Introduction
Spinal cord injury (SCI) affects ~2.5 million people
all over the world and there are about 130,000 new cases of SCI
each year (1). Body segments
below the injury level in SCI patients suffer from chronic
paralysis and autonomic dysfunctions, accompanied by several
distressful secondary complications, such as spasticity, bowel and
bladder dysfunction (2). The
bladder dysfunction can result in incontinence and negatively
affects the life quality of SCI patients (3). It may also bring about upper urinary
tract deterioration and cause the death of SCI patients (4). So far, the therapies for the
neurogenic bladder secondary to SCI mainly include catheterization,
anti-cholinergic medications, botulinum toxin A and acupuncture,
but no fully restorative treatments have been exploited (5,6).
Therefore, more basic scientific research should be performed and
try to provide theoretical bases to preferable clinical therapies
for the neurogenic bladder due to SCI. Under the premise that it is
so difficult to achieve neural regeneration as yet, we should focus
on exploring the role of spontaneous excited system in bladder of
SCI patients.
Interstitial cells of Cajal (ICCs) were initially
identified in 1893 and were demonstrated to act as pacemakers
involved in generating slow wave activity and driving peristalsis
in the gastrointestinal (GI) tract (7). In recent years, cells resembling the
ICC in GI tract have been discovered in the urinary bladder of
humans and animals and termed as ICC-like cells (ICC-LCs) (8). ICC-LCs in bladders are deemed to
participate in different cellular processes such as pacemaker
activity or transmitting neural inputs to detrusor smooth muscle,
due to its various locations in the bladder wall (9). Abundant evidence showed that
alterations in the distribution and quantity of bladder ICC-LCs
were associated with several pathological conditions, such as
obstructed bladder, bladder pain syndrome/interstitial cystitis
(BPS/IC) or diabetic bladder (10). Our previous study suggested that
ICC-LCs may be involved in the pathogenesis of SCI induced
neurogenic bladder (11).
Therefore, the functional role of ICC-LCs in SCI induced neurogenic
bladder should be clarified by further research.
Hyperpolarization-activated cyclic nucleotide gated
(HCN) channels, which include four subtypes (HCN1-4) in mammals,
can generate Ih and participate in multiple systemic
functions such as heart contractility, hormonal regulation, central
pattern generation, sensory perception, learning and memory
(12). In a previous study, we
detected that all four HCN subtypes are expressed exclusively in
bladder ICC-LCs and the HCN1 channel is the prominent one. HCN
channels were deemed to be involved in controlling the bladder
pacemaker activity via ICC-LCs (13). Abundant evidence suggests that
altered HCN channels are associated with multiple excitatory
disorders in heterologous systems, such as various types of
epilepsy, sinus bradycardia, atrial fibrillation and Hirschsprung's
disease (14–16). Furthermore, we found that HCN
channel expression and function in bladder ICC-LCs were
significantly increased in detrusor overactive (DO) bladders with
partial bladder outlet obstruction (PBOO) (17). However, the role of HCN channels
in bladder ICC-LCs of SCI induced neurogenic bladder has not been
clarified as yet. In the present study, the altered expression and
function, as well as the possible functional roles of HCN channels
were investigated in rats with SCI induced neurogenic bladder.
Materials and methods
Animals
Sixty female Sprague-Dawley rats weighing 150–180 g
were used in our study. All the rats were randomly assigned to
three groups: control group, sham group and SCI group. All animal
experiments were performed according to the Guide for Care and Use
of Laboratory Animals issued by the National Institutes of Health,
and were authorized by the Research Council and Animal Care and Use
Committee of the Third Military Medical University (Chongqing,
China).
SCI
Rats were anesthetized by intraperitoneal injection
of pentobarbital sodium (40 mg/kg). Under aseptic conditions, we
incised the skin and muscle layers in the rat's back and conducted
a laminectomy to expose the spinal cord. Subsequently, we performed
complete spinal cord transection at vertebral level S2 in rats, as
previously described (18). Rats
were subjected to a laminectomy at S2 level but no SCI in sham
group and received no treatment in control group. After injury,
muscle layers and skin were sutured orderly and rats were injected
with 2 ml sterile saline subcutaneously. Then the rats were placed
into warmed cages with free access to food and water under a
standard 12-h/12-h light/dark cycle. Rats were injected daily with
gentamicin subcutaneously (5 mg/kg) for the first postoperation
week. Manual bladder expression was performed twice daily and
dehydration was monitored daily. After being maintained for 6
weeks, rats were sacrificed for further research. Three rats in SCI
group died of unknown cause before deadline and were excluded from
the experiments.
Hematoxylin and eosin (H&E)
staining
The bladders were isolated from the sacrificed rats
and fixed with 4% paraformaldehyde (Boster, Wuhan, China)
overnight. Then paraffin-embedded blocks were made and 4-µm
sections were manufactured. The H&E staining was conducted
according to a standard protocol (19). Briefly, the sections were first
stained in hematoxylin for 30 sec and then 10–30 sec in 1% eosin
solution. Afterwards, the sections were dehydrated in ethanol and
fixed with mounting medium. All sections were viewed using a
optical microscope (Olympus, Tokyo, Japan).
Urodynamic measurements
As previously described (20), urodynamic measurements were
performed in unconscious rats with urethane anesthesia (1 g/kg body
weight, intraperitoneally). PE-50 polyethylene tubing (Becton,
Dickinson and Company, Franklin Lakes, NJ, USA) was ducted into the
bladder through the urethra. The catheter was connected to a 3-way
valve linked with an infusion pump (AVI 270; 3M, Maplewood, MN,
USA) and a pressure transducer (Chengyi Co., Chengdu, China), which
was calibrated before each experiment. Room temperature saline was
infused into the bladder at a constant rate (10 ml/h) and the
pressure transducer output was amplified and visualized using a
multi-channel signal processing system (RM6240C; Chengyi Co.).
