significant
difference was indicated among the age groups. It was identified
that the expression of LMO3 was upregulated in 81.10% of GC tissues
(Fig. 1C). Furthermore, the
expression of LMO3 was closely associated with the tumor size,
differentiation, vascular embolism and tumor-nodes-metastasis (TNM)
stage (Table I), and high
expression of LMO3 was positively correlated with poor overall
survival (OS; P=0.014) or disease-free survival (DFS; P=0.049;
Fig. 1D and E, respectively). The
above results were confirmed by analyzing the datasets (Affy ID
204424_s_at) with the software Kaplan Meier-plotter (http://kmplot.com/analysis/index.php?p=service&cancer=gastric),
revealing that LMO3 expression was positively correlated with poor
OS (P<0.001) and DFS (P<0.001) of GC patients (Fig. 1F and G).
 | Table ICorrelation of clinicopathological
factors with LMO3 expression. |
Table I
Correlation of clinicopathological
factors with LMO3 expression.
| Variable | LMO3
| P-value |
|---|
| High (n) (%) | Low (n) (%) |
|---|
| Age (years) | | | 0.734 |
| ≤62 | 24 (14.63) | 53 (32.32) | |
| >62 | 25 (15.24) | 62 (37.80) | |
| Gender | | | 0.668 |
| Female | 6 (3.66) | 17 (10.37) | |
| Male | 43 (26.22) | 98 (59.76) | |
| Smoking history | | | 0.276 |
| Yes | 37 (22.56) | 77 (46.95) | |
| No | 12 (7.32) | 38 (23.17) | |
| Lauren subtype | | | 0.787 |