Western blot analyses
For western blot analysis, rat bladders were lysed
in RIPA lysis buffer (Beyotime Institute of Biotechnology,
Shanghai, China) to extract total protein and protein concentration
was measured using the Bio-Rad DC Protein Assay kit (Bio-Rad
Laboratories Inc., Hercules, CA, USA). Protein (50 µg) was
separated by SDS-PAGE gels and then transferred to PVDF membranes
(Merck Millipore, Darmstadt, Germany). Membranes were blocked with
5% bovine serum albumin in Tris-buffered saline for 2 h at room
temperature and incubated with primary antibodies overnight at 4°C
as follows: HCN1 (ab84816, 1:1,000), HCN2 (ab65704, 1:1,000), HCN3
(ab84818, 1:1,000) and HCN4 (ab69054, 1:1,000) (all from Abcam,
Cambridge, MA, USA), GAPDH (AG019, 1:1,000; Beyotime Institute of
Biotechnology), tetratricopeptide repeat-containing
Rab8b-interacting protein (Trip8b) (constant, 75–243, 1:800;
NeuroMab, Davis, CA, USA), filamin A (4762, 1:1,000) and neural
precursor cell-expressed developmentally downregulated (Nedd)4-2
(4013, 1:1,000) (both from Cell Signaling Technology, Inc.,
Danvers, MA, USA) neuronal restrictive silencing factor (NRSF)
(ab21635, 1:1,000; Abcam), tubulin (AT819, 1:1,000; Beyotime
Institute of Biotechnology), then primary antibodies was bound by
horseradish peroxidase-conjugated secondary antibodies (ZB-2301,
ZB-2305, 1:5,000; Zhongshan Golden Bridge Biotechnology Co., Ltd.,
Beijing, China). Signal was detected using ECL substrate
(Millipore, Billerica, MA, USA) and imaged by ImageQuant LAS 4000
Bio-Imaging system (GE Healthcare Life Sciences, Stockholm,
Sweden).
Quantitative RT-PCR
Total RNA was extracted from rat bladders using
TRIzol reagent (Takara Biotechnology Co., Ltd., Dalian, China) and
cDNA was synthesized by PrimeScript™ RT reagent kit (Takara
Biotechnology Co., Ltd.), according to the manufacturer's
instructions. Primers used in this study are listed in Table I. Quantitative RT-PCR was
performed with a StepOnePlus Real-Time PCR system (Invitrogen Life
Technologies, Carlsbad, CA, USA) using SYBR-Green Real-time PCR
Master mix (Toyobo, Osaka, Japan). The thermocycling program
consisted of 95°C for 1 min, 95°C for 15 sec, 60°C for 15 sec and
72°C for 45 sec (40 cycles). Subsequently, the amplified product
was subjected to melting curve analysis. Negative control reactions
were conducted using the same amount of RNA without reverse
transcription. Expression of each gene was normalized to GAPDH.
 | Table IPrimers used for quantitative
RT-PCR. |
Table I
Primers used for quantitative
RT-PCR.
| Genes | Species | Primer
sequences | Accession no. |
|---|
| HCN1 | Rat | F:
5′-CATGCCACAGCTTTGATCCAGTCT-3′
R: 5′-CGCATGTCAGCTGGTAACTTGTG-3′ | NM_053375 |
| HCN2 | Rat | F:
5′-AAGGGCAACAAGGAGATGAAGC-3′
R: 5′-CGCCGCATCATGGGGTACTC-3′ | NM_053684 |
| HCN3 | Rat | F:
5′-GCAGCGCATCCACGAGTACTACG-3′
R: 5′-CCCCGGCAGGTGAAGTTAATAATC-3′ | NM_053685 |
| HCN4 | Rat | F:
5′-CGCTCGCTCTCCACAGGCTG-3′
R: 5′-CGGGGGTGTCTCTGGTGTACTTG-3′ | NM_021658 |
| GAPDH | Rat | F:
5′-GGCCCCTCTGGAAAGCTGTG-3′
R: 5′-CCAGGCGGCATGTCAGATC-3′ | NM_017008 |
Preparation of isolated bladder
cells
After sacrificing the rat, the whole bladder was
aseptically isolated and washed in sterile phosphate-buffered
saline (PBS) solution. Then, the urothelium was dissected away in
the sterile Ca2+-free Hank's solution (Boster). The
bladder was minced and incubated for 10 min at 37°C in 4 ml enzyme
solution. To terminate digestion, 4 ml RMPI-1640 medium (HyClone,
Logan, UT, USA) containing 10% fetal bovine serum (FBS; Gibco, Life
Technologies, Grand Island, NY, USA) was applied. The tissue
suspension was blowed and the cells were collected using a 200-mesh
cell strainer. The digestive process was repeated several times to
harvest more cells. Finally, cells were plated onto sterile
polylysin-coated glass coverslips in RMPI-1640 medium containing
10% FBS and 1% antibiotics/antimycotics (Beyotime Institute of
Biotechnology), and cultured at 37°C in a 95% O2 and 5%
CO2 incubator. The enzyme solution contained the
following components (in mg/ml): 1.0 type II collagenase, 1.0 BSA
and 1.0 trypsin inhibitor (all from Sigma-Aldrich, St. Louis, MO,
USA).
Immunofluorescence
Cells were fixed in 4% paraformaldehyde (Boster) for
30 min, and washed in PBS (10 min × 3 times). To block non-specific
epitopes, cells were incubated with immunostaining blocking buffer
(Beyotime Institute of Biotechnology) for 60 min. Then cells were
incubated overnight at 4°C with primary antibodies: c-kit (sc-1494,
1:50; Santa Cruz Biotechnology, Inc., Santa Cruz, CA, USA), HCN1
(ab84816, 1:100), HCN2 (ab65704, 1:100), HCN3 (ab84818, 1:100) and
HCN4 (ab69054, 1:100) (all from Abcam), followed by the appropriate
fluorescence-conjugated secondary antibodies: Alexa Fluor 488 mouse
anti-goat IgG (bs-0294M, 1:200; Bioss, Beijing, China), Alexa Fluor
647 goat anti-mouse IgG (P0191, 1:200) and Alexa Fluor 647 goat
anti-rabbit IgG (P0180, 1:200) (both from Beyotime Institute of
Biotechnology). Next, cells were incubated with
2-(4-amidinophenyl)-6-indolecarbamidine dihydrochloride (DAPI;
Beyotime Institute of Biotechnology) to label the cell nucleus.
Negative control was performed by omitting the primary antibody.
All incubation steps were followed by washes with PBS (10 min × 3
times). Cells were visualized and photographed using a confocal
laser scanning microscope (Leica, Wetzlar, Germany). The mean
fluorescence density was measured by Image-Pro Plus version 6.0
software (Media Cybernetics, Inc., Rockville, MD, USA).
Patch-clamp
After regularly culturing for 1–2 days, the
patch-clamp technique was performed on the bladder ICC-LCs, which
were distinguished by distinctive cell morphology, stellate or
spindle shapes with multiple branches (9). Electrodes were pulled by model P-97
Flaming/Brown micropipette puller (Sutter Instrument, Novato, CA,
USA), and possessed resistance of 4–6 MΩ. In the whole-cell
voltage-clamp mode, the cells were held at −60 mV and followed by a
stepped potential from −60 to −120 mV in −10 mV increments. Then,
the cells were subjected a voltage jump to −120 mV to fully
activate the HCN channels. Ih was amplified and recorded
by the HEKA EPC10 USB amplifier (HEKA Elektronik, Lambrecht,
Germany) and filtered at a threshold frequency of 2.9 kHz. To
calculate the current density, Ih was normalized to cell
capacitance. The elements of intracellular pipette solution were
(in mM): 130 K aspartate, 0.1 Na2GTP, 5
Na2ATP, 2 MgCl2, 5 CaCl2, 11 EGTA,
10 HEPES, pH 7.2 with KOH. The bath solution included (in mM): 140
NaCl, 1.2 MgCl2, 5.4 KCl, 1.8 CaCl2, 10
glucose, 5 HEPES, pH 7.4 with NaOH. BaCl2 (1 mM) and
0.001 mM TTX (Tocris Bioscience, Bristol, UK) were added into the
bath solution to pharmacologically isolate the Ih. The
specific inhibitor of HCN channels, ZD7288 (50 µM;
Sigma-Aldrich), was applied to further validate the specificity of
Ih.
Measurement of intracellular calcium ion
concentration ([Ca2+]i)
The [Ca2+]i measurement was
tested on the bladder cells that were cultured for 1–2 days. The
primary isolated bladder cells were washed with Hank's solution
(Boster) for 5 min and incubated with Fluo-4 AM (10 µM;
Molecular Probes, Eugene, OR, USA) for 30 min at 37°C. Then,
bladder cells were washed with Hank's solution (5 min × 3 times).
Ca2+ imaging was performed on the bladder ICC-LCs using
the laser scanning confocal microscope (Leica) at an emission
wavelength of 488 nm. Forskolin (FSK) (50 µM; Sigma-Aldrich)
and 8-bromoadenosine 3′,5′-cyclic mono-phosphate (8-Br-cAMP) (50
µM; Tocris Bioscience) were administered into the vessel,
respectively. ZD7288 (50 µM; Sigma-Aldrich) was applied to
further confirm the reliability of isolated bladder ICC-LCs. The
results were presented as the relative fluorescence intensity (RFI
= F1/F0, where F0 is the baseline fluorescence intensity, and F1 is
the real-time fluorescence intensity after drug
administration).
Contractility studies
The dissected rat bladders were softly stripped, the
urothelium layer was then longitudinally sheared into ~3×3×8 mm
strips. The strips were settled into a tissue bath filled with 15
ml aerated Kreb's solution (95% O2 and 5% CO2
at 37°C), and suspended perpendicularly between two crooked hooks
connected to the stretch transducer (Chengyi Co.) and fixed on the
bottom of the bath, respectively. Stretch transducer was calibrated
before every experiment. The strip was equilibrated for 30 min and
stretched with 0.75 g tension. Two doses of the ZD7288 (10 and 50
µM; Sigma-Aldrich) were orderly added into the bathing
solution at 6-min intervals. The continuous contraction curves were
visualized with the RM6240C (Chengyi Co.). The Kreb's solution
contained the following components (in mM): 119 NaCl, 4.7 KCl, 1.2
KH2PO4, 1.2 MgSO4•7H2O,
25 NaHCO3, 2.5 CaCl2 and 11 glucose, adjusted
to pH 7.2 with NaOH.
Statistical analysis
All experimental data are presented as mean ± SD.
Significance between each group was analyzed using one-way analysis
of variance (ANOVA) in SPSS version 13.0 software (SPSS Inc.,
Chicago, IL, USA). P-values <0.05 were regarded as significant.
All experiments were conducted with a minimum of three independent
replications.
Results
Changed morphology and histology of
bladder after SCI
The changes of bladder histology after SCI were
assessed by H&E staining. Under ×100 magnification, we found
that the muscle layers manifested obvious hypertrophy and
urothelium were noticeably incrassated, accompanied by abundant
mesenchyme matter in SCI rats compared with control and sham rats.
Macroscopically, bladders in SCI rats were much greater than that
in control and sham rats (Fig.
1A).
SCI rats exhibited remarkable urodynamics
changes
As shown in Fig.
1B, in control and sham rats, we detected the disciplinary
voiding cycle or contractility of the bladder. However, we did not
observe common voiding contractions of the bladder in SCI rats,
which were replaced by several irregular micturition waves with low
amplitude (black arrows).