| Diffuse | 23 (14.11) | 61 (37.42) | |
| Intestine | 25 (15.34) | 54 (33.13) | |
| Location | | | 0.447 |
| Upper stomach | 11 (6.71) | 20 (12.20) | |
| Middle stomach | 22 (13.41) | 65 (39.63) | |
| Lower stomach | 13 (7.93) | 24 (14.63) | |
| Remnant stomach | 3 (1.83) | 6 (3.66) | |
| Tumor size (cm) | | | <0.001a |
| ≤5 | 17 (10.37) | 78 (47.56) | |
| >5 | 32 (19.51) | 37 (22.56) | |
| Differentiation | | | 0.004a |
| Well | 1 (0.61) | 2 (1.23) | |
| Moderate | 11 (6.75) | 59 (36.20) | |
| Poor | 36 (22.09) | 54 (33.13) | |
| Vascular
embolism | | | 0.001a |
| Yes | 35 (21.34) | 105 (64.02) | |
| No | 14 (8.54) | 10 (6.10) | |
| TNM stage | | | 0.004a |
| I | 30 (18.40) | 87 (53.37) | |
| II | 4 (2.45) | 18 (11.04) | |
| III/IV | 14 (8.59) | 10 (6.13) | |
Knockdown of LMO3 in MGC-803 and SGC-7901
cells
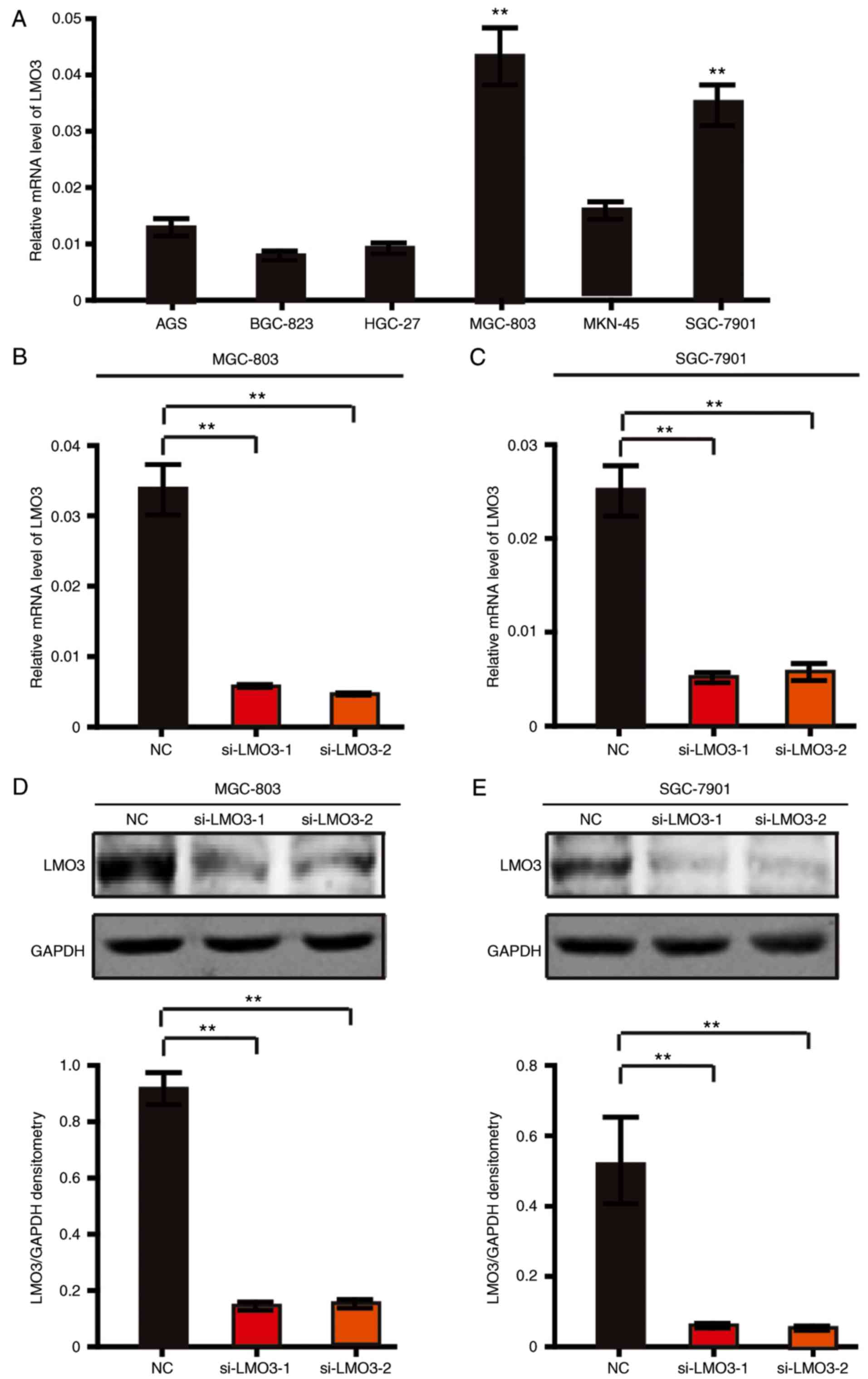
To further investigate the biological functions of
LMO3 in GC, the expression level of LMO3 were first detected in 6
GC cell lines. As presented in Fig.
2A, LMO3 was highly expressed in MGC-803 and SGC-7901 cells
compared with the other cell lines. Therefore, the MGC-803 and
SGC-7901 cell lines were subjected to knockdown of LMO3 by using
siRNA (labeled as si-LMO3-1 and si-LMO3-2). Through RT-qPCR and
western blot analysis, it was demonstrated that LMO3 was
successfully silenced in MGC-803 and SGC-7901 cells (Fig. 2B–E).