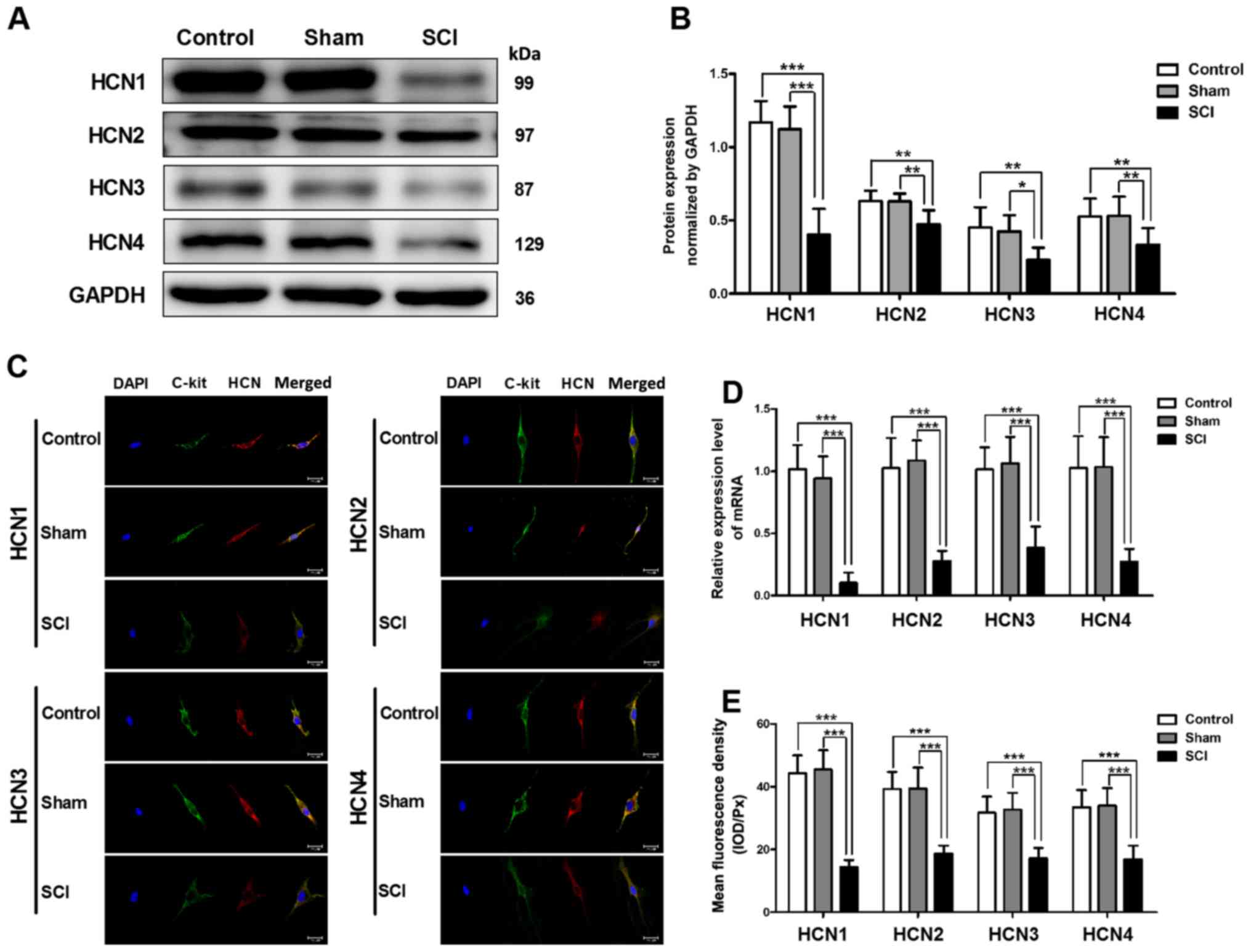
Decreased mRNA and protein expression
levels of HCN channels in SCI rat bladders
Using quantitative RT-PCR and western blot analysis,
we detected the changes in HCN channel expression in SCI rat
bladders. The mRNA and protein expression levels of four HCN
subtypes were significantly decreased in SCI rat bladders, and the
alteration of the HCN1 channel was the most significant (Fig. 2A, B and D). To test the HCN
channel expression levels in single bladder ICC-LC, we utilized
immunofluorescent staining. The results revealed that all four HCN
subtypes were expressed in single bladder ICC-LC which was labeled
by c-kit primary antibody, but the mean fluorescence density of
four HCN subtypes was significantly decreased in SCI rat bladders
and the HCN1 channel also exhibited most significant decrease
(Fig. 2C and E).
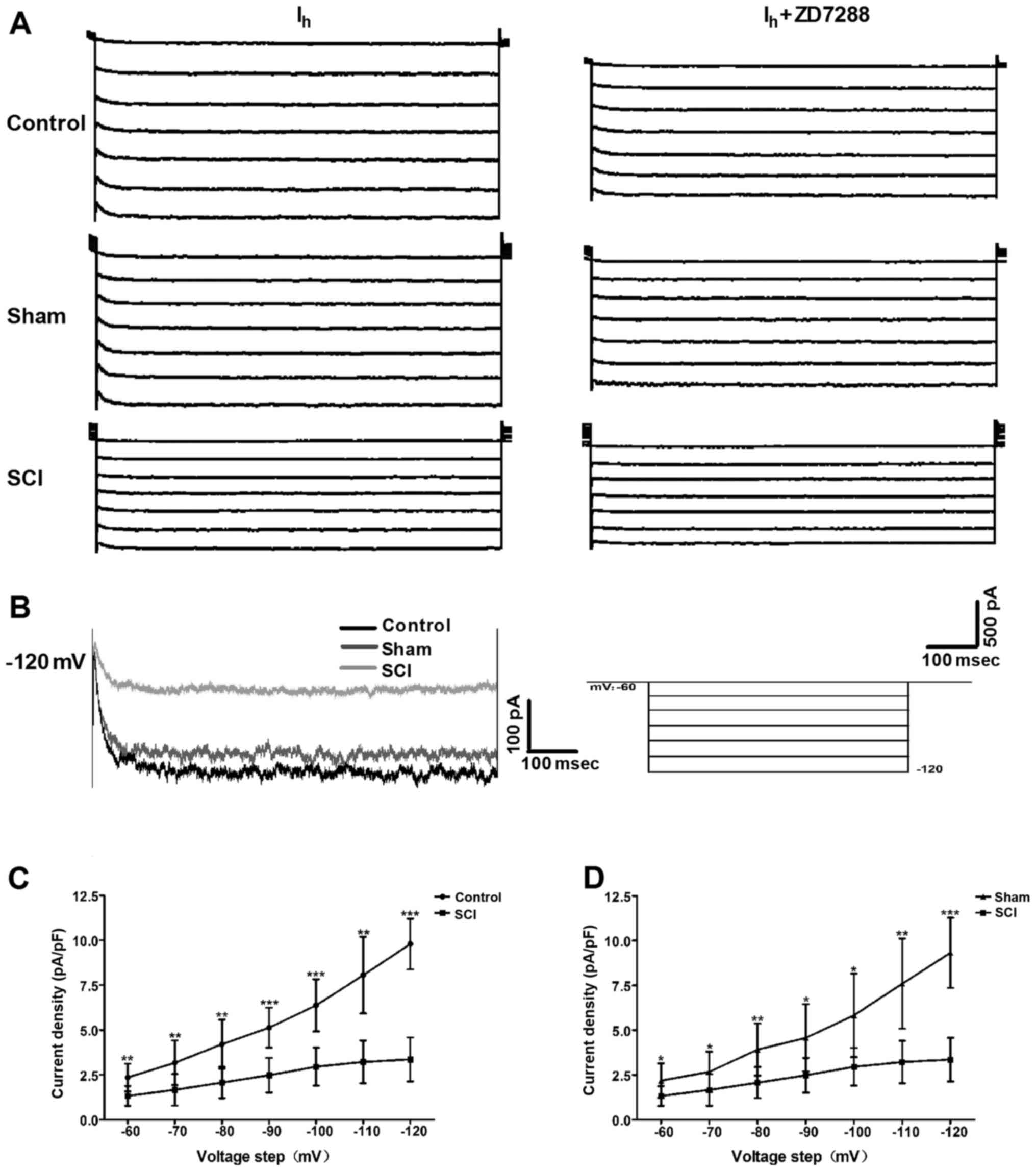
Changes in Ih properties in
bladder ICC-LCs after SCI
To evaluate whether the HCN channel function was
changed after SCI, we utilized the whole-cell patch clamp to record
Ih in isolated bladder ICC-LCs. As shown in Fig. 3A, Ih was recorded in
all three groups. Moreover, the specificity of these recorded
Ih was validated by ZD7288 that reduced the