LMO3 knockdown suppresses the invasion
and proliferation of GC cells
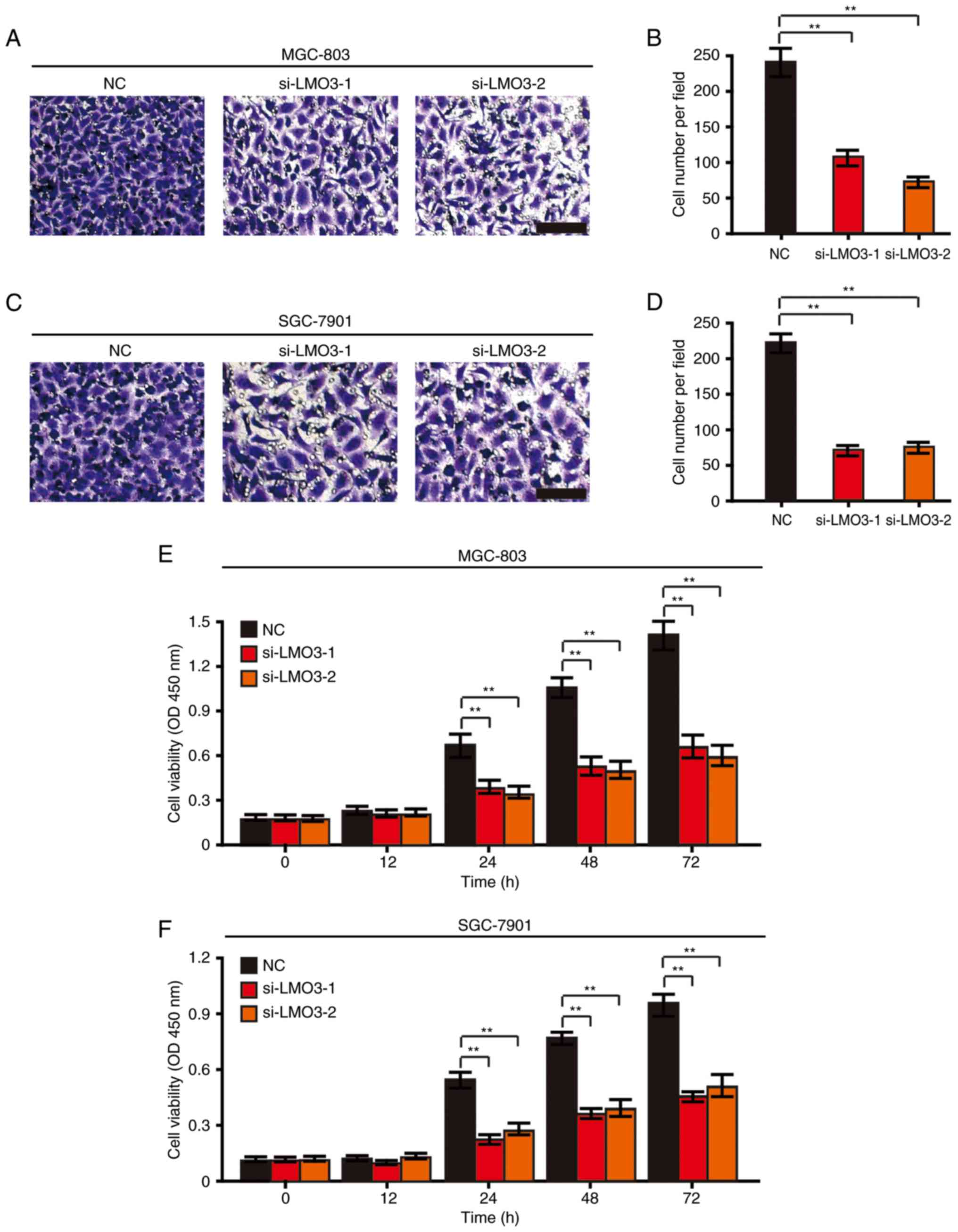
The present study investigated the role of LMO3 in
the invasion of GC cells. A Transwell Matrigel invasion assay
demonstrated that knockdown of LMO3 suppressed the invasive
capacity of MGC-803 (Fig. 3A and
B) and SGC-7901 (Fig. 3C and
D) cells after 48 h.