Ih amplitudes. Fig. 3B
shows that Ih amplitude and HCN channel gating rate in
SCI rats were significantly decreased at the potential of −120 mV.
When normalized to cell capacitance, Ih density in SCI
rats was significantly decreased compared with that in control and
sham rats over the voltage range of −60 to −120 mV, and the
decrease of Ih density at the potential of −120 mV in
SCI rats was the most significant (Fig. 3C and D).
Altered expression levels of HCN1 channel
regulatory proteins in SCI rat bladders
To investigate whether the expression levels of HCN1
channel regulatory proteins were changed in SCI rat bladders, we
detected the protein expression of Trip8b, filamin A, Nedd4-2 and
NRSF. We found that the protein expression levels of Trip8b,
Nedd4-2 and NRSF were significantly upregulated, while the protein
expression level of filamin A was significantly downregulated in
SCI rat bladders (Fig. 4).
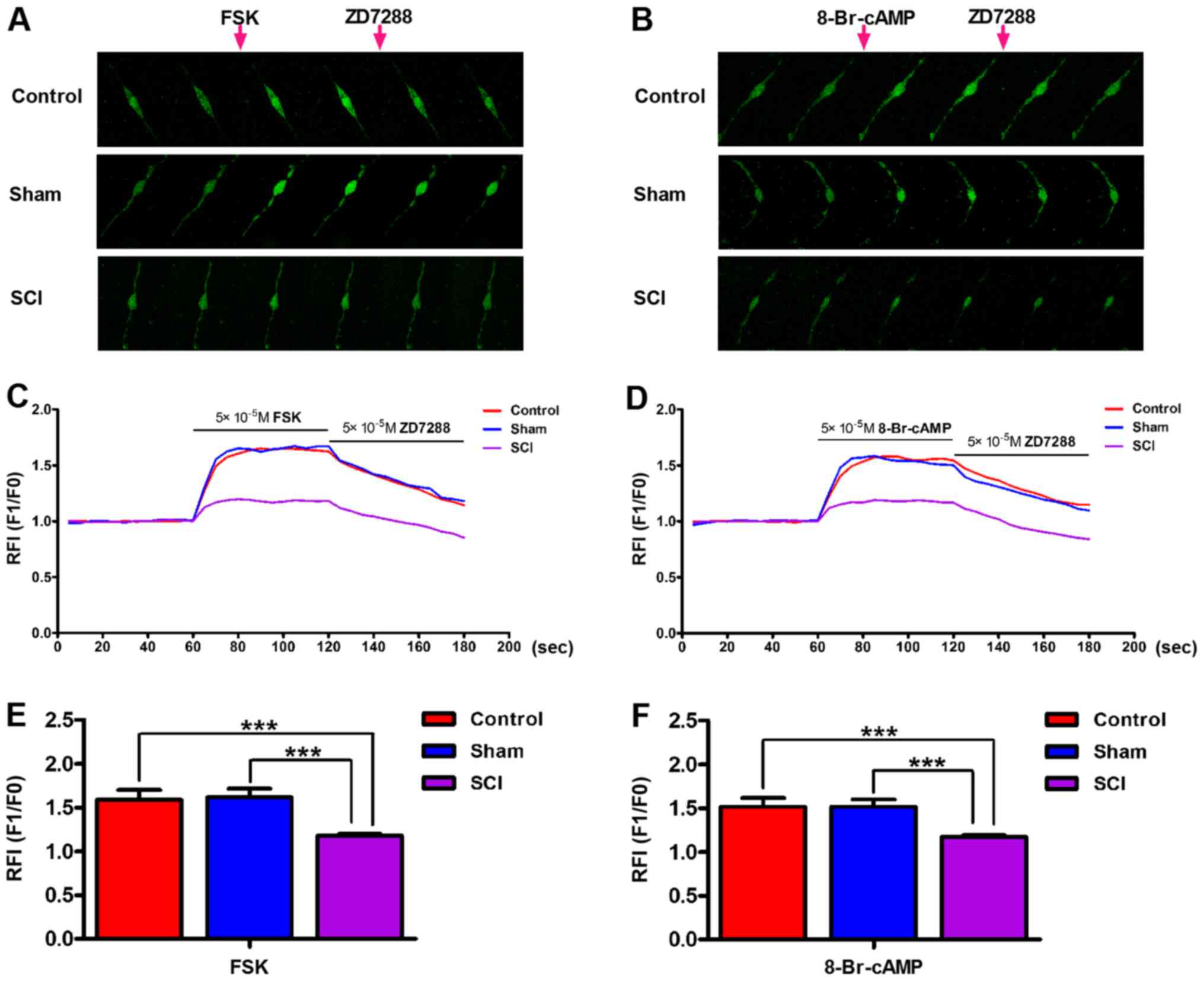
Decreased bladder ICC-LC excitability in
SCI rats
We conducted ([Ca2+]i)
measurements to estimate the bladder ICC-LC excitability in each
group. Upon the application of FSK (50 µM) and 8-Br-cAMP (50
µM), [Ca2+]i in isolated bladder
ICC-LCs from all three groups was significantly elevated.