Furthermore, a CCK-8 cell viability assay was
employed to investigate the role of LMO3 in the proliferation of GC
cells. It was revealed that the viability of MGC-803 and SGC-7901
cells was significantly suppressed by knockdown of LMO3 at the 24,
48 and 72 h time-points (Fig. 3E and
F).
LMO3 knockdown suppresses Akt, mTOR and
GSK3β signaling in GC cells
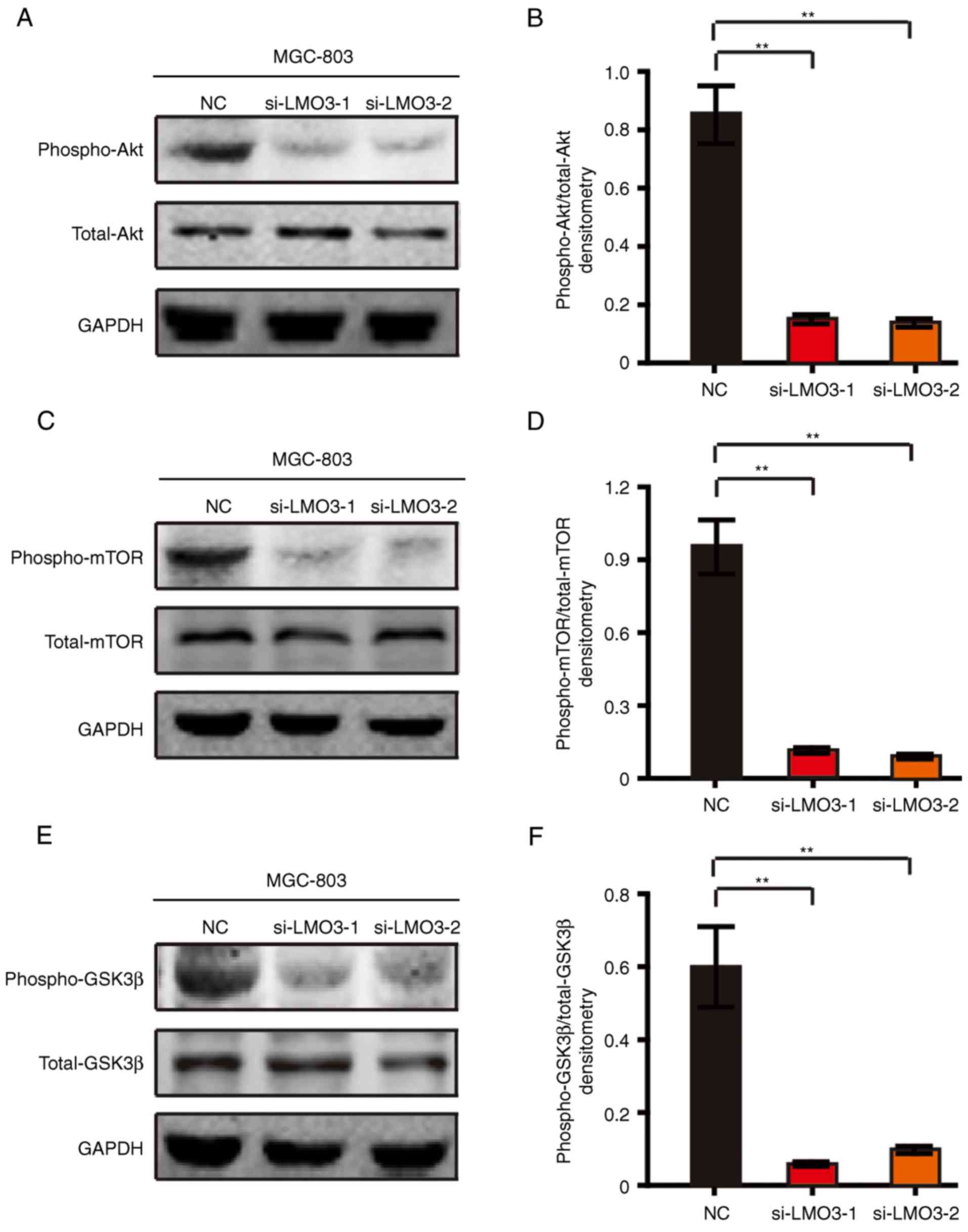
To investigate the underlying mechanism of the role
of LMO3 in GC, the Akt/mTOR and Akt/GSK3β signaling pathways were
assessed in MGC-803 cells by western blot analysis. Of note, LMO3
knockdown significantly suppressed the phosphorylation of Akt
(Fig. 4A and B). The
phosphorylation of mTOR, a downstream signaling event of Akt, was
also suppressed by silencing LMO3 (Fig. 4C and D). Furthermore, the
phosphorylation of GSK3β, another downstream signaling event of
Akt, was also suppressed by silencing LMO3 (Fig. 4E and F).