Furthermore, the increased [Ca2+]i in
isolated bladder ICC-LCs could be reversed by the ZD7288 (50
µM) administration (Fig. 5A
and B). Therefore, the authenticity of isolated bladder ICC-LCs
which were chosen for testing by morphology was further confirmed
by this consequence. The enhancements of
[Ca2+]i facilitated by FSK or 8-Br-cAMP in
SCI rats were significantly weaker than that in control and sham
rats (Fig. 5C–F).
Decreased tolerance of detrusor strip to
ZD7288 in SCI rats
In the contractility studies, isolated detrusor
strips from all three groups generated spontaneous contractions.
The contraction amplitude and frequency of detrusor strips were
significantly reduced in SCI rats (Fig. 6A). The low ZD7288 (10 µM)
concentration did not influence the phasic contractions of detrusor
strips in control and sham groups, but significantly decreased the
contraction amplitude (Fig. 6B),
and increased the frequency of detrusor strips in SCI rats
(Fig. 6C). Only at the high
ZD7288 (50 µM) concentration, the phasic contractions of
detrusor strips in all three groups were significantly inhibited
(Fig. 6).
Discussion
In the present study, we demonstrated that the HCN
channel expression and function were significantly reduced in rats
with SCI induced neurogenic bladder, and the expression levels of
several HCN1 channel regulatory proteins were also significantly
altered. Then, we found that the bladder ICC-LC excitability and
detrusor strip contractility were significantly declined due to the
decrease of HCN channels. These findings indicate that decreased
HCN channels and impaired bladder ICC-LCs are involved in the
pathogenesis of SCI induced neurogenic bladder.
As we know, the primary micturition center locates
in the sacral spinal cord level S2–S4 and damage at this level
commonly leads to a highly compliant acontractile bladder (21). This is supported by our results
detected in histomorphology test and urodynamic measurement.
Moreover, we found that although the regular voiding contraction
was disappeared in SCI rat bladders, several weaken and irregular
micturition waves were existed. This finding prompts us that the
bladder possess the capacity of spontaneous excitement to modulate
bladder contraction under the condition that lost disciplinary
neural control. Thus, in this study, we focused on the
investigation of the changes of spontaneous excitability in SCI
induced neurogenic bladder.
HCN channels which can generate the pacemaker
current, Ih, are deemed to play an important role in the
bladder pacemaker activity (13).
In our study, we demonstrated that the gene and protein expression
levels of four HCN subtypes were significantly decreased in SCI
induced neurogenic bladder. Given that bladder ICC-LC quantities
were significantly decreased in rat with sacral spinal cord
(17), therefore, we investigated
whether the HCN channel expression levels in individual bladder
ICC-LC were changed. To solve our confusion, we conducted
immunofluorescence staining, in which we observed that the protein
expression levels of HCN channels in single bladder ICC-LC was also
signicantly reduced. Furthermore, these data were supported by the
results we detected in patch-clamp experiments that the
Ih density was significantly decreased in individual
ICC-LC of SCI induced neurogenic bladder. The HCN channel gating
rate was obviously decelerated after SCI. In conclusion, we suggest
that the decreased HCN channel expression and function are
associated with the pathogenesis of SCI induced neurogenic
bladder.
In addition, we validated that the HCN1 channel
exhibited the most significant alterations in gene and protein
expression levels after SCI, highlighting the crucial role of HCN1
channels in SCI induced bladder dysfunction. Hence, we mainly
explored whether the expression levels of HCN1 channel regulatory
proteins were changed in SCI induced neurogenic bladder. As we
expected, we found that the expression levels of four HCN1 channel
regulatory proteins including Trip8b, filamin A, Nedd4-2 and NRSF
in bladder were all significantly altered after SCI. We ascribe the
significant reduction of HCN1 channel expression to the
significantly increased transcription factor, NRSF, which can bind
to the HCN1 gene promoter and reduce the HCN1 channel gene
transcription (22). Furthermore,
Trip8b, which contains abundant alternative N-terminal splice
variants, is an important regulatory subunit of HCN1 channel in the
mammalian brain. Different Trip8b splice variants generate various
effects on HCN1 channel membrane trafficking, while all splice
isoforms suppress HCN1 channel gating by antagonizing the function
of cyclic nucleotides (23). The
cytoplasmic scaffolding protein filamin A can interact with the
C-terminal of HCN1 channel through a 22-amino acid region and
enhance HCN1 channel surface expression (24). In contrast, the Nedd4-2 interacts
with the C-terminus of HCN1 channel, then decreases its surface
expression and negatively modulates the channel gating (25). Taken together, we speculate that
the decrease in Ih density and HCN channel gating rate
in SCI rat bladders may be largely attributed to the alterations in
HCN1 channel regulatory proteins. Moreover, we believe that the
altered regulatory proteins of HCN1 channel and even of other HCN
subtypes play an important role in SCI induced neurogenic
bladder.