Taken together, the abovementioned results suggested
that LMO3 promotes the invasion and proliferation of GC cells
through Akt/mTOR and Akt/GSK3β signaling.
LMO3 promotes GC cell invasion through
Akt/mTOR signaling, and supports GC cell proliferation via Akt/mTOR
and Akt/GSK3β signaling
Since mTOR and GSK3β are different downstream
signaling events of Akt, the effects of LMO3 on GC cell invasion
and proliferation were investigated in the presence of dactolisib
(inhibitor of mTOR) and CHIR-98014 (inhibitor of GSK3β).
Furthermore, rLMO3 protein was added to BGC-823 and HGC-27 cells,
which had low LMO3 expression levels, and dactolisib and CHIR-98014
were added after 2 h. It was revealed that dactolisib treatment
abrogated rLMO3 protein-induced GC cell invasion (Fig. 5A and B). Furthermore, dactolisib
and CHIR-98014 abrogated rLMO3 protein-induced proliferation
(Fig. 5C and D).
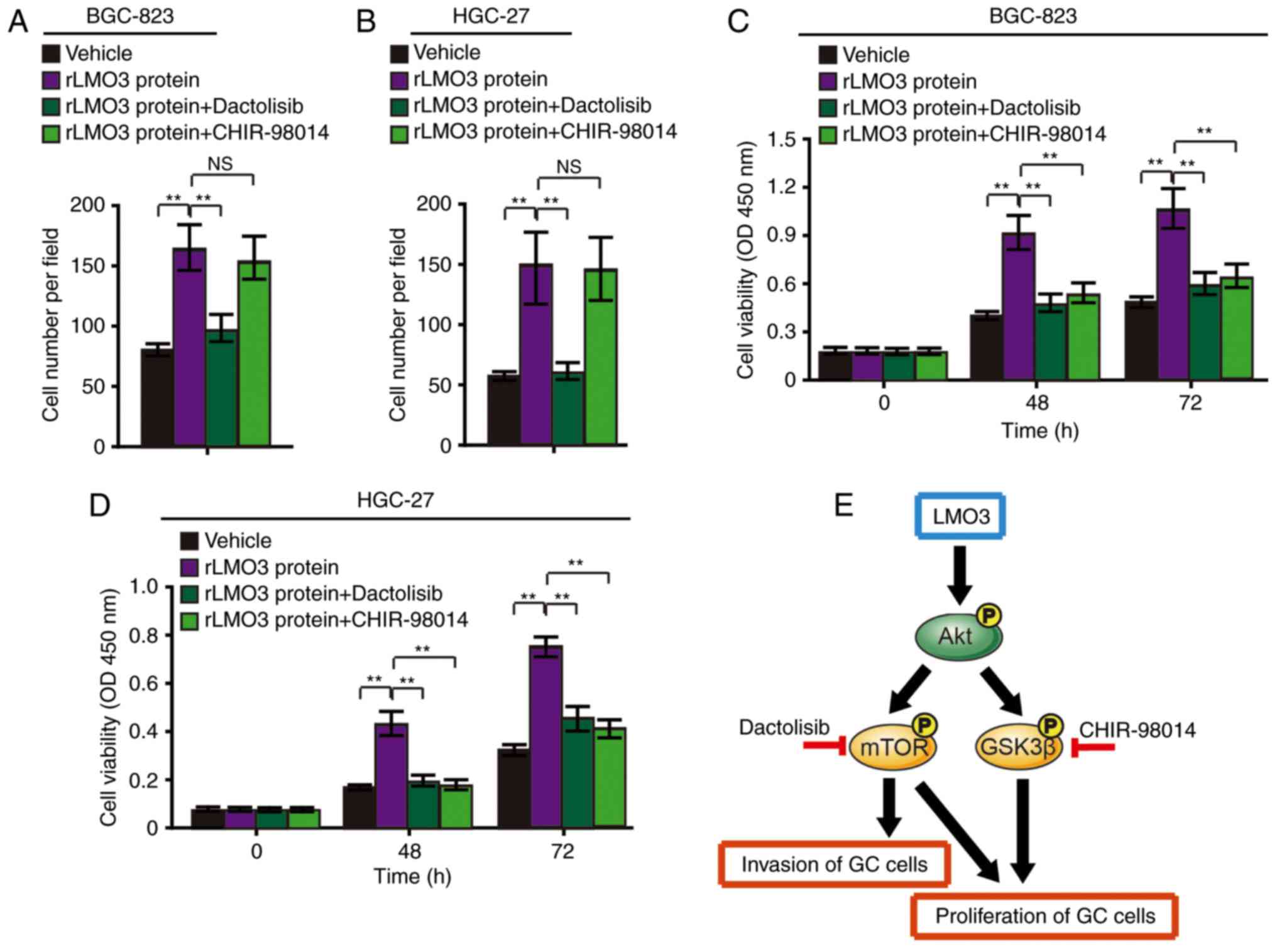 | Figure 5LMO3 promotes GC-cell invasion via
Akt-mTOR signaling and promotes GC cell proliferation via Akt-mTOR
and Akt-GSK3β signaling. (A) BGC-823 and (B) HGC-27 cells were
treated with 50 nM rLMO3 protein, 50 nM rLMO3 protein plus 50 nM
Dactolisib (an inhibitor of mTOR), or 50 nM rLMO3 protein plus 50
nM CHIR-98014 (an inhibitor of GSK3β). Cell invasion was analyzed
after 48 h. (C) BGC-823 and (D) and HGC-27 cells were subjected to
the abovementioned treatments and cell viability was detected with
a Cell Counting Kit-8 at 0, 48 and 72 h time-points.
**P<0.01. (E) Schematic depicting the mechanism of
LMO3-induced GC cell invasion and proliferation. rLMO3, recombinant
LIM domain only 3; NC, negative control; siRNA, small interfering
RNA; mTOR, mammalian target of rapamycin; GSK, glycogen synthase
kinase; GC, gastric cancer; P, phosphate; NS, no significance; OD,
optical density. |
These results indicated that LMO3-induced GC cell
invasion is mainly dependent on Akt/mTOR signaling. Furthermore,
LMO3-induced proliferation is mainly dependent on Akt/mTOR and
Akt/GSK3β signaling. The signaling cascades are outlined in a
schematic in Fig. 5E.
Discussion
Numerous studies have assessed LMO3 in various
cancer types. It has been reported that LMO3 collaborates with HEN2
to enhance neuroblastoma cell growth (17,18). Notably, microRNA-630 targets LMO3
to regulate lung cancer cell growth and metastasis (19,20). However, the detailed biological
functions of LMO3 in GC and the underlying mechanisms have remained
to be elucidated. In the present study, the exact roles of LMO3 in
GC were deeply investigated for the first time, to the best of our
knowledge. By analyzing GC tissue microarray and KMplot datasets,
it was identified that the expression of LMO3 was closely
associated with the tumor size, differentiation, vascular embolism
and TNM stage, as well as poor patient prognoses. These results
suggested that LMO3 may have important roles in the development of
GC.