To our knowledge, the functional role of HCN
channels in regulating cytomembrane electrophysiology has been well
clarified. HCN channels can be activated by membrane
hyper-polarization or cyclic nucleotides, particularly cyclic
adenosine monophosphate (cAMP), and depolarize membrane potential
toward the threshold for firing action potential by generating an
inward current, Ih (26). Moreover, such HCN channels induced
pacemaker depolarization subsequently activate the
low-voltage-activated Ca2+ channels, such as T-type or
L-type Ca2+ channels, and facilitate robust
Ca2+ influx into cells (27). Therefore, HCN channels may not
only act as pacemaker channels to initiate rhythmic burst firing in
neurons and to control cardiac rhythm (28), but also regulate the spontaneous
pacemaker potentials in cultured colonic ICCs (29). In this study, we performed
[Ca2+]i measurements to detect the effects of
HCN channels on the excitability of bladder ICC-LCs, which also
possess the ability to generate spontaneous action potential
(30). Upon the administration of
the two HCN channel agonists (FSK and 8-Br-cAMP), prominent
Ca2+ influx was observed in bladder ICC-LCs from normal
and sham rats, indicating that HCN channels may also participate in
modulating the bladder ICC-LC pacemaker activity. However, the
effects of FSK and 8-Br-cAMP on the bladder ICC-LCs in SCI group
were significantly weakened, further highlighting the important
role of HCN channels in regulating bladder ICC-LC excitability.
Furthermore, we suggest that the impaired bladder ICC-LCs ascribed
to decreased HCN channel expression and function are involved in
SCI induced neurogenic bladder.
Bladder ICC-LCs are deemed to control the
spontaneous contractions of detrusor smooth muscle (31). Thus, in our study, corresponding
to the decreased bladder ICC-LC excitability, we found that
detrusor strips in SCI rats generated much weaker spontaneous
phasic contractions than that in normal and sham rats. We further
detected that the tolerance of detrusor strips to ZD7288 were
decreased in SCI rats. These results could also be attributed to
the decreased HCN channel expression and function. Accordingly, we
conclude that HCN channels can control the spontaneous contraction
of detrusor strips via regulating bladder ICC-LC excitability, and
correspondingly, altered HCN channel expression and function in
pathologic conditions may lead to disordered detrusor strip
contractility by damaging the bladder ICC-LC function.
Compounds targeting HCN channels are not only widely
utilized to treat several heart diseases and neuropathic pain, but
also used as anticonvulsant and anaesthetic drugs (32). For example, ivabradine is the
first clinically approved medication that specifically targets HCN
channels to treat chronic stable angina pectoris in patients with
contraindication or intolerance for β-blockers (33). In the present study, our data
indicate that HCN channels in bladder ICC-LCs may be considered as
a novel therapeutic target to improve bladder function in SCI
induced neurogenic bladder. The limitation of our study is that we
only tested the functional role of bladder HCN channels in rats
subjected with sacral cord injury. To more completely characterize
the role of HCN channels in SCI induced neurogenic bladder, animal
models with suprasacral cord injury should be used in further
studies.
In conclusion, our findings illustrate the possible
mechanism that decreases in HCN channel expression and function
induced by altered regulatory protein expression, can impair the
bladder ICC-LC excitability and damage the detrusor strip
contractility, and then lead to bladder dysfunction. The present
mechanism may be involved in the pathological process of SCI
induced neurogenic bladder.
Abbreviations:
|
SCI
|
spinal cord injury
|
|
HCN
|
hyperpolarization-activated cyclic
nucleotide-gated
|
|
FSK
|
forskolin
|
|
8-Br-cAMP
|
8-bromoadenosine 3′,5′-cyclic
monophosphate
|
|
ICC-LCs
|
interstitial cells of Cajal-like
cells
|
|
BPS/IC
|
bladder pain syndrome/interstitial
cystitis
|
|
DO
|
detrusor overactive
|
|
PBOO
|
partial bladder outlet obstruction
|
|
NRSF
|
neuronal restrictive silencing
factor
|
|
Trip8b
|
tetratricopeptide repeat-containing
Rab8b-interacting protein
|
|
Nedd
|
neural precursor cell-expressed
developmentally downregulated
|
Acknowledgments
Not applicable.
Notes
[1]
Funding
The present study was supported by the National
Natural Science Foundation of China (NSFC) (grant nos. 81230017 and
81300630).
[2] Availability
of data and material
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
[3] Authors'
contributions
QL performed the major experiments and drafted the
manuscript. CW and SH analyzed the data. QW and TZ participated in
the hematoxylin and eosin staining, western blot analyses,
quantitative RT-PCR and immunofluorescence. XL, XL and XH were
involved in the patch-clamp experiments,
[Ca2+]i measurements, contractility studies
and urodynamic measurements. LL was responsible for designing the
experiments and editing the paper. All authors read and approved
the final manuscript.
[4] Ethics
approval and consent to participate
All animal experiments were performed according to
the Guide for Care and Use of Laboratory Animals issued by the
National Institutes of Health, and were authorized by the Research
Council and Animal Care and Use Committee of the Third Military
Medical University, China.(approval no. SYXK20070002).
[5] Consent for
publication
Not applicable.
[6] Competing
interests
The authors declare that they have no competing
interests.
References
|
1
|
Thuret S, Moon LD and Gage FH: Therapeutic
interventions after spinal cord injury. Nat Rev Neurosci.
7:628–643. 2006. View
Article : Google Scholar : PubMed/NCBI
|
|
2
|
Kim JH, Shim SR, Doo SW, Yang WJ, Yoo BW,
Kim JM, Ko YM, Song ES, Lim IS, Lee HJ, et al: Bladder recovery by
stem cell based cell therapy in the bladder dysfunction induced by
spinal cord injury: systematic review and meta-analysis. PLoS One.
10:e01134912015. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Ku JH: The management of neurogenic
bladder and quality of life in spinal cord injury. BJU Int.
98:739–745. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Cetinel B, Onal B, Can G, Talat Z, Erhan B
and Gunduz B: Risk factors predicting upper urinary tract
deterioration in patients with spinal cord injury: a retrospective
study. Neurourol Urodyn. 36:653–658. 2017. View Article : Google Scholar
|
|
5
|
Harris CJ and Lemack GE: Neurourologic
dysfunction: evaluation, surveillance and therapy. Curr Opin Urol.