We further revealed the biological functions of LMO3
in GC. The invasion and cell viability of GC cells were
significantly suppressed by LMO3 knockdown. These results indicated
that LMO3 is involved in the invasion and proliferation of GC
cells.
Invasion and metastasis are major concerns during
the prognosis and progression of cancer. The Akt/mTOR pathway is
pivotal in modulating the invasion and migration of tumor cells
(22,23). The present study indicated that
knockdown of LMO3 decreases the phosphorylation of Akt and mTOR. By
using an inhibitor of mTOR, dactolisib, it was further determined
that LMO3-induced GC-cell invasion and proliferation are dependent
on Akt-mTOR signaling.
Furthermore, phosphoinositide-3 kinase/Akt/GSK3β
signaling has key roles in the regulation of numerous biological
functions in various cancer types, including cell migration,
invasion, proliferation and apoptosis. The present study identified
that LMO3 knockdown decreased the phosphorylation of GSK3β, and an
inhibitor of GSK3β, CHIR-98014, was able to abrogate rLMO3
protein-induced proliferation. Furthermore, the present findings
suggested that LMO3-induced GC cell proliferation was dependent on
the Akt/GSK3β pathway.
In conclusion, the present study revealed that LMO3
has an important role in GC cell invasion and proliferation. LMO3
promotes GC cell invasion and proliferation through Akt/mTOR and
Akt/GSK3β signaling. LMO3 may be used as a potential prognostic or
therapeutic target for GC in the future.
Notes
[1] Competing
interests
The authors declare that they have no competing
interests.
References
|
1
|
Hansson LE, Sparén P and Nyrén O: Survival
in stomach cancer is improving: Results of a nationwide
population-based Swedish study. Ann Surg. 230:162–169. 1999.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Sun X, Mu R, Zhou Y, Dai X, Qiao Y, Zhang
S, Huangfu X, Sun J, Li L and Lu F: (1990–1992) mortality of
stomach cancer in China). Zhonghua Zhongliu Zazhi. 24:4–8. 2002.In
Chinese.
|
|
3
|
Werner M, Becker KF, Keller G and Höfler
H: Gastric adenocarcinoma: Pathomorphology and molecular pathology.
J Cancer Res Clin Oncol. 127:207–216. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Becker KF, Keller G and Hoefler H: The use
of molecular biology in diagnosis and prognosis of gastric cancer.
Surg Oncol. 9:5–11. 2000. View Article : Google Scholar
|
|
5
|
Wong SS, Kim KM, Ting JC, Yu K, Fu J, Liu
S, Cristescu R, Nebozhyn M, Gong L, Yue YG, et al: Genomic
landscape and genetic heterogeneity in gastric adenocarcinoma
revealed by whole-genome sequencing. Nat Commun. 5:54772014.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Kang G, Hwang WC, Do IG, Wang K, Kang SY,
Lee J, Park SH, Park JO, Kang WK, Jang J, et al: Exome sequencing
identifies early gastric carcinoma as an early stage of advanced
gastric cancer. PLoS One. 8:e827702013. View Article : Google Scholar :
|
|
7
|
Hu B, El Hajj N, Sittler S, Lammert N,
Barnes R and Meloni-Ehrig A: Gastric cancer: Classification,
histology and application of molecular pathology. J Gastrointest
Oncol. 3:251–261. 2012.PubMed/NCBI
|
|
8
|
Zang ZJ, Cutcutache I, Poon SL, Zhang SL,
McPherson JR, Tao J, Rajasegaran V, Heng HL, Deng N, Gan A, et al:
Exome sequencing of gastric adenocarcinoma identifies recurrent
somatic mutations in cell adhesion and chromatin remodeling genes.
Nat Genet. 44:570–574. 2012. View
Article : Google Scholar : PubMed/NCBI
|
|
9
|
Cancer Genome Atlas Research Network:
Comprehensive molecular characterization of gastric adenocarcinoma.
Nature. 513:202–209. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Ding Y, Yang Q, Wang B, Ye G and Tong X:
The correlation of MGMT promoter methylation and
clinicopathological features in gastric cancer: A systematic review
and meta-analysis. PLoS One. 11:e01655092016. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Baroudi O and Benammar-Elgaaied A:
Involvement of genetic factors and lifestyle on the occurrence of
colorectal and gastric cancer. Crit Rev Oncol Hematol. 107:72–81.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Zhou J, Shen J, Seifer BJ, Jiang S, Wang
J, Xiong H, Xie L, Wang L and Sui X: Approaches and genetic
determinants in predicting response to neoadjuvant chemotherapy in
locally advanced gastric cancer. Oncotarget. 8:30477–30494.
2017.
|
|
13
|
Qi J, Zhang P, Wang Y, Chen H and Li Y:
Does total gastrectomy provide better outcomes than distal subtotal
gastrectomy for distal gastric cancer? A systematic review and
meta-analysis. PLoS One. 11:e01651792016. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Rabbitts TH: LMO T-cell translocation
oncogenes typify genes activated by chromosomal translocations that
alter transcription and developmental processes. Genes Dev.
12:2651–2657. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Sum EY, Segara D, Duscio B, Bath ML, Field
AS, Sutherland RL, Lindeman GJ and Visvader JE: Overexpression of
LMO4 induces mammary hyperplasia, promotes cell invasion, and is a
predictor of poor outcome in breast cancer. Proc Natl Acad Sci USA.
102:7659–7664. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Visvader JE, Venter D, Hahm K, Santamaria
M, Sum EY, O'Reilly L, White D, Williams R, Armes J and Lindeman
GJ: The LIM domain gene LMO4 inhibits differentiation of mammary
epithelial cells in vitro and is overexpressed in breast cancer.
Proc Natl Acad Sci USA. 98:14452–14457. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Aoyama M, Ozaki T, Inuzuka H, Tomotsune D,
Hirato J, Okamoto Y, Tokita H, Ohira M and Nakagawara A: LMO3
interacts with neuronal transcription factor, HEN2, and acts as an
oncogene in neuroblastoma. Cancer Res. 65:4587–4597. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Isogai E, Ohira M, Ozaki T, Oba S,
Nakamura Y and Nakagawara A: Oncogenic LMO3 collaborates with HEN2
to enhance neuroblastoma cell growth through transactivation of
Mash1. PLoS One. 6:e192972011. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Song YF, Hong JF, Liu DL, Lin QA, Lan XP
and Lai GX: miR-630 targets LMO3 to regulate cell growth and
metastasis in lung cancer. Am J Transl Res. 7:1271–1279.
2015.PubMed/NCBI
|
|
20
|
Watanabe H, Francis JM, Woo MS, Etemad B,
Lin W, Fries DF, Peng S, Snyder EL, Tata PR, Izzo F, et al:
Integrated cistromic and expression analysis of amplified NKX2-1 in
lung adenocarcinoma identifies LMO3 as a functional transcriptional
target. Genes Dev. 27:197–210. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Schmittgen TD and Livak KJ: Analyzing
real-time PCR data by the comparative C(T) method. Nat Protoc.
3:1101–1108. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Hennessy BT, Smith DL, Ram PT, Lu Y and
Mills GB: Exploiting the PI3K/AKT pathway for cancer drug
discovery. Nat Rev Drug Discov. 4:988–1004. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Lin HP, Jiang SS and Chuu CP: Caffeic acid
phenethyl ester causes p21C ip1 induction, Akt signaling reduction,
and growth inhibition in PC-3 human prostate cancer cells. PLoS
One. 7:e312862012. View Article : Google Scholar
|