26:290–294. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Zhang T, Liu H, Liu Z and Wang L:
Acupuncture for neurogenic bladder due to spinal cord injury: a
systematic review protocol. BMJ Open. 4:e0062492014. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Gfroerer S and Rolle U: Interstitial cells
of Cajal in the normal human gut and in Hirschsprung disease.
Pediatr Surg Int. 29:889–897. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Sanders KM, Ward SM and Koh SD:
Interstitial cells: regulators of smooth muscle function. Physiol
Rev. 94:859–907. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
McCloskey KD: Interstitial cells in the
urinary bladder - localization and function. Neurourol Urodyn.
29:82–87. 2010. View Article : Google Scholar
|
|
10
|
McCloskey KD: Bladder interstitial cells:
an updated review of current knowledge. Acta Physiol (Oxf).
207:7–15. 2013. View Article : Google Scholar
|
|
11
|
Deng J, Zhang Y, Wang L, Zhao J, Song B
and Li L: The effects of Glivec on the urinary bladder excitation
of rats with suprasacral or sacral spinal cord transection. J Surg
Res. 183:598–605. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Yang Q, Kuzyk P, Antonov I, Bostwick CJ,
Kohn AB, Moroz LL and Hawkins RD: Hyperpolarization-activated,
cyclic nucleotide-gated cation channels in Aplysia: contribution to
classical conditioning. Proc Natl Acad Sci USA. 112:16030–16035.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
He P, Deng J, Zhong X, Zhou Z, Song B and
Li L: Identification of a hyperpolarization-activated cyclic
nucleotide-gated channel and its subtypes in the urinary bladder of
the rat. Urology. 79:1411.e7–1411.e13. 2012. View Article : Google Scholar
|
|
14
|
He C, Chen F, Li B and Hu Z:
Neurophysiology of HCN channels: from cellular functions to
multiple regulations. Prog Neurobiol. 112:1–23. 2014. View Article : Google Scholar
|
|
15
|
Sartiani L, Romanelli MN, Mugelli A and
Cerbai E: Updates on HCN channels in the heart: function,
dysfunction and pharmacology. Curr Drug Targets. 16:868–876. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
O'Donnell AM, Coyle D and Puri P:
Decreased expression of hyperpolarisation-activated cyclic
nucleotide-gated channel 3 in Hirschsprung's disease. World J
Gastroenterol. 21:5635–5640. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Deng T, Zhang Q, Wang Q, Zhong X and Li L:
Changes in hyperpolarization-activated cyclic nucleotide-gated
channel expression and activity in bladder interstitial cells of
Cajal from rats with detrusor overactivity. Int Urogynecol J Pelvic
Floor Dysfunct. 26:1139–1145. 2015. View Article : Google Scholar
|
|
18
|
Lucin KM, Sanders VM, Jones TB, Malarkey
WB and Popovich PG: Impaired antibody synthesis after spinal cord
injury is level dependent and is due to sympathetic nervous system
dysregulation. Exp Neurol. 207:75–84. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Fischer AH, Jacobson KA, Rose J and Zeller
R: Hematoxylin and eosin staining of tissue and cell sections. CSH
Protoc. pdb.prot4986. 2008. View Article : Google Scholar
|
|
20
|
Andersson KE, Soler R and Füllhase C:
Rodent models for urodynamic investigation. Neurourol Urodyn.
30:636–646. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Goldmark E, Niver B and Ginsberg DA:
Neurogenic bladder: from diagnosis to management. Curr Urol Rep.
15:4482014. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Bender RA and Baram TZ: Hyperpolarization
activated cyclic-nucleotide gated (HCN) channels in developing
neuronal networks. Prog Neurobiol. 86:129–140. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Santoro B, Piskorowski RA, Pian P, Hu L,
Liu H and Siegelbaum SA: TRIP8b splice variants form a family of
auxiliary subunits that regulate gating and trafficking of HCN
channels in the brain. Neuron. 62:802–813. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Gravante B, Barbuti A, Milanesi R, Zappi
I, Viscomi C and DiFrancesco D: Interaction of the pacemaker
channel HCN1 with filamin A. J Biol Chem. 279:43847–43853. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Wilkars W, Wollberg J, Mohr E, Han M,
Chetkovich DM, Bähring R and Bender RA: Nedd4-2 regulates surface
expression and may affect N-glycosylation of
hyperpolarization-activated cyclic nucleotide-gated (HCN)-1
channels. FASEB J. 28:2177–2190. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Biel M, Wahl-Schott C, Michalakis S and
Zong X: Hyper-polarization-activated cation channels: from genes to
function. Physiol Rev. 89:847–885. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Robinson RB and Siegelbaum SA:
Hyperpolarization-activated cation currents: from molecules to
physiological function. Annu Rev Physiol. 65:453–480. 2003.
View Article : Google Scholar
|
|
28
|
Benarroch EE: HCN channels: function and
clinical implications. Neurology. 80:304–310. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Shahi PK, Choi S, Zuo DC, Kim MY, Park CG,
Kim YD, Lee J, Park KJ, So I and Jun JY: The possible roles of
hyper-polarization-activated cyclic nucleotide channels in
regulating pacemaker activity in colonic interstitial cells of
Cajal. J Gastroenterol. 49:1001–1010. 2014. View Article : Google Scholar
|
|
30
|
Wu Y, Shi C, Deng J, Zhang X, Song B and
Li L: Expression and function of muscarinic subtype receptors in
bladder interstitial cells of cajal in rats. Urol J. 11:1642–1647.
2014.PubMed/NCBI
|
|
31
|
Kubota Y, Biers SM, Kohri K and Brading
AF: Effects of imatinib mesylate (Glivec) as a c-kit tyrosine
kinase inhibitor in the guinea-pig urinary bladder. Neurourol
Urodyn. 25:205–210. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Postea O and Biel M: Exploring HCN
channels as novel drug targets. Nat Rev Drug Discov. 10:903–914.
2011.PubMed/NCBI
|
|
33
|
Riccioni G, Vitulano N and D'Orazio N:
Ivabradine: beyond heart rate control. Adv Ther. 26:12–24. 2009.
View Article : Google Scholar : PubMed/NCBI
